

Abstract
The natural extract artemisinin and its derivatives have good anticancer activity. The present study aimed to investigate the in vitro inhibitory effects of combined dihydroartemisinin (DHA) and doxorubicin (DOX) treatment on a variety of tumor cell lines (HeLa, OVCAR-3, MCF-7, PC-3 and A549), as well as the underlying mechanisms. In addition, the in vivo effects of DHA and DOX were evaluated using a mouse HeLa tumor model. The HeLa, OVCAR-3, MCF-7, PC-3 and A549 cells were treated with a combination of DHA and DOX, and the effect on cell viability was detected by Cell Counting kit-8. The cells were observed under a fluorescence microscope after staining with Hoechst 33258 dye to observe morphological changes in the nuclei in order to determine whether the cells in the treatment group exhibited apoptosis. Apoptosis of the cells was further detected by flow cytometry, and statistical analysis was performed. The specific inhibitors of caspase-3, −8 and −9 were used to determine the intrinsic and extrinsic pathways of cell apoptosis. The cervical cancer HeLa cells treated with the combination of DHA and DOX showed up to a 91.5% decrease in viability, which was higher than that of the same cells treated with DHA or DOX alone at the same concentration, respectively (P<0.01). The optimal concentrations of the drugs used in combination were DHA at 10 µg/ml and DOX at 10 µg/ml. DHA + DOX also had a significant inhibitory effect on the ovarian cancer (OVCAR-3), breast cancer (MCF-7), lung cancer (A549) and prostate cancer (PC-3) cells. The images observed under fluorescence microscope after Hoechst 33258 staining showed marked pyknosis in the cells treated with DHA + DOX, similar to that when treated with DHA or DOX alone, which is typical in apoptosis. As determined by flow cytometry, the apoptotic rate of the cells treated with DHA + DOX at optimal concentrations was up to 90%, which was significantly higher than that of the cells treated with DHA or DOX alone at the same concentration. Caspase-9 and −3 inhibitors significantly increased the viability of the cells treated with DHA + DOX. At 6 days post-intratumoral injection of DHA + DOX, the tumor volume was markedly reduced. In vivo toxicity results revealed that the combination of the drugs had basically no effect on the body weight of the mice and had no significant toxicity on the liver, spleen, kidneys and heart of the animals. Overall, the combination of DHA and DOX markedly inhibited the viability of the HeLa, OVCAR-3, MCF-7, PC-3 and A549 cells, and acted on the HeLa cells through the intrinsic apoptotic pathway mediated by caspase-9 and caspase-3. DHA + DOX also had a significant treatment effect in vivo. This study provides a novel idea for the development of a clinical medication against several types of cancer.
Keywords: dihydroartemisinin, doxorubicin, drug combination, apoptosis, tumor cells
Introduction
Cancer is the biggest killer threatening human health globally (1). At present, the main treatment approaches include radiotherapy, chemotherapy and combination therapy (2). However, radiotherapy has significant toxicity and side effects on healthy tissues of the body, and the toxicity and side effects of chemotherapy adversely affect the impact of the treatment to a certain degree (3). Therefore, the choice of drugs for chemotherapy has become key in tumor treatment. In recent years, an emphasis has been placed upon individualized medication, and improvement of drug sensitivity and treatment effect through combination therapy in tumor treatment (4).
Dihydroartemisinin (DHA) is an active derivative of the first-line anti-malarial drug artemisinin, which has been extracted from the goldenrod Artemisia annua in China for the first time (5,6). Due to the quick activity, effectiveness and safety of artemisinin and its derivative, the World Health Organization promotes their application in the treatment of severe and drug-resistant malaria (7). It has been demonstrated that artemisinin and its derivative show an antitumor effect and that they act on a series of cell biochemical processes, inhibiting proliferation, inducing apoptosis and oxidative stress, and acting in anti-angiogenesis (8). Doxorubicin (DOX), also a currently-used antitumor antibiotic, can inhibit the synthesis of RNA and DNA, with the strongest inhibitory effect on RNA. The drug is efficacious against multiple types of tumor, with a wide antitumor spectrum (9,10).
In the present study, the natural drug DHA and the chemical drug DOX were combined to act on HeLa tumor cells, and the survival rate and the mechanism of death were detected. The activity of the DHA + DOX combination in the OVCAR-3, MCF-7, PC-3 and A549 cells was also investigated in the study. In addition, the in vivo treatment effect of DHA + DOX was studied with an intratumoral method.
