

Abstract
Background
Inherited retinal diseases (IRDs) are a clinically and genetically heterogeneous set of disorders, for which diagnostic second-generation sequencing (next-generation sequencing, NGS) services have been developed worldwide.
Methods
We present the molecular findings of 537 individuals referred to a 105-gene diagnostic NGS test for IRDs. We assess the diagnostic yield, the spectrum of clinical referrals, the variant analysis burden and the genetic heterogeneity of IRD. We retrospectively analyse disease-causing variants, including an assessment of variant frequency in Exome Aggregation Consortium (ExAC).
Results
Individuals were referred from 10 clinically distinct classifications of IRD. Of the 4542 variants clinically analysed, we have reported 402 mutations as a cause or a potential cause of disease in 62 of the 105 genes surveyed. These variants account or likely account for the clinical diagnosis of IRD in 51% of the 537 referred individuals. 144 potentially disease-causing mutations were identified as novel at the time of clinical analysis, and we further demonstrate the segregation of known disease-causing variants among individuals with IRD. We show that clinically analysed variants indicated as rare in dbSNP and the Exome Variant Server remain rare in ExAC, and that genes discovered as a cause of IRD in the post-NGS era are rare causes of IRD in a population of clinically surveyed individuals.
Conclusions
Our findings illustrate the continued powerful utility of custom-gene panel diagnostic NGS tests for IRD in the clinic, but suggest clear future avenues for increasing diagnostic yields.
Keywords: next-generation sequencing, Molecular genetics, inherited retinal dystrophy, bioinformatics, retinitis pigmentosa
Introduction
Inherited retinal diseases (IRDs) are a diverse set of Mendelian disorders that are a major cause of inherited blindness across the world. They are caused by the progressive deterioration or the early loss of cells fundamental for the normal function of the retina,1 the component of the eye responsible for converting light energy into electrical signals.
IRDs are clinically heterogeneous and vary widely in their severity, age of onset, pathogenesis, manner of disease progression and inheritance pattern. IRDs distinguishable through clinical and electrophysiological investigation include: rod-cone dystrophy and retinitis pigmentosa (RCD/RP); cone dystrophy (CD); cone-rod dystrophy (CRD); macular dystrophy and Stargardt disease (MD/STGD); early-onset retinal disease and Leber congenital amaurosis (EORD/LCA); congenital stationary night blindness (CSNB); and familial exudative vitreo-retinopathy. Notably, IRD can be a feature of a multisystem disorder such as Usher syndrome, Bardet–Biedl syndrome (BBS), Senior–Loken syndrome and Joubert syndrome.
Decades of research have elucidated the genetic basis of IRD, revealing a suite of mutations in over 200 genes.2 The genes associated with IRDs can be involved in a variety of processes and functions,3 and their tissue expression can range from exclusive expression in the retina to ubiquitous expression across the body. IRDs are predominantly monogenic but a range of inheritance patterns have been described, including: autosomal-dominant, autosomal-recessive, X-linked, digenic and mitochondrial inheritance.
The identification of the genetic basis of IRD can greatly assist the clinical diagnosis, counselling, treatment and management received on a patient-by-patient basis. Since the inception of second-generation DNA sequencing technologies (commonly referred to as next-generation sequencing, NGS)—techniques permitting the surveillance of genetic variation within multiple genes through a single experiment4—many clinical laboratories have adopted NGS as a tool for the diagnosis of rare genetic diseases,5–7 including IRD.8 9 We previously described a custom-gene panel NGS diagnostic service for individuals with IRD.10 This follow-up report outlines the success of this 105 gene diagnostic test for the first 537 individuals with IRD referred from worldwide institutions and describes 131 new mutations in 45 genes as a potential cause of disease.
Materials and methods
Patient referrals
Patient referrals were made from worldwide clinical institutions. There were no specific requirements for clinical phenotypic descriptions of IRD before diagnostic genomic testing was undertaken, although the reason for referral was requested. All experiments had been approved by the Greater Manchester West Research Ethics Committee and were performed in a UK Accreditation Service Clinical Pathology Accredited Medical Laboratory.
Enrichment and sequencing
DNA samples were enriched for the coding regions ±50 bp of 105 genes (see online supplementary table S1) and a specified intronic region of the CEP290 gene, using an Agilent SureSelect Custom Design target-enrichment kit (Agilent, Santa Clara, California, USA). DNA samples were indexed with a unique paired-end barcode and then subjected to multiplexed high-throughput parallel sequencing using either the ABI SOLiD 5500 platform (n=235; Life Technologies, Carlsbad, California, USA) or the Illumina HiSeq 2000/2500 (n=302; Illumina, San Diego, California, USA), following manufacturer's protocols.
jmedgenet-2016-103837supp_tables.pdf (3.9MB, pdf)
Variant calling
Sequencing reads were demultiplexed and aligned to the hg19 reference genome before duplicate read removal and variant calling was performed. For ABI SOLiD sequencing reads, demultiplexing, alignment and variant calling were all performed using Lifescope Genomic Analysis software. For Illumina HiSeq sequencing reads, demultiplexing was performed using CASAVA V.1.8.2. (Illumina), alignment was performed using Burrows-Wheeler Aligner short read (BWA-short V.0.6.2) software,11 and variant calling was performed using the UnifiedGenotyper12 within the Genome Analysis Tool Kit13 (GATK-lite V.2.0.39), after base quality score recalibration and indel realignment. To reduce the number of potential false-positive variants from NGS, we primarily limited the clinical analysis of genomic variants to those with sequencing quality metrics above specific criteria. These metrics were calculated as a result of pilot and control sample analyses, which have been previously published.10 14 For the ABI SOLiD, we considered single-nucleotide variants (SNVs) with ≥18× sequencing depth and a minimum mean quality value (MQV) >18, and considered indels with support from >5× independent sequencing reads. For the Illumina HiSeq, we considered SNVs with ≥50× independent sequencing reads and ≥45 MQV, and considered indels with support from >25% of the aligned and independent sequencing reads.
