

Abstract
Background
Climate change is predicted to be a serious threat to agriculture due to the need for crops to be able to tolerate increased heat stress. Desert plants have already adapted to high levels of heat stress so they make excellent systems for identifying genes involved in thermotolerance. Rhazya stricta is an evergreen shrub that is native to extremely hot regions across Western and South Asia, making it an excellent system for examining plant responses to heat stress. Transcriptomes of apical and mature leaves of R. stricta were analyzed at different temperatures during several time points of the day to detect heat response mechanisms that might confer thermotolerance and protection of the plant photosynthetic apparatus.
Results
Biological pathways that were crosstalking during the day involved the biosynthesis of several heat stress-related compounds, including soluble sugars, polyols, secondary metabolites, phenolics and methionine. Highly downregulated leaf transcripts at the hottest time of the day (40–42.4 °C) included genes encoding cyclin, cytochrome p450/secologanin synthase and U-box containing proteins, while upregulated, abundant transcripts included genes encoding heat shock proteins (HSPs), chaperones, UDP-glycosyltransferase, aquaporins and protein transparent testa 12. The upregulation of transcripts encoding HSPs, chaperones and UDP-glucosyltransferase and downregulation of transcripts encoding U-box containing proteins likely contributed to thermotolerance in R. stricta leaf by correcting protein folding and preventing protein degradation. Transcription factors that may regulate expression of genes encoding HSPs and chaperones under heat stress included HSFA2 to 4, AP2-EREBP and WRKY27.
Conclusion
This study contributed new insights into the regulatory mechanisms of thermotolerance in the wild plant species R. stricta, an arid land, perennial evergreen shrub common in the Arabian Peninsula and Indian subcontinent. Enzymes from several pathways are interacting in the biosynthesis of soluble sugars, polyols, secondary metabolites, phenolics and methionine and are the primary contributors to thermotolerance in this species.
Electronic supplementary material
The online version of this article (doi:10.1186/s12870-016-0938-6) contains supplementary material, which is available to authorized users.
Keywords: Thermotolerance, HSP, Chaperones, HSF, Cyclin, U-box, Aquaporine, Protein transparent testa 12, AP2-EREBP, WRKY27
Background
Predicted climate changes are likely to represent serious threats to agriculture and food safety [1–3]. By the end of this century maximum temperature is expected to increase by more than 2 °C depending on industrial emmissions [4]. Heat stress poses one of the greatest detrimental effects on the growth and productivity of crop plants [5]. These effects include some physiological alterations in the leaf such as low photosynthetic rate and changes in metabolite accumulation [6–8]. The overall effect of the combined stress is altered protein homeostasis [9], including the control of protein synthesis, intracellular sorting, folding and degradation [10].
Tolerance to heat stress is a multigenic process with many regulatory mechanisms acting during plant development [11, 12]. Heat stress injury and response are more evident in plant leaves [1] and pollen [13] during sexual reproduction than other tissues. Plants respond to heat stress by synthesizing heat shock proteins [14] (HSPs). Transcript abundance of HSPs along with chaperones has been shown to be involved in heat stress tolerance [15, 16]. Heat shock proteins are considered molecular chaperones (e.g., HSP90, HSP70 and HSP60) that control stability and folding of other proteins to protect misfolded proteins from irreversible aggregation [17–20].
In general, plant cells tolerate heat stress by orchestrating energy metabolism between dissimilation and assimilation [21], by scavenging antioxidant enzymes [22] and by reducting detoxification of reactive oxygen species (ROS) responsible for the peroxidation of membrane lipids and pigments, which causes loss of membrane permeability [23, 24]. The latter action requires high levels of expression of antioxidant genes to help confer heat tolerance in plants.
Omics has been used extensively to provide valuable information for breeding programs to improve plant thermotolerance. In recent reports, ~5% of plant transcripts were highly upregulated due to heat stress [25–27]. Upregulated transcripts include those encoding chaperones [7, 25], while others are involved in calcium/phytohormone/lipid signaling, phosphorylation, sugar accumulation, secondary metabolism and many other biological processes [28].
Transcription factors (TFs) represent key proteins required for the regulation of almost all biosynthetic pathways in life [29]. They are important for the development of organisms and for all cellular functions and responses to biotic and abiotic stresses [30]. In a previous study [31], a number of important TF families were identified in the perennial evergreen C3 desert shrub Rhazya stricta by Mapman analysis. This shrub grows well in its arid environment under high temperatures and vapor pressure deficits. The expression of gene families encoding the basic helix-loop-helix (bHLH), homeobox domain (HB), MYB as well as AP2-ERF significantly decreased at midday [31].
In the present study, we extended previous efforts of Yates et al. [31] by studying the transcriptomes of the apical and mature leaves of Rhazya stricta at differenent day time temperatures to gain new insights into heat response mechanisms that are involved in thermotolerance and protection of the plant photosynthetic apparatus. Such mechanisms might be a target for improving thermotolerance of economically important crop plants via transgenesis.
Results and discussion
Clusters of gene expression at different temperatures across times of the day
RNA-Seq analysis was used to analyse apical and mature leaves to test if heat responsive genes are expressed similarly in the two different leaf types. These two types of leaves differ in their developmental stages and status of cell division, which might affect heat-responsive genes differently. We speculated that this plant organ would provide a wealth of information in terms of the responsive gene families and biological pathways under heat stress. Temperatures (40–42.4 °C) at the three midday time points (13:25, 14:05, 14:30) were 12.6–15 °C higher than the morning time point (07:10) temperatures (27.4 °C), confirming that Rhazya stricta was experiencing heat stress during midday time points as compared to the morning. We speculate that more accurate results will be gained when comparing transcriptomes of the same plant across different time points, e.g., dawn (non-stressed) vs midday (stressed), rather than comparing transcriptomes of stressed vs. non-stressed plants at a given time point, e.g., midday. Furthermore, it is difficult to control environmental conditions for plants growing in the field. Hierarchical cluster analysis of gene expression based on log ratio RPKM data for transcripts of R. stricta SRA database in the apical and mature leaves at different time points of the day indicated the high quality of sampling and RNA-Seq analysis as evidenced by within timepoint clustering of replicates in 37 of the 40 samples (Fig. 1). Similar conclusions were reached when studying the genes with different expression patterns in the apical and mature leaves (Additional file 1: Table S1 & Additional file 2: Table S2 and Additional file 3: Figure S1 & Additional file 4: Figure S2, respectively). The only non-concordant samples (Fig. 1, red arrows) were the apical leaf samples F2, G1 and L3; F2 clustered with the apical leaf samples at dusk (L), G1 with apical leaf samples at time point F at midday and L3 with mature leaf samples at dusk. In general, the sampling of mature leaves resulted in more homogenous data than the apical leaves. The number of DE transcripts resulting from the RNA-Seq analysis of apical leaves across different time points was 2507 in 32 clusters (Additional file 1: Table S1). The number of DE transcripts across time points in mature leaves was 4853 in 38 clusters (Additional file 2: Table S2). We can infer that a key reason for the larger number of genes enriched in the mature leaves across the day compared to apical meristimic leaves is that the latter is more active in cell division and cell differentiation [32]. Clusters with up or downregulation starting at midday that were utilized frequently for both leaf types are shown in Fig. 2.
Fig. 1.
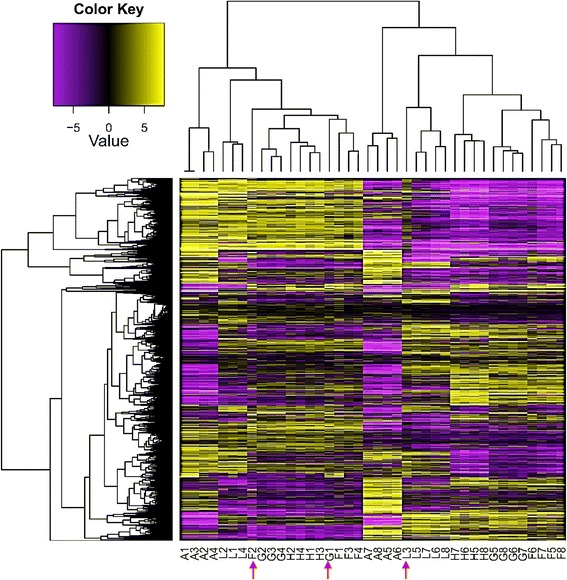
Hierarchical cluster analysis of gene expression based on log ratio RPKM data for transcriptome of R. stricta SRA database in the apical (A1-L4) and mature leaves (A5-L8) at different time points of the day (A, morning; F-H, midday & L, dusk). Red arrows indicate the misplaced samples in the cluster analysis
Fig. 2.
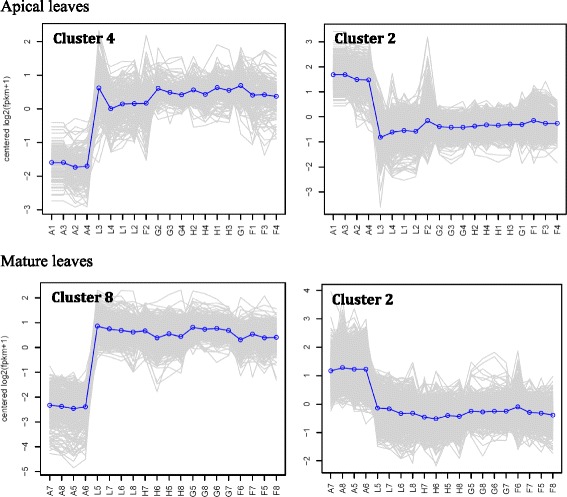
Selected clusters of up or downregulated genes of R. stricta from apical (A1-L4) and mature (A5-L8) leaves at different time points of the day (A, morning; F-H, midday and L, dusk). Clusters 4 and 2 of apical leaves = up and downregulation starting midday, respectively. Clusters 8 and 2 of mature leaves = up and downregulation starting midday, respectively. Blue lines indicate overall expression pattern across different transcripts of a given cluster
Semi-quantitative RT-PCR of 10 randomly selected genes was used to validate the RNA-Seq data with three replicates of both types of leaves across the three time points, e.g., morning (A), midday (F-H) and dusk (L) (Additional file 5: Figure S3). Expression patterns of these 10 genes included upregulation starting at midday and gradual downregulation (Additional file 6: Table S3). The results of semi-quantitative RT-PCR for the selected genes confirmed the fold change in the RNA-Seq data across the two types of leaves and three time points.
