

Abstract
Cancer cells have a variety of interactions with neighboring connective tissue, and this activity primarily involves fibroblasts. Co-culture of fibroblasts derived from human skin with cancer cells results in the conversion of fibroblasts into cancer-associated fibroblasts, which are known to support tumor growth and invasiveness. To evaluate the effect of radiation on tumor-fibroblast interactions, the present study co-cultivated fibroblasts from pre-irradiated and non-irradiated human skin with FaDu head and neck squamous cell carcinoma cells for 3 days. Subsequently, cells were analyzed for tumor viability, apoptosis, and secretion of interleukin (IL)-6 and −8 by performing an MTT assay, Annexin V-propidium iodide test and enzyme-linked immunosorbent assay, respectively. Co-culture of FaDu cells with pre-irradiated fibroblasts resulted in a significant decrease in tumor viability, a notable increase in apoptosis and significantly lower levels of IL-8 compared with FaDu cells cultured with non-irradiated fibroblasts. Therefore, we propose that pre-irradiation changes the properties of fibroblasts and their effects on co-cultivated tumor cells, and, thus may lead to an improved understanding of the therapeutic options for patients that have already undergone irradiation.
Keywords: fibroblasts, head and neck squamous cell carcinoma, tumor stroma, co-culture, irradiation
Introduction
Squamous cell carcinoma is a major cause of cancer morbidity and mortality, and is one of the most commonly occurring malignancies worldwide. The incidence of head and neck squamous cell carcinoma (HNSCC) is estimated at 500,000 per year in the United States (1). Changes have been made regarding the therapeutic strategies applied for such patients, including different surgical approaches, radiotherapy and chemotherapy either in alone or in various combinations; however, despite this multi-modal treatment strategy, survival rates have not improved significantly over the past several decades, with an overall 5-year survival rate of 40–50% (2).
There is increasing evidence that tumor growth is not only determined by malignant cancer cells, but also by the tumor stroma (3). The tumor stroma is partially composed of fibroblasts and the connective tissue they produce. Physiologically, due to their close proximity, continuous crosstalk between the stroma and epithelia controls tissue differentiation (4). In the pathology of a tissue wound, the stroma takes on the role of repair, while paracrine signaling alters epithelial proliferation and differentiation (4,5). As cancer functionally resembles a chronic wound, these mechanisms of wound repair are useful to examine from an oncological point of view. In this regard, fibroblasts, which are the primary component of tumor stroma, have been shown to be prominent modifiers of cancer progression (6). It has also become increasingly clear that there are different subpopulations of fibroblasts, as a result of their continuous interaction with epithelial or cancer cells. One of these subpopulations, termed cancer-associated fibroblasts (CAFs), has been shown to be an important promoter of tumor growth and progression (7). CAFs induce epithelial-mesenchymal transition (EMT) in epithelial tumor cells, which is a key factor in the invasion of squamous cell carcinomas (8).
It is also known that cytokines are involved in tumor progression. For example, interleukin-6 (IL-6) has been shown to promote cancer resistance against chemotherapeutic agents, such as cisplatin (9), while elevated IL-8 expression is associated with an enhanced metastatic potential in different cancer entities (9,10). Numerous therapeutic strategies concerning HNSCC involve the use of radiation, either combined with chemotherapy as the primary therapy or via adjuvant radiation following surgery. However, there have been few studies analyzing the impact of radiation on the tumor stroma. Recent studies showed no significant changes in the growth or proliferation of CAFs themselves when exposed to radiation in vitro (11,12). Instead, the application of low doses of radiation (<20 Gy) appears to enhance the capability of fibroblasts to promote survival of co-cultured cancer cells (13). However, in these aforementioned studies, irradiation was delivered to cells as a monolayer culture in vitro. Thus, the complex interactions of pre-irradiated tissue in vivo and their influence on CAFs is still largely unknown.
