

ABSTRACT
HIV-1 is able to elicit broadly potent neutralizing antibodies in a very small subset of individuals only after several years of infection, and therefore, vaccines that elicit these types of antibodies have been difficult to design. The RV144 trial showed that moderate protection is possible and that this protection may correlate with antibody-dependent cellular cytotoxicity (ADCC) activity. Our previous studies demonstrated that in an HIV vaccine phase I trial, the DP6-001 trial, a polyvalent Env DNA prime-protein boost formulation could elicit potent and broadly reactive, gp120-specific antibodies with positive neutralization activities. Here we report on the production and analysis of HIV-1 Env-specific human monoclonal antibodies (hMAbs) isolated from vaccinees in the DP6-001 trial. For this initial report, 13 hMAbs from four vaccinees in the DP6-001 trial showed broad binding to gp120 proteins of diverse subtypes both autologous and heterologous to vaccine immunogens. Equally cross-reactive Fc receptor-mediated functional activities, including ADCC and antibody-dependent cellular phagocytosis (ADCP) activities, were present with both immune sera and isolated MAbs, confirming the induction of nonneutralizing functional hMAbs by the DNA prime-protein boost vaccination. Elicitation of broadly reactive hMAbs by vaccination in healthy human volunteers confirms the value of the polyvalent formulation in this HIV vaccine design.
IMPORTANCE The roles of Fc receptor-mediated protective antibody responses are gaining more attention due to their potential contribution to the low-level protection against HIV-1 infection that they provided in the RV144 trial. At the same time, information about hMabs from other human HIV vaccine studies is very limited. In the current study, both immune sera and monoclonal antibodies from vaccinated humans showed not only high-level ADCC and ADCP activities but also cross-subtype ADCC and ADCP activities when a polyvalent DNA prime-protein boost vaccine formulation was used.
INTRODUCTION
One important finding from the RV144 ALVAC-HIV (vCP1521) prime-AIDSVAX B/E boost vaccine efficacy trial is the identification of immune correlates of vaccine-induced protection from acquisition of human immunodeficiency virus type 1 (HIV-1). The presence of antibodies targeting the Env gp120 V1/V2 region was inversely correlated with infection risk, while the presence of IgA Env-binding antibodies directly correlated with infection risk (1). Further studies indicated that the presence of low plasma IgA Env antibody levels in association with high levels of antibody-dependent cellular cytotoxicity (ADCC) was inversely correlated with infection (1).
While broadly neutralizing antibodies (bnAbs) have been the main focus in HIV vaccine development efforts for several decades, ADCC activities have also been suggested in the literature to play important roles in the control of established HIV-1 infection as well as protection from initial infection (2). The majority of such studies have shown an inverse correlation between ADCC and the progression of disease or viral load (2). There was an inverse correlation between titers of ADCC activity in milk and transmission to newborns which was not correlated with neutralization (3).
Along with ADCC, an inverse correlation exists between antibody-dependent cell-mediated viral inhibition (ADCVI) and progression to simian AIDS (4). Furthermore, phagocytosis is increased in acute infection and is impaired in chronic infection for monocytes and dendritic cells. Blockage of Fc gamma receptor II (FcγRII) results in the loss of antibody-dependent cellular phagocytosis (ADCP) activity (5). It was shown in a mucosal protection study in a nonhuman primate (NHP) model that nonneutralizing antibodies can reduce the plasma viral load, and phagocytosis was implicated in passive protection (6).
When investigating how well an adenovirus prime-gp140 boost vaccination regimen compared to an unboosted vaccination in macaques, it was discovered that improved ADCC and ADCVI levels correlated with reduced acute viremia (7). Animals that had greater prechallenge ADCC, ADCVI, and transcytosis inhibitory activities had greater antibody avidity and reduced acute viremia (7). In humans, differences in vaccination efficacy may have been due to the Fc-mediated functions elicited by each vaccine. The RV144 trial had an immunogen similar to that in the VAX003 trial, yet the outcomes were different. The RV144 vaccine was able to elicit highly functional IgG3 antibodies, while the repeated immunization of a subunit vaccine pushed the VAX003 vaccine repertoire only into the less functional IgG4 subclass (8). Mounting evidence indicates that more complex interrogations of serology are needed to understand vaccine trial outcomes and that these interrogations include Fc-mediated activities (9).
In recent years, molecularly cloned human monoclonal antibodies (hMAbs) from HIV-1-infected patients have been used to provide a more in-depth understanding of monoclonal antibody (MAb)-mediated ADCC activity. However, there are only a few human MAbs that have been demonstrated to have potent ADCC activities. MAb A32 is a nonneutralizing antibody isolated from a chronically infected individual that has been shown to bind to Env on the surface of CD4 cells and mediate potent ADCC activity. Most importantly, an A32 Fab fragment can block most of the ADCC activity in a broad range of HIV-infected plasma samples (10). Two A32-like MAbs with distinct ADCC potencies were shown to have overlapping epitopes but differed in their binding angle and ability to mediate ADCC (11). Nonneutralizing epitopes are usually buried on the trimer surface and accessible only after engagement by CD4, which is the case with the Fc-mediated function of MAb A32 (10).
There is only limited study with ADCC-mediating MAbs produced from human volunteers who received experimental HIV vaccines, and these MAbs were almost exclusively from the RV144 trial. In the present study, we assessed the Fc-mediated effector functions of a panel of HIV-1 Env-specific MAbs isolated from four volunteers who received a DNA prime-protein boost human vaccine in a previously reported phase I clinical trial, the DP6-001 trial (12). These MAbs could recognize a wide range of primary HIV-1 Env glycoproteins from multiple major subtypes. While these MAbs did not exhibit significant neutralizing activity, they demonstrated potent Fc-mediated effector functions, including ADCC and ADCP activities. Our results provide valuable information on the magnitude and quality of ADCC activities mediated by MAbs produced from the second human HIV-1 vaccine trial, in addition to the RV144 trial. The information learned here can be used as a baseline to guide the design of future HIV-1 vaccines to elicit high-level, broadly reactive Fc-mediated effector functions.
MATERIALS AND METHODS
Plasma and PBMC samples from DP6-001 vaccine recipients.
All participants in the completed multigene polyvalent HIV vaccine DP6-001 phase I clinical trial gave written informed consent, as previously described (12). Samples were collected and tested according to protocols approved by the institutional review board (IRB) at the University of Massachusetts Medical School.
Plasma and peripheral blood mononuclear cell (PBMC) samples involved in the current study were from subjects ABL-034 (group A), ABL-001 and ABL-003 (group B), and ABL-040 (group C). The selection of samples was random in this pilot study, but samples were collected from at least one volunteer from each of the three study groups in which the dosing and route of DNA prime immunization were tested. The plasma samples used for ADCP assays were collected at week 0 (prebleed) and week 30 (post-protein boosts). The PBMCs used for Env-specific MAb isolation were collected from subjects ABL-001, ABL-003, and ABL-034 after two protein boosts and from subject ABL-040 after one protein boost due to the early termination of group C after one protein boost (13).
Isolation of Env-specific single B cells.
The cryopreserved PBMCs from the above-described vaccinees were thawed and processed in two ways for gp120-specific single B cell sorting. In the first method, freshly thawed PMBCs were cultured in RPMI 1640 with 15% fetal bovine serum (FBS), 1% l-glutamine, and 1% penicillin-streptomycin (P/S) overnight at 37°C with 5% CO2. Then, B cells were isolated by magnetic bead separation (magnetically activated cell sorting human B cell isolation kit II; Miltenyi Biotec, San Diego, CA). The gp120 protein probe for antigen-specific B cell staining was a mixture of 5 gp120 proteins from HIV-1 strains A2 (clade A strain 92UG037.8), Bal and B (clade B strains Bal and 92US715.6, respectively), Czm (clade C strain 96ZM651), and AE (clade AE strain 93TH976.17), which were autologous to gp120 DNA and proteins in the DP6-001 vaccine formulation. The B cells were stained with phycoerythrin (PE)-conjugated gp120 protein, anti-human-CD19 allophycocyanin (APC)-Cy7, and Via-Probe solution (a dead cell marker) (BD Biosciences, San Jose, CA). gp120-positive, CD19+, and 7-aminoactinomycin D-negative cells were sorted with a FACSAria flow cytometer (BD) into 96-well PCR plates containing a lysis buffer (RNaseOut recombinant ribonuclease inhibitor, Invitrogen 5× first-strand buffer, dithiothreitol, Igepal detergent). The plates were immediately spun down and frozen on dry ice.