Analysis of differentially expressed genes
KEGG analysis
To identify the biological pathways that are active in the apical and mature leaves of R. stricta during the day, we mapped the detected genes to reference canonical pathways in the Kyoto Encyclopedia of Genes and Genomes (KEGG) (http://www.genome.ad.jp/kegg/). Heat tolerance is a multigenic process with different metabolic pathways affecting plant growth [12]. Enzymes with roles in the pathways that showed regulation during the day under heat stress were examined in apical and mature leaves (Table 1 and Figs. 3 and 4 and Additional file 7: Figure S4, Additional file 8: Figure S5, Additional file 9: Figure S6, Additional file 10: Figure S7 and Additional file 11: Figure S8).
Table 1.
Description of the differentially responding enzymes in apical and mature leaves to changing environments at two time points (e.g., A, morning and G, midday). Activated (blue), repressed (orange)
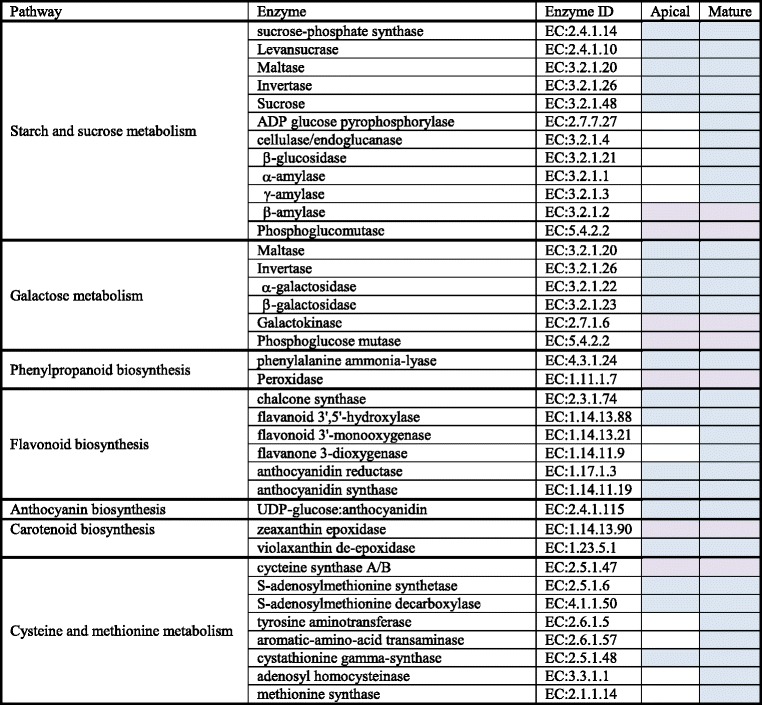
Fig. 3.
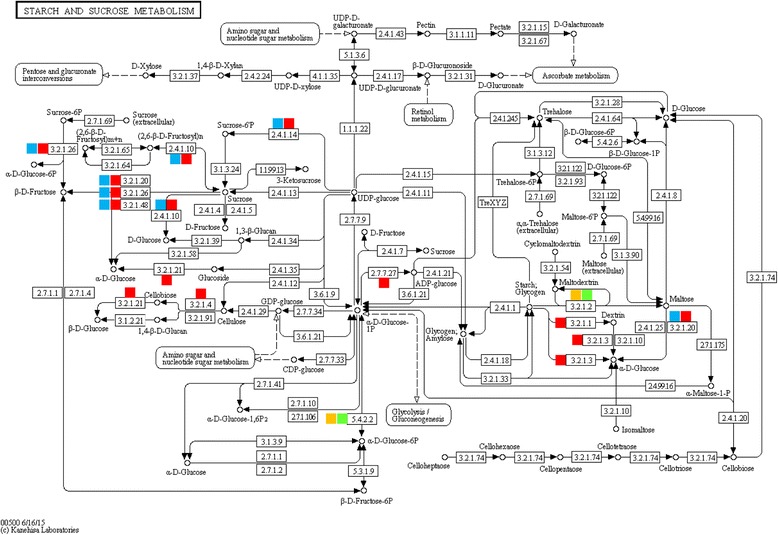
Enzymes in the starch and sucrose metabolic pathway in apical and mature leaves responded differentially to changing environment at two time points (morning, A and midday. G). Upregulated (activated) in apical leaves (blue), upregulated in mature leaves (red), downregulated (repressed) in apical leaves (orange box), downregulated in mature leaves (green box)
Fig. 4.
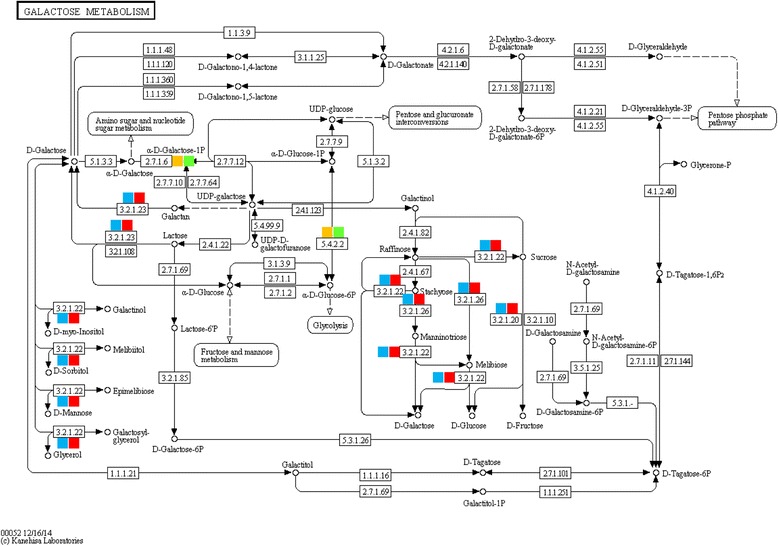
Enzymes in the galactose metabolic pathway in apical and mature leaves responded differentially to changing environments at two time points (morning, A and midday, G). Upregulated (activated) in apical leaves (blue), upregulated in mature leaves (red), downregulated (repressed) in apical leaves (orange box), downregulated in mature leaves (green box)
In general, KEGG analysis indicated that the biosynthesis of soluble sugars, polyols, secondary metabolites, phenolics and methionine are involved in conferring thermotolerance in R. stricta. The results of starch and sucrose metabolism pathway indicated the involvement of 12 enzymes in the response to heat stress (Table 1 and Fig. 3). The most evident responses are the synthesis of several soluble sugars, e.g., sucrose, fructose and glucose, and the depletion of starch and maltose mostly by the action of sucrose-phosphate synthase, levansucrase, maltase, sucrase and invertase. Earlier reports on sucrose phosphate synthase and invertase in mulberry and soybean documented their repression under heat stress [33, 34]. Hence, soluble sugars were not accumulated as a response to heat stress during the day in these two plant species. Depletion of starch in R. stricta during the day towards the production of soluble sugars can be considered a favorable action only at night. Therefore, the activation of ADP glucose pyrophosphorylase (AGPase) under heat stress during the day in leaves of R. stricta leads to the synthesis of glycogen, which provides a continuous supply of starch during the day. This transition maximizes cytosolic carbon-sink strength in the cell [35]. Krasensky and Jonak [36] also indicated an active role of AGPase and other enzymes in starch production in the plastid during photosynthesis. It is unlikely that glycogen is converted to maltose during the day in leaves of R. stricta as the enzyme responsible for this action, β-amylase [37, 38], was repressed. Therefore, we can conclude that both soluble sugars and starch are favorably accumulated in R. stricta during the day.
Lawson et al. [39] found evidence for thermotolerance while studying photosynthetic capacity in R. stricta at the same time period and under the same field conditions as our study. The evidence involved the occurrence of a maximum in vivo carboxylation capacity of the thermostable Rubisco [40] (up to 50 °C). The recorded temperature during Lawson’s and our experiments was 43 °C. Salvucci and Crafts-Brandner [41] indicated that the thermal instability of the two Rubisco activase (RCA) isoforms at such high temperatures is a major limitation to photosynthetic capacity. The enzyme plays an important regulatory role in photosynthesis as it catalyses the removal of the sugar phosphates from the Rubisco catalytic sites [42]. Sugar phosphates are known for their action in retarding photosynthesis as they bind to Rubisco and prevent the carbamylation process [42]. The results of the present study support the results of Lawson et al. [39] because we detected a gene encoding rubisco subunit binding-protein alpha that was upregulated in the two leaf types during midday (Fig. 5). The encoded protein binds Rubisco small and large subunits and is implicated in the assembly of the enzyme oligomer. Upregulation of this gene during midday secures the continuous supply of the thermostable Rubisco during photosynthesis. In addition, the two RCA forms (RCA1 and RCA2), which represent the weak link to appropriate photosynthetic capacity under heat stress, were detected in the mature leaf of R. stricta, while only one form was detected in the apical leaf. These enzyme isoforms were downregulated in the present study only at dusk (Fig. 6). The continuous expression of the two RCA genes during the day secures the biosynthesis of the enzyme isoforms under heat stress, thus promoting photosynthesis. These results add to the understading of the mechanisms of thermotolerance in R. stricta.