The primary focus of the present study was the influence of pre-irradiation on the interactions between fibroblasts and HNSCC. The aim of the present study was to analyze the in vitro effects pre-irradiation of fibroblasts on HNSCC cell lines compared with non-irradiated fibroblasts in terms of morphological changes, viability and apoptosis.
Materials and methods
Culture of the FaDu cell line
The FaDu HNSCC cell line was established from a human hypopharyngeal squamous cell carcinoma (14) and was obtained from DSMZ (Braunschweig, Germany). Cells were cultivated in RPMI-1640 medium with 10% fetal calf serum (FCS) (Biochrom GmbH, Berlin, Germany), 100 µg/ml streptomycin, 100 U/ml penicillin, 1% sodium pyruvate (100 mM; Biochrom GmbH, Berlin, Germany) and 1% non-essential amino acids [RPMI-expansion medium (EM); 100-fold concentration]. The culture conditions included a temperature of 37°C with 5% CO2 in culture flasks. Medium was replaced every other day and passaging was performed by trypsinization (0.25% trypsin; Gibco, Thermo Fisher Scientific, Inc., Waltham, MA, USA) once cells reached 70–80% confluencey. Subsequently, cells were washed and seeded in new flasks or treatment wells. Cells in the exponential growth phase were used for performing experiments.
Acquisition and culture of fibroblasts
Fibroblasts were obtained from skin samples of voluntary patients undergoing neck surgery (n=20) at the University Hospital of Würzburg (Würzburg, Germany) between August 2011 and October 2012, with 10/20 patients having been previously irradiated with intensity modulated irradiation with 60–70 Gy for 6 weeks during head and neck cancer therapy 6–16 months previously. Approval was obtained from the Ethics Committee of the Medical Faculty, University of Würzburg (approval no., 12/06) and evidence of informed consent was received from all individuals included in the present study. The skin samples were cleared of fat and cut into small pieces (2–3 mm), which were then seeded on 6-plate wells. After 60 min of culture without medium, tissue pieces were attached to the bottom of the plates strongly enough not to be washed away by the addition of Dulbecco's modified Eagle medium (DMEM; Gibco, Thermo Fisher Scientific, Inc.) with 10% FCS, 100 U/ml penicillin and 100 µg/ml streptomycin [DMEM-expansion medium (DMEM-EM)] at 37°C in 5% CO2. Fibroblasts grew out from these tissue pieces into the periphery. Medium was replaced every other day and passaging was performed by trypsinization (0.25% trypsin; Gibco, Thermo Fisher Scientific, Inc.) once cells reached 70–80% confluencey. Subsequently, cells were washed with phosphate buffered saline (PBS) and seeded in new flasks or treatment wells.
Group composition
The experiment included 10 fibroblast cultures from pre-irradiated tissue and 10 from non-irradiated samples. Simultaneously, FaDu cells were cultivated alone in parallel to serve as the control group. A schematic of all groups are shown in Fig. 1.
Figure 1.

Schematic of group composition and abbreviations. FB, fibroblasts; irr. irradiated.
Transwell culture
A Transwell system (Costar; Corning, Inc., Corning, NY, USA) was used to analyze the effects of fibroblasts on the FaDu HNSCC cell line. First, a co-culture was generated, as follows: FaDu cells (5×104) were added to 1 ml RPMI-EM on the bottom well of the Transwell 12-well plate; subsequently, fibroblasts (5×104) with 0.5 ml RPMI-EM were added to the upper wells of the Transwell system and transferred to the wells containing FaDu cells. Following co-culture for 3 days at 37°C, the fibroblasts were discarded, and the co-culture of FaDu cells was used in additional analysis.