The second method involved bulk sorted B cells isolated from freshly thawed volunteer PBMCs. After B cell isolation, the cells were incubated with PE-conjugated gp120 and cell markers as described above. The cells were then sorted into a single tube by use of the FACSAria flow cytometer under the same conditions described above. Sorted cells were then seeded at clonal dilutions in 96-well round-bottom culture plates with 5 × 104 irradiated EL4C5 feeder cells, medium B (RMPI 1640, 15% FBS, 2% HEPES, 0.1% 2-mercaptoethanol, 1% P/S), and 3% activated rabbit splenocyte supernatant. After a week of culture, the supernatants were tested for gp120 binding by enzyme-linked immunosorbent assay (ELISA), and the cells in the gp120-positive wells were harvested for cloning of Ig variable regions.
Isolation of V(D)J immunoglobulin regions.
The single B cells sorted into PCR plates were directly amplified by reverse transcription (RT) and nested PCR of the variable Ig region. In contrast, for the wells from the clonal culture positive for gp120 binding, RNA was extracted for RT-PCR. The Ig heavy chain-variable (VH), and lambda light chain-variable (VL), or kappa light chain-variable (VK) regions were isolated by RT-PCR using the RT and nested PCR primers described in a previous report (14).
RT was performed using SuperScript III reverse transcriptase (Invitrogen) according to the manufacturer's instructions, with primer concentrations of 10 μM being used for each heavy chain primer and 25 μM being used for each light chain primer. The nested primers were situated on the edge of the VH, VL, or VK region of the immunoglobulin genes with primers that contain a sequence that overlaps an Ig overlap cassette system (14). For the VH nested PCR, external (Ext) and internal (Int) forward primers covered VH1 to VH6 and reverse primers covered IgA, IgD, IgG, and IgM. For the VK nested PCR, external and internal forward primers covered VK1 to VK7 and reverse primers covered the kappa chain constant (IgK). For the VL nested PCR, external and internal forward primers covered VL1 to VL10 and reverse primers covered the lambda chain constant (IgL). The internal primers for VH, VK, or VL had a 5′ tag overlapping the linear cassette fragments expressing the leader sequence (C fragment), the IgG1 constant region (H fragment), the IgK constant region (K fragment), or the IgL constant region (L fragment) for overlapping PCR to construct the linear expression cassettes (14).
The nested PCR utilized Qiagen's HotStarTaq Plus kit. The first PCR used 5 μl of the RT product and followed the manufacturer's instructions with the addition of MgCl2. The second PCR used 2.5 μl of the product of the first PCR and replaced MgCl2 with Qiagen Q buffer in the reaction mixture. The PCR products were then cleaned up with a Qiagen PCR cleanup kit for sequencing analysis.
The variable region sequence from heavy and light chain cloning was analyzed by the IMGT/V-QUEST software program (15). Parameters that were collected included the functionality of the sequence; the heavy, kappa, or lambda chain designation; the allele designation for V, D, and J; CDR (complementarity-determining region) and FR (fragment region) lengths; percent identity to the germ line; and the junction amino acid sequence.
Construction of linear Ig expression cassettes.
After identification of 400- to 500-bp bands from RT-PCR, sequencing was done with primer CL-F681 (TCTGGGTTCCAGGTTCCACTGGTGAC) to confirm that the Ig sequence was functional. After sequence verification, variable region fragments were cloned into an immunoglobulin-expressing linear cassette by overlapping PCR (14). The 5′ fragment of the cassette contained a cytomegalovirus promoter and an Ig leader sequence, while the IgH, IgK, or IgL 3′ fragment contained the constant region of the Ig heavy chain (IgG1), kappa chain, or lambda chain, respectively, and a polyadenylation signal. The overlap PCR was done with iProof DNA polymerase (Bio-Rad, CA) according to the manufacturer's instructions. Equal amounts of the VH, VK, or VL PCR product, 5′ promoter region fragment, and the 3′ constant region CH, CK, or CL fragment were mixed with deoxynucleoside triphosphates, HF buffer, forward and reverse primers, and iProof DNA polymerase (Bio-Rad, CA). After the overlap PCR product of the heavy, kappa, or lambda chain was verified by agarose gel electrophoresis, the paired IgH/IgK or IgH/IgL fragments were used for transient transfection of 293T cells to produce antibodies in the transfection supernatant. The supernatant was used for screening for gp120 binding by ELISA, as described below. Clones of interest were further cloned into a mammalian expression vector, pJW4303, for large-scale production.
Expression and production of recombinant MAbs.
For large-scale production, equal copy numbers of heavy and light chain plasmids were transfected with polyethyleneimine into Freestyle 293F cells (Invitrogen). After 2 days of expression, the cell culture supernatant was harvested and human IgG was purified by Äkta fast-performance liquid chromatography with protein A HP columns (GE Healthcare).
HIV-1 gp120 protein production and conjugation.
Recombinant HIV-1 gp120 was produced from transient transfection of Freestyle 293F cells. Plasmids expressing the gp120 proteins from HIV-1 strains 92UG037.8 (A2), 92US715.6 (B), Ba-L (Bal), 96ZM651 (Czm), 93TH976.17 (AE), JR-FL, 93MW965 (C2), 92UG021 (D), and consensus AE sequences were used to transfect 293F cells by use of the 293Fectin transfection reagent (Invitrogen), and after 2 days of expression, the cell culture supernatant was harvested and the gp120 protein was purified over a lectin column.
Biotinylation was performed by transferring biotin to the lysine residue in the Avitag sequence (LNDIFEAQKIEWHE) at the C terminus of gp120. A commercially available biotin ligase system (Avidity, CO) was used to biotinylate the Avitag sequence of the DP6-001 autologous gp120s (from strains A2, B, Bal, Czm, and AE). The biotinylated gp120 proteins were then further conjugated to streptavidin-APC (Invitrogen) or ExtrAvidin reagent-PE (Sigma) as previously reported (14, 16).
ELISA.
Supernatant from the B cell clonal culture, supernatant from cultures of 293T cells transiently transfected with paired IgH/IgK or IgH/IgL linear cassettes, or purified MAb was screened for binding in a sandwich ELISA format in 384-well microtiter plates (Corning, NY). First, 20 μl/well of goat anti-human IgG Fc (catalog no. 2048-01; Southern Biotech) in phosphate-buffered saline (PBS; 1 μg/ml) was incubated overnight at 4°C. Then, the plate was blocked for 1 h at 37°C in 40 μl/well of blocking buffer (PBS containing 4% whey and 5% powdered milk). The plates were further incubated with 20-μl/well serial dilutions of antibody in dilution buffer (PBS containing 4% whey) for 1 h at room temperature. Biotinylated gp120 or peptide (20 μl/well) at 1 μg/ml in dilution buffer was incubated for 1 h at room temperature. Streptavidin-conjugated horseradish peroxidase (Vector Laboratories, CA) was diluted in dilution buffer to 500 ng/ml, added to the plate, and incubated for 1 h at room temperature. The plates were washed five times with wash buffer (PBS containing 0.1% Triton-X) between steps. In the last step, the plates were developed for 5 min at 37°C in 20 μl of a 3,3′5,5′-tetramethylbenzidine substrate solution (Sigma). The reaction was stopped with 20 μl of 2 N H2SO4. The plates were read at 450 nm.
For testing the breadth of binding in the ELISA, a concanavalin A (ConA) ELISA format was utilized. In 384-well microtiter plates (Corning, NY), 20 μl of ConA (5 μg/ml in PBS) was incubated at room temperature for 1 h. Then, 20 μl gp120 (1 μg/ml) was added and the plates were incubated overnight at 4°C. After blocking for 1 h at 37°C in blocking buffer, serial dilutions of MAbs diluted in dilution buffer were added to the wells and the plates were incubated for 1 h at room temperature. Biotinylated goat anti-human IgG (H/L; Vector Laboratories) was diluted in dilution buffer to 1 μg/ml and added to the wells, and the plates were incubated for 1 h at room temperature. Streptavidin-conjugated horseradish peroxidase (Vector Laboratories) was diluted in dilution buffer to 500 ng/ml and added to the plate, and the plate was incubated for 1 h at room temperature. The plates were washed five times with wash buffer (PBS containing 0.1% Triton-X) between steps. Finally, the plates were developed for 3 min at room temperature in 20 μl of a 3,3′5,5′-tetramethylbenzidine substrate solution (Sigma). The reaction was stopped with 20 μl of 2 N H2SO4. The plates were read at 450 nm.
Measurement of binding affinity with an Octet Qke instrument.
The kinetics of human MAb binding to gp120 proteins were measured on an Octet Qke instrument (manufactured by ForteBio), based on biolayer interferometry. The antibody being tested was loaded onto protein A sensor tips diluted to 10 μg/ml in ForteBio kinetics buffer. After capture, the tips were washed in kinetics buffer and a baseline measurement was recorded. The tips were then incubated in wells containing serial dilutions of gp120 protein (300 nM to 0.4 nM) to measure the association rate (Kon) and dissociation rate (Koff). The antibody binding kinetics and equilibrium dissociation constant (KD; Koff/Kon) values were analyzed by the ForteBio Data Analysis software package (version 7.1) using a 1:1 binding model.
Neutralization assays.