Materials and methods
Experimental materials and reagents
The cervical cancer (HeLa), ovarian cancer (OVCAR-3), breast cancer (MCF-7), lung cancer (A549) and prostate cancer (PC-3) cells were purchased from the American Type Culture Collection (Manassas, VA, USA). Flasks and 96-well plates for cell culture were purchased from Corning Inc. (Corning, NY, USA). The Dulbecco's modified Eagle's medium (DMEM) was obtained from Thermo Fisher Scientific Inc., (Gibco; Waltham, MA, USA) and the new-born calf serum was purchased from Hangzhou Sijiqing Biological Engineering Materials Co., Ltd. (Hangzhou, China). Cell Counting kit-8 (CCK-8) was bought from Dojindo Molecular Technologies Inc. (Kumamoto, Japan). The DHA, DOX, Hoechst 33258 dye, broad-spectrum caspase inhibitors Z-IETD-fmk, Z-LEHD-fmk and Z-DQMD-fmk, and staurosporin (STS) were purchased from Sigma-Aldrich (St. Louis, MO, USA). The fluoroscein isothiocyanate (FITC)-Annexin V/propidium iodide (PI) apoptosis kit was purchased from eBioscience Inc. (San Diego, CA, USA).
Experimental instruments
The flow cytometer (FACSCanto™ II) was obtained from BD Biosciences (Franklin Lakes, NJ, USA). The laser scanning confocal microscope (LSM510/ConfoCor2) was purchased from Zeiss GmbH (Jena, Germany). The microplate reader (Infinite M200) was purchased from Tecan Austria GmbH (Grödig, Austria).
Cell culture
The HeLa, OVCAR-3, MCF-7, A549 and PC-3 cells were cultivated in the DMEM liquid medium containing 10% new-born calf serum. Once 80–90% confluence was reached, 0.25% trypsin was used to digest the cells for passaging and then the cells were placed into a 5% CO2 incubator at 37°C for culture, ready for use in subsequent experiments.
Determination of cell viability with CCK-8
Cell suspension (100 µl; 4,000–5,000 cells/well) was inoculated into the 96-well plate and then placed into a 37°C, 5% CO2, saturated humidity incubator for 24-h culture. Next, the medium was replaced by fresh medium, and the samples were assigned into the control group and the treatment groups, with 4 parallel wells in each group. Cells in the treatment groups were treated with 0.5, 1, 2.5, 5, 10 or 20 µg/ml DHA and/or DOX. After treatment, the medium in the 96-well plate was replaced with fresh medium, and 10% CCK-8 working fluid was added to each well. After a 30-min culture in the incubator, a microplate reader was used to determine absorbance [optical density at a wavelength of 450 nm (OD450)], and the OD450 value was positively correlated with the cell survival rate.
Hoechst 33258 staining
After the HeLa cells underwent serial subcultivation for 24 h, and were treated with DHA alone (10 µg/ml), DOX alone (10 µg/ml) or DHA (10 µg/ml) + DOX (10 µg/ml) for 24 h, PBS was used for washing three times. Next, Hoechst 33258 dye at a final concentration of 1 µM was added for 20 min for staining, PBS was used for washing three times and the cell karyotype was observed under a laser scanning confocal microscope. The excitation light source was a mercury lamp, and the amplification factor of the fluorescence microscope was 400-fold.
Flow cytometry with FITC-Annexin V/PI double staining
Flow cytometry with FITC-Annexin V/PI double staining is considered ideal for the quantitative determination of cell apoptosis. The cells were inoculated in a six-well plate, ensuring that the number of cells was 1×106/well. After the cells were treated with DHA alone (10 µg/ml), DOX alone (10 µg/ml) or DHA (10 µg/ml) + DOX (10 µg/ml) for 24 h, 5 µl FITC-Annexin V (at a concentration of 10 µM) was added and then the cells were incubated for 30 min in the dark. PI (5 µl; at a concentration of 10 µg/ml) was then added and the cells were incubated again for 10 min in the dark. A 300-mesh sieve was used to filter the cells and analysis was performed on a flow cytometer. The light source was a 488-nm argon ion laser.