MTT assay
After 3 days of co-culture, the MTT (Sigma-Aldrich) colorimetric staining method was performed, according to Mosmann (15), to determine cell viability. Cells were seeded at 10,000 cells per well in a 12-well plate. All wells were incubated with 1 ml MTT (1 mg/ml) for 5 h at 37°C and 5% CO2. MTT was then removed and 1 ml isopropanol was added, followed by another incubation period of 1 h at 37°C and 5% CO2. Measurement of the color conversion of the blue formazan dye was performed using a multi-plate reader (Titertek Multiskan PLUS MK II; Thermo Labsystems, Thermo Fisher Scientific, Inc.) at a wavelength of 570 nm.
Annexin V-propidium iodide analysis
A BD Pharmingen Annexin V-APC kit from (BD Biosciences, Heidelberg, Germany) was used to evaluate apoptosis. After 3 days of co-culture, cells in suspension and adherent cells were harvested then washed twice with PBS, followed by resuspension in 1:10 binding buffer [0.1 M HEPES (pH 7.4), 1.4 M NaCl, 25 mM CaCl2) at a concentration of 1×106 cells/ml. Aliquots of this cell suspension (100 µl; 1×105 cells) were then transferred to a 5 ml culture tube. Propidium iodide (5 µl) and Annexin V-APC (5 µl) were added to each aliquot. After 15 min of incubation at room temperature in the dark, the cells were resuspended with 400 µl 1:10 binding buffer. A FACSCanto flow cytometer was used to analyze the samples with BD FACSDiva version 5.0.3 software (BD Biosciences). Only cells with damaged membranes were stained by propidium iodide.
IL-6/8 enzyme-linked immunosorbent assay (ELISA)
For measurement of the secretion of IL-6 and IL-8, the supernatants were collected (centrifugation, 150 × g for 5 min at 37°C) after 3 days of co-culture and stored at −20°C in sterile tubes until further use. DMEM-EM served as control. Human IL-6 and IL-8 kits (catalog nos., 950.030.192 and 950.050.192, respectively; Diaclone SAS, Besançon, France) were used and the experiments were performed in duplicate. The ELISA plate was read at 450 nm (Titertek Multiskan PLUS MK II; Thermo Labsystems, Thermo Fisher Scientific, Inc.). The concentration of IL-6 and IL-8 were determined by constructing a standard curve using recombinant IL-6 and IL-8. For statistical analysis, mono-cultures [FaDu or fibroblasts (non-irradiated or pre-irradiated)] were compared with co-cultures [FaDu and fibroblasts (non-irradiated or pre-irradiated)].
Statistical analysis
The data collected was transferred to standard spreadsheets and statistically analyzed using GraphPad Prism software (version 4.0; GraphPad Software, Inc., San Diego, CA, USA). Data are presented as the mean ± standard deviation of three experiments, unless otherwise stated. The Gaussian distribution was tested via first column analysis. In cases of Gaussian distribution, one-way analysis of variance followed by Tukey's multiple comparison test was used, and in cases of a non-Gaussian distribution, the Kruskal-Wallis test was performed. P<0.05 was used to indicate a statistically significant difference.
Results
MTT assay
A co-culture of FaDu cells with fibroblasts from non-irradiated skin samples showed a significant increase in FaDu cell viability compared with the mono-culture of FaDu cancer cells alone (P=0.0001). By contrast, when co-cultured with fibroblasts from pre-irradiated tissue, there was a significant decrease in FaDu cell viability compared with the non-irradiated co-culture and the mono-culture (P=0.0001; Fig. 2).
Figure 2.
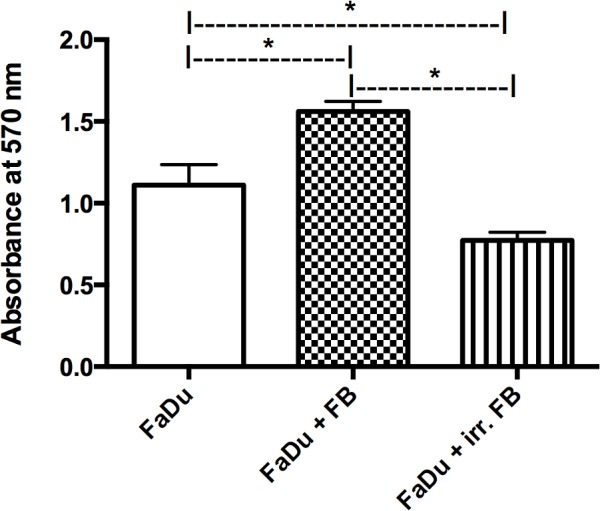
MTT assay of FaDu cells in mono-culture and co-culture. Significant enhancement in cell viability was observed in the co-culture with non-irradiated FBs compared with irr. FBs. *P<0.05 vs. FaDu. FB, fibroblasts; irr. irradiated.