Pseudovirus neutralization assays were performed using a system previously optimized in TZM-bl cells (17). Pseudovirus was generated by cotransfection of a gp160 envelope with the pSG3ΔEnv backbone. Pseudotyped virus was added at 200 tissue culture infective doses per well and incubated with antibody at 37°C for 1 h. TZM-bl cells were then seeded at 10,000 cells/well in DEAE-dextran at a final concentration of 20 μg/ml. The plates were incubated at 37°C for 48 h and then developed with a luciferase assay reagent per the manufacturer's instructions (Promega). The numbers of relative light units (RLUs) were collected by running of the plates on a Wallac luminometer. Neutralization was calculated as the percent reduction in luciferase activity in the presence of antibody compared to the luciferase activity induced by the virus control without antibody, calculated as follows: {[1 − (average number of sample RLUs − number of cell control RLUs)/(number of virus control RLUs − number of cell control RLUs)]} × 100.
ADCC assays. (i) ADCC against gp120-coated target cells.
The GranToxiLux (GTL) assay was used to detect the ADCC activities of human MAbs and plasma directed against CEM.NKRCCR5 cells (NIH AIDS Reagent Program, Division of AIDS) coated with recombinant gp120 proteins (from strain A2, B, Bal, Czm, or AE) which were included as part of the vaccine immunogens as described previously (18). Cryopreserved human PBMCs from an HIV-seronegative donor with the homozygous 158V/V genotype for Fc gamma receptor IIIa were used as the source of effector cells (19) at an effector-to-target cell ratio of 30:1. Data are reported as the proportion of cells positive for proteolytically active granzyme B (GzB) out of the total viable target cell population (percent GzB activity) after subtraction of the background activity observed in wells containing effector and target cells in the absence of antibodies or plasma. ADCC endpoint titers were determined by interpolation of the dilutions of sera that intercepted the positive cutoff (8% GzB activity) using GraphPad Prism (version 6.0) software (GraphPad Software, Inc.).
(ii) ADCC against HIV-1-infected target cells. The ability of DP6-001 hMAbs and plasma to direct the killing of CEM.NKRCCR5 cells infected with subtype AE HIV-1 isolate CM235 (GenBank accession no. AF259954.1; Agnès Chenine, U.S. Military HIV Research Program) and subtype B isolate Bal (GenBank accession no. AY426110) infectious molecular clone (IMC) viruses containing a Renilla luciferase (Luc) reporter gene (20) was measured using a Luc-based ADCC assay according to a previously described procedure (21, 22). Cryopreserved PBMCs were used as a source of effector cells (effector-to-target cell ratio, 30:1). Killing was measured as a reduction in luminescence (ViviRen assay; Promega) compared to that of control wells containing target and effector cells in the absence of antibodies. ADCC endpoint titers were determined by interpolating the concentrations of serum that intercepted the positive cutoff (15% specific killing).
ADCP assays.
ADCP was measured by a modified form of a previously described method (23). Biotinylated gp120 (5 μg) was incubated with 1 μl streptavidin-fluorescent beads (FluoSpheres NeutrAvidin-labeled microspheres, 1.0 μm, yellow-green fluorescent [505/515 nm], 1% solids; Invitrogen) that were diluted 100-fold for 25 min at room temperature in the dark. THP-1 cells were plated in a 96-well U-bottom plate (250,000 cells per well), and serial dilutions of the MAbs were added. The bead-gp120 mix was then further diluted 5-fold in PBS-bovine serum albumin, 50 μl of this mixture was added to cells, the cells were incubated for 3 h at 37°C and washed at low speed. The cells were then assayed for fluorescent bead uptake by flow cytometry using a BD Biosciences LSR II flow cytometer. The phagocytic score was obtained by the formula (percent phagocytosis/mean fluorescence intensity) and normalized by division by 106.
RESULTS
Volunteers receiving the DP6-001 vaccine and their serum antibody responses.
In the current study, a total of 13 human Env-specific MAbs from four DP6-001 volunteers (ABL-001, ABL-003, ABL-034, and ABL-040) were produced by the single B cell cloning approach (Fig. 1). In the DP6-001 trial, volunteers in groups A and B first received three low-dose DNA prime immunizations by the intradermal (i.d.) and intramuscular (i.m.) routes (1.2 mg each time), respectively, with a polyvalent formulation including five gp120-expressing plasmids and one Gag-expressing DNA plasmid, followed by two immunizations with a polyvalent preparation of five gp120 proteins produced from CHO cells formulated with the adjuvant QS-21 (12). Group C volunteers received the same formulation, except that they received (i) a higher-dose DNA prime immunization by the i.m. route (7.2 mg each time) and (ii) only a single gp120 protein boost immunization due to the early stop of the clinical trial in this group of volunteers (13). In the current pilot study, MAbs were generated from one group A volunteer, two group B volunteers, and one group C volunteer (Fig. 1; Table 1).
FIG 1.

Serum antibody responses from subjects in the DP6-001 phase 1 vaccine trial. (A) gp120-specific IgG titers for select volunteers. The gp120 protein used to coat the ELISA plates for detection of the IgG titer was a proportionally mixed solution of the 5 autologous gp120s used in the DP6-001 immunization. Solid arrows, immunization with DNA prime; open arrows, immunization with gp120 protein boosts. In group A (GrpA), the DNA prime was done i.d., while in group B (GrpB) and group C (GrpC), the DNA prime was done i.m. The volunteer in group C received a high-dose DNA prime and received only the first boost. (B) Serum neutralization titers (ID50) determined by the PhenoSense assay at Monogram Biosciences. NA, not available; -, no detectable titer.
TABLE 1.
Characteristics of hMAbs from volunteers in the DP6-001 trial
| Volunteer | hMAb | Wk | Epitope | Heavy chain |
Light chain |
||||
|---|---|---|---|---|---|---|---|---|---|
| Germ line | Mutation % | CDR3 | Germ line (chaina) | Mutation % | CDR3 | ||||
| ABL-034 (group A) | HH1G9 | 36 | C2/C5 | 1-2*02 | 3 | 16 | 2-28*01 (K) | 1 | 9 |
| HH2D11 | 36 | NDb | 1-2*02 | 3 | 19 | 3-11*01 (K) | 0 | 8 | |
| HH4E4 | 36 | ND | 1-46*01 | 5 | 18 | 7-43*01 (L) | 2 | 11 | |
| ABL-001 (group B) | GB1 | 30 | C2 | 1-69*9 | 6 | 18 | 1-5*03 (K) | 4 | 6 |
| EA1 | 32 | C5 | 1-69*01 | 2 | 19 | 3-10*01 (K) | <1 | 11 | |
| EA7 | 32 | ND | 1-69*01 | 5 | 14 | 3-19*01 (L) | 4 | 11 | |
| EA8 | 32 | ND | 4-39*01 | 9 | 20 | 1-40*01 (L) | 3 | 11 | |
| ABL-003 (group B) | CC3C2 | 36 | ND | 3-30*03 | 2 | 23 | 1-44*01 (L) | 2 | 11 |
| CC6B5 | 36 | C2 | 1-46*01 | 3 | 12 | 1-26*01 (K) | 4 | 9 | |
| CC6C11 | 36 | ND | 1-46*01 | 5 | 12 | 1D-17*02 (K) | 1 | 9 | |
| ABL-040 (group C) | TA6 | 30 | V3 | 1-3*01 | 5 | 18 | 3-10*01 (L) | 3 | 11 |
| TA7 | 30 | V3 | 1-3*01 | 4 | 17 | 3-10*01 (L) | 1 | 11 | |
| TB7 | 30 | C2/C5 | 1-69*06 | 2 | 23 | 2-14*01 (L) | 2 | 10 | |
K, kappa chain; L, lambda chain.
ND, not determined.
Initial DNA priming immunization did not elicit a detectable gp120-specific IgG titer in these volunteers' sera, with the exception of that from the volunteer in group C who received the higher-dose DNA prime. However, after the first protein boost, gp120-specific IgG responses were observed to rise quickly to high titers (Fig. 1A). Serum neutralization was tested against a diverse panel of pseudotyped viruses from multiple clades (clades A, B, C, D, and AE). For the volunteers included in the current study, positive neutralization activity against this panel of pseudotyped viruses could be seen in immune sera after one or two protein boost immunizations, with the titers for the 50% inhibitory dose (ID50) ranging from 1:20 to 1:977 (Fig. 1B).
HIV-1 Env-specific hMAbs from DP6-001 trial volunteers.
Frozen PBMCs from the four DP6-001 vaccine trial volunteers were used for antigen-specific cloning of human monoclonal antibodies (hMAbs). The 13 hMAbs in this panel were from single B cells isolated from frozen PBMCs collected at 30 to 36 weeks after the initial DNA immunization (i.e., after one or two protein immunizations). Immunogenetics analysis showed that except for two Ig HC variable regions (IGHV) (hMAb CC3C2 and EA8), which were subtypes 3 and 4, respectively, the IGHVs of these MAbs were from germ line subtype 1, while the Ig LC variable regions (IGLVs) were more diverse, with a variety of variable gene subgroups being utilized. The mutation rates of IGHVs from germ lines were low, usually 3 to 6%, but the mutation rate for one was 9%. The CDR3 length ranged from 12 to 23 amino acids (Table 1).