Tumor inoculation and treatment
A total of 35 male BALB/c nude mice (age, 6–8 weeks; weight, ~25 g) were obtained from the Experimental Animal Center of Southern Medical University (Guangzhou, China). The present study was approved by the institutional review board. The mice were maintained at 25°C and 70% relative humidity, under a 12-h light/dark cycle, with ad libitum access to food and water. To establish an animal HeLa tumor model, 100 µl PBS containing 2×106 HeLa cells was injected subcutaneously into the back of each mouse. The tumor volume was measured once every 2 days and calculated using the formula: Tumor volume = 0.5 × long axis × short axis2. The weight of the mice was also measured once every 2 days. When the tumor volume was ~100 mm3, the mice were divided into five groups (7 mice/group), as follows: i) The control group (no injection); ii) the saline group, in which the mice were intratumorally injected with 100 µl normal saline on day 1; iii) the DHA group, in which mice received intratumoral injection with 15 mg/kg DHA; iv) the DOX group, in which mice received intratumoral injection with 15 mg/k DOX; and v) the DHA + DOX group, in which mice received intratumoral injection with 15 mg/kg DHA and 15 mg/kg DOX.
Hematoxylin and eosin (HE) staining of vital organs in mice
After 35 days, the mice were anesthetized with 10% chloral hydrate (300 mg/kg; Chengdu Kelong Chemicals, Co., Ltd., Chengdu, China) and sacrificed by cervical dislocation. The organs of the mice were isolated and placed into 4% formaldehyde solution for fixation, then gradually dehydrated and finally embedded in paraffin. Sections (5-µm) were cut for HE staining and organ toxicity analysis.
Statistical analysis
Data are presented as the mean ± standard deviation. One-way analysis of variance followed by the Holm-Šídák test was used to determine differences in cell viability and the ratios of apoptotic cells among the different groups. Each experiment was performed in triplicate, and statistical analysis was performed with the SPSS 13.0 software (SPSS Inc., Chicago, IL, USA). P≤0.05 was used to indicate a statistically significant difference.
Results
Effect of the combination of DHA and DOX at different concentrations on cell viability
CCK-8 is commonly used to detect cell viability. As shown in Fig. 1A, the viability of the HeLa cells had a tendency to decrease as the concentrations of DHA and DOX increased. After the cells were treated with DHA alone at 0.5, 1, 2.5, 5, 10 and 20 µg/ml for 24 h, the viability ranged between 45 and 95%, and after treatment with DOX alone at 0.5, 1, 2.5, 5, 10 and 20 µg/ml for 24 h, the viability ranged between 42 and 94%. After treatment with DHA + DOX, the cell viability decreased up to 90%. The combination of 10 µg/ml DHA and 10 µg/ml DOX resulted in a decrease of 89% in cell viability, which was significantly different from that caused by the combination of 5 µg/ml DHA and 5 µg/ml DOX (P=0.008; Fig. 1), and was comparable with that caused by the combination of 20 µg/ml DHA and 20 µg/ml DOX (P=0.643). Therefore 10 µg/ml DHA plus 10 µg/ml DOX (DHA + DOX) was the optimal concentration combination. In Fig. 1B, combination of the drugs at the optimal concentrations was used to treat the OVCAR-3, MCF-7, PC-3 and A549 cells for 24 h, and a good inhibitory effect was shown for these tumor cells, with statistically significant differences compared with the control group (P=0.002, 0.001, 0.003 and 0.001, respectively).
Figure 1.
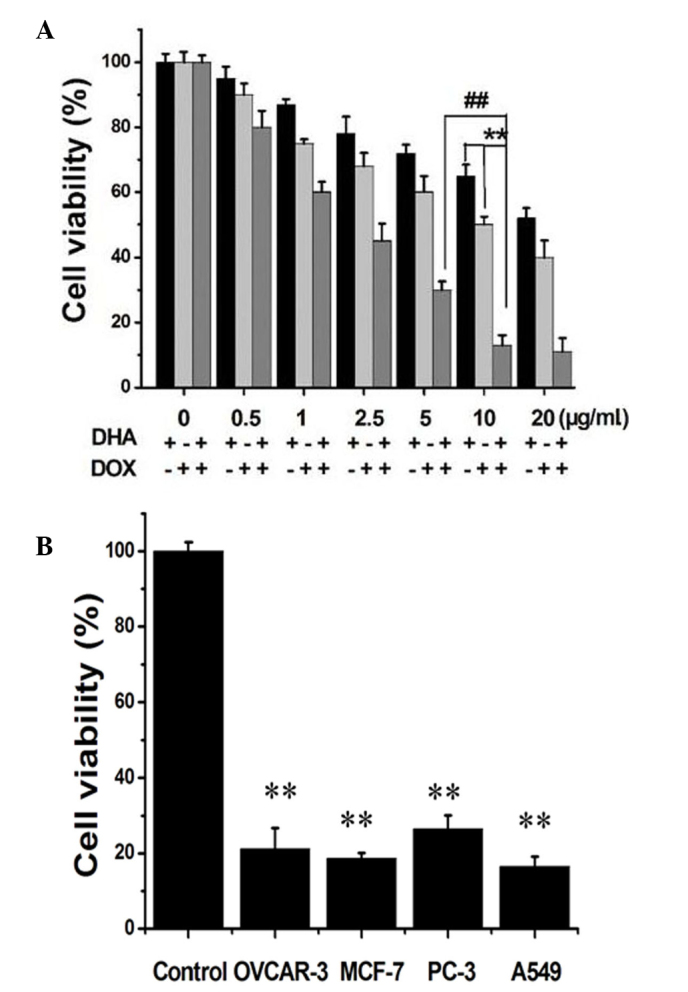
(A) Effect on the viability of the HeLa cells after 24 h of treatment with DHA and DOX at different concentrations. (B) The effect on the viability of the OVCAR-3, MCF-7, PC-3 and A549 cells after 24 h of treatment with DHA + DOX (##P<0.01 and **P<0.05; n=4). DHA, dihydroartemisinin; DOX, doxorubicin.