Variant interpretation
A summary of our strategy to clinically interpret genetic variants is provided in online supplementary table S2. Clinical interpretation was restricted to variants within coding regions ±5 bp of the 105 genes included in online supplementary table S1 and an intronic variant, c.2991+1655A>G, in the CEP290 gene. We considered variants with a frequency >1% in control population databases (Exome Variant Server, ESP-6500; dbSNP V.135) as benign polymorphisms, providing the cohort size was sufficiently large and diverse, and the sequencing read depth exceeded an average of 18× in the Exome Variant Server (EVS). Variants with a frequency below 1% in SNP databases but with a high recurrence rate in-house were classified as neutral or proven to be NGS run artefacts by Sanger sequencing. The pathogenicity of the remaining genetic variants was determined through extensive appraisal of the variant's predicted consequence (annotations performed against the Ensembl V.68 database), the scientific literature, the patient's clinical referral (genotype–phenotype correlations) and in silico modelling including SIFT15 and PolyPhen-2.16 These criteria for variant interpretation are consistent with the guidelines recently outlined by the American College of Medical Genetics.17
Clinical decision making
A confirmed molecular diagnosis was provided for individuals with variants determined as ‘clearly pathogenic’ or ‘likely pathogenic’ (see online supplementary table S2) in a disease-causing state (eg, homozygous or compound heterozygous in a gene known to cause recessive IRD). A provisional molecular diagnosis was reported for individuals with ‘variants of unknown clinical significance’ (see online supplementary table S2) that were found in a disease-causing state (eg, compound heterozygous with another ‘clearly pathogenic’ variant in a gene known to cause recessive IRD). If ‘clearly pathogenic’ or ‘likely pathogenic’ variants were identified in more than one gene in a disease-causing state the family history, phenotypic presentation and evidence for variant pathogenicity was discussed in detail within a clinical multidisciplinary team meeting. Variants determined as ‘clearly pathogenic’ or ‘likely pathogenic’ in a carrier state (eg, heterozygous in a gene known to cause recessive IRD) were clinically reported as carrier findings. Cascade testing was offered for relevant family members to assist with the interpretation of variant pathogenicity and to clarify the risks to additional family members.
Variant validation
We performed PCR and bidirectional capillary sequencing to confirm the zygosity and the presence of disease-causing and carrier variants before they were clinically reported.
Retrospective variant analysis
All clinically analysed SNVs were also retrospectively compared with the Exome Aggregation Consortium (ExAC) database in October 2015 to determine their frequency within a large and ethnically diverse population data set. Novel and potentially disease-causing variants, absent from the literature at the time of clinical analysis, were retrospectively compared with the Human Gene Mutation Database, HGMD,18 in October 2015. Mutated genes determined to account for molecular diagnoses were compared with their dates of discovery, available in the RetNet database.2
Results
Clinical details
Demographics and classifications
We performed diagnostic NGS testing for 537 patients (287 males and 250 females) referred with clinical indications of IRD. All of the 537 individuals were initially referred on a singleton basis.
Phenotype classification
IRDs have a diverse spectrum of overlapping clinical presentation, which can vary by age of onset, extent of visual impairment, nature of disease progression and involvement of additional clinical features. We grouped all referred patients into 10 distinct clinical classifications (see online supplementary table S3). The most common referral was non-syndromic RCD/RP (n=250). The most common referral of syndromic disease was Usher syndrome (n=38), a disorder characterised by neurosensory hearing loss and visual impairment. There were eight cases of suspected syndromic ciliopathies referred, including seven cases of BBS.
Molecular results
Sequencing statistics
A total of 2 089 243 343 aligned, ontarget and unique sequencing reads were generated by NGS. For the 235 patients analysed through ABI SOLiD sequencing, an average of 6 537 522 aligned, ontarget and unique sequencing reads were generated for each patient. For the 302 patients analysed through Illumina HiSeq sequencing, an average of 1 830 879 aligned, ontarget and unique sequencing reads were generated for each patient. The diagnostic assay achieved an average coverage at ≥20× for 98.4% and at ≥50× for 97.3% of the region clinically analysed for the 537 referred patients (see online supplementary figure S1). Sequencing data generated through the Illumina HiSeq covered significantly more of the clinically analysed region than sequencing data generated through the ABI SOLiD at ≥20× coverage (see online supplementary figure S1; HiSeq=99.0±0.01%, SOLiD=97.6±0.05%, p<0.0001) and ≥50× coverage (see online supplementary figure S1; HiSeq=98.3±0.03%, SOLiD=95.9±0.09%, p<0.0001).