Because the volunteers from whom the gp120-specific hMAbs were isolated were immunized with a polyvalent gp120 vaccine, a wide selection of gp120 antigens was used to test the breadth of binding of this panel of MAbs. Two patterns of binding were evident, with one group of hMAbs broadly binding to multiple clades of gp120 proteins and the second group specifically binding to gp120 antigens from B clade. Nine hMAbs (hMAbs HH1G9, HH2D11, and HH4E4 from volunteer ABL-034 of group A; MAbs GB1, EA7, and EA8 from volunteer ABL-001 of group B; MAbs CC6B5 and CC6C11 from volunteer ABL-003 of group B; and MAb TB7 from volunteer ABL-040 of group C) belonged to the first pattern, although some of them had higher relative binding than the other hMAbs. Four hMAbs (hMAb EA1 from volunteer ABL-001 of group B, hMAb CC3C2 from volunteer ABL-003 of group B, and hMAbs TA6 and TA7 from volunteer ABL-040 of group C) belonged to the second pattern. This pattern was consistent between the gp120 antigens autologous to vaccine components used in the ELISA (Fig. 2) and the gp120 antigens heterologous to gp120 vaccine components used in the ELISA (Fig. 3). Those hMAbs with broad reactivity against autologous gp120 antigens were also able to recognize a wide range of heterologous gp120 antigens. One noticeable exception was the gp120 antigen from a subtype D virus. Given the fact that the gene sequences of subtype D viruses are more different from the consensus sequences than the gene sequences of viruses of other major subtypes (such as subtypes A, B, C, and AE), it is not surprising that the broadly reactive hMAbs cannot recognize the subtype D gp120. In addition, no gp120 immunogen from subtype D was included in the vaccine formulation. However, one broadly reactive hMAb, hMAb HH1G9 (from group A), was able to bind all gp120 antigens tested, including the one from subtype D.
FIG 2.

Reactivities of 13 hMAbs against a panel of five autologous gp120 proteins included in the DP6-001 trial measured by ELISA. HIV-1 Env gp120 proteins were derived from the following isolates: 92UG037.8 (A2), 92US715.6 (B), Ba-L (Bal), 96ZM651 (Czm), and 93TH976.17 (AE). (A) hMAbs from one group A volunteer; (B) hMAbs from two group B volunteers; (C) hMAbs from one group C volunteer; (D) well-characterized hMAbs as controls. OD450 nm, optical density at 450 nm.
FIG 3.

Reactivities of 13 hMAbs against a panel of heterologous gp120 proteins revealed by ELISA. HIV-1 Env gp120 proteins were derived from the following isolates: JR-FL, 93MW965 (C2), 92UG021 (D), and consensus clade AE (AE Con). (A) hMAbs from one group A volunteer; (B) hMAbs from two group B volunteers; (C) hMAbs from one group C volunteer; (D) well-characterized hMAbs as controls.
Several human bnAbs well characterized by researchers in the HIV field were included as controls in the binding antibody analysis. MAb VRC01 is able to bind to gp120 proteins from different subtypes but binds less well to the gp120 protein from subtype D. MAb PGT128 can bind strongly only to gp120 from subtype A and binds less well to gp120 proteins from other subtypes, reflecting its overall reduced binding to gp120 monomers. Not surprisingly, V3-specific hMAb 447-52D mainly binds to subtype B gp120 proteins, confirming its preferred subtype specificity. In contrast, MAb A32 is able to bind to all gp120 proteins except the one from subtype D, which may also explain its high ADCC activity against a wide range of HIV-1 isolates of different subtypes.
The kinetics of binding, including affinities for the panel of hMAbs from volunteers in the DP6-001 trial, to the same autologous gp120 proteins used in the ELISA were also measured (Table 2; Fig. 2). The levels of binding affinity are comparable to the binding profile seen in the ELISA. A few hMAbs that were subtype B specific (hMAbs EA1 and CC3C2) could also be seen (Table 2).
TABLE 2.
Kinetics of binding of DP6-001 hMAbsa
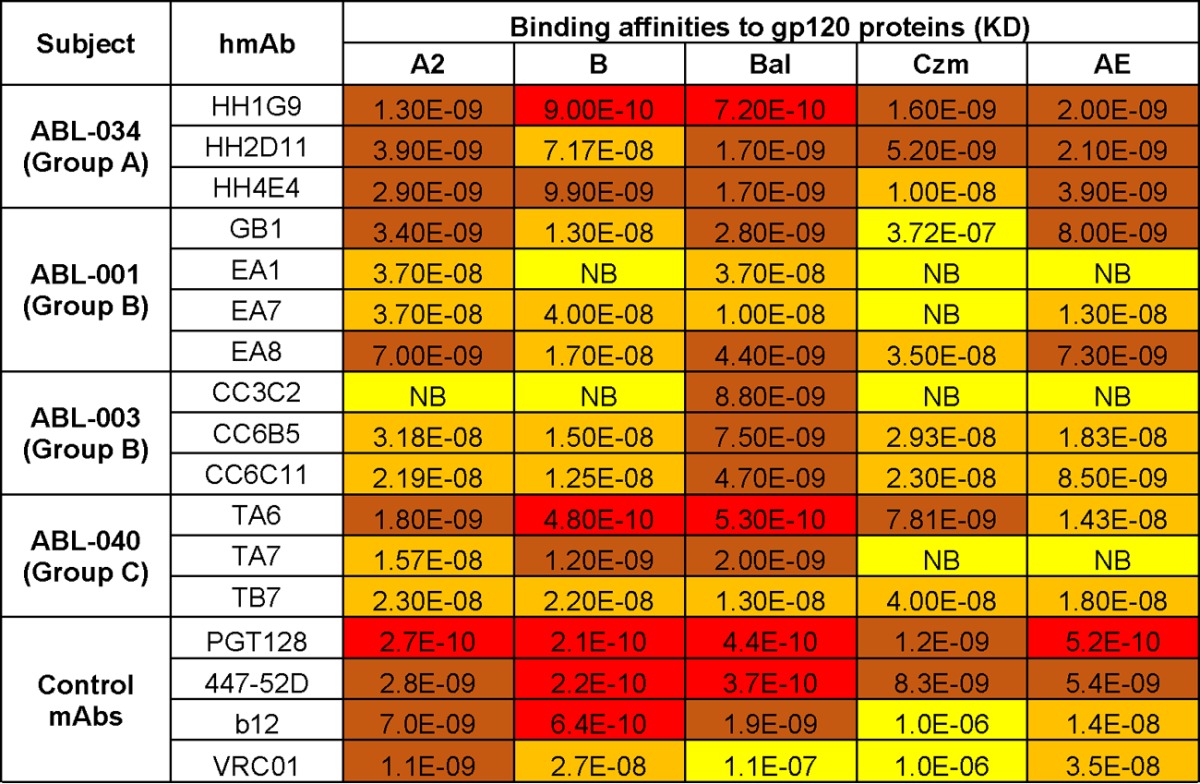
NB, no binding; KD, equilibrium dissociation constant (M); red, equilibrium dissociation constants of <1.0E−10; brown, equilibrium dissociation constants of 1.0E−9 to 1.0E−10; orange, equilibrium dissociation constants of 1.0E−8 to 1.0E−9; yellow, equilibrium dissociation constants of >1.0E−8.
Using a peptide microarray system (JPT Peptide Technologies, Berlin, Germany), linear epitopes were determined for about half of the hMAbs from volunteers in the DP6-001 trial included in this study (Table 1). The other half did not give clear dominant binding peaks, indicating that their epitopes may be conformational. The linear epitopes were mainly in the constant regions, including C2 (hMAbs GB1 and CC6C11), C5 (hMAb EA1), and C2/C5 (hMAbs HH1G9 and TB7) (Table 1). Representative JPT Peptide Technologies mapping results are shown in Fig. 4A. Two hMAbs from the DP6-001 trial (TA6 and TA7) had specific binding to peptides in the V3 region (Table 1). For the V3-specific hMAbs, a peptide ELISA using cyclic V3 was further used to confirm TA6 and TA7 binding, along with positive-control V3-specific MAb 447-52D and negative-control non-V3-specific hMAb EA1 and TB7 binding (Fig. 4B).
FIG 4.

Multiple epitopes elicited by vaccination in human volunteers revealed by use of a peptide microarray. Purified hMAbs were tested on JPT Peptide Technologies slides containing overlapping peptides for a diverse set of gp120s from multiple clades (identified on the right), including consensus sequences. (A) Peptide microarray against multiple gp120 protein peptides, focused on epitopes in clades C2, C5, and V3. The numbers on the x axes of panel A are amino acid positions. (B) ELISA against a consensus clade B V3 circular peptide.