Fig. 5.
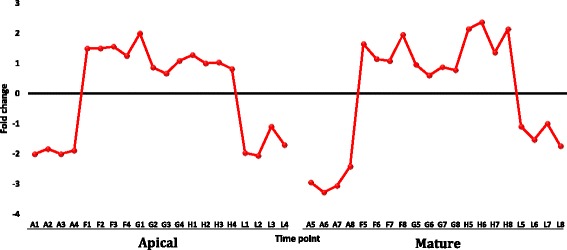
Fold change values for the gene encoding Rubisco in apical (A1-L4) and mature (A5-L8) leaves during the day (A, morning; F-H, midday & L, dusk) in R. stricta
Fig. 6.
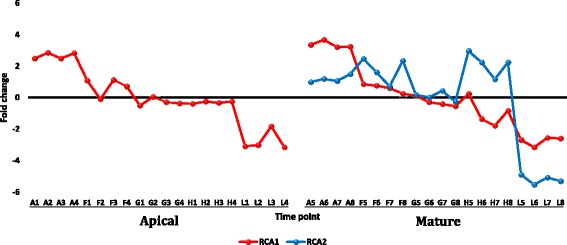
Fold change values for the genes encoding the two Rubisco activase isoforms (RCA1 and RCA2) in apical (A1-L4) and mature (A5-L8) leaves during the day (A, morning; F-H, midday & L, dusk) in R. stricta
Results of the enzyme activity in the galactose metabolism pathway under heat stress in leaves of R. stricta support the accumulation of soluble sugars (e.g., sucrose, glucose and galactose), as well as in the synthesis of several polyols (e.g., myo-inositol, sorbitol, mannose, glycerol) due to the activity of α-galactosidase (or melibiase) (Table 1 and Fig. 4). The analogue of this enzyme, i.e., β-galactosidase (or lactase), is involved in the synthesis of galactose via the conversion of galactan and lactose. Starch and glycogen are known for their sensitivity to changing environments [43–46]. The metabolism of either compound is important for the storage of carbon and energy in the cell [47]. Activities of enzymes involved in starch and sucrose metabolism and galactose metabolism during the day in leaves of R. stricta resulted in the accumulation of soluble sugars that can act as osmolytes to maintain cell turgor and protect membranes and proteins from damage caused by different abiotic stresses. Polyols are compatible solutes with the ability to stabilize proteins and scavenge hydroxyl radicals towards the prevention of oxidative damage of membranes and enzymes under abiotic stresses, including heat stress [48]. In agreement with our results, many reports indicated that stress tolerant plants accumulate larger amounts of protective metabolites, such as soluble sugars and polyols, under adverse conditions [40, 44]. Rosa et al. [49] also found that sucrose and hexoses upregulate growth-related genes, while downregulating stress-related genes. This dual response likely assures proper growth under unfavorable conditions in R. stricta.
Many secondary metabolites are synthesized from the intermediates of primary carbon metabolism [50]. Important enzymes in the phenylpropanoid metabolic pathway crosstalk with many downstream secondary metabolite pathways such as flavonoid and anthocyanin biosynthesis. It is well known that high temperature stress induces the production of phenolic compounds such as flavonoids and phenylpropanoids for thermotolerance [9]. The key enzyme in the phenylpropanoid metabolism pathway, phenylalanine ammonia-lyase (PAL), was activated during the day in leaves of R. stricta (Table 1 and Additional file 7: Figure S4). Activity of PAL in response to heat stress was reported earlier as the main acclimatory response [9] where the enzyme induces the biosynthesis of other phenolics in the pathway. Phenolics, including flavonoids and anthocyanins, were reported earlier as the key secondary metabolites in abiotic stress tolerance [50, 51]. In contrast, peroxidase enzyme was repressed in the phenylpropanoid metabolism pathway indicating the suppression of oxidation of phenolics in apical and mature leaves of R. stricta during the day. This action can help reduce detoxification of ROS to maintain cell membrane permeability [24].
The enzyme chalcone synthase, the first enzyme in flavonoid biosynthesis pathway, was activated during the day in leaves of R. stricta (Table 1 and Additional file 8: Figure S5). This enzyme is also important in the orchestration of several other pathways, including flavone and flavonol biosynthesis and anthocyanin biosynthesis. Three other enzymes in the flavonoid biosynthesis pathway involved in the synthesis of several important intermediate flavonoids, flavonoid 3′,5′-hydroxylase, flavonoid 3′-monooxygenase and naringenin 3-dioxygenase, were also activated in leaves of R. stricta under heat stress. Two other key enzymes, leucocyanidin reductase (LAR) and leucocyanidin oxygenase, were activated in leaves of R. stricta towards the production of important phenolics. The first enzyme acts in the formation of proanthocyanidins (PAs), polymers of flavan-3-ol subunits, while the action of the second enzyme is linked directly through many avenues to the anthocyanin biosynthesis pathway (Table 1 and Additional file 9: Figure S6). Earlier reports in grape indicated that increased temperature enhances the production of PAs [52], which act in protecting plants against herbivores and UV radiation during the day [53]. The KEGG analysis in the anthocyanin biosynthesis pathway indicated the activation of only one enzyme, UDP-glucose:anthocyanidin (Table 1 and Additional file 9: Figure S6). This key enzyme catalyzes the first step of the pathway towards the eventual synthesis of many anthocyanins in the cell.
Two light-responsive enzymes in the carotenoid biosynthesis pathway were also regulated in leaves of R. stricta (Table 1 and Additional file 10: Figure S7). The first, zeaxanthin epoxidase, was repressed under heat stress, while the second, violaxanthin de-epoxidase, was activated. The two enzymes act as a shuttle for the reversible interconversion of the two carotenoids zeaxanthin and violaxanthin and their activities are light regulated [9]. It is evident that zeaxanthin biosynthesis was enhanced, while violaxanthin biosynthesis was repressed. Zeaxanthin is known for its role in photoprotection in the cells as it also acts to prevent peroxidative damage to the membrane lipids triggered by ROS under abiotic stresses [24, 54].
The pathway of cysteine and methionine metabolism is regulated in mature leaf cells of R. stricta under heat stress towards the oversynthesis of methionine (Table 1 and Additional file 11: Figure S8) due to the activation of three enzymes, methionine synthase, tyrosine aminotransferase and aromatic-amino-acid transaminase. Two other enzymes, S-adenosylmethionine synthetase and S-adenosylmethionine decarboxylase, were also activated in both apical and mature leaves towards the depletion of methionine. However, this can be compensated for in mature leaves by the action of the three enzymes indicated earlier for oversynthesis of methionine. Cysteine seems negatively regulated in both apical and mature leaves due to the possible repression of cycteine synthase A/B enzyme in the cell under heat stress. Methionine is a major amino acid in chloroplast small heat shock proteins (sHSPs), which act in plant adaptation to severe heat stress by protecting the process of photosystem II electron transport [55]. Gustavsson et al. [56] also reported that methionine residues in HSP21 mediate protein repair under heat stress.
Regulated gene families under heat stress with ≥ 5 fold change
Transcripts selected from the datasets of apical and mature leaves of R. stricta that showed down or upregulation with fold change (FC) of ≥ 5 are shown in Additional file 12: Table S4. Analysis was selectively done for gene families whose members were frequently up or down regulated in leaves of R. stricta or those with prior information on their response to heat stress. The selected highly downregulated transcripts at highest midday temperatures in leaves of R. stricta included genes encoding cyclin, cytochrome p450/secologanin synthase and U-box containing proteins (Additional file 12: Table S4 and Figs. 7, 8 and 9, respectively). Upregulated, abundant transcripts included genes encoding HSPs/chaperones, UDP-glycosyltransferase, aquaporins and protein transparent testa 12 (Additional file 12: Table S4 and Figs. 10, 11, 12 and 13, respectively). Some upregulated transcripts showed extreme downregulation at dusk, while none of them showed downregulation at midday with no differential regulation among the three time points of the midday (e.g., F, G & H).
Fig. 7.
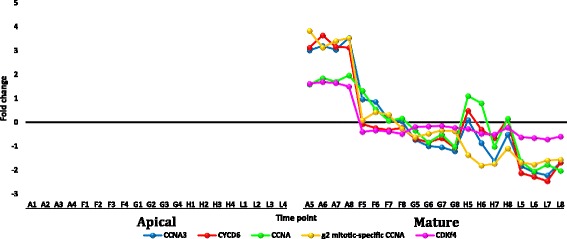
Fold change values for the downregulated genes encoding cyclin in apical (A1-L4) and mature (A5-L8) leaves during the day (A, morning; F-H, midday & L, dusk) in R. stricta
Fig. 8.
Fold change values for the downregulated genes encoding cytochrome P-450 in apical (A1-L4) and mature (A5-L8) leaves during the day (A, morning; F-H, midday & L, dusk) in R. stricta
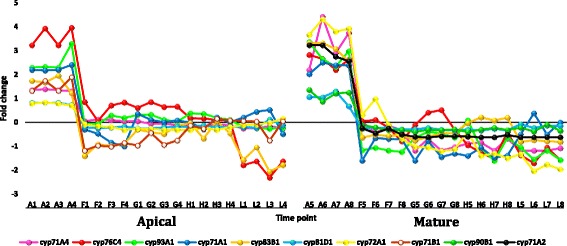
Fig. 9.