Variant analysis burden
A total of 143 675 variants were initially identified in the 537 patients (table 1). Using a bioinformatics pipeline, we filtered the variants for their quality and frequency in control populations. This restricted the number of variants for analysis to 4542, an average of 8.4 variants per referred individual. Most of the clinically analysed variants were single-nucleotide alterations (SNVs), with an average of 8.1 per patient. Heterozygous SNVs provided the largest burden for analysis, with 4172 detected (an average of 7.8 per patient), but only 6% (n=252) reported as a cause of disease after extensive clinical analysis (table 1). There were 2266 unique SNVs clinically analysed, with 574 identified in more than 1 individual. An indel event was clinically analysed, on average, in a third of the referred patients.
Table 1.
The total number of genetic variants found during the diagnostic screening process
| Type | Hom | Het | Hemi | Het–het | Total |
|---|---|---|---|---|---|
| Raw calls | |||||
| SNVs | 46 405 | 90 981 | 338 | 82 | 137 806 |
| Indels | 2182 | 3653 | 12 | 22 | 5869 |
| Total | 48 587 | 94 634 | 350 | 104 | 143 675 |
| Clinically analysed | |||||
| SNVs | 166 | 4172 | 20 | 0 | 4358 |
| Indels | 27 | 150 | 7 | 0 | 184 |
| Total | 193 | 4322 | 27 | 0 | 4542 |
| Clinically reported | |||||
| SNVs | 58 | 252 | 9 | 0 | 319 |
| Indels | 31 | 48 | 4 | 0 | 83 |
| Total | 89 | 300 | 13 | 0 | 402 |
The zygosity of raw calls and clinically analysed variants is estimated from the sequencing read pileups of next-generation sequencing (NGS) data. The zygosity of clinically reported variants is confirmed through an alternative technique. hom, homozygous variants; het, heterozygous variants, hemi, hemizygous variants found on chrX in males; het–het, variants with two unique alternative alleles differing from the reference allele (hg19).
At the time of analysis, 30% of SNVs (1311/4358) and 71% of indels (131/184) were neither present in dbSNP or in EVS. This included 811 missense variants, 70 nonsense variants, 105 coding region indel events, 26 splice region indel events, 350 synonymous variants, 2 start codon variants, 32 canonical splice site variants and 46 splice region variants (novel; figure 1). We clinically analysed 135 deletion events (105 heterozygous and 30 homozygous), 48 insertion events (46 heterozygous and 2 homozygous) and 1 heterozygous deletion–insertion event. Eighty percent (147/184) of the indel events were present in the coding regions of genes, and 110 of these were expected to cause premature termination of the protein product (84 heterozygous and 26 homozygous). A small number of clinically analysed variants were determined as false positives after analysis through PCR and bidirectional capillary sequencing techniques (see online supplementary table S4).
Figure 1.

The variant analysis burden. The numbers of variants analysed by clinically accredited scientists by their expected consequences on the encoded protein and frequency in control populations. Annotations are performed against the specified transcripts in online supplementary table S1 using V.68 of the Ensembl database and population frequencies available in dbSNP and EVS.
Clinical decision making
We identified a molecular diagnosis for 271 of the 537 referred patients (51%). The molecular findings underpinning these diagnoses are reported in online supplementary table S5. We have provided a molecular diagnosis for recessive IRD in 208 cases (homozygous, n=88; compound heterozygous, n=120), dominant IRD in 50 cases and X-linked IRD in 13 cases. We found no clear instances of digenic or polygenic inheritance. We informed clinicians of 154 individuals who carry ‘clearly pathogenic’ or ‘likely pathogenic’ variants in a carrier state, that is, heterozygous in a gene known to underpin disease inherited in a recessive manner, 59 of whom received a molecular diagnosis after the identification of ‘clearly pathogenic’ or ‘likely pathogenic’ variants in other disease-causing genes (dominant IRD, n=11; recessive IRD, n=48). Eighty families were referred for cascade testing after receiving molecular results from NGS testing for the referred proband. We confirmed compound heterozygous variants to be in-trans in 36 cases, confirmed homozygosity in 11 cases, identified de novo variants in 3 cases and identified 16 cases of additional variant segregation with IRD. In a single case, we identified heterozygous variants underpinning a provisional molecular diagnosis to be in-cis, altering the counselling that was received by the referred individual.
Several individuals were found to have ‘clearly pathogenic’ or ‘likely pathogenic’ variants in a disease-causing state that would not be expected to cause the IRD subtype declared by the referring clinician (see online supplementary table S6). For example, two individuals referred with clinical indications of RP were found to have ‘likely pathogenic’ mutations in the CHM gene, a cause of choroideremia. These genomic findings led to the re-evaluation of initial clinical descriptions, and in some cases refined or altered the initial clinical diagnosis. In a single case (13005797; online supplementary table S5), bidirectional capillary sequencing targeted at a region of GUCY2D, which had been poorly covered through NGS (460 nucleotides of a coding exon with <50× coverage) identified a novel heterozygous missense variant, c.380C>T p.(Pro127Arg), that had not been identified through diagnostic NGS. This large protein-coding exon of GUCY2D (chr17:7906361–7907091) has a GC content of 76.20%. Homozygous and compound heterozygous mutations in GUCY2D are known to cause EORD/LCA.19 When considered in combination with a heterozygous frameshift mutation in GUCY2D, c.2595delG, the identified missense variant was concluded to likely account for a molecular diagnosis of autosomal-recessive EORD/LCA.