Annexin V-propidium iodide
Annexin V-propidium iodide analysis revealed no statistically significant differences between a co-culture with non-irradiated fibroblasts and pre-irradiated fibroblasts regarding the rate of apoptosis in FaDu cells (P=0.21). This indicates that apoptosis may not be the dominant mechanism involved (Fig. 3).
Figure 3.

Representative flow cytogramme of Annexin V binding (abscissa) versus propidium iodide uptake (ordinate) in FaDu cells incubated with pre-irradiated or non-irradiated fibroblasts. There is no statistically significant difference between the two groups, indicating that a mechanism other than apoptosis may be involved (data not shown). Q1, % of damaged cells; Q2, % of necrotic cells; Q3, % of viable cells; Q4, % of apoptotic cells. APC-A, allophycocyanin-A.
Quantitative analysis of IL-6 and IL-8 expression
In mono-culture, IL-6-expression was significantly higher in normal fibroblasts than in fibroblasts from pre-irradiated tissue (P=0.0001). FaDu cells in mono-culture showed negligible secretion of IL-6 (data not shown). When in co-culture, the level of secretion in both groups (pre-irradiated or non-irradiated) was significantly higher (P=0.001) compared with mono-culture, yet there was no significant difference between the two groups (P=0.0787) (Fig. 4).
Figure 4.

Concentration of IL-6 in different mono-cultures and co-cultures. Significantly higher levels of IL-6 were observed in mono-cultures of non-irradiated FBs compared with irr. FBs. In addition, the increase in IL-6 concentration in co-culture of both groups compared with the matched mono-culture was statistically significant. No significant difference was observed when comparing the two co-culture groups. Data for FaDu mono-culture not shown. *P<0.05. IL-6, interleukin-6; FB, fibroblasts; irr. irradiated.
IL-8 expression in mono-culture was only markedly present in non-irradiated fibroblasts, while FaDu cells (data not shown) and pre-irradiated fibroblasts showed negligible expression. When in co-culture, both groups demonstrated an increase in IL-8 secretion compared with their respective mono-cultures (P=0.0015), although the non-irradiated fibroblasts still exhibited a significantly higher level of expression compared with the pre-irradiated fibroblasts (P=0.001; Fig. 5).
Figure 5.
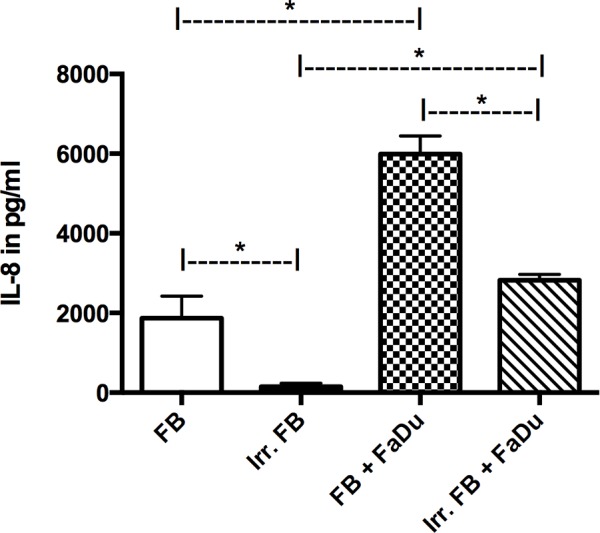
Concentration of IL-8 in different mono-cultures and co-cultures. In mono-culture and in co-culture, significantly higher levels of IL-8 were observed in non-irradiated FBs compared with irr. FBs. An increase in IL-8 production was observed in both co-culture groups compared with the matched mono-culture groups, while the difference between the two co-culture groups remained statistically significant. Data for FaDu mono-culture not shown. *P<0.05. IL-8, interleukin-8; FB, fibroblasts; irr. irradiated.