The majority of cloned DP6-001 hMAbs did not exhibit a neutralization titer, with the exception of the V3-specific hMAb TA6, which could weakly neutralize tier 1 subtype B pseudoviruses based on the neutralization assay conducted in-house at the University of Massachusetts Medical School or at M. Seaman's laboratory (Table 3).
TABLE 3.
Neutralization titers of hMAb TA6 by TZM-bl cell assaya
| Study location, neutralization result, and virus | Clade | IC50 (μg/ml) |
|---|---|---|
| UMass | ||
| Positive neutralization | ||
| MN | B | 27.5 |
| SF162 | B | 3.67 |
| SS1196 | B | 2.23 |
| QH0692.42 | B | 31.8 |
| Negative neutralization | ||
| NL4-3 | B | >50 |
| SC422661 | B | >50 |
| WITO4160 | B | >100 |
| ZM109F.PB4 | C | >50 |
| Du156 | C | >50 |
| Du422.1 | C | >100 |
| ZM197.pB7 | C | >50 |
| CM235.c11 | CRF01_AE | >50 |
| 816367.CO2 | CRF01_AE | >50 |
| MLV | >50 | |
| Harvard | ||
| Positive neutralization | ||
| SS1196.1 | B | 30.08 |
| T271-11 | CRF02_AG | 59.08 |
| Negative neutralization | ||
| MS208.A1 | A | >100 |
| Q23.17 | A | >100 |
| MN-3 | B | >100 |
| SF162.LS | B | >100 |
| BaL.26 | B | >100 |
| 6535.3 | B | >100 |
| MW965.26 | C | >100 |
| TV1.21 | C | >100 |
| ZM109F.PB4 | C | >100 |
| DJ263.8 | CRF02_AG | >100 |
| 242-14 | CRF02_AG | >100 |
| TH023.6 | CRF01_AE | >100 |
IC50, 50% inhibitory concentration; UMass, University of Massachusetts Medical School; Harvard, Harvard Medical School; MLV, murine leukemia virus.
ADCC activities with sera and hMAbs from volunteers in the DP6-001 trial.
We next studied the presence and levels of Fc-mediated antibody activities in either immune sera or hMAbs from volunteers in the DP6-001 trial (Fig. 5 and 6). After multiple immunizations, including a protein boost, gp120-specific ADCC activities could be seen in all volunteers and were sustained out to 36 weeks, which was the last time point that we included in the ADCC study. ADCC activity against both subtype B and subtype AE gp120 antigens was seen (Fig. 5A). Most significantly, ADCC activities could be detected by using either the assay format with target cells coated with recombinant gp120 antigens or the infection format with viruses expressing HIV-1 Env antigens (Fig. 5A and B). ADCC levels in cells with viruses expressing HIV-1 Bal Env were lower than those in cells with viruses expressing HIV-1 CM235 (Fig. 5B). The ADCC activities in the serum from volunteer ABL-040 (group C of the DP6-001 trial) were lower than those in the sera from the other three volunteers, most likely due to the fact that group C volunteers did not receive the second protein boost (Fig. 5B).
FIG 5.

Titers of ADCC activity in serum against gp120-coated cells for select volunteers from a GranToxiLux ADCC assay (A) and infected cells from a Luc-based ADCC assay (B).
FIG 6.

ADCC reactivities of 13 hMAbs with a panel of DP6-001 gp120 proteins. HIV-1 Env gp120 proteins were derived from the following isolates: 92UG037.8 (A2), 92US715.6 (B), Ba-L (Bal), 96ZM651 (Czm), and 93TH976.17 (AE). (A) hMAbs from volunteer ABL-034 (group A); (B) hMAbs from volunteer ABL-001 (group B); (C) hMAbs from volunteer ABL-003 (group B); (D) hMAbs from volunteer ABL-040 (group C); (E) hMAb controls.
Most of the hMAbs generated by these volunteers in the DP6-001 trial also showed high levels of ADCC activity (Fig. 6A to D). The hMAbs that showed broad gp120 binding activity also showed broad ADCC activities. Those with narrow binding specificity (mainly B subtype; hMAbs EA1, CC3C2, TA6, and TA7) also had poor ADCC activity. TB7 was the one hMAb that had a broad gp120 binding profile but lacked ADCC activity, indicating that all hMAbs produced from this group C volunteer had poor ADCC activity (Fig. 6D). When the relative ADCC activities were compared with the activity of the positive-control hMAb A32, some of the hMAbs from the volunteers in the DP6-001 trial (MAbs GB1, EA7, and CC685) had ADCC activities against selected subtypes of gp120 antigens close to the ADCC activity achieved by A32 (Fig. 6E and 7A). The hMAbs from the DP6-001 trial were also tested against cells infected with two infectious molecular clone (IMC) viruses, Bal and CM235. The levels of specific killing were lower than those seen with gp120-coated cells, though hMAb GB1 was still able to mediate decent ADCC activity against both IMC viruses (Fig. 7B).
FIG 7.

ADCC activity of human MAbs. (A) The maximum percent GzB activity of 13 hMAbs with a panel of DP6-001 gp120 proteins. HIV-1 Env gp120 proteins were derived from the following isolates: 92UG037.8 (A2), 92US715.6 (B), Ba-L (Bal), 96ZM651 (Czm), and 93TH976.17 (AE). (B) Maximum percent specific killing of 11 hMAbs with cells infected with IMC viruses Bal and CM235.
Another important Fc-mediated function that was explored in the current study was antibody-dependent cellular phagocytosis (ADCP). As seen with the antibody binding and ADCC analyses, sera from the volunteers in the DP6-001 trial were able to mediate ADCP against all 5 autologous gp120 proteins (Fig. 8). Also consistent with the ADCC results was the observation that group C volunteer ABL-040 has reduced ADCP activity, most likely due to the missing second protein boost in the original human vaccine study (12).
FIG 8.

Plasma ADCP response of MAbs from four DP6-001 trial volunteers against gp120-coated fluorescent beads. (A) Representative histograms of ADCP assay. Results for Bal gp120-coated fluorescent beads assayed with THP-1 cells using prebleed (day 0) and day 210 sera from volunteer ABL-001 as examples are shown. Bars indicate percent positive cell population bracketed by the horizontal bar. (B) ADCP with sera from day 0 and day 210 bleeds from 4 volunteers against beads coated with the following gp120 isolates: 92UG037.8 (A2), 92US715.6 (B), Ba-L (Bal), 96ZM651 (Czm), and 93TH976.17 (AE).
The majority of hMAbs from the DP6-001 trial were capable of mediating ADCP against the entire range of vaccine autologous gp120 antigens. Like the results of the ADCC study, two hMAbs with narrow gp120 binding (hMAbs EA1 and CC3C2) also had lower levels of ADCP activity. Also similar to the ADCC results, while hMAb TB7 had a relatively broad binding antibody profile, its ADCP activity was also low. Interestingly, hMAbs TA6 and TA7 still showed strong ADCP activity against multiple subtypes of gp120 antigens, even though they had narrow binding breadths and low levels of ADCC activity (Fig. 9).
FIG 9.

ADCP reactivity of 13 hMAbs with a panel of DP6-001 gp120 proteins. HIV-1 Env gp120 proteins were derived from the following isolates: 92UG037.8 (A2), 92US715.6 (B), Ba-L (Bal), 96ZM651 (Czm), and 93TH976.17 (AE). (A) hMAbs from volunteer ABL-034 (group A); (B) hMAbs from volunteer ABL-001 (group B); (C) hMAbs from volunteer ABL003 (group B); (D) hMAbs from volunteer ABL-040 (group C); (E) normal human Ig (NHIgG) used as a negative control.
When the endpoint titer for ADCP or the maximum ADCP score was used, hMAbs from volunteers in the DP6-001 trial showed quite impressive ADCP activities (Fig. 10). They were even higher than the ADCP activity of hMAb A32, which is known for having extremely high ADCC activity against many subtypes of gp120 antigens. In contrast, hMAbs VRC01 and PGT128 showed higher ADCP activities than hMAb A32.
FIG 10.

The maximum (Max) ADCP reactivity of 13 hMAbs with a panel of DP6-001 gp120 proteins. HIV-1 Env gp120 proteins were derived from the following isolates: 92UG037.8 (A2), 92US715.6 (B), Ba-L (Bal), 96ZM651 (Czm), and 93TH976.17 (AE). Normal human Ig (NHIgG) was used as a negative control.
In summary, it can be seen that the antibody binding profiles and the ADCP activities of the hMAbs from volunteers in the DP6-001 trial are the factors that most closely correlate (Table 4). The broad gp120 binding reactivity was mirrored by the ADCP activity. The position where we see ADCC activity drop off is with the MAbs that have B clade-specific gp120 binding (hMAbs EA1, CC3C2, TA6, and TA7). Two B clade-specific MAbs, EA1 and CC3C2, had low to no ADCC activity against gp120 from any clade, including subtype B ADCC targets. The one exception to this was hMAb TB7, which had moderate but broad gp120 binding activity but no ADCC activity (Table 4).