Genetic heterogeneity
In total, 402 genetic variants are reported to account for a molecular diagnosis of IRD (273 variants with a single occurrence and 38 variants reported multiple times). Variants reported in multiple individuals ranged from presence in 2 (n=22) to 11 patients (n=1). The most commonly reported variant is CERKL c.375C>G p.(Cys125Trp), which is reported to coincide with clinical presentations of IRD from four disease subgroups: one case of CD; three cases of CRD; four cases of RCD/RP and three cases of MD/STGD.
Variants accounting for a molecular diagnosis of IRD were found in 62 of the 105 genes included in the diagnostic NGS assay (figure 2). Most of the genes with disease-causing variants are restricted to a single disease referral category (see online supplementary table S7; 66%, n=41), the most frequent of which are mutations causing recessively inherited RCD/RP in the EYS gene (n patients=7) and dominantly inherited RCD/RP in the RHO gene (n patients=6). The remaining 21 genes are reported with pathogenic variants in patients referred from two or more disease referral categories (see online supplementary table S7). The gene associated with the most highly variable phenotype is ABCA4, which was concluded to contain pathogenic variants causing IRD in 23 patients referred from five discrete referral categories, including: 12 patients with MD/STGD; 5 with CRD; 4 with RCD/RP and 1 individual with CD. Disruption of the ABCA4 gene was also concluded to be the cause of visual impairment in a single patient referred with Usher syndrome but is not expected to contribute to the clinical presentation of hearing loss.
Figure 2.

The genetic basis of inherited retinal disease in 271 individuals with a confirmed or provisional molecular diagnosis. Each segment illustrates the number of individuals with genetic variants determined as a cause of inherited retinal disease.
Mutation consequences
We determined 311 unique mutations to be the cause of disease. These included 137 missense variants, 20 canonical splice site variants, 76 nonsense variants, 54 out-of-frame insertion or deletion events, 1 variant disrupting the start codon, 6 synonymous variants, 9 splice region variants, 1 intronic variant and 7 inframe deletions. Fifty-two percent (162/311) of the variants reported to account for a molecular diagnosis were neither present in dbSNP or EVS. Specifically, 71% (97/137) of the clinically reported missense variants, 55% (42/76) of clinically reported nonsense SNVs and 63% (34/54) of clinically reported out-of-frame indel events were neither present in dbSNP or EVS.
Of the 137 clinically reported missense variants, 30 had functional support of their disruption to the encoded protein, 49 had disease segregation evidence in the literature without functional support, 6 had evidence for an alternative amino acid substitution at the same residue as a cause of disease and 52 were determined as a novel cause of disease (see online supplementary table S8). One hundred and thirty clinically reported variants are expected to cause premature termination during protein synthesis, 76 of these are SNVs and 54 are out-of-frame small insertion or deletion events. Fifty-one percent (66/130) of these loss-of-protein-function events were determined as novel causes of disease; this included 35 nonsense SNVs and 31 small indels (see online supplementary table S8).
Retrospective variant analysis
The average allele frequency in ExAC of the 2266 unique clinically analysed SNVs was 0.12; however, 12% (517/4358) of all clinically analysed SNVs were found to be at a greater frequency than 1% in the ExAC database. The average allele frequency in ExAC of variants clinically reported as a cause of disease was 0.02 (median=0 and max=0.78).
One hundred and forty-four variants were determined as novel causes of disease at the time of clinical analysis. Ninety-one percent (131/144) of these variants remained novel causes of disease after comparison with the HGMD in October 2015. This included 46 missense variants, 12 canonical splice site variants, 33 nonsense variants, 28 out-of-frame insertion or deletion events, 4 synonymous variants, 5 splice region variants and 3 inframe deletions. All of these variants are reported in online supplementary table S8 and have been submitted to the ClinVar database.
Sixty-five of the 105 genes surveyed (62%) were identified between 1995 and 2004, and mutations in these genes accounted for a molecular diagnosis in 214 of the 271 cases (79%; figure 3). This equates to a rate of 3.3 diagnoses per gene analysed. In contrast, the 33 genes included in our analysis that were identified post-2005 only accounted for 41 of the 271 molecular diagnoses (15%; figure 3), a rate of 1.2 diagnoses per gene analysed.
Figure 3.
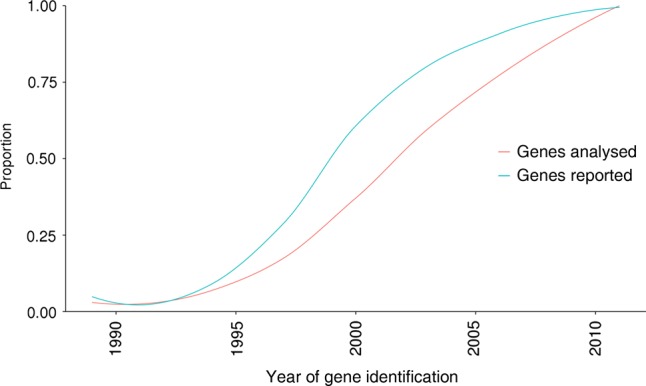
The relationship between the genes accounting for molecular diagnoses and the year of their discovery. The curves illustrate the trends in the proportion of 271 molecular diagnoses accounted for by the analysis of 105 genes through diagnostic next-generation sequencing.
Discussion
Developments in DNA sequencing technology have enabled the transition from single gene and known mutation diagnostic assays to comprehensive surveys of variation within all genes known to cause disease phenotypes. This has shifted the diagnostics bottleneck from the detection of disease-causing variation to the clinical interpretation of the plethora of genetic variants identified. Here, we discuss the molecular findings from a NGS diagnostic test that has been employed in a clinical context to survey genes known to be associated with IRD, a clinically and genetically heterogeneous set of Mendelian disorders that affect millions of individuals worldwide.20 The findings derived from a large cohort of individuals referred specifically for diagnostic genomic testing.