Discussion
The present study focused on the effects of prior radiation on the properties of fibroblasts in co-culture with FaDu HNSCC cells. Over the past few decades, the effects of the tumor stroma on cancer cells has been thoroughly investigated, and CAFs have been shown to promote the growth of cancer cells, many of which have already been extracted from a number of types of invasive human carcinoma (16). CAFs have been induced from regular fibroblasts, and even from mesenchymal stem cells, via treatment with carcinoma cell-derived medium (17) or a co-culture model (18). In addition, previous studies have demonstrated that co-culture with CAFs changes the level of secretion of numerous cytokines from carcinoma cells (19,20), thus, giving them a higher potential for invasion and metastasis. The possibility of the two cell types mutually influencing each other is one of the advantages of the Transwell co-culture model, and the background for selecting it for use in the present study.
The results of the co-culture of FaDu cells with fibroblasts from non-irradiated skin samples in the present study revealed a significant increase in FaDu cell viability when compared with a mono-culture of FaDu cells. This is consistent with the data available in the literature regarding various other carcinoma cell types, such as prostate carcinoma (20) or breast cancer (21). Thus, although data regarding the effect of co-cultured fibroblasts on the proliferation of HNSCC is scarce, particularly on a quantitative basis, the current results were as anticipated.
In a co-culture of FaDu cells with fibroblasts derived from irradiated tissue, a significant decrease in tumor cell viability was found compared with the non-irradiated fibroblast co-culture and compared with the mono-culture of FaDu cells. According to the actual literature, there is no data available regarding a co-culture of irradiated fibroblasts with HNSCC cells. However, such data in regard to other carcinoma cell types differs from the present study. For example, a study by Ohuchida et al demonstrated an increase in cancer invasion and progression for pancreatic cancer cells co-cultivated with irradiated fibroblasts (22). This result was predominantly explained by tumor-stromal interactions, particularly via increased phosphorylation of c-Met, although no increase in hepatocyte growth factor, a mediator of tumor-stromal interactions, was found (22). In addition, Tsai et al showed that a low-dose radiation of 20 Gy could promote the invasiveness of co-cultivated breast cancer cells, and even render the cancer cells radioresistant (13). Both studies concluded that irradiation of stromal fibroblasts is a promoting factor for cancer cell growth. This is in accordance with the clinical findings of increased cases of metastasis in previously irradiated tissue following definitive radiotherapy, as described for nasopharyngeal carcinoma (23), as well as in an animal model of squamous cell carcinoma (24). However, this has yet to be investigated for HNSCC cells. Furthermore, the radiation in the aforementioned studies was applied at a dose of only 5–10 Gy in vitro immediately prior to the experiments, whereas the fibroblasts in the present study were cultivated from skin that had been radiated with 60–70 Gy a number of months prior to the experiment. Therefore, a prolonged effect of radiation on the tissue samples in vivo may explain the differing results. A comparative study on fibroblasts cultivated from pre-irradiated skin and fibroblasts that received radiation in vitro in a co-culture should be conducted in the future for further clarification.