Pattern of DHA + DOX-induced cell death
In order to verify the pattern of cell death induced by the combination of the drugs, Hoechst 33258 staining was used first to observe the morphological changes in the cell nuclei. Under confocal fluorescence microscope, it was observed that the cells treated with DHA + DOX and DHA or DOX alone showed marked apoptosis, with chromatin agglutination and karyopyknosis (Fig. 2). In the blank control group, the cell fluorescence was relatively superficial and uniform, and the structure of the cell nuclei was normal. When compared with the cells treated with the apoptosis-positive control reagent, STS, the results shown with the combination of the drugs was similar to those of the positive control. These results indicated that DHA + DOX may have induced a type of programmed cell death, which may have been apoptosis induced by the synergistic action of DHA and DOX.
Figure 2.
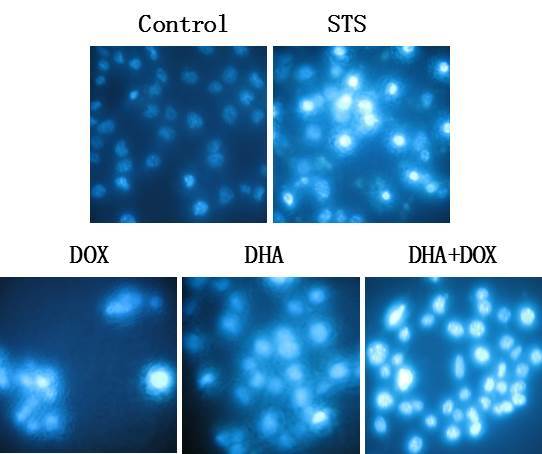
Hoechst 33258 dye cell images after the HeLa cells were treated with DOX or DHA alone, or DOX+DHA for 24 h. HeLa cells treated with STS were used as a positive control. DHA, dihydroartemisinin; DOX, doxorubicin; STS, staurosporin.
In order to further confirm that the cell death induced by DHA + DOX was apoptosis, Annexin V/PI double staining was used to detect the extroversion of phosphatidyl serine on the cell membrane and the PI-specific stained cell nuclei after rupture of the membrane, indicating apoptosis (11,12). As shown in Fig. 3, after treatment with DHA + DOX, the percentage of apoptotic cells (often considered to be the total of cells of Q2 and Q4) increased from 3% (control) to 90% (24 h), but the percentage of necrotic cells increased inconspicuously. The positive control indicated that Q2 + Q4 did represent apoptosis. Among the cells treated with DHA or DOX alone, there were also apoptotic cells, but the apoptotic rate was significantly lower than that of the combination group (P=0.005 and 0.006, respectively; Fig. 4). These results indicated that the DHA-induced cell death was apoptosis.
Figure 3.