We have clinically reported 402 mutations as a cause or a potential cause of disease in 62 of the 105 genes surveyed. These variants account or likely account for the clinical diagnosis of IRD in 51% of the 537 individuals referred for diagnostic testing, including 208 autosomal-recessive, 50 autosomal-dominant and 13 X-linked recessive cases. Other studies employing NGS have shown higher molecular diagnostic rates than reported in this study, including Eisenberger and colleagues,21 Zhao and colleagues22 and the Saudi Mendeliome group.23 We attribute differences in diagnostic success rates to a number of factors. First, the diagnostic service is provisioned for worldwide referral institutions on a singleton basis and there are no prespecified criteria for the clinical phenotyping of referred individuals. Detailed clinical phenotyping in complement to genomic analysis has been shown to discover and accelerate diagnoses in the clinic.24 25 Second, the 105 gene enrichment does not include all genes that are now known as a cause of IRD, and application of whole exome sequencing (WES) or whole genome sequencing (WGS) techniques to individuals without a molecular diagnosis has identified variants in other genes as a cause of disease, for example, mutations in IQCB1 identified through WES.26 It is of note that we have recently expanded the diagnostic service to include an additional 71 genes and pathogenic intronic variants associated with IRD. This expansion intends to meet the clinical need for the surveillance of genes known to underpin CSNB. Third, there were technical limitations with early versions of the diagnostic service, for example, insufficient coverage achieved by the ABI SOLiD sequencing platform over all 288 087 DNA bases which are clinically surveyed (see online supplementary figure S1).
The use of NGS in the clinic has drastically increased the diagnostic yields routinely achieved for IRD. However, there still remains an inability to clinically survey the final AG-rich exon of RPGR (orf15) through the NGS techniques described in this study and by others. Mutations in RPGRorf15 are well documented as a major cause of X-linked RP,27 but bidirectional capillary sequencing techniques remain the gold standard clinical screening tool for this region of the genome.28 Complementing diagnostic NGS services with bidirectional capillary sequencing of regions which remain elusive to NGS will increase diagnostic yields. Moreover, genetic variant detection in clinical diagnostics requires clinically validated bioinformatics techniques. Such techniques have evolved with the clinical requirements of diagnostic services. Validation of bioinformatics techniques to detect complex insertion and deletion events, for example, Pindel,29 and complete exon deletions and duplications, for example, ExomeDepth,30 will undoubtedly increase the diagnostic yields described in this study, and in others. Finally, the expansion of analyses to non-coding and regulatory regions of known disease-causing genes will increase diagnostic yields for IRD. We observed an interesting trend in our cohort of predominantly Western European and South Asian individuals, which suggests that newly identified genes are becoming increasingly rarer causes of disease (figure 3). The striking observation that 79% of cases with molecular diagnoses are accounted for by mutations in genes discovered between 1995 and 2004 suggests that while the continued addition of new disease-causing genes will of course improve diagnostic services, the comprehensive evaluation of protein-coding and regulatory variants affecting known disease-causing genes may have a greater influence on diagnostic yield. For example, the genomic sequencing of genes known to cause IRD has identified large deletions,14 deeply intronic disease-causing mutations31 and variants in regulatory regions,32 which would elude traditional custom panel NGS analysis.
A significant complication of implementing NGS as a diagnostic service is the number of variants, which require clinical interpretation on a patient-by-patient basis (table 1), many of which have not previously been defined in control populations (figure 1). However, a clear advantage of NGS diagnostic approaches over other high-throughput techniques is the capability to detect novel disease-causing variation,33 defined as variants that have not previously been reported as disease causing in the literature or in the HGMD. We have reported 144 novel disease-causing variants as a possible cause of IRD (see online supplementary table S8). Thirteen of these variants determined as novel at the time of clinical analysis have since been described as disease causing in other studies, for example, USH2A c.11713C>T p.(Arg3905Cys)34 and PRPF31 c.1060C>T p.(Arg354Ter).35 Many of the reported novel variants are expected to cause disruption of normal translation, including 35 nonsense variants, 6 disruptions of canonical splice sites and 31 out-of-frame coding indels. Most novel variants are clinically reported on a single occasion; however, some are reported to cause disease in multiple individuals, for example, CERKL c.193G>T p.(Glu65Ter) in five individuals and EYS c.1155T>A p.(Cys385Ter) in three individuals. None of the novel missense variants, splice region variants or inframe indels reported in this study are determined to cause disease on multiple occasions, and further disease segregation and/or functional study will greatly assist their clinical interpretation.
In summary, we illustrate the powerful clinical utility of a diagnostic NGS test for individuals with inherited visual impairment. We add to evidence of known pathogenic mutations through further correlation in individuals who present with clinical indications of IRD (see online supplementary table S5) and outline 131 novel disease-causing variants that are not previously described in individuals with IRD.
Acknowledgments
This work was supported by Fight for Sight, RP Fighting Blindness, the Biotechnology and Biological Sciences Research Council, the Manchester Biomedical Research Centre and the DST UK-India Education and Research Initiative. We thank referring clinicians and acknowledge contributions from the UK Inherited Retinal Disease Consortium.