TABLE 4.
Comparison of relative activity for gp120 ELISA binding and ADCC and ADCP activities among hMAbs induced by DP6-001
| Volunteer | MAb | Function | Activitya for gp120 proteins from the following strains: |
||||
|---|---|---|---|---|---|---|---|
| A2 | B | Bal | Czm | AE | |||
| ABL-034 (group A) | HH1G9 | Binding | ++++ | ++++ | ++++ | ++++ | ++++ |
| ADCC | ++ | +++ | +++ | +++ | ++ | ||
| ADCP | +++ | +++ | +++ | +++ | +++ | ||
| HH2D11 | Binding | ++++ | +++ | ++++ | +++ | ++++ | |
| ADCC | ++ | ++ | ++ | +++ | + | ||
| ADCP | ++ | ++ | ++ | +++ | +++ | ||
| HH4E4 | Binding | ++++ | ++++ | ++++ | ++++ | ++++ | |
| ADCC | ++ | +++ | ++ | +++ | ++ | ||
| ADCP | +++ | +++ | +++ | +++ | +++ | ||
| ABL-001 (group B) | GB1 | Binding | ++++ | ++++ | ++++ | +++ | ++++ |
| ADCC | +++ | ++++ | +++ | ++++ | +++ | ||
| ADCP | ++ | ++ | ++ | ++ | + | ||
| EA1 | Binding | − | + | ++ | − | − | |
| ADCC | + | − | − | − | − | ||
| ADCP | ++ | + | + | − | − | ||
| EA7 | Binding | +++ | ++++ | ++++ | − | +++ | |
| ADCC | ++ | ++++ | ++++ | + | ++ | ||
| ADCP | ++ | ++ | ++ | + | + | ||
| EA8 | Binding | +++ | ++++ | ++++ | +++ | ++++ | |
| ADCC | ++ | ++ | ++ | +++ | + | ||
| ADCP | ++ | ++ | ++ | +++ | ++ | ||
| ABL003 (group B) | CC3C2 | Binding | − | − | ++++ | − | − |
| ADCC | − | − | + | − | − | ||
| ADCP | − | + | ++ | − | − | ||
| CC6B5 | Binding | + | +++ | +++ | ++ | ++ | |
| ADCC | + | +++ | ++ | ++++ | + | ||
| ADCP | + | ++ | ++ | ++ | ++ | ||
| CC6C11 | Binding | ++ | +++ | +++ | ++ | +++ | |
| ADCC | + | +++ | +++ | ++++ | ++ | ||
| ADCP | ++ | ++ | ++ | +++ | ++ | ||
| ABL040 (group C) | TA6 | Binding | + | ++++ | ++++ | − | − |
| ADCC | − | ++ | + | − | − | ||
| ADCP | ++ | +++ | ++++ | ++++ | − | ||
| TA7 | Binding | + | +++ | ++ | − | − | |
| ADCC | − | ++ | + | − | − | ||
| ADCP | + | +++ | ++++ | +++ | − | ||
| TB7 | Binding | ++ | +++ | ++ | + | ++ | |
| ADCC | − | + | − | − | − | ||
| ADCP | + | ++ | ++ | ++ | ++ | ||
−, dissociation complex 50 (DC50) for binding of >5 nM, <5% maximum GzB activity for ADCC, and maximum ADCP score of <5; +, DC50 for binding of 0.5 to 5 nM, 5% to 10% maximum GzB activity for ADCC, and maximum ADCP score of 5 to 10; ++, DC50 for binding of 0.05 to 0.5 nM, 10% to 15% maximum GzB activity for ADCC, and maximum ADCP score of 10 to 15; +++, DC50 for binding of 0.005 to 0.05 nM, 15% to 20% maximum GzB activity for ADCC, and maximum ADCP score of 15 to 20; ++++, DC50 for binding of <0.005 nM, >20% maximum GzB activity for ADCC, and maximum ADCP score of >20.
DISCUSSION
Analysis of nonneutralizing antibody responses in immune sera from volunteers in the RV144 trial showed that ADCC was a correlate of reduced infection risk in the group with low IgA C1 binding titers (1). At the same time, knowledge in this area is limited due to the limited availability of human MAbs that are generated from non-HIV-infected individuals. More studies are needed to understand Fc effector functions in the responses of antibodies from volunteers who received different experimental HIV vaccines.
The current study produced another panel of human Env-specific MAbs from a vaccine trial in healthy volunteers. While these MAbs, with the exception of TA6, did not exhibit significant neutralization activity, they did display a broad breadth of gp120 binding, along with substantial Fc-mediated effector activity of similar breadths against various subtypes of HIV-1. Multiple linear epitopes along with some potential conformational epitopes contributed to the activities present in this newest panel of vaccine-elicited hMAbs. One such region could be classified as being adjacent to the epitope for cluster A antibodies to which nonneutralizing monoclonal antibodies such as C11 belong. The activity directed against the cluster A region was associated with a reduced risk of infection in the RV144 trial. This region has previously been shown to be a target of ADCC-mediated viral escape (24–26). hMAbs HH1G9, EA1, and TB7 all have an epitope that is adjacent to the cluster A epitope.
One of the central activities that was associated with protection in the RV144 trial was ADCC. The panel of MAbs isolated from volunteers vaccinated in the RV144 trial lacked neutralization activity but had a significant level of ADCC activity against autologous antigen (27). This pattern was also observed among MAbs characterized in the current study. Another similarity between MAbs from the RV144 and DP6-001 trials was that both panels preferentially used the VH1 germ line locus and recognized multiple clades, while both sets had a low level of somatic hypermutations (27). Some key differences between the two panels of MAbs were that the MAbs from the RV144 trial lacked linear epitopes and had consistent ADCC activity. For the subset of MAbs obtained in the current study that had the most consistent ADCC activity, gp120 binding was universally broad and potent, with potent ADCC being present in the absence of the enhancing modifications introduced in the Fc region as suggested by Shields et al. (28). These MAbs recognize a mixture of linear and possible conformational epitopes. These characteristics are possibly due to the vaccine formulation utilized in the DP6-001 trial, which included a 5-valent gp120 prime and boost, while in the RV144 trial, only two envelope proteins were included in the vaccine. Although there were other differences between the vaccine formulations, such as a viral vector prime compared to a DNA prime, the most likely factor involved in the epitope differences would be the antigens. The one linear epitope seen in the MAbs with ADCC activity from the RV144 trial was V3 (CH23), which was also present in our panel (hMAbs TA6 and TA7). Both sets of V3-specific MAbs were clade specific in their ADCC activities.
A smaller panel of RV144 MAbs (MAbs CH58, CH59, HG107, and HG120) was specifically isolated because of their V2 epitope, which was correlated with protection in vaccine recipients and their ability to mediate ADCC against RV144 vaccine strains (1, 22). Like the MAbs from the DP6-001 trial, these MAbs from the RV144 trial developed from the germ line with a low mutation rate. In contrast, MAbs from the RV144 trial utilized multiple immunoglobulin gene families, while the MAbs from the DP6-001 trial had a VH1 focus. Another difference between the two panels of MAbs was that the MAbs from the RV144 trial were specific to the isolates with the conserved critical binding residues by Liao and collaborators (22) and able to mediate neutralization against a couple of tier 1 clade AE viruses (22). Epitopes that can mediate ADCC are diverse, and as seen in the RV144 and DP6-001 trials, vaccines are able to elicit ADCC to multiple epitopes (1, 22, 27, 29). When antibodies against the two regions implicated in protection in the RV144 trial (C1 and V2) were tested together, synergy that increased the potency of CH58 to clinically relevant concentrations was observed (30). Because the epitope and activity of Fc-mediated functions like ADCC and ADCP are not as dependent on functional and well-formed trimers, elicitation of these types of MAbs could be an important component in a successful HIV vaccine.