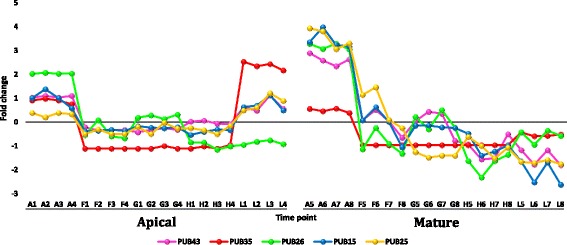
Fold change values for the downregulated genes encoding U-box containing proteins in apical (A1-L4) and mature (A5-L8) leaves during the day (A, morning; F-H, midday & L, dusk) in R. stricta
Fig. 10.
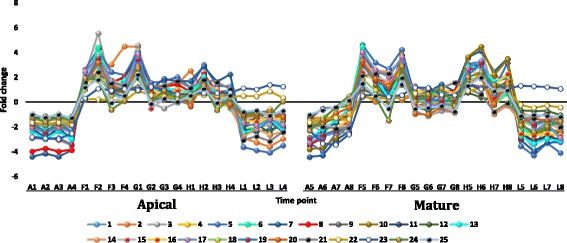
Fold change values for the upregulated genes encoding HSPs in apical (A1-L4) and mature (A5-L8) leaves during the day (A, morning; F-H, midday & L, dusk) in R. stricta. Numbers refer to those in Additional file 13: Table S5
Fig. 11.
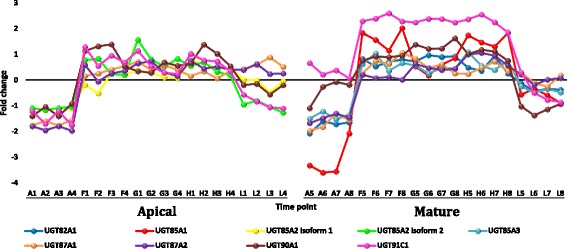
Fold change values for the upregulated genes encoding UDP-glycosyltransferase in apical (A1-L4) and mature (A5-L8) leaves during the day (A, morning; F-H, midday & L, dusk) in R. stricta
Fig. 12.
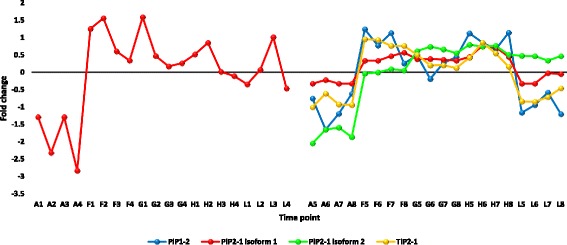
Fold change values for the upregulated genes encoding aquaporins (AQPs) or major intrinsic proteins (MIPs) in apical (A1-L4) and mature (A5-L8) leaves during the day (A, morning; F-H, midday & L, dusk) in R. stricta
Fig. 13.
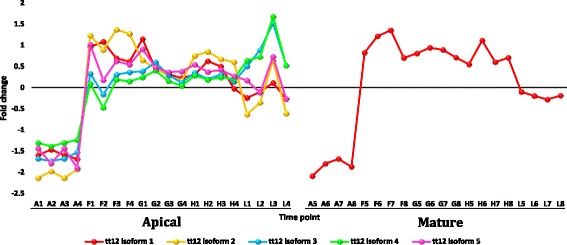
Fold change values for the upregulated genes encoding protein transparent testa 12 (tt12) in apical (A1-L4) and mature (A5-L8) leaves during the day (A, morning; F-H, midday & L, dusk) in R. stricta
Transcripts encoding cyclin proteins A (CCNA), A3 (CCNA3) and D6 (CYCD6) were downregulated only in mature leaves of R. stricta (Additional file 12: Table S4). The plant cyclin gene family has 10 types (A, B, C, D, H, L, T, U, SDS and J18; Zhang et al. [57]). The A and D types are involved in regulation of cell division during phases S to M and G1 to S, respectively [58]. Thus, it is likely that mature leaf cells of R. stricta were arrested at G1-S phases due to heat stress. In agreement with these findings, transcripts encoding cyclin-dependent kinase (CDK) class f4-like, a regulator of cell cycle progression through the binding to cyclin, were also highly downregulated at midday only in mature leaves, while upregulated in apical leaves of R. stricta (Additional file 12: Table S4). This should result in prompt inhibition of cell division in mature leaves only, which may be a mechanism of tolerance by avoiding or escaping heat stress. Based on these results, the stress avoidance mechanism is not likely applicable to apical leaves of R. stricta whose major process is cell division. Recent studies indicated the indirect role of CDKs in plant tolerance to heat stress via a sophisticated mechanism of stress avoidance [59].
Highly downregulated transcripts encoding cytochrome P450 (cyt P450) in response to heat stress were identified in leaves of R. stricta (Additional file 12: Table S4). This involved 10 genes belonging to seven gene families, cyp71A1, cyp71A2, cyp71A4, cyp71B1, cyp72A1 (encoding secologanin synthase), cyp76C4, cyp81D1, cyp83B1, cyp90b1 and cyp93A1. There are no previous reports implicating cyt P450 genes in thermotolerance, however, Larkindale and Vierling [60] indicated the downregulation of 18 different cyp genes in Arabidopsis under high temperature stress. Other reports indicated the involvement of some of these genes in other biological processes. For example, cyp71A1 and cyp72A1 (encoding secologanin synthase) genes are involved in the synthesis of indole alkaloid secologanin, which is important in mevalonate pathway for the production of the two anticancer bisindole alkaloids vinblastine and vincristine [61]. cyp83B1 is involved in the biosynthesis of glucosinolates, which have anticancer and flavoring functions [62]. cyp71A4 is involved in the defense response to pathogen attacks [63]. In conclusion, the high levels of downregulation of a large number of cyp genes in response to heat stress in leaves of R. stricta is not fully understood.
Large numbers of upregulated, abundant transcripts of genes encoding HSPs and chaperones (or chaperonin) were detected in leaves of R. stricta (Additional file 12: Table S4 & Additional file 13: Table S5). These genes are frequently reported as being involved in plant thermotolerance (e.g., Hu et al. [58]). HSPs are protective proteins acting as molecular chaperones that prevent protein misfolding and aggregation or denaturation during heat stress [64]. Recent reports indicated that ATP-independent chaperones act with sHSPs as “holdases” to suppress the aggregation of proteins and delay their folding under heat stress [65]. ATP-independent chaperones also assist with protein refolding under heat stress to recover original protein structures [66]. There are two major groups of HSPs, high molecular mass or HMM-HSPs ranging from 60 to 100 KDa and small sHSPs ranging from 15 to 30 kDa [64]. Genes within these two groups were classified into five gene families based on intracellular localization. Classes I and II are cytosolic, while classes III, IV and V are localized in the chloroplast, mitochondrion or endoplasmic reticulum [64, 67]. In the present study, upregulated, abundant transcripts encoding HMM-HSPs in leaves of R. stricta during the day were cytosolic of class I, while those encoding sHSPs were either cytosolic of class II or chloroplastic of classes III or IV (Additional file 12: Table S4). Transgenics of different plant species with hsp genes, especially chloroplastic (CP), showed enhanced tolerance to heat stress [68–72] mainly via the protection of Photosystem II (PSIIHSP) [55]. The amount of CP-sHSPs in per unit protein correlated positively with the level of tolerance in Chenopodium album and Lycopersicum esculentum [73]. Upregulated, abundant transcripts encoding UDP-glycosyltransferase also occurred in leaves of R. stricta (Additional file 12: Table S4). They involve eight genes belonging to five gene families, ugt82A1, ugt85A1, ugt85A2, ugt85A3, ugt87A1, ugt87A2, ugt90A1 and ugt91C1. The enzyme is a key player in the quality control mechanism for newly synthesized glycoproteins in the endoplasmic reticulum (ER). This organelle hosts the synthesis/folding of proteins secreted extracellularly or delivered to endomembrane system [74]. Quality control includes the calnexin (CNX)/calreticulin (CRT) cycle, which involves lectin-chaperones retaining N-glycosylated proteins at the ER while they undergo the folding process. UDP-glucosyltransferase acts on unglycosylated proteins during folding and catalyzes reglucosylation to allow protein binding to CNX/CRT and retention in the ER to continue the folding process [75]. In contrast, several highly downregulated transcripts encoding U-box containing proteins occurred in leaves of R. stricta, such as E3 ubiquitin-protein ligase atl42. The genes include pub15, 25, 26, 35 and 43 (Additional file 12: Table S4). The U-box protein family represents a class of E3 enzymes (e.g., CHIP) acting as a degradatory co-chaperone of HSP70 and HSP90 [76]. This is another mechanism of protein quality control under heat stress where the interaction of HSPs with co-chaperones that have either folding or degradatory activity determines the fate of HSP proteins. In transgenic Arabidopsis with overexpressed pub22 and 23 genes, two homologous U-box E3 ubiquitin ligases caused hypersensitivity to drought stress [77, 78]. In contrast, loss-of-function of these two genes resulted in the recovery of drought-tolerant plants. In our analysis, the high levels of downregulation of pub23 as well as the other genes encoding U-box containing proteins in leaves of R. stricta indicates that the degradatory activity of these enzymes on target proteins under heat stress is unlikely (Additional file 12: Table S4). In conclusion, the occurrence of upregulated, abundant transcripts encoding HSPs, chaperones and UDP-glucosyltransferase and downregulated transcripts encoding U-box containing proteins likely contribute to thermotolerance in leaf cells of R. stricta by maintaining proper protein folding and preventing protein degradation.