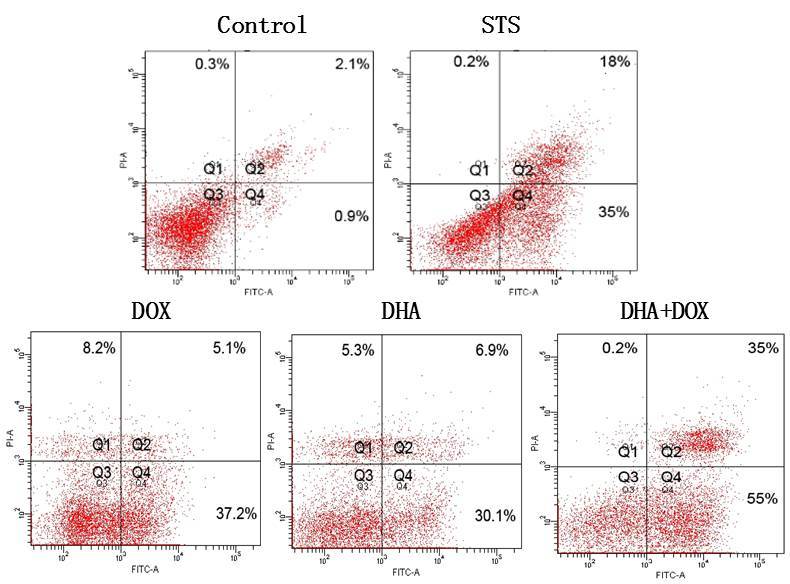
Cell death analysis. Following treatment with DOX or DHA alone, or DOX+DHA for 24 h, the HeLa cells were stained with Annexin V-FITC and then analyzed by flow cytometry for the extroversion of phosphatidyl serine on the cell membrane. DHA, dihydroartemisinin; DOX, doxorubicin; STS, staurosporin; FITC, fluoroscein isothiocyanate.
Figure 4.
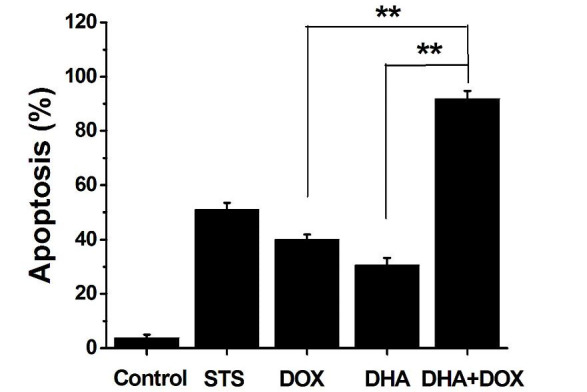
Ratio of apoptotic cells to the total number of cells. The number of apoptotic cells equals the sum of the cells in the Q2 and Q4 regions in Fig. 3 (**P<0.01; n=4). DHA, dihydroartemisinin; DOX, doxorubicin; STS, staurosporin.
Intrinsic and extrinsic pathways through which DHA + DOX causes cell apoptosis
The specific inhibitors of caspase-3, −8 and −9, namely, z-DQMD-fmk, z-IETD-fmk and z-LEHD-fmk, respectively, were used to determine whether caspase-3, −8 or −9 were involved in the induction of cell apoptosis by DHA + DOX. Caspase-8 is an essential apoptotic factor in the extrinsic apoptotic pathway, and caspase-3 and −9 are factors in the intrinsic apoptotic pathway (13,14). As shown in Fig. 5, unlike the caspase-8 inhibitor z-IETD-fmk, treatment with z-DQMD-fmk and z-LEHD-fmk increased the viability of the cells treated with DHA + DOX (P=0.036 and 0.039, respectively; Fig. 5), suggesting that caspase-3 and −9 were involved in the induction of cell apoptosis by DHA + DOX. Based on this, it was speculated that DHA + DOX acted through the intrinsic apoptotic pathway.
Figure 5.
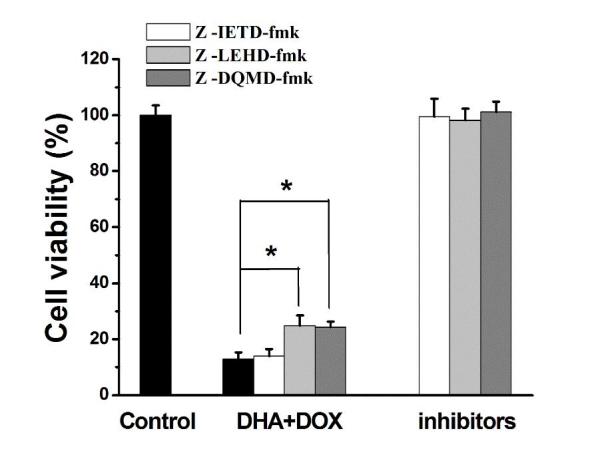
Effect of caspase inhibitors on DHA + DOX-induced cell apoptosis. *P<0.05 compared with the group treated with DHA + DOX only. DHA, dihydroartemisinin; DOX, doxorubicin; STS, staurosporin.