Regarding the levels of cytokine secretion, significantly higher levels of IL-6 and IL-8 were observed in a mono-culture of non-irradiated fibroblasts compared with in a mono-culture of pre-irradiated fibroblasts. When in co-culture with FaDu cells, IL-6 secretion levels were similar in both groups, while the levels of IL-8 remained significantly lower in the pre-irradiated group compared with the non-irradiated group. Various studies have shown that patients with HNSCC commonly overexpress cytokines, including IL-1, IL-6, tumor necrosis factor (TNF)-α and TNF-β (25,26). Among others, IL-6 has been shown to be elevated in HNSCC cell lines of metastatic origin compared with those cultivated from the primary tumor lesion (19). Therefore, an IL-6 assay was integrated into the present study, as increased levels of IL-6 indicate a higher potential for tumor proliferation and metastasis. IL-8 was also included due to its known tumor-promoting properties (27), primarily through an increase in angiogenesis. The increased secretion observed in both co-cultured groups correlates well with previous clinical studies that describe increased levels of IL-6 and IL-8 in HNSCC tumor patients compared with healthy controls (25,26). The lower levels of IL-6 and IL-8 in the pre-irradiated mono-culture group compared with the non-irradiated mono-culture group may be explained by the irradiation damage caused to the tissue from which the fibroblasts were cultivated, indicating a long-term effect on the stromal tissue. However, when co-cultured with FaDu cells, the pre-irradiated fibroblasts appeared to increase cytokine production; increasing to the same level as the non-irradiated group for IL-6 and at least partially increasing for IL-8. This raises the question of whether the differences in the co-cultures are time-dependent, or if there is a limit on how much of the productive capabilities the pre-irradiated fibroblasts can regain.
It should be noted that there are certain drawbacks to the present study design, as the setup is not completely physiological. The fibroblasts used were not CAFs, which define the tumor stroma, as they only develop in the vicinity of cancer cells. Thus, we can only postulate the effects of regular fibroblasts with and without previous irradiation on cancer cells rather than changes in the actual tumor stroma caused by irradiation. To evaluate changes in the tumor stroma caused by irradiation, irradiation of tumor cells as well as with the surrounding fibroblasts would be necessary. Thus, an in vitro model would not be possible; instead, such experiments would have to be conducted, for example, in an animal model with injection of tumor cells, and subsequent irradiation of the tumor cells and the surrounding skin. Therefore, the present study provides good preliminary data, although further research into the aforementioned mechanisms is of much interest.
Another interesting question raised by this setup is, if a pre-irradiation of tissues has an effect on the efficacy of cytostatic agents on co-cultured tumor cells. As a cytostatic therapy often is the only treatment available for already irradiated and inoperable tumors, more information concerning the interactions of irradiated tissue and cytostatic agents would be of much interest and is the aim of a study being undertaken at the University Hospital of Würzburg.
In conclusion, pre-irradiation of tissue changes the secretory abilities of the fibroblasts cultured from it as well as the way these fibroblasts affect the growth of co-cultured tumor cells in vitro. While major differences in the secretion of IL-6 and IL-8 were observed, the exact mechanisms of the changes caused by previous irradiation are still largely unknown. Thus, irradiated tumor stroma should be examined more closely to gain insight into the long-term effects of irradiation that may not yet have become obvious. Future studies should be conducted to specifically examine differences between irradiated fibroblast cell lines in vitro and fibroblasts cultured from pre-irradiated skin, as well as the effect of irradiated fibroblasts on the efficiency of cytostatic agents in a co-culture and the involvement of EMT in pre-irradiated tissue. Furthermore, to evaluate these findings in a more clinical setting, an animal study with irradiation of cancer cells and the surrounding tissue should be conducted.