Upregulated, abundant transcripts encoding aquaporins (AQPs) or major intrinsic proteins (MIPs) also occurred in leaves of R. stricta (Additional file 12: Table S4). The main types of MIPs include plasma membrane intrinsic proteins (PIPs) and tonoplast intrinsic proteins (TIPs). In R. stricta, three MIP genes, pip2-1, pip1-2 and tip2-1, were detected and may be involved in thermotolerance in leaves. AQPs are classes of membrane proteins that facilitate water diffusion across cell membranes during the response/tolerance to adverse environmental stimuli [79, 80] including heat stress [81]. They are also involved in the opening and closing of cellular gates and in the physiology of water balance and water use efficiency under abiotic stresses [82]. We propose that the upregulation of the three genes encoding AQPs may contribute to thermotolerance of R. stricta leaves. Finally, upregulated, abundant transcripts encoding protein transparent testa 12 (tt12) occurred in leaves of R. stricta (Additional file 12: Table S4). Debeaujon et al. [83] indicated that flavonoids were sequestered in seed coat endothelium due to the action of the tt12 gene but our results suggest that upregulation of this gene in leaves is a response to heat stress. This conclusion complements the results of KEGG analysis of flavonoid biosynthesis under heat stress because this process is likely followed by sequestering of accumulated flavonoids in the vacuole of leaf cells. As indicated earlier, flavonoids are key secondary metabolites in abiotic stress tolerance [50].
Transcription factors co-expressed with Hsp or chaperone genes
Co-expression analysis was done to investigate the transcription factors that may regulate genes encoding HSPs or chaperones in apical and mature leaves of R. stricta with special emphasis on those upregulated at the sampling times with the highest temperatures at midday (Additional file 13: Table S5). The clusters of DE transcripts selected for co-expression analysis in either leaf type were those with the highest number of the target genes (Additional file 14: Table S6). They are clusters 4, 7, 10, 18 and 26 for apical leaves (Additional file 1: Table S1), and 8 and 18 for mature leaves (Additional file 2: Table S2). Selected TFs co-expressed with genes encoding HSPs and chaperones are shown in Additional file 14: Table S6. These TFs are known for their response to abiotic stresses, including HSFAs, AP2/ERF (or AP2-EREBP), WRKY, bHLH, zinc finger and MYB (Additional file 14: Table S6). Co-expressed hsp genes encode HSP10, HSP20, HSP60, HSP70, HSP83 and HSP90. The results indicated several highly concordant co-expressions of TFs and genes encoding either HSPs or chaperones during the day. Five pairs of these co-expressing genes in both apical and mature leaves were selected for further analysis (Fig. 14). The expression pattern of each gene pair was the same in both types of leaves. The highly similar co-expression across the day in each of the five gene pairs is unlikely to be coincidental. Expression patterns of these five pairs included upregulation starting midday (clusters 7 of apical leaves and 8 of mature leaves) and upregulation at midday (clusters 10, 18 and 26 of apical leaves and 18 of mature leaves). The first pattern indicated co-expression of genes encoding HSFA3 and endoplasmin/HSP90, and co-expression of genes encoding HSFA4 and HSP83 (Additional file 14: Table S6 and Fig. 14). The second expression pattern involved co-expression of genes encoding AP2/ERF (or AP2-EREBP) and 10 KDa chaperonin (clusters 10 of apical leaves and 18 of mature leaves, Additional file 14: Table S6 and Fig. 14), co-expression of genes encoding WRKY27-like and HSP20/α-crystallin (clusters 18 of both apical and mature leaves, Additional file 14: Table S6 and Fig. 14) and co-expression of genes encoding HSFA2 and HSP70 (clusters 26 of apical leaves and 18 of mature leaves, Additional file 14: Table S6 and Fig. 14).
Fig. 14.
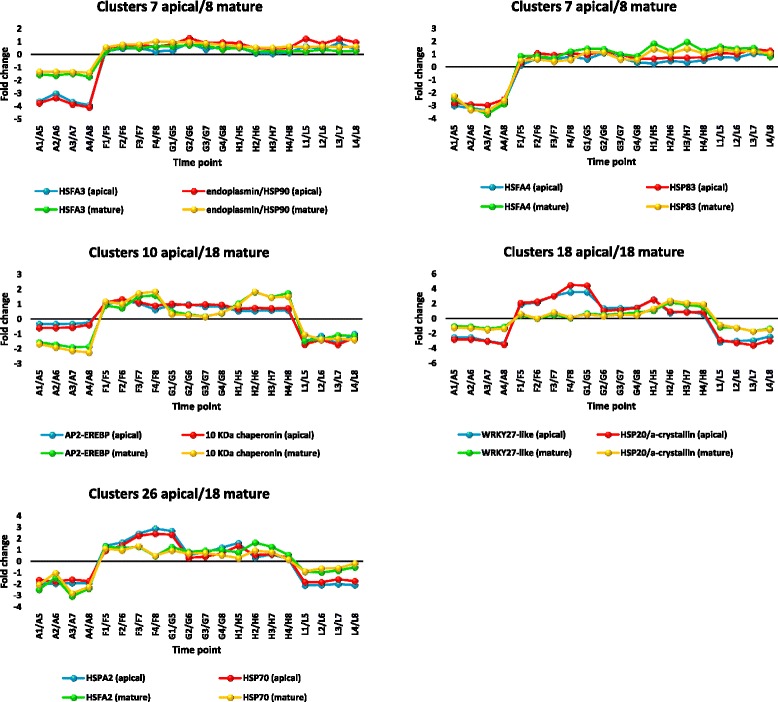
Selected transcription factors with very similar co-expression patterns with HSP or chaperone genes in apical (A1-L4) and mature leaves (A5-L8) of R. stricta at different time points of the day (A, morning; F-H, midday & L, dusk). Expression patterns involved upregulation starting midday (clusters 7 (apical)/8 (mature)) and upregulation at midday (clusters 10, 18 or 26 (apical)/18 (mature)). Transcript fold change refers to that described in Additional file 14: Table S6
The expression of hsp genes in response to various stimuli is regulated by HSFs [84]. Our results indicated the co-expression of the upregulated, abundant transcripts encoding HSP70, HSP90 and HSP83 with the upregulated, abundant transcripts encoding HSFA2, HSFA3 and HSFA4, respectively (Additional file 14: Table S6 and Fig. 14). There are three clases of HSFs in Arabidopsis, A, B and C [85]. Only class HSF A harbors an AHA motif essential for transcription activation of hsp genes [86, 87]. We observed five upregulated, abundant HSF transcripts in leaves of R. sticta that encode HSFA1, HSFA2, HSFA3, HSFA4 and HSFB1 (Additional file 12: Table S4). Earlier reports on the gene encoding HSFA1 indicated that it functions as a master regulator of early heat stress response in tomato [88] and plays an important role in the induction of several hsp genes in Arabidopsis [89]. The gene encoding HSFA2 is a heat-inducible transactivator sustaining the expression of hsp genes and promoting an extended duration of acquired thermotolerance in Arabidopsis [90]. Disruption of the gene encoding HSFA2 resulted in a reduction in expression of shsp genes Hsp18.1, Hsp25.3-P and Hsa32 under stress, while overexpression resulted in enhanced thermotolerance [91]. However, our results indicated the co-expression of the transcript encoding HSFA2 with a HMM-hsp transcript encoding HSP70 in leaves of R. stricta. As for the gene encoding HSFA3, Yoshida et al. [92] found that it is highly upregulated under heat stress in transgenic plants overexpressing dreb2A gene. This indicates that dreb2A acts upstream of the gene encoding HSFA3. In turn, hsfA3 was reported to regulate expression of many heat-inducible genes acting downstream in the transcriptional cascade as this gene acts as a potent activator on the hsp gene promoters [93]. hsfA3-knockout mutant lines of Arabidopsis showed reduced levels of hsp101 and shsps under heat stress. We have no evidence for the expression of hsp101 in leaves of R. stricta. Rather, our results indicated the co-expression of genes encoding HSFA3 and the HMM-hsp gene encoding a HSP90. The gene encoding HSFA4 in leaves of R. stricta co-expressed with the hsp83 gene (Fig. 14). This TF was reported recently to enhance transient transactivation of hsp17.6A transcription under heat stress [94]. It was also reported to confer tolerance to salt and oxidative stresses. Our results contradict those of Pérez-Salamó et al. [94] as this TF co-expressed with a gene encoding a HMM-HSP rather than the sHSP17.6A protein. In general, we can conclude that the HSF machinery in R. stricta basically acts on HMM-hsp genes. None of the detected HSFAs involved in co-expression in the present study drives expression of shsp genes as reported in other plant species, such as Arabidopsis.
Our results indicate that one upregulated, abundant transcript encoding AP2-EREBP was co-expressed with an upregulated, abundant transcript encoding 10 KDa chaperonin (Additional file 14: Table S6 and Fig. 14) in leaves of R. stricta. A large number of transcripts encoding AP2/ERFs or AP2-EREBP were also upregulated during the day in R. stricta leaves (Additional file 14: Table S6). They include genes encoding AP2-ERF17, AP2-ERF34, AP2-ERF98, AP2-ERF109 and AP2-EREBP (AT2G41710 in Arabidopsis). Two more genes encoding AP2-ERF were differentially expressed in R. stricta including the upregulated gene encoding AP2-ERF23 and the downregulated gene encoding AP2-ERF10 (Additional file 2: Table S2 & Additional file 6: Table S3). Yates et al. [31] indicated that the gene family encoding AP2-EREBP showed a decrease in expression at midday. These results apply in the present study to genes encoding AP2-ERF04, AP2-ERF5, AP2-ERF12, AP2-ERF13, AP2-ERF1b, AP2-ERFabr1 and AP2-ERFwin1 (Additional file 2: Table S2 & Additional file 6: Table S3). In general, ERFs were thought to act upstream of HSFs in the response to heat stress especially at night [95], but no previous reports indicate that ERF drives expression of genes encoding chaperonin. AP2/ERFs contain at least one DNA binding domain, AP2 domain [96]. They were induced by biotic and abiotic stresses including drought, heat, salt, osmotic, wounding, etc. [97]. The most extensively studied ERFs in abiotic stress responses were the DREB proteins. Among them, DREB2A in Arabidopsis was induced by heat in an ABA-ethylene-independent manner and plants overexpressing it exhibited improved tolerance to heat stress [98].