In vivo treatment effect of DHA + DOX
The heterologous HeLa tumor model was established to investigate whether DHA + DOX had the effect of tumor elimination in vivo. DHA + DOX (15 mg/kg) was administered by intratumoral injection, and the treatment effect was evaluated through the measurement of tumor volume. As shown in Fig. 6, there was a significant inhibitory effect after 6 days of intratumoral injections with DHA + DOX (n=7). After injection of DHA or DOX alone, the tumor size was significantly inhibited, as compared with the control (P=0.042 and 0.043, respectively) and saline groups (P=0.040 and 0.042, respectively), although tumor size was inhibited to the greatest extent in the DHA + DOX group (P=0.031, 0.033, 0.035 and 0.036 vs. the control, saline, DHA and DOX groups, respectively).
Figure 6.
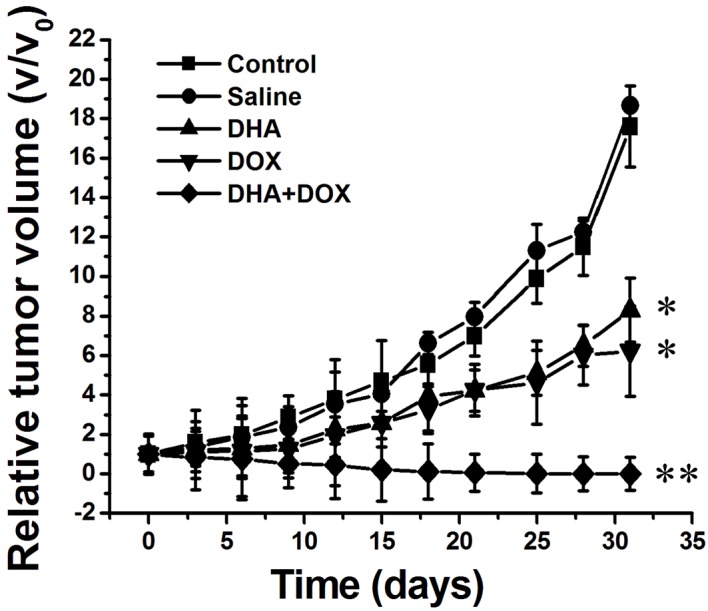
Changes in the relative tumor volume of tumor-bearing mice following injection of normal saline, DHA, DOX and DHA + DOX, respectively. DHA, dihydroartemisinin; DOX, doxorubicin; STS, staurosporin. *P<0.05, **P<0.01 vs. the control and saline groups.
In vivo toxicity detection after intratumoral injection of DHA + DOX
The in vivo toxicity of drugs has always been a clinical concern. In the present study, it was determined through monitoring changes in the weight of the mice after drug administration and through pathological analysis of the sections of vital organs after 30 days of administration. As shown in Fig. 7A, compared with the control group, after injection of DHA or DOX alone, or DHA + DOX, the weight of the mice did not significantly decrease, but was actually increasing slowly. The HE staining images (Fig. 7B) showed that after intratumoral injection in all treatment groups, no toxicity or side effects were observed in the liver, kidneys, spleen and heart.
Figure 7.

In vivo toxicity detection after intratumoral injection of normal saline, DHA, DOX or DHA + DOX. (A) Weight changes in the mice. (B) Hematoxylin and eosin-stained images of the spleen, liver, kidney and heart sections of the mice (magnification, ×400). DHA, dihydroartemisinin; DOX, doxorubicin.
Discussion
DHA, extracted from goldenrod Artemisia annua in China for the first time, is a novel first-line anti-malarial drug that has been also been revealed to exhibit antitumor effects (15–17). Park et al (18) found that DHA exhibited no clear cytotoxicity on normal human breast cells, but that it exhibited significant cytotoxicity on human breast cancer cells. DOX is a wide-spectrum antitumor antibiotic and a cell cyclic non-specific agent that is mostly combined with other anticancer drugs (19). In the present study, four experiments were designed to investigate the lethal effect of DHA + DOX on human cervical cancer cells (HeLa) and the corresponding mechanism. The first investigation studied the effect of DHA + DOX at different concentrations on the viability of the HeLa cells to determine the optimal concentrations and synergistic lethal effect of the combination. The lethal effect of DHA + DOX at optimal concentrations on the OVCAR-3, MCF-7, PC-3 and A549 cells was also assessed. These results indicated that, as compared with either drug alone, combined DHA and DOX treatment showed an enhanced lethal effect on HeLa cells. Furthermore, this combination inhibited the viability of OVCAR-3, MCF-7, PC-3 and A549 cells.