Along with ADCC, another Fc-mediated function that was explored was ADCP activity. Recent studies have started to look into comparisons between vaccine trials and the differences in serology that may help with distinguishing which protocol will be best (9). With the group of MAbs that displayed the greatest breadth of gp120 binding and ADCC activity, ADCP activity was also broad and potent in the current study. Our study is one of the few to start looking into a more complete description of the Fc-mediated functions of MAbs elicited by vaccines in humans. Although the value of nonneutralizing activity is still being explored, some evidence indicates that there is immune pressure that HIV tries to circumvent during chronic infection. It has been shown that during chronic infection, FcγRII is downregulated on immune cells, and this downregulation is specifically associated with a loss of ADCP activity (5). While groups have compared ADCP activity in sera, our study is among the first to start to look more specifically at MAbs (31). Even without neutralization, the ability of MAbs to broadly bind gp120 was associated with the breadth of ADCP activity in the current study. This elicitation of multiclade MAbs may be an important factor to consider in future vaccine design. Recent studies in macaques have given evidence for this stance. Utilizing both simian immunodeficiency virus and HIV immunogens, polyfunctional antibody responses and antibody titers were correlated with increased sterilizing immunity (32). Both the protocol with macaques and the DP6-001 vaccine regimen utilized a protein boost. In the study with macaques, the protective efficacy was linked to the protein boost, which conferred both polyfunctional antibodies and protective efficacy. The groups that lacked a protein boost had reduced levels of both of these parameters (32). Therefore, it is exciting to demonstrate in the current report that broadly cross-reactive and polyfunctional MAbs can indeed be elicited in healthy human volunteers with the polyvalent DNA prime-protein boost HIV vaccine formulation DP6-001.
ACKNOWLEDGMENTS
We acknowledge the fluorescence-activated cell sorting core at the University of Massachusetts Medical School for help with the isolation of B cells from PBMCs. We thank Aaron Wallace, Diego Farfan, Shuying Liu, and Rachel Gerstein for helpful discussions during the experiments.
This study is partially supported by NIH/NIAID grants R21/R33 AI087191, HIVRAD P01 AI082274, R01 AI065250, IPCAVD U19 AI082676, and R01 AI112546 and PAVEG grant HHSN272201100016C.
REFERENCES
- 1.Haynes BF, Gilbert PB, McElrath JM, Zolla-Pazner S, Tomaras GD, Alam SM, Evans DT, Montefiori DC, Karnasuta C, Sutthent R, Liao H- X, DeVico AL, Lewis GK, Williams C, Pinter A, Fong Y, Janes H, DeCamp A, Huang Y, Rao M, Billings E, Karasavvas N, Bailer RT, Soderberg KA, Andrews C, Berman PW, Frahm N, De Rosa SC, Alpert MD, Yates NL, Shen X, Koup RA, Pitisuttithum P, Kaewkungwal J, Nitayaphan S, Rerks-Ngarm S, Michael NL, Kim JH. 2012. Immune-correlates analysis of an HIV-1 vaccine efficacy trial. N Engl J Med 366:1275–1286. doi: 10.1056/NEJMoa1113425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Lewis GK. 2014. Role of Fc-mediated antibody function in protective immunity against HIV-1. Immunology 142:46–57. doi: 10.1111/imm.12232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Mabuka J, Nduati R, Odem-Davis K, Peterson D, Overbaugh J. 2012. HIV-specific antibodies capable of ADCC are common in breastmilk and are associated with reduced risk of transmission in women with high viral loads. PLoS Pathog 8:e1002739. doi: 10.1371/journal.ppat.1002739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Asmal M, Sun Y, Lane S, Yeh W, Schmidt SD, Mascola JR, Letvin NL. 2011. Antibody-dependent cell-mediated viral inhibition emerges after simian immunodeficiency virus SIVmac251 infection of rhesus monkeys coincident with gp140-binding antibodies and is effective against neutralization-resistant viruses. J Virol 85:5465–5475. doi: 10.1128/JVI.00313-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Dugast A-S, Tonelli A, Berger CT, Ackerman ME, Sciaranghella G, Liu Q, Sips M, Toth I, Piechocka-Trocha A, Ghebremichael M, Alter G. 2011. Decreased Fc receptor expression on innate immune cells is associated with impaired antibody-mediated cellular phagocytic activity in chronically HIV-1 infected individuals. Virology 415:160–167. doi: 10.1016/j.virol.2011.03.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Moog C, Dereuddre-Bosquet N, Teillaud J-L, Biedma ME, Holl V, Van Ham G, Heyndrickx L, Van Dorsselaer A, Katinger D, Vcelar B, Zolla-Pazner S, Mangeot I, Kelly C, Shattock RJ, Le Grand R. 2014. Protective effect of vaginal application of neutralizing and nonneutralizing inhibitory antibodies against vaginal SHIV challenge in macaques. Mucosal Immunol 7:46–56. doi: 10.1038/mi.2013.23. [DOI] [PubMed] [Google Scholar]
- 7.Xiao P, Zhao J, Patterson LJ, Brocca-Cofano E, Venzon D, Kozlowski PA, Hidajat R, Demberg T, Robert-Guroff M. 2010. Multiple vaccine-elicited nonneutralizing antienvelope antibody activities contribute to protective efficacy by reducing both acute and chronic viremia following simian/human immunodeficiency virus SHIV89.6P challenge in rhesus macaques. J Virol 84:7161–7173. doi: 10.1128/JVI.00410-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Chung AW, Ghebremichael M, Robinson H, Brown EP, Choi I, Lane S, Dugast A-S, Schoen MK, Rolland M, Suscovich TJ, Mahan AE, Liao L, Streeck H, Andrews C, Rerks-Ngarm S, Nitayaphan S, de Souza MS, Kaewkungwal J, Pitisuttithum P, Francis D, Michael NL, Kim JH, Bailey-Kellogg C, Ackerman ME, Alter G. 2014. Polyfunctional Fc-effector profiles mediated by IgG subclass selection distinguish RV144 and VAX003 vaccines. Sci Transl Med 6:228ra38. doi: 10.1126/scitranslmed.3007736. [DOI] [PubMed] [Google Scholar]
- 9.Chung AW, Kumar MP, Arnold KB, Yu WH, Schoen MK, Dunphy LJ, Suscovich TJ, Frahm N, Linde C, Mahan AE, Hoffner M, Streeck H, Ackerman ME, McElrath MJ, Schuitemaker H, Pau MG, Baden LR, Kim JH, Michael NL, Barouch DH, Lauffenburger DA, Alter G. 2015. Dissecting polyclonal vaccine-induced humoral immunity against HIV using systems serology. Cell 163:988–998. doi: 10.1016/j.cell.2015.10.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Ferrari G, Pollara J, Kozink D, Harms T, Drinker M, Freel S, Moody MA, Alam SM, Tomaras GD, Ochsenbauer C, Kappes JC, Shaw GM, Hoxie JA, Robinson JE, Haynes BF. 2011. An HIV-1 gp120 envelope human monoclonal antibody that recognizes a C1 conformational epitope mediates potent antibody-dependent cellular cytotoxicity (ADCC) activity and defines a common ADCC epitope in human HIV-1 serum. J Virol 85:7029–7036. doi: 10.1128/JVI.00171-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Acharya P, Tolbert WD, Gohain N, Wu X, Yu L, Liu T, Huang W, Huang C-C, Do Kwon Y, Louder RK, Luongo TS, McLellan JS, Pancera M, Yang Y, Zhang B, Flinko R, Foulke JS, Sajadi MM, Kamin-Lewis R, Robinson JE, Martin L, Kwong PD, Guan Y, DeVico AL, Lewis GK, Pazgier M. 2014. Structural definition of an antibody-dependent cellular cytotoxicity response implicated in reduced risk for HIV-1 infection. J Virol 88:12895–12906. doi: 10.1128/JVI.02194-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Wang S, Kennedy JS, West K, Montefiori DC, Coley S, Lawrence J, Shen S, Green S, Rothman AL, Ennis FA, Arthos J, Pal R, Markham P, Lu S. 2008. Cross-subtype antibody and cellular immune responses induced by a polyvalent DNA prime-protein boost HIV-1 vaccine in healthy human volunteers. Vaccine 26:3947–3957. doi: 10.1016/j.vaccine.2007.12.060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Kennedy JS, Co M, Green S, Longtine K, Longtine J, O'Neill MA, Adams JP, Rothman AL, Yu Q, Johnson-Leva R, Pal R, Wang S, Lu S, Markham P. 2008. The safety and tolerability of an HIV-1 DNA prime-protein boost vaccine (DP6-001) in healthy adult volunteers. Vaccine 26:4420–4424. doi: 10.1016/j.vaccine.2008.05.090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Liao H- X, Levesque MC, Nagel A, Dixon A, Zhang R, Walter E, Parks R, Whitesides J, Marshall DJ, Hwang K-K, Yang Y, Chen X, Gao F, Munshaw S, Kepler TB, Denny T, Moody MA, Haynes BF. 2009. High-throughput isolation of immunoglobulin genes from single human B cells and expression as monoclonal antibodies. J Virol Methods 158:171–179. doi: 10.1016/j.jviromet.2009.02.