We also observed the co-expression of the upregulated genes encoding WRKY27 and HSP20/α-crystallin in apical and mature leaves of R. stricta (Fig. 14). This TF was reported in Arabidopsis to influence wilt disease symptom caused by Ralstonia solanacearum [99], but there are no reports supporting its involvement in heat stress. Three other genes encoding WRKY28, WRKY35 and WRKY56 were also upregulated in leaves of R. stricta but with lower rates. Previous reports indicated that one WRKY transcription factor and four HSFs were induced by heat stress in switchgrass [100]. Yates et al. [31] indicated that the gene family encoding WRKY showed decreased expression at midday. We confirmed these results for genes encoding WRKY6, WRKY7, WRKY12, WRKY33, WRKY40, WRKY44, WRKY46, WRKY48, WRKY49, WRKY50, WRKY53 and WRKY55 (Additional file 2: Table S2 & Additional file 6: Table S3). There are reports indicating that the constitutive expression of genes encoding WRKY25, WRKY26 or WRKY33 enhanced tolerance to heat stress [101]. No previous reports have indicated the direct involvement of WRKY in driving hsp genes, including the shsp20. Calmodulin-dependent protein kinases were induced in switchgrass by heat stress [100]. A number of transcripts encoding calmodulin-dependent protein kinase were detected in leaves of R. stricta at different rates across the day (Additional file 2: Table S2 & Additional file 6: Table S3). Investigations of the role of calmodulin in cells of Arabidopsis indicated that it was required for heat stress signaling and can serve as an activator of WRKY39 and a number of HSFs [102]. It seems that this is not the case in leaves of R. stricta.
Conclusion
In general, we conclude that enzymes in several pathways are interacting in the biosynthesis of soluble sugars, polyols, secondary metabolites, phenolics and methionine. Genes encoding these enzymes can be primary contributors to thermotolerance in the wild plant species R. stricta. A number of heat-responsive genes are regulated under heat stress and can add to the thermotolerance of this plant species. Our analyses also indicate the relationship of individual genes and transcription factors during heat stress. Overall, our results contribute to the knowledge of the regulatory mechanisms underlying heat stress response at the molecular level in R. stricta. This is particularly important because the native habitat of this species is extremely hot, making it an excellent system for examining plant responses to heat stress. In the future, manipulating the expression of genes affecting protein folding and degradation, such as those encoding HSPs, chaperones, UDP-glucosyltransferase and U-box, holds great potential for improving thermotolerance of economically important crop plants.
Methods
Plant sampling and RNA-Seq data
Apical and mature leaves of R. stricta were collected from the Bahrah region, Jeddah, Saudi Arabia. The voucher specimen was deposited in the Department of Biological Sciences Herbarium at King Abdulaziz University (Number 1150/M/75 collected by N. Baeshen, M. Baeshen and J. Sabir). Samples of the two leaf types were taken in four replicates at seven time points of the day from which the following five time points of the day were selected; morning (A, 07:10), midday (F, 13:25; G, 14:05 & H, 14:30) and dusk (L, 18:27). The temperatures at the five time points were 27.4 (A), 42.4 (F), 42.2 (G), 40.0 (H) and 33.5 °C (L). The total number of samples across the two leaf types was 40 (2 leaf types X 5 time points X 4 replicates). The morning and dusk samples were taken immediately after sunrise and immediately before sunset, respectively. The first time point is considered as the control condition for heat stress, while time points F-H were collectively designated midday. RNA samples were isolated [31] and sequenced by Genome Enterprise Limited (GEL) at The Genome Analysis Centre (Norwich, UK) and the resulting raw reads from the RNA-Seq data were deposited in the short read archive (SRA) of the NCBI (study SRP028238).
Bioinformatics analysis
Relative abundance of reads was calculated by RSEM v1.1.6 with the R. stricta nuclear genome [103] used as a reference. By default, RSEM uses the Bowtie aligner [104] (Bowtie v0.12.1) to map the reads against the transcripts. Expected read counts were used as input to DE analysis by EdgeR (version 3.0.0, R version 2.1.5). The median of these values was used as the common dispersion factor for DE analyses. DE transcripts were annotated and KEGG pathway analyses were performed using Blast2GO software [105] (version 2.3.5, https://www.blast2go.com). Further, Blastx was performed for DE transcripts in selected clusters against the NCBI non-redundant protein database with an E value cut-off of 1e−5. HMMER v3.1b2 was used to identify protein domains common in TFs to detect TF genes possibly involved in regulating expression of DE transcripts encoding HSPs and chaperones in the selected clusters. Differential expression data were introduced in fold change of transcript levels of either leaf type under heat stress at midday as compared to those of the control condition at the morning.
Validation of RNA-Seq datasets
An experiment was conducted to validate the RNA-Seq data for selected genes whose expression pattern in apical leaves was similar to that in mature leaves, and whose expression rate was mostly consistent within the four replicates of each time point. Expression patterns suitable for the validation experiment across the two types of leaves included upregulation starting at midday and gradual downregulation across the day in which 10 transcripts were randomly selected and actin was used as the unregulated housekeeping gene (Additional file 6: Table S3). Three out of the four replicates of RNA samples for the original RNA-Seq study [31] were utilized in the experiment to validate expression patterns of the selected genes across the different time points of the day (A, morning; F-H, midday; L, dusk) via semi-quantitative RT-PCR. Primers were designed using Netprimer software (http://www.premierbiosoft.com/netprimer/index.html) with the following criteria: length ~20 bases, GC content ~50%, minimal secondary structure, comparable annealing temperatures (55 °C) of the primer pairs, and PCR products of ~350–450 bp (Additional file 6: Table S3).
Acknowledgments
The authors gratefully acknowledge the financial support from King Abdulaziz University (KAU) Director Prof. Dr. Abdulrahman O. Alyoubi and the KAU Deanship of Scientific Research, Jeddah, Saudi Arabia, represented by the Unit of Strategic Technologies Research through the Project number 431/008-D for the Project entitled: “Identification and Isolation of Salt and Heat-Tolerance Genes of Rhazya stricta and Detection of Metabolites and their Therapeutic Effects via Cheminformatics”.
Funding
King Abdulaziz University (KAU) Director Prof. Dr. Abdulrahman O. Alyoubi and the KAU Deanship of Scientific Research, Jeddah, Saudi Arabia, represented by the Unit of Strategic Technologies Research through the Project number 431/008-D.
Availability of data and materials
The raw reads from the RNA-Seq data were deposited in the short read archive (SRA) of the NCBI (study SRP028238). Permission to collect Rhazya stricta plant material and to perform fieldwork at the site was granted by the Governor of Makkah Province, Prince Khalid Al- Faisal.
Authors’ contributions
AYO, JSMS, AA, SE, FMD, NH, RKJ, AB conceived and designed the experiments. AA, XL, MZM, NOG, NHH, MAK performed the experiments. AA, SE, FMD, AB analyzed the data. AYO, JSMS, FMD, NH, RKJ, AB wrote the manuscript. All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Ethics approval and consent to participate
Not applicable.