References
- 1.Parkin DM, Bray F, Ferlay J, Pisani P. Global cancer statistics,2002. CA Cancer J Clin. 2005;55:74–108. doi: 10.3322/canjclin.55.2.74. [DOI] [PubMed] [Google Scholar]
- 2.Chan GG, Tai BC, Liang S, Lim DT, Soo KC. Squamous cell carcinoma of the head and neck (HNSCC)-multi-modality treatment and impact on survival. Asian J Surg. 2002;25:35–40. [PubMed] [Google Scholar]
- 3.Kalluri R. Basement membranes: Structure, assembly and role in tumour angiogenesis. Nat Rev Cancer. 2003;3:422–433. doi: 10.1038/nrc1094. [DOI] [PubMed] [Google Scholar]
- 4.Tripathi M, Billet S, Bhowmick NA. Understanding the role of stromal fibroblasts in cancer progression. Cell Adh Migr. 2012;6:231–235. doi: 10.4161/cam.20419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Joyce JA, Pollard JW. Microenvironmental regulation of metastasis. Nat Rev Cancer. 2009;9:239–252. doi: 10.1038/nrc2618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Tlsty TD, Hein PW. Know thy neighbor: Stromal cells can contribute oncogenic signals. Curr Opin Genet Dev. 2001;11:54–59. doi: 10.1016/S0959-437X(00)00156-8. [DOI] [PubMed] [Google Scholar]
- 7.Mueller MM, Fusenig NE. Friends or foes-bipolar effects of the tumour stroma in cancer. Nat Rev Cancer. 2004;4:839–849. doi: 10.1038/nrc1477. [DOI] [PubMed] [Google Scholar]
- 8.Higashikawa K, Yoneda S, Taki M, Shigeishi H, Ono S, Tobiume K, Kamata N. Gene expression profiling to identify genes associated with high-invasiveness in human squamous cell carcinoma with epithelial-to-mesenchymal transition. Cancer Lett. 2008;264:256–264. doi: 10.1016/j.canlet.2008.01.045. [DOI] [PubMed] [Google Scholar]
- 9.Hamada H, Kobune M, Nakamura K, Kawano Y, Kato K, Honmou O, Houkin K, Matsunaga T, Niitsu Y. Mesenchymal stem cells (MSC) as therapeutic cytoreagents for gene therapy. Cancer Sci. 2005;96:149–156. doi: 10.1111/j.1349-7006.2005.00032.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Yang F, Cho SW, Son SM, Bogatyrev SR, Singh D, Green JJ, Mei Y, Park S, Bhang SH, Kim BS, et al. Genetic engineering of human stem cells for enhanced angiogenesis using biodegradable polymeric nanoparticles. Proc Natl Acad Sci USA. 2010;107:3317–3322. doi: 10.1073/pnas.0905432106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Affolter A, Schmidtmann I, Mann J, Brieger J. Cancer-associated fibroblasts do not respond to combined irradiation and kinase inhibitor treatment. Oncol Rep. 2013;29:785–790. doi: 10.3892/or.2012.2180. [DOI] [PubMed] [Google Scholar]
- 12.Grey B, Coppey J, Little JB. Modulation of clonogenicity, growth, and radiosensitivity of three human epidermoid tumor cell lines by a fibroblastic environment. J Rad Onc Biol Phys. 1996;34:1061–1071. doi: 10.1016/0360-3016(95)02175-2. [DOI] [PubMed] [Google Scholar]
- 13.Tsai KK, Chuang EY, Little JB, Yuan ZM. Cellular mechanisms for low-dose ionizing radiation-induced perturbation of the breast tissue microenvironment. Cancer Res. 2005;65:6734–6744. doi: 10.1158/0008-5472.CAN-05-0703. [DOI] [PubMed] [Google Scholar]
- 14.Rangan SR. A new human cell line (FaDu) from a hypopharyngeal carcinoma. Cancer. 1972;29:117–121. doi: 10.1002/1097-0142(197201)29:1<117::AID-CNCR2820290119>3.0.CO;2-R. [DOI] [PubMed] [Google Scholar]
- 15.Mosmann T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J Immunol Methods. 1983;65:55–63. doi: 10.1016/0022-1759(83)90303-4. [DOI] [PubMed] [Google Scholar]