Footnotes
Contributors: Design and coordination of the study: GCMB, JME, SCR. Contributing genetic and/or phenotypic data: JME, StB, SaB, JOS, SGW, PIS, RLG, SPD, GH, TG, ICL, PNB, SCR, GCMB. Writing of the manuscript: JME and GCMB. Revision of the manuscript with important intellectual content: all authors.
Funding: UK-India Education and Research Initiative, Fight for Sight UK, Biotechnology and Biological Sciences Research Council, RP Fighting Blindness.
Competing interests: None declared.
Ethics approval: Greater Manchester West Research Ethics Committee.
Provenance and peer review: Not commissioned; externally peer reviewed.
Data sharing statement: Novel variants, identified in an NHS laboratory are shared through ClinVar.
References
- 1.den Hollander AI, Black A, Bennett J, Cremers FPM. Lighting a candle in the dark: advances in genetics and gene therapy of recessive retinal dystrophies. J Clin Invest 2010;120:3042–53. 10.1172/JCI42258 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Daiger SP, Sullivan LS, Bowne SJ. RetNet: Retinal Information Network, 1996. https://sph.uth.edu/retnet/ (Accessed April 2016).
- 3.Wright AF, Chakarova CF, Abd El-Aziz MM, Bhattacharya SS. Photoreceptor degeneration: genetic and mechanistic dissection of a complex trait. Nat Rev Genet 2010;11:273–84. 10.1038/nrg2717 [DOI] [PubMed] [Google Scholar]
- 4.Metzker ML. Sequencing technologies—the next generation. Nat Rev Genet 2010;11:31–46. 10.1038/nrg2626 [DOI] [PubMed] [Google Scholar]
- 5.Yang Y, Muzny DM, Xia F, Niu Z, Person R, Ding Y, Ward P, Braxton A, Wang M, Buhay C, Veeraraghavan N, Hawes A, Chiang T, Leduc M, Beuten J, Zhang J, He W, Scull J, Willis A, Landsverk M, Craigen WJ, Bekheirnia MR, Stray-Pedersen A, Liu P, Wen S, Alcaraz W, Cui H, Walkiewicz M, Reid J, Bainbridge M, Patel A, Boerwinkle E, Beaudet AL, Lupski JR, Plon SE, Gibbs RA, Eng CM. Molecular findings among patients referred for clinical whole-exome sequencing. JAMA 2014;312:1870–9. 10.1001/jama.2014.14601 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Gillespie RL, O'Sullivan J, Ashworth J, Bhaskar S, Williams S, Biswas S, Kehdi E, Ramsden SC, Clayton-Smith J, Black GC, Lloyd IC. Personalized diagnosis and management of congenital cataract by next-generation sequencing. Ophthalmology 2014;121:2124–37.e2. 10.1016/j.ophtha.2014.06.006 [DOI] [PubMed] [Google Scholar]
- 7.Lee H, Deignan JL, Dorrani N, Strom SP, Kantarci S, Quintero-Rivera F, Das K, Toy T, Harry B, Yourshaw M, Fox M, Fogel BL, Martinez-Agosto JA, Wong DA, Chang VY, Shieh PB, Palmer CGS, Dipple KM, Grody WW, Vilain E, Nelson SF. Clinical Exome Sequencing for Genetic Identification of Rare Mendelian Disorders. JAMA 2014;312:1880–7. 10.1001/jama.2014.14604 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Lee K, Berg JS, Milko L, Crooks K, Lu M, Bizon C, Owen P, Wilhelmsen KC, Weck KE, Evans JP, Garg S. high diagnostic yield of whole exome sequencing in participants with retinal dystrophies in a clinical ophthalmology setting. Am J Ophthalmol 2015;160:354–63. 10.1016/j.ajo.2015.04.026 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Wang H, Wang X, Zou X, Xu S, Li H, Soens ZT, Wang KQ, Li YM, Dong FT, Chen R, Sui RF. comprehensive molecular diagnosis of a large Chinese Leber congenital amaurosis cohort. Invest Ophthalmol Vis Sci 2015;56:3642–55. 10.1167/iovs.14-15972 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.O'Sullivan J, Mullaney BG, Bhaskar SS, Dickerson JE, Hall G, O'Grady A, Webster A, Ramsden SC, Black GC. A paradigm shift in the delivery of services for diagnosis of inherited retinal disease. J Med Genet 2012;49:322–6. 10.1136/jmedgenet-2012-100847 [DOI] [PubMed] [Google Scholar]
- 11.Li H, Durbin R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009;25:1754–60. 10.1093/bioinformatics/btp324 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.DePristo MA, Banks E, Poplin R, Garimella KV, Maguire JR, Hartl C, Philippakis AA, del Angel G, Rivas MA, Hanna M, McKenna A, Fennell TJ, Kernytsky AM, Sivachenko AY, Cibulskis K, Gabriel SB, Altshuler D, Daly MJ. A framework for variation discovery and genotyping using next-generation DNA sequencing data. Nat Genet 2011;43:491–8. 10.1038/ng.806 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.McKenna A, Hanna M, Banks E, Sivachenko A, Cibulskis K, Kernytsky A, Garimella K, Altshuler D, Gabriel S, Daly M, DePristo MA. The Genome Analysis Toolkit: a MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res 2010;20:1297–303. 