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Lefranc M-P, Giudicelli V, Ginestoux C, Jabado-Michaloud J, Folch G, Bellahcene F, Wu Y, Gemrot E, Brochet X, Lane J, Regnier L, Ehrenmann F, Lefranc G, Duroux P. 2009. IMGT, the international ImMunoGeneTics information system. Nucleic Acids Res 37:D1006–D1012. doi: 10.1093/nar/gkn838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Wu X, Yang Z-Y, Li Y, Hogerkorp C-M, Schief WR, Seaman MS, Zhou T, Schmidt SD, Wu L, Xu L, Longo NS, McKee K, O'Dell S, Louder MK, Wycuff DL, Feng Y, Nason M, Doria-Rose N, Connors M, Kwong PD, Roederer M, Wyatt RT, Nabel GJ, Mascola JR. 2010. Rational design of envelope identifies broadly neutralizing human monoclonal antibodies to HIV-1. Science 329:856–861. doi: 10.1126/science.1187659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Montefiori DC. 2005. Evaluating neutralizing antibodies against HIV, SIV, and SHIV in luciferase reporter gene assays. Curr Protoc Immunol Chapter 12:Unit 12.11. doi: 10.1002/0471142735.im1211s64. [DOI] [PubMed] [Google Scholar]
- 18.Pollara J, Hart L, Brewer F, Pickeral J, Packard BZ, Hoxie JA, Komoriya A, Ochsenbauer C, Kappes JC, Roederer M, Huang Y, Weinhold KJ, Tomaras GD, Haynes BF, Montefiori DC, Ferrari G. 2011. High-throughput quantitative analysis of HIV-1 and SIV-specific ADCC-mediating antibody responses. Cytometry A 79:603–612. doi: 10.1002/cyto.a.21084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Koene HR, Kleijer M, Algra J, Roos D, von dem Borne AE, de Haas M. 1997. Fc gammaRIIIa-158V/F polymorphism influences the binding of IgG by natural killer cell Fc gammaRIIIa, independently of the Fc gammaRIIIa-48L/R/H phenotype. Blood 90:1109–1114. [PubMed] [Google Scholar]
- 20.Edmonds TG, Ding H, Yuan X, Wei Q, Smith KS, Conway JA, Wieczorek L, Brown B, Polonis V, West JT, Montefiori DC, Kappes JC, Ochsenbauer C. 2010. Replication competent molecular clones of HIV-1 expressing Renilla luciferase facilitate the analysis of antibody inhibition in PBMC. Virology 408:1–13. doi: 10.1016/j.virol.2010.08.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Pollara J, McGuire E, Fouda GG, Rountree W, Eudailey J, Overman RG, Seaton KE, Deal A, Edwards RW, Tegha G, Kamwendo D, Kumwenda J, Nelson JAE, Liao H-X, Brinkley C, Denny TN, Ochsenbauer C, Ellington S, King CC, Jamieson DJ, van der Horst C, Kourtis AP, Tomaras GD, Ferrari G, Permar SR. 2015. Association of HIV-1 envelope-specific breast milk IgA responses with reduced risk of postnatal mother-to-child transmission of HIV-1. J Virol 89:9952–9961. doi: 10.1128/JVI.01560-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Liao H-XX, Bonsignori M, Alam SM, McLellan JS, Tomaras GD, Moody MA, Kozink DM, Hwang K-K, Chen X, Tsao C-Y, Liu P, Lu X, Parks RJ, Montefiori DC, Ferrari G, Pollara J, Rao M, Peachman KK, Santra S, Letvin NL, Karasavvas N, Yang Z-Y, Dai K, Pancera M, Gorman J, Wiehe K, Nicely NI, Rerks-Ngarm S, Nitayaphan S, Kaewkungwal J, Pitisuttithum P, Tartaglia J, Sinangil F, Kim JH, Michael NL, Kepler TB, Kwong PD, Mascola JR, Nabel GJ, Pinter A, Zolla-Pazner S, Haynes BF. 2013. Vaccine induction of antibodies against a structurally heterogeneous site of immune pressure within HIV-1 envelope protein variable regions 1 and 2. Immunity 38:176–186. doi: 10.1016/j.immuni.2012.11.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Tuero I, Mohanram V, Musich T, Miller L, Vargas-Inchaustegui DA, Demberg T, Venzon D, Kalisz I, Kalyanaraman VS, Pal R, Ferrari MG, LaBranche C, Montefiori DC, Rao M, Vaccari M, Franchini G, Barnett SW, Robert-Guroff M. 2015. Mucosal B cells are associated with delayed SIV acquisition in vaccinated female but not male rhesus macaques following SIVmac251 rectal challenge. PLoS Pathog 11:e1005101. doi: 10.1371/journal.ppat.1005101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Chung AW, Navis M, Isitman G, Wren L, Silvers J, Amin J, Kent SJ, Stratov I. 2011. Activation of NK cells by ADCC antibodies and HIV disease progression. J Acquir Immune Defic Syndr 58:127–131. doi: 10.1097/QAI.0b013e31822c62b9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Guan Y, Pazgier M, Sajadi MM, Kamin-Lewis R, Al-Darmarki S, Flinko R, Lovo E, Wu X, Robinson JE, Seaman MS, Fouts TR, Gallo RC, DeVico AL, Lewis GK. 2013. Diverse specificity and effector function among human antibodies to HIV-1 envelope glycoprotein epitopes exposed by CD4 binding. Proc Natl Acad Sci U S A 110:E69–E78. doi: 10.1073/pnas.1217609110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Tomaras GD, Ferrari G, Shen X, Alam SM, Liao H-X, Pollara J, Bonsignori M, Moody MA, Fong Y, Chen X, Poling B, Nicholson CO, Zhang R, Lu X, Parks R, Kaewkungwal J, Nitayaphan S, Pitisuttithum P, Rerks-Ngarm S, Gilbert PB, Kim JH, Michael NL, Montefiori DC, Haynes BF. 2013. Vaccine-induced plasma IgA specific for the C1 region of the HIV-1 envelope blocks binding and effector function of IgG. Proc Natl Acad Sci U S A 110:9019–9024. doi: 10.1073/pnas.1301456110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Bonsignori M, Pollara J, Moody MA, Alpert MD, Chen X, Hwang KK, Gilbert PB, Huang Y, Gurley TC, Kozink DM, Marshall DJ, Whitesides JF, Tsao CY, Kaewkungwal J, Nitayaphan S, Pitisuttithum P, Rerks-Ngarm S, Kim JH, Michael NL, Tomaras GD, Montefiori DC, Lewis GK, DeVico AL, Evans DT, Ferrari G, Liao HX, Haynes BF. 2012. Antibody-dependent cellular cytotoxicity-mediating antibodies from an HIV-1 vaccine efficacy trial target multiple epitopes and preferentially use the VH1 gene family. J Virol 86:11521–11532. doi: 10.1128/JVI.01023-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Shields RL, Namenuk AK, Hong K, Meng YG, Rae J, Briggs J, Xie D, Lai J, Stadlen A, Li B, Fox JA, Presta LG. 2001. High resolution mapping of the binding site on human IgG1 for Fc gamma RI, Fc gamma RII, Fc gamma RIII, and FcRn and design of IgG1 variants with improved binding to the Fc gamma R. J Biol Chem 276:6591–6604. doi: 10.1074/jbc.M009483200. [DOI] [PubMed] [Google Scholar]
- 29.Pollara J, Bonsignori M, Moody MA, Pazgier M, Haynes BF, Ferrari G. 2013. Epitope specificity of human immunodeficiency virus-1 antibody dependent cellular cytotoxicity [ADCC] responses. Curr HIV Res 11:378–387. doi: 10.2174/1570162X113116660059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Pollara J, Bonsignori M, Moody MA, Liu P, Alam SM, Hwang K-K, Gurley TC, Kozink DM, Armand LC, Marshall DJ, Whitesides JF, Kaewkungwal J, Nitayaphan S, Pitisuttithum P, Rerks-Ngarm S, Robb ML, O'Connell RJ, Kim JH, Michael NL, Montefiori DC, Tomaras GD, Liao H-X, Haynes BF, Ferrari G. 2014. HIV-1 vaccine-induced C1 and V2 Env-specific antibodies synergize for increased antiviral activities. J Virol 88:7715–7726. doi: 10.1128/JVI.00156-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ackerman ME, Dugast A-S, McAndrew EG, Tsoukas S, Licht AF, Irvine DJ, Alter G. 2013. Enhanced phagocytic activity of HIV-specific antibodies correlates with natural production of immunoglobulins with skewed affinity for FcγR2a and FcγR2b. J Virol 87:5468–5476. doi: 10.1128/JVI.03403-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Barouch DH, Alter G, Broge T, Linde C, Ackerman ME, Brown EP, Borducchi EN, Smith KM, Nkolola JP, Liu J, Shields J, Parenteau L, Whitney JB, Abbink P, Ng'ang'a DM, Seaman MS, Lavine CL, Perry JR, Li W, Colantonio AD, Lewis MG, Chen B, Wenschuh H, Reimer U, Piatak M, Lifson JD, Handley SA, Virgin HW, Koutsoukos M, Lorin C, Voss G, Weijtens M, Pau MG, Schuitemaker H. 2015. HIV-1 vaccines. Protective efficacy of adenovirus/protein vaccines against SIV challenges in rhesus monkeys. Science 349:320–324. [DOI] [PMC free article] [PubMed] [Google Scholar]