Apoptosis has important biological significance in the development and progression of tumors, and numerous chemotherapy drugs exert an antitumor effect by inducing the apoptosis of tumor cells. A previous study revealed that DHA could induce apoptosis through the intrinsic pathway (8). The second experiment in the present study investigated whether DHA + DOX also induced tumor cell apoptosis. The nuclei-specific Hoechst 33258 dye was used to check the changes in cell nuclei after administration of DHA + DOX, and flow cytometry was employed to determine the mode of cell death. The results indicated that after administration of DHA + DOX, karyopyknosis of the HeLa cells occurred, inducing apoptosis, with an apoptotic rate of 90%. This indicated that the administration of DHA + DOX synergistically improved the rate of apoptosis in the cells.
Caspase proteins are the primary factor that induces changes in the intracellular biochemical and morphological properties, with caspase-8 as a typical apoptotic factor in the extrinsic pathway, and caspase-3 and −9 as typical apoptotic factors in the intrinsic pathway (20). The third experiment in the present study used specific inhibitors of caspase-3, −8 and −9 to detect whether these caspases were involved in the apoptosis induced by the administration of DHA + DOX. The results indicated that caspase-8 was not involved in the apoptosis, but that caspase-3 and −9 were involved in the apoptosis of the HeLa cells, suggesting that the administration of DHA + DOX induced apoptosis through the intrinsic pathway.
The present study also investigated the in vivo treatment effect of DHA + DOX on mice with HeLa tumors. Intratumoral injection is increasingly being applied in the treatment of tumors (21). In the present study, intratumoral injection was employed to treat the tumor-bearing mice, and it was observed that the tumor elimination effect of the combined medication was significantly better than that of separate administration. The results of the in vivo experiment indicated that combined medication exhibited no clear toxicity in the mice. Previous studies have demonstrated a high toxicity for DOX at high concentrations in the hearts of mice. However, the present study showed that DHA + DOX could not only reduce drug toxicity shortly after administration, but that it could also considerably increase the treatment effect of the drugs.
In conclusion, DHA + DOX exhibits a significant synergistic inhibitory effect against a variety of tumor cell lines. The present study showed that, in Hela cells, the lethal mechanism behind the combined DHA + DOX medication was apoptosis induced by the intrinsic pathway, with involvement of caspase-3 and −9. The in vivo tumor elimination effect of the combined medication was also significant. The results of this study provide a novel idea for clinical chemotherapy and the experimental research into multiple cancers, including human cervical cancer.
References
- 1.Parkin DM, Bray F, Ferlay J, Pisani P. Estimating the world cancer burden: Globocan 2000. Int J Cancer. 2001;94:153–156. doi: 10.1002/ijc.1440. [DOI] [PubMed] [Google Scholar]
- 2.Chou TC. Theoretical basis, experimental design, and computerized simulation of synergism and antagonism in drug combination studies. Pharmacol Rev. 2006;58:621–681. doi: 10.1124/pr.58.3.10. [DOI] [PubMed] [Google Scholar]
- 3.Roth AD, Maibach R, Martinelli G, Fazio N, Aapro MS, Pagani O, Morant R, Borner MM, Herrmann R, Honegger H, et al. Docetaxel (Taxotere)-cisplatin (TC): An effective drug combination in gastric carcinoma. Swiss group for clinical cancer research (SAKK), and the European institute of oncology (EIO) Ann Oncol. 2000;11:301–306. doi: 10.1023/A:1008342013224. [DOI] [PubMed] [Google Scholar]
- 4.Martello LA, McDaid HM, Regl DL, Yang CP, Meng D, Pettus TR, Kaufman MD, Arimoto H, Danishefsky SJ, Smith AB, III, Horwitz SB. Taxol and discodermolide represent a synergistic drug combination in human carcinoma cell lines. Clin Cancer Res. 2000;6:1978–1987. [PubMed] [Google Scholar]