10.1101/gr.107524.110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Ellingford JM, Barton S, Bhaskar S, Williams SG, Sergouniotis PI, O'Sullivan J, Lamb JA, Perveen R, Hall G, Newman WG, Bishop PN, Roberts SA, Leach R, Tearle R, Bayliss S, Ramsden SC, Nemeth AH, Black GCM. Whole genome sequencing increases molecular diagnostic yield compared with current diagnostic testing for inherited retinal disease. Ophthalmology 2016;123:1143–50. 10.1016/j.ophtha.2016.01.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kumar P, Henikoff S, Ng PC. Predicting the effects of coding non-synonymous variants on protein function using the SIFT algorithm. Nat Protoc 2009;4:1073–82. 10.1038/nprot.2009.86 [DOI] [PubMed] [Google Scholar]
- 16.Adzhubei IA, Schmidt S, Peshkin L, Ramensky VE, Gerasimova A, Bork P, Kondrashov AS, Sunyaev SR. A method and server for predicting damaging missense mutations. Nat Methods 2010;7:248–9. 10.1038/nmeth0410-248 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Richards S, Aziz N, Bale S, Bick D, Das S, Gastier-Foster J, Grody WW, Hegde M, Lyon E, Spector E, Voelkerding K, Rehm HL. Standards and guidelines for the interpretation of sequence variants: a joint consensus recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genet Med 2015;17:405–23. 10.1038/gim.2015.30 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Stenson PD, Mort M, Ball EV, Shaw K, Phillips AD, Cooper DN. The Human Gene Mutation Database: building a comprehensive mutation repository for clinical and molecular genetics, diagnostic testing and personalized genomic medicine. Hum Genet 2014;133:1–9. 10.1007/s00439-013-1358-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Perrault I, Rozet JM, Calvas P, Gerber S, Camuzat A, Dollfus H, Chatelin S, Souied E, Ghazi I, Leowski C, Bonnemaison M, LePaslier D, Frezal J, Dufier JL, Pittler S, Munnich A, Kaplan J. Retinal-specific guanylate cyclase gene mutations in Leber's congenital amaurosis. Nature Genet 1996;14:461–4. 10.1038/ng1296-461 [DOI] [PubMed] [Google Scholar]
- 20.Berger W, Kloeckener-Gruissem B, Neidhardt J. The molecular basis of human retinal and vitreoretinal diseases. Prog Retin Eye Res 2010;29:335–75. 10.1016/j.preteyeres.2010.03.004 [DOI] [PubMed] [Google Scholar]
- 21.Eisenberger T, Neuhaus C, Khan AO, Decker C, Preising MN, Friedburg C, Bieg A, Gliem M, Issa PC, Holz FG, Baig SM, Hellenbroich Y, Galvez A, Platzer K, Wollnik B, Laddach N, Ghaffari SR, Rafati M, Botzenhart E, Tinschert S, Borger D, Bohring A, Schreml J, Kortge-Jung S, Schell-Apacik C, Bakur K, Al-Aama JY, Neuhann T, Herkenrath P, Nurnberg G, Nurnburg P, Davis JS, Gal A, Bergmann C, Lorenz B, Bolz HJ. Increasing the yield in targeted next-generation sequencing by implicating CNV analysis, non-coding exons and the overall variant load: the example of retinal dystrophies. PLoS ONE 2013;8:e78496 10.1371/journal.pone.0078496 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Zhao L, Wang F, Wang H, Li YM, Alexander S, Wang KQ, Willoughby CE, Zaneveld JE, Jiang LC, Soens ZT, Earle P, Simpson D, Silvestri G, Chen R. Next-generation sequencing-based molecular diagnosis of 82 retinitis pigmentosa probands from Northern Ireland. Hum Genet 2015;134:217–30. 10.1007/s00439-014-1512-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Abdulwahab F, Abouelhoda M, Abouthuraya R, Imam A, Ahmed SO, Al Rubeaan K, Al Tassan N, AlAbdulaziz B, AlAbdulrahman K, Alamer FH, Alazami A, Al-Baik LA, Aldahmesh M, Al-Dhekri H, AlDusery H, Algazlan S, Al-Ghonaium A, Alhamed M, Alhashem A, Alhissi SA, AlIssa A, Aljurf MD, Alkuraya FS, Alkuraya H, Allam R, Almasharawi IJ, Almoisheer A, AlMostafa A, Al-Mousa H, Al-Muhsen S, Almutairy EA, Alnader N, AlNaqeb D, AlOtaibi AB, Alotibi A, Al-Qattan S, Al-Saud B, Al-Saud H, Alshammari M, Alsheikh H, Aisheikh AH, Al-Sulaiman A, Altamimi AS, Al-Tayeb H, Alwadaee SM, Al-Younes B, Alzahrani F, Anazi S, Arnaout R, Fahad B, Yousef B, Binhumaid FS, Boholega S, Broering D, Burdelski M, Dasouki MJ, Dzimiri NF, Elamin T, El-Kalioby M, Elsiesy H, Faqeih E, Faquih T, Hagos S, Hagr AA, Hashem M, Hawwari A, Hazzaa S, Ibrahim N, Imtiaz F, Jabr A, Kattan R, Kaya N, Kentab A, Khalil D, Khan AO, Khier O, Meyer B, Mohamed J, Monies D, Muiya PN, Murad H, Naim EA, Owaidah T, Patel N, Ramzan K, Salih MA, Shagrani M, Shaheen R, Shamseldin H, Sameera S, Shazia T, Khalid W, Salma M, Saudi Mendeliome Group. Comprehensive gene panels provide advantages over clinical exome sequencing for Mendelian diseases. Genome Biol 2015;16:134 10.1186/s13059-015-0581-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Akawi