Abbreviations
- AQP
Aquaporin
- bHLH
Basic helix-loop-helix
- CDK
Cyclin-dependent kinase
- CNX
Calnexin
- CRT
Calreticulin
- Cyt 450
Cytochromome P450
- DE
Differential expression
- ER
Endoplasmic reticulum
- FC
Fold change
- HB
Homeobox domain
- HSP
Heat shock protein
- KDa
Kilodalton
- KEGG
Kyoto encyclopedia of genes and genomes
- LAR
Leucocyanidin reductase
- MIP
Major intrinsic protein
- PA
Proanthocyanidin
- PAL
Phenylalanine ammonia-lyase
- PIP
Plasma membrane intrinsic protein
- PSII
Photosystem II
- ROS
Reactive oxygen species
- sHSP
Small heat shock protein
- SRA
Short read archive
- TF
Transcription factor
- TIP
Tonoplast intrinsic protein
Additional files
Fold change values of assembled transcripts of R. stricta SRA database in different clusters in the apical leaves (A1-L4) at different time points of the day (A, morning; F-H, midday & L, dusk). (XLSX 775 kb)
Fold change values of assembled transcripts of R. stricta SRA database in different clusters in the mature leaves (A5-L8) at different time points of the day (A, morning; F-H, midday & L, dusk). (XLSX 1523 kb)
Clusters of assembled transcripts of R. stricta in apical leaves (A1-L4) at different time points of the day (A, morning; F-H, midday & L, dusk). Grey lines indicate expression patterns of individual transcripts in a given cluster. Blue lines indicate overall expression pattern across different transcripts of a given cluster. (PDF 223 kb)
Clusters of assembled transcripts of R. stricta SRA in mature leaves (A5-L8) at different time points of the day (A, morning; F-H, midday & L, dusk). Grey lines indicate expression patterns of individual transcripts in a given cluster. Blue lines indicate overall expression pattern across different transcripts of a given cluster. (PDF 397 kb)
Semi-quantitative RT-PCR and profiles of fold change values resulting from RNA-Seq analysis for selected upregulated genes starting midday (1–5) and gradually downregulated genes (6-10) used for validating RNA-Seq data of apical (A1-L4) and mature (A5-L7) leaves of R. stricta collected at different time points of the day. Serial numbers 1–10 refer to genes described in Additional file 6: Table S3. The “actin” gene was used as the unregulated house-keeping gene. (DOCX 9867 kb)
Fold change values of transcripts in two expression patterns (e.g., upregulation starting midday and gradual downregulation) in the apical (A1-L4) and mature (A5-L8) leaves of R. stricta at different time points of the day (A, morning; F-H, midday & L, dusk) selected for validation experiment along with primer information for semi-quantitative RT-PCR. The “actin” was used as the unregulated house-keeping gene. (XLSX 35 kb)
Enzymes in the phenylpropanoid metabolic pathway in apical and mature leaves responded differentially to changing environment at two time points (morning, A and midday, G). Upregulated (activated) in apical leaves (blue), upregulated in mature leaves (red), downregulated (repressed) in apical leaves (orange box), downregulated in mature leaves (green box). (DOCX 174 kb)
Enzymes in the flavonoid metabolic pathway in apical and mature leaves responded differentially to changing environments at two time points (morning, A and midday, G). Upregulated (activated) in apical leaves (blue), upregulated in mature leaves (red). (DOCX 193 kb)
Enzymes in the anthocyanin metabolic pathway in apical and mature leaves responded differentially to changing environment at two time points (morning, A and midday, G). Upregulated (activated) in apical leaves (blue), upregulated in mature leaves (red). (DOCX 210 kb)
Enzymes in the carotenoid metabolic pathway in apical and mature leaves responded differentially to changing environment at two time points (morning, A and midday, G). Upregulated (activated) in apical leaves (blue), upregulated in mature leaves (red), downregulated (repressed) in apical leaves (orange box), downregulated in mature leaves (green box). (DOCX 242 kb)
Enzymes in the cysteine and methionine metabolic pathways in apical and mature leaves responded differentially to changing environment at two time points (morning, A and midday, G). Upregulated (activated) in apical leaves (blue), upregulated in mature leaves (red), downregulated (repressed) in apical leaves (orange box), downregulated in mature leaves (green box). (DOCX 201 kb)
Comparative differential expression of genes of R. stricta transcriptomes of apical (a) and mature (b) leaves with fold change of ≥ 5 in different time points (A, morning; F-H, midday & L, dusk). Blue box = upregulation, orange box = downregulation. Numbers between parentheses represent no. analogs of a given gene. Green boxes indicate genes selected for further analysis. (DOCX 80 kb)
Fold change values of selected upregulated genes encoding HSPs in apical (A1-L4) and mature (A5-L8) leaves during the day (A, morning; F-H, midday & L, dusk) in R. stricta (XLSX 25 kb)
Selected differentially expressed genes in selected clusters of apical (a, A1-L4) and mature (b, A5-L8) leaves indicating the TFs co-expressed with Hsp and chaperone genes during the day (A, morning; F-H, midday & L, dusk) in R. stricta. (XLSX 42 kb)
Contributor Information
Abdullah Y. Obaid, Email: aobaid@kau.edu.sa
Jamal S. M. Sabir, Email: jsabir2622@gmail.com
Ahmed Atef, Email: ahmed_atefaig2@yahoo.com.
Xuan Liu, Email: Xuan.Liu@liverpool.ac.uk.
Sherif Edris, Email: sedris@aucegypt.edu.
Fotouh M. El-Domyati, Email: fm_domyati@hotmail.com
Mohammed Z. Mutwakil, Email: mmutwakil@kau.edu.sa
Nour O. Gadalla, Email: nouromar71@yahoo.com
Nahid H. Hajrah, Email: nhajrah260@gmail.com
Magdy A. Al-Kordy, Email: m_alkordy@yahoo.com
Neil Hall, Email: neil.hall@tgac.ac.uk.
Ahmed Bahieldin, Email: bahieldin55@gmail.com.
Robert K. Jansen, Email: jansen@austin.utexas.edu
References
- 1.Zhang Y, Mian MR, Chekhovskiy K, So S, Kupfer D, et al. Differential gene expression in Festuca under heat stress conditions. J Exp Bot. 2005;56:897–907. doi: 10.1093/jxb/eri082. [DOI] [PubMed] [Google Scholar]
- 2.Xu C, Huang B. Root proteomic responses to heat stress in two Agrostis grass species contrasting in heat tolerance. J Exp Bot. 2008;59:4183–94. doi: 10.1093/jxb/ern258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Shah F, Huang J, Cui K, Nie L, Shah T, et al. Impact of high temperature stress on rice plant and its traits related to tolerance. J Agr Sci. 2011;149:545–56. doi: 10.1017/S0021859611000360. [DOI] [Google Scholar]
- 4.IPCC et al. Managing the risks of extreme events and disasters to advance climate change adaptation. In: Field CB, Barros V, Stocker TF, Qin D, Dokken DJ, et al., editors. A special report of working groups I and II of the intergovernmental panel on climate change. Cambridge: Cambridge University Press; 2012. p. 582. [Google Scholar]
- 5.Jiang YW, Huang BR. Drought and heat stress injury to two cool-season turfgrasses in relation to antioxidant metabolism and lipid peroxidation. Crop Sci. 2001;41:436–42. doi: 10.2135/cropsci2001.412436x. [DOI] [Google Scholar]
- 6.Rizhsky L, Liang H, Mittler R. The combined effect of drought stress and heat shock on gene expression in tobacco. Plant Physiol. 2002;130:1143–51. doi: 10.1104/pp.006858. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Rizhsky L, Liang HJ, Shuman J, Shulaev V, Davletova S, et al. When defense pathways collide. The response of Arabidopsis to a combination of drought and heat stress. Plant Physiol. 2004;134:1683–96. doi: 10.1104/pp.103.033431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Szabados L, Savouré A. Proline: a multifunctional amino acid. Trends Plant Sci. 2010;15:89–97. doi: 10.1016/j.tplants.2009.11.009. [DOI] [PubMed] [Google Scholar]
- 9.Wahid A, Close TJ. Expression of dehydrins under heat stress and their relationship with water relations of sugarcane leaves. Biol Plant. 2007;51:104–9. doi: 10.1007/s10535-007-0021-0. [DOI] [Google Scholar]
- 10.Hartl FU, Bracher A, Hayer-Hartl M. Molecular chaperones in protein folding and proteostasis. Nature. 2011;475:324–32. doi: 10.1038/nature10317. [DOI] [PubMed] [Google Scholar]
- 11.Suzuki N, Rizhsky L, Liang H, Shuman J, Shulaev V, et al. Enhanced tolerance to environmental stress in transgenic plants expressing the transcriptional coactivator multiprotein bridging factor 1c. Plant Physiol. 2005;139:1313–22. doi: 10.1104/pp.105.070110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kotak S, Larkindale J, Lee U, von Koskull-Döring P, Vierling E, et al. Complexity of the heat stress response in plants. Curr Opin Plant Biol. 2007;10:310–6. doi: 10.1016/j.pbi.2007.04.011. [DOI] [PubMed] [Google Scholar]
- 13.Zinn KE, Tunc-Ozdemir M, Harper JF. Temperature stress and plant sexual reproduction: uncovering the weakest links. J Exp Bot. 2010;61:1959–68. doi: 10.1093/jxb/erq053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Howarth C. Molecular responses of plants to an increased incidence of heat shock. Plant Cell Environ. 1991;1:831–41. doi: 10.1111/j.1365-3040.1991.tb01446.x. [DOI] [Google Scholar]
- 15.Tian J, Belanger FC, Huang B. Identification of heat stress-responsive genes in heat-adapted thermal Agrostis scabra by suppression subtractive hybridization. J Plant Physiol. 2009;166:588–601. doi: 10.1016/j.jplph.2008.09.003. [DOI] [PubMed] [Google Scholar]