- 5.Huang FY, Lee TW, Chang CH, Chen LC, Hsu WH, Chang CW, Lo JM. Evaluation of (188) Re-labeled PEGylated nanoliposome as a radionuclide therapeutic agent in an orthotopic glioma-bearing rat model. Int J Nanomedicine. 2015;10:463–473. doi: 10.2147/IJN.S75955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Karavitis J, Zhang M. COX2 regulation of breast cancer bone metastasis. Oncoimmunology. 2013;2:e23129. doi: 10.4161/onci.23129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Haynes RK. Artemisinin and derivatives: The future for malaria treatment. Curr Opin Infect Dis. 2001;14:719–726. doi: 10.1097/00001432-200112000-00010. [DOI] [PubMed] [Google Scholar]
- 8.Guo Y, Shi X, Li L. Effects of dihydroartemisinin on ovarian cancer HO-8910 cells and expression of Akt. Zhong Guo Yi Xue Qian Yan Za Zhi. 2012;7:17–18. [Google Scholar]
- 9.Howe LR. Inflammation and breast cancer. Cyclooxygenase/prostaglandin signaling and breast cancer. Breast Cancer Res. 2007;9:210. doi: 10.1186/bcr1678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Farh KK, Grimson A, Jan C, Lewis BP, Johnston WK, Lim LP, Burge CB, Bartel DP. The widespread impact of mammalian MicroRNAs on mRNA repression and evolution. Science. 2005;310:1817–1821. doi: 10.1126/science.1121158. [DOI] [PubMed] [Google Scholar]
- 11.Schutte B, Nuydens R, Geerts H, Ramaekers F. Annexin V binding assay as a tool to measure apoptosis in differentiated neuronal cells. J Neurosci Methods. 1998;86:63–69. doi: 10.1016/S0165-0270(98)00147-2. [DOI] [PubMed] [Google Scholar]
- 12.Cheung JY, Ong RC, Suen YK, Ooi V, Wong HN, Mak TC, Fung KP, Yu B, Kong SK. Polyphyllin D is a potent apoptosis inducer in drug-resistant HepG2 cells. Cancer Lett. 2005;217:203–211. doi: 10.1016/j.canlet.2004.06.042. [DOI] [PubMed] [Google Scholar]
- 13.Zhang JM, Wang HC, Wang HX, Ruan LH, Zhang YM, Li JT, Tian S, Zhang YC. Oxidative stress and activities of caspase-8, −9, and-3 are involved in cryopreservation-induced apoptosis in granulosa cells. Eur J Obstet Gynecol Reprod Biol. 2013;166:52–55. doi: 10.1016/j.ejogrb.2012.09.011. [DOI] [PubMed] [Google Scholar]
- 14.Hsiao PC, Lee WJ, Yang SF, Tan P, Chen HY, Lee LM, Chang JL, Lai GM, Chow JM, Chien MH. Nobiletin suppresses the proliferation and induces apoptosis involving MAPKs and caspase-8/-9/-3 signals in human acute myeloid leukemia cells. Tumor Biol. 2014;35:11903–11911. doi: 10.1007/s13277-014-2457-0. [DOI] [PubMed] [Google Scholar]
- 15.Kong R, Jia G, Cheng ZX, Wang YW, Mu M, Wang SJ, Pan SH, Gao Y, Jiang HC, Dong DL, Sun B. Dihydroartemisinin enhances Apo2 L/TRAIL-mediated apoptosis in pancreatic cancer cells via ROS-mediated up-regulation of death receptor 5. PLoS One. 2012;7:e37222. doi: 10.1371/journal.pone.0037222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Sun Q, Teong B, Chen IF, Chang SJ, Gao J, Kuo SM. Enhanced apoptotic effects of dihydroartemisinin-aggregated gelatin and hyaluronan nanoparticles on human lung cancer cells. J Biomed Mater Res B Appl Biomater. 2014;102:455–462. doi: 10.1002/jbm.b.33023. [DOI] [PubMed] [Google Scholar]
- 17.Posobiec LM, Clark RL, Bushdid PB, Laffan SB, Wang KF, White TE. Dihydroartemisinin (DHA) treatment causes an arrest of cell division and apoptosis in rat embryonic erythroblasts in whole embryo culture. Birth Defects Res B Dev Reprod Toxicol. 2013;98:445–458. doi: 10.1002/bdrb.21088. [DOI] [PubMed] [Google Scholar]
- 18.Park J, Lai HC, Singh M, Sasaki T, Singh NP. Development of a dihydroartemisinin-resistant Molt-4 leukemia cell line. Anticancer Res. 2014;34:2807–2810. [PubMed] [Google Scholar]
- 19.Wang Y, Xu L, Duan Z, Cao K, Luo J, Xu Y, Shi P. Effects of ADM on cell apoptosis and expression of FAK mRNA in K562. J Trop Med. 2012;12:1195–1198. [Google Scholar]
- 20.Brentnall M, Rodriguez-Menocal L, De Guevara RL, Cepero E, Boise LH. Caspase-9, caspase-3 and caspase-7 have distinct roles during intrinsic apoptosis. BMC Cell Biol. 2013;14:32. doi: 10.1186/1471-2121-14-32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Wan L, Zheng X. Expression pattern and function of SATB1 in the invasiveness of thyroid carcimoma. Chin J Biochem Pharm. 2014;34:18–20. [Google Scholar]