N, McRae J, Ansari M, Balasubramanian M, Blyth M, Brady AF, Clayton S, Cole T, Deshpande C, Fitzgerald TW, Foulds N, Francis R, Gabriel G, Gerety SS, Goodship J, Hobson E, Jones WD, Joss S, King D, Klena N, Kumar A, Lees M, Lelliott C, Lord J, McMullan D, O'Regan M, Osio D, Piombo V, Prigmore E, Rajan D, Rosser E, Sifrim A, Smith A, Swaminathan GJ, Turnpenny P, Whitworth J, Wright CF, Firth HV, Barrett JC, Lo CW, FitzPatrick DR, Hurles ME, Study DDD. Discovery of four recessive developmental disorders using probabilistic genotype and phenotype matching among 4,125 families. Nature Genet 2015;47:1363 10.1038/ng.3410 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Smedley D, Jacobsen JOB, Jäger M, Köhler S, Holtgrewe M, Schubach M, Siragusa E, Zemojtel T, Buske OJ, Washington NL, Bone WP, Haendel MA, Robinson PN. Next-generation diagnostics and disease-gene discovery with the Exomiser. Nat Protoc 2015;10:2004–15. 10.1038/nprot.2015.124 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Ellingford JM, Sergouniotis PI, Lennon R, Bhaskar S, Williams SG, Hillman KA, O'Sullivan J, Hall G, Ramsden SC, Lloyd IC, Woolf AS, Black GC. Pinpointing clinical diagnosis through whole exome sequencing to direct patient care: a case of Senior-Loken syndrome. Lancet 2015;385:1916 10.1016/S0140-6736(15)60496-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ruddle JB, Ebenezer ND, Kearns LS, Mulhall LE, Mackey DA, Hardcastle AJ. RPGR ORF15 genotype and clinical variability of retinal degeneration in an Australian population. Br J Ophthalmol 2009;93:1151–4. 10.1136/bjo.2008.153908 [DOI] [PubMed] [Google Scholar]
- 28.Bader I, Brandau O, Achatz H, Apfelstedt-Sylla E, Hergersberg M, Lorenz B, Wissinger B, Wittwer B, Rudolph G, Meindl A, Meitinger T. X-linked retinitis pigmentosa: RPGR mutations in most families with definite X linkage and clustering of mutations in a short sequence stretch of exon ORF15. Invest Ophthalmol Vis Sci 2003;44:1458–63. 10.1167/iovs.02-0605 [DOI] [PubMed] [Google Scholar]
- 29.Ye K, Schulz MH, Long Q, Apweiler R, Ning Z. Pindel: a pattern growth approach to detect break points of large deletions and medium sized insertions from paired-end short reads. Bioinformatics 2009;25:2865–71. 10.1093/bioinformatics/btp394 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Plagnol V, Curtis J, Epstein M, Mok KY, Stebbings E, Grigoriadou S, Wood NW, Hambleton S, Burns SO, Thrasher AJ, Kumararatne D, Doffinger R, Nejentsev S. A robust model for read count data in exome sequencing experiments and implications for copy number variant calling. Bioinformatics 2012;28: 2747–54. 10.1093/bioinformatics/bts526 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Mayer AK, Rohrschneider K, Strom TM, Glockle N, Kohl S, Wissinger B, Weisschuh N. Homozygosity mapping and whole-genome sequencing reveals a deep intronic PROM1 mutation causing cone-rod dystrophy by pseudoexon activation. Eur J Hum Genet 2015;24:459–62. 10.1038/ejhg.2015.144 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Small KW, DeLuca AP, Whitmore SS, Rosenberg T, Silva-Garcia R, Udar N, Puech B, Garcia CA, Rice TA, Fishman GA, Heon E, Folk JC, Streb LM, Haas CM, Wiley LA, Scheetz TE, Fingert JH, Mullins RF, Tucker BA, Stone EM. North Carolina Macular Dystrophy Is Caused by Dysregulation of the Retinal Transcription Factor PRDM13. Ophthalmology 2016;123:9–18. 10.1016/j.ophtha.2015.10.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.de Castro-Miro M, Pomares E, Lores-Motta L, Tonda R, Dopazo J, Marfany G, Gonzalez-Duarte R. Combined genetic and high-throughput strategies for molecular diagnosis of inherited retinal dystrophies. PLoS ONE 2014;9:e88410 10.1371/journal.pone.0088410 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Krawitz PM, Schiska D, Krüger U, Appelt S, Heinrich V, Parkhomchuk D, Timmermann B, Millan JM, Robinson PN, Mundlos S, Hecht J, Gross M. Screening for single nucleotide variants, small indels and exon deletions with a next-generation sequencing based gene panel approach for Usher syndrome. Mol Genet Genomic Med 2014;2:393–401. 10.1002/mgg3.92 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Sullivan LS, Bowne SJ, Reeves MJ, Blain D, Goetz K, Ndifor V, Vitez S, Wang XJ, Tumminia SJ, Daiger SP. Prevalence of mutations in eyeGENE Probands with a diagnosis of autosomal dominant retinitis pigmentosa. Invest Ophthalmol Vis Sci 2013;54:6255–61. 10.1167/iovs.13-12605 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
jmedgenet-2016-103837supp_tables.pdf (3.9MB, pdf)