

The p53 tumor suppressor plays pivotal roles in normal growth, development, and tumor suppression. A key function of p53 as a transcription factor is to induce target genes involved in cell cycle arrest, apoptosis, and senescence.1 In its active form, p53 is tetrameric, which is controlled by the tetramerization domain. Indeed, the DNA-binding activity of a p53 tetramer is ~100-fold higher than that of a p53 monomer. Thus, the tetramerization domain is essential for p53-mediated tumor suppression.2
Most studies on the tetramerization domain are focused on its ability to regulate p53 oligomerization and DNA-binding activity. Interestingly, an elegant study recently published in Cell Cycle by Dr. Prives and her colleagues showed that 2 lysine residues (K351 and K357) in the tetramerization domain differentially modulate p53-dependent cell cycle arrest and apoptosis.3 Mutations of these 2 lysine residues to arginines (2KR-p53), which preserve electric charge but block acetylation, or to glutamines (2KQ-p53), which mimic acetylation but neutralize electric charge, have little effect on the tetramerization, localization, and stability of p53. Indeed, the 2KR-p53 mutant exerts similar activities as wild-type p53 in DNA binding and transcriptional regulation of downstream targets. Surprisingly, the 2KQ-p53 mutant is deficient in transactivation of growth-arrest targets, including p21 and Cyclin G1, but not proapoptotic targets, such as PUMA, PIG3, and Noxa. Most importantly, Dr. Prives and her colleagues showed that the 2KQ-p53 mutant, but not the 2KR-p53 mutant, fails to induce cell cycle arrest. These data indicate that acetylation at K351 and/or K357 may serve as a “molecular switch” of p53 to drive apoptosis vs. cell cycle arrest. Herein, a novel function of p53 tetramerization domain in cell fate control has been identified by Dr. Prives and her colleagues (Fig. 1).
Figure 1.
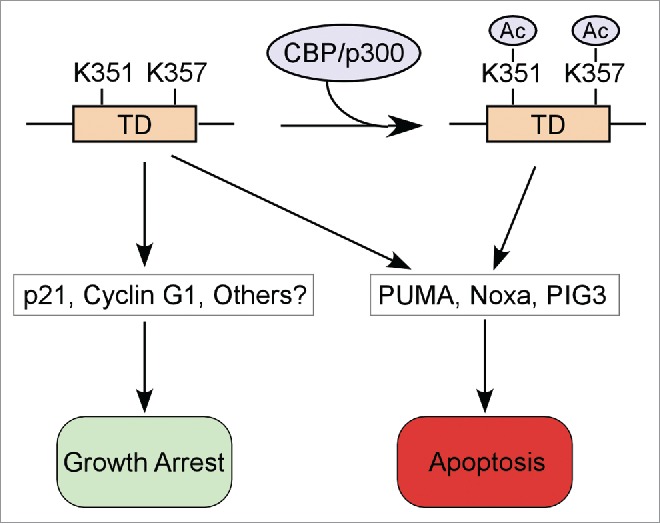
A proposed model for the role of p53 tetramerization domain in cell fate control.
In spite of extensive efforts, how p53 controls cell fate, namely cell cycle arrest, apoptosis, and/or senescence, is still not well understood. One potential mechanism for such control has been ascribed to the post-translational modifications of p53, which may affect the p53 molecule and its selectively binding to the promoters of target genes. For example, previous studies have shown that several p53 mutants, including K120R and R175P, which are defective in apoptotic induction, are still able to regulate cell cycle arrest.4-7 Notably, all of these sites are located in the DNA-binding domain and have an opposing effect on cell cycle arrest as observed with the K351 and K357 residues.
The new function of p53 tetramerization domain raises a few intriguing questions and possibly new research directions: Is there any interplay between the modifications of lysine K351/357 and other site(s)? Does acetylation of p53 at K351/357 affect other cellular outcomes, such as senescence, metabolism, and autophagy? Are there any other targets involved in cell cycle arrest that are blocked by acetylation of p53 at K351/357? The answers to these and other questions will provide an insight into p53 biology, and lay a foundation for developing new therapies by turning off cell cycle arrest to maximize p53-induced cell death.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
References
- [1].Vousden KH, Prives C. Blinded by the Light: The Growing Complexity of p53. Cell 2009; 137(3):413-31; PMID:19410540; http://dx.doi.org/ 10.1016/j.cell.2009.04.037 [DOI] [PubMed] [Google Scholar]
- [2].Chene P. The role of tetramerization in p53 function. Oncogene 2001; 20(21):2611-7; PMID:11420672; http://dx.doi.org/ 10.1038/sj.onc.1204373 [DOI] [PubMed] [Google Scholar]
- [3].Beckerman R, et al.. Lysines in the tetramerization domain of p53 selectively modulate G1 arrest. Cell Cycle 2016; 15(11):1425-38; PMID:27210019; http://dx.doi.org/ 10.1080/15384101.2016.1170270 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [4].Rowan S, et al.. Specific loss of apoptotic but not cell-cycle arrest function in a human tumor derived p53 mutant. EMBO J 1996; 15(4):827-38; PMID:8631304 [PMC free article] [PubMed] [Google Scholar]
- [5].Sykes SM, et al.. Acetylation of the p53 DNA-binding domain regulates apoptosis induction. Mol Cell 2006; 24(6):841-51; PMID:17189187; http://dx.doi.org/ 10.1016/j.molcel.2006.11.026 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [6].Tang Y, et al.. Tip60-dependent acetylation of p53 modulates the decision between cell-cycle arrest and apoptosis. Mol Cell 2006; 24(6):827-39; PMID:17189186; http://dx.doi.org/ 10.1016/j.molcel.2006.11.021 [DOI] [PubMed] [Google Scholar]
- [7].Rokudai S, et al.. MOZ increases p53 acetylation and premature senescence through its complex formation with PML. Proc Natl Acad Sci U S A 2013; 110(10):3895-900; PMID:23431171; http://dx.doi.org/ 10.1073/pnas.1300490110 [DOI] [PMC free article] [PubMed] [Google Scholar]