- 16.Orimo A, Weinberg RA. Stromal fibroblasts in cancer: A novel tumor-promoting cell type. Cell Cycle. 2006;5:1597–1601. doi: 10.4161/cc.5.15.3112. [DOI] [PubMed] [Google Scholar]
- 17.Mishra PJ, Mishra PJ, Humeniuk R, Medina DJ, Alexe G, Mesirov JP, Ganesan S, Glod JW, Banerjee D. Carcinoma-associated fibroblast-like differentiation of human mesenchymal stem cells. Cancer Res. 2008;68:4331–4339. doi: 10.1158/0008-5472.CAN-08-0943. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Dudás J, Bitsche M, Schartinger V, Falkeis C, Sprinzl GM, Riechelmann H. Fibroblasts produce brain-derived neurotrophic factor and induce mesenchymal transition of oral tumor cells. Oral Oncol. 2011;47:98–103. doi: 10.1016/j.oraloncology.2010.11.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Koontongkaew S, Amornphimoltham P, Yapong B. Tumor-stroma interactions influence cytokine expression and matrix metalloproteinase activities in paired primary and metastatic head and neck cancer cells. Cell Biol Int. 2009;33:165–173. doi: 10.1016/j.cellbi.2008.10.009. [DOI] [PubMed] [Google Scholar]
- 20.Olumi AF, Grossfeld GD, Hayward SW, Carroll PR, Tlsty TD, Cunha GR. Carcinoma-associated fibroblasts direct tumor progression of initiated human prostatic epithelium. Cancer Res. 1999;59:5002–5011. doi: 10.1186/bcr138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.van Roozendaal KE, Klijn JG, van Ooijen B, Claassen C, Eggermont AM, Henzen-Logmans SC, Foekens JA. Differential regulation of breast tumor cell proliferation by stromal fibroblasts of various breast tissue sources. Int J Cancer. 1996;65:120–125. doi: 10.1002/(SICI)1097-0215(19960103)65:1<120::AID-IJC20>3.0.CO;2-J. [DOI] [PubMed] [Google Scholar]
- 22.Ohuchida K, Mizumoto K, Murakami M, Qian LW, Sato N, Nagai E, Matsumoto K, Nakamura T, Tanaka M. Radiation to stromal fibroblasts increases invasiveness of pancreatic cancer cells through tumor-stromal interactions. Cancer Res. 2004;64:3215–3222. doi: 10.1158/0008-5472.CAN-03-2464. [DOI] [PubMed] [Google Scholar]
- 23.Meltzer J, Ahmed SA, Archambeau JO. The development of metastases within a field of previous irradiation: A case report. Cancer. 1981;48:717–720. doi: 10.1002/1097-0142(19810801)48:3<717::AID-CNCR2820480310>3.0.CO;2-G. [DOI] [PubMed] [Google Scholar]
- 24.Baker DG, Morgan EP, Persing JA. Squamous cell carcinoma growth in irradiated tissue: A murine model for quantitative assessment of treatment. Ann Plast Surg. 1995;35:171–177. doi: 10.1097/00000637-199508000-00010. [DOI] [PubMed] [Google Scholar]
- 25.Chen Z, Malhotra PS, Thomas GR, Ondrey FG, Duffey DC, Smith CW, Enamorado I, Yeh NT, Kroog GS, Rudy S, et al. Expression of proinflammatory and proangiogenic cytokines in patients with head and neck cancer. Clin Cancer Res. 1999;5:1369–1379. [PubMed] [Google Scholar]
- 26.Lathers DM, Young MR. Increased aberrance of cytokine expression in plasma of patients with more advanced squamous cell carcinoma of the head and neck. Cytokine. 2004;25:220–228. doi: 10.1016/j.cyto.2003.11.005. [DOI] [PubMed] [Google Scholar]
- 27.Todorović-Raković N, Milovanović J. Interleukin-8 in breast cancer progression. J Interferon Cytokine Res. 2013;33:563–570. doi: 10.1089/jir.2013.0023. [DOI] [PMC free article] [PubMed] [Google Scholar]