- 16.Giorno F, Wolters-Arts M, Grillo S, Scharf KD, Vriezen WH, et al. Developmental and heat stress-regulated expression of HsfA2 and small heat shock proteins in tomato anthers. J Exp Bot. 2010;61:453–62. doi: 10.1093/jxb/erp316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Nakamoto H, Vigh L. The small heat shock proteins and their clients. Cell Mol Life Sci. 2007;64:294–306. doi: 10.1007/s00018-006-6321-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Hartl FU, Hayer-Hartl M. Converging concepts of protein folding in vitro and in vivo. Nat Struct Mol Biol. 2009;16:574–81. doi: 10.1038/nsmb.1591. [DOI] [PubMed] [Google Scholar]
- 19.Pratt WB, Morishima Y, Peng HM, Osawa Y. Proposal for a role of the Hsp90/Hsp70-based chaperone machinery in making triage decisions when proteins undergo oxidative and toxic damage. Exp Biol Med. 2010;235:278–89. doi: 10.1258/ebm.2009.009250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Taipale M, Jarosz DF, Lindquist S. HSP90 at the hub of protein homeostasis: emerging mechanistic insights. Nat Rev Mol Cell Biol. 2010;11:515–28. doi: 10.1038/nrm2918. [DOI] [PubMed] [Google Scholar]
- 21.Huner N, Öquist G, Sarhan F. Energy balance and acclimation to light and cold. Trends Plant Sci. 1998;3:224–30. doi: 10.1016/S1360-1385(98)01248-5. [DOI] [Google Scholar]
- 22.Hu T, Li H, Zhang X, Luo H, Fu J. Toxic effect of NaCl on ion metabolism, antioxidative enzymes and gene expression of perennial ryegrass. Ecotox Environ Safe. 2011;74:2050–6. doi: 10.1016/j.ecoenv.2011.07.013. [DOI] [PubMed] [Google Scholar]
- 23.Sairam RK, Tyagi A. Physiology and molecular biology of salinity stress tolerance in plants. Curr Sci. 2004;86:407–21. [Google Scholar]
- 24.Xu HW, LuY TSY, Song FB. Lipid peroxidation, antioxidant enzyme activity and osmotic adjustment changes in husk leaves of maize in black soils region of Northeast China. Afr J Agric Res. 2011;6:3098–102. [Google Scholar]
- 25.Qin D, Wu H, Peng H, Yao Y, Ni Z, et al. Heat stress-responsive transcriptome analysis in heat susceptible and tolerant wheat (Triticum aestivum L.) by using wheat genome array. BMC Genomics. 2008;9:432. doi: 10.1186/1471-2164-9-432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Finka A, Mattoo RU, Goloubinoff P. Meta-analysis of heat and chemically upregulated chaperone genes in plant and human cells. Cell Stress Chap. 2011;16:15–31. doi: 10.1007/s12192-010-0216-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Bokszczanin KL. Perspectives on deciphering mechanisms underlying plant heat stress response and thermotolerance. Frontiers in Plant Sci. 2013;4:315. doi: 10.3389/fpls.2013.00315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Mittler R, Finka A, Goloubinoff P. How do plants feel the heat? Trends Biochem Sci. 2012;37:118–25. doi: 10.1016/j.tibs.2011.11.007. [DOI] [PubMed] [Google Scholar]
- 29.Latchman DS. Transcription factors: an overview. Int J Biochem Cell Biol. 1997;29:1305–12. doi: 10.1016/S1357-2725(97)00085-X. [DOI] [PubMed] [Google Scholar]
- 30.Lindemose S, O’Shea C, Jensen MK, Skriver K. Structure, function and networks of transcription factors involved in abiotic stress responses. Int J Mol Sci. 2013;14:5842–78. doi: 10.3390/ijms14035842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Yates SA, Chernukhin I, Alvarez-Fernandez R, Bechtold U, Baeshen M, et al. The temporal foliar transcriptome of the perennial C3 desert plant Rhazya stricta in its natural environment. BMC Plant Biol. 2014;14:2. doi: 10.1186/1471-2229-14-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Bowman JL, Eshed Y. Formation and maintenance of the shoot apical meristem. Trends Plant Sci. 2000;5:110–5. doi: 10.1016/S1360-1385(00)01569-7. [DOI] [PubMed] [Google Scholar]
- 33.Chaitanya KV, Sundar D, Reddy AR. Mulberry leaf metabolism under high temperature stress. Biol Plant. 2001;44:379–84. doi: 10.1023/A:1012446811036. [DOI] [Google Scholar]
- 34.Vu JCV, Gesch RW, Pennanen AH, Allen LHJ, Boote KJ, et al. Soybean photosynthesis, rubisco and carbohydrate enzymes function at supra-optimal temperatures in elevated CO2. J Plant Physiol. 2001;158:295–307. doi: 10.1078/0176-1617-00290. [DOI] [Google Scholar]
- 35.Cenci U, Nitschke F, Steup M, Minassian BA, Colleoni C, et al. Transition from glycogen to starch metabolism in Archaeplastida. Trends Plant Sci. 2014;19:18–28. doi: 10.1016/j.tplants.2013.08.004. [DOI] [PubMed] [Google Scholar]
- 36.Krasensky J, Jonak C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J Exp Bot. 2012;63:1593–1608. doi: 10.1093/jxb/err460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Tetlow IJ, Morell MK, Emes MJ. Recent developments in understanding the regulation of starch metabolism in higher plants. J Exp Bot. 2004;55:2131–45. doi: 10.1093/jxb/erh248. [DOI] [PubMed] [Google Scholar]
- 38.Kotting O, Kossmann J, Zeeman SC, Lloyd JR. Regulation of starch metabolism: the age of enlightenment? Curr Opin Plant Biol. 2010;13:321–9. doi: 10.1016/j.pbi.2010.01.003. [DOI] [PubMed] [Google Scholar]
- 39.Lawson T, Davey PA, Yates SA, Bechtold U, Baeshen M, et al. C3 photosynthesis in the desert plant Rhazya stricta is fully functional at high temperatures and light intensities. New Phytol. 2014;201:862–73. doi: 10.1111/nph.12559. [DOI] [PubMed] [Google Scholar]
- 40.Crafts-Brandner SJ, Salvucci ME. Rubisco activase constrains the photosynthetic potential of leaves at high temperature and CO2. Proc Natl Acad Sci U S A. 2000;97:13430–5. doi: 10.1073/pnas.230451497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Salvucci ME, Crafts-Brandner SJ. Inhibition of photosynthesis by heat stress: the activation state of Rubisco as a limiting factor in photosynthesis. Plant Physiol. 2004;120:179–86. doi: 10.1111/j.0031-9317.2004.0173.x. [DOI] [PubMed] [Google Scholar]
- 42.Portis AR. Rubisco activase–Rubisco’s catalytic chaperone. Photosynthesis Res. 2003;75:11–27. doi: 10.1023/A:1022458108678. [DOI] [PubMed] [Google Scholar]
- 43.Todaka D, Matsushima H, Morohashi Y. Water stress enhances beta-amylase activity in cucumber cotyledons. J Exp Bot. 2000;51:739–45. doi: 10.1093/jexbot/51.345.739. [DOI] [PubMed] [Google Scholar]
- 44.Kaplan F, Guy CL. Beta-Amylase induction and the protective role of maltose during temperature shock. Plant Physiol. 2004;135:1674–84. doi: 10.1104/pp.104.040808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Basu PS, Ali M, Chaturvedi SK. Osmotic adjustment increases water uptake, remobilization of assimilates and maintains photosynthesis in chickpea under drought. Indian J Exp Biol. 2007;45:261–7. [PubMed] [Google Scholar]
- 46.Kempa S, Krasensky J, Dal Santo S, Kopka J, Jonak C. A central role of abscisic acid in stress-regulated carbohydrate metabolism. PLoS One. 2008;3:e3935. doi: 10.1371/journal.pone.0003935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Gründel M, Scheunemann R, Lockau W, Zilliges Y. Impaired glycogen synthesis causes metabolic overflow reactions and affects stress responses in the cyanobacterium Synechocystis sp. PCC 6803. Microbiol. 2012;158:3032–43. doi: 10.1099/mic.0.062950-0. [DOI] [PubMed] [Google Scholar]
- 48.Shen B, Jensen RG, Bohnert HJ. Increased resistance to oxidative stress in transgenic plants by targeting mannitol biosynthesis to chloroplasts. Plant Physiol. 1997;113:527–32. doi: 10.1104/pp.115.2.527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Rosa M, Prado C, Podazza G, Interdonato R, González JA, et al. Soluble sugars—Metabolism, sensing and abiotic stress. A complex network in the life of plants. Plant Signal Behav. 2009;4:388–93. doi: 10.4161/psb.4.5.8294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Wahid A, Ghazanfar A. Possible involvement of some secondary metabolites in salt tolerance of sugarcane. J Plant Physiol. 2006;163:723–30. doi: 10.1016/j.jplph.2005.07.007. [DOI] [PubMed] [Google Scholar]
- 51.Winkel-Shirley B. Biosynthesis of flavonoids and effects of stress. Curr Opin Plant Biol. 2002;5:218–23. doi: 10.1016/S1369-5266(02)00256-X. [DOI] [PubMed] [Google Scholar]
- 52.Cohen SD, Tarara JM, Gambetta GA, Matthews MA, Kennedy JA. Impact of diurnal temperature variation on grape berry development, proanthocyanidin accumulation, and the expression of flavonoid pathway genes. J Exp Bot. 2012;63:2655–65. doi: 10.1093/jxb/err449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Rienth M, Torregrosa L, Luchaire N, Chatbanyong R, Lecourieux D, et al. Day and night heat stress trigger different transcriptomic responses in green and ripening grapevine (Vitis vinifera) fruit. BMC Plant Biol. 2014;14:108. doi: 10.1186/1471-2229-14-108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Horton P. Crop improvement through alteration in the photosynthetic membrane. Blacksburg: Virginia Tech; 2002. [Google Scholar]
- 55.Heckathorn SA, Downs CA, Sharkey TD, Coleman JS. The small, methionine-rich chloroplast heat-shock protein protects photosystem II electron transport during heat stress. Plant Physiol. 1998;116:439–44. doi: 10.1104/pp.116.1.439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Gustavsson N, Kokke BP, Harndahl U, Silow M, Bechtold U, et al. A peptide methionine sulfoxide reductase highly expressed in photosynthetic tissue in Arabidopsis thaliana can protect the chaperone-like activity of a chloroplast-localized small heat shock protein. Plant J. 2002;29:545–53. doi: 10.1046/j.1365-313X.2002.029005545.x. [DOI] [PubMed] [Google Scholar]
- 57.Zhang T, Wang X, Lu Y, Cai X, Ye Z, et al. Genome-wide analysis of the cyclin gene family in tomato. Int J Mol Sci. 2014;15:120–40. doi: 10.3390/ijms15010120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Hu T, Sun X, Zhang X, Nevo E, Fu J. An RNA sequencing transcriptome analysis of the high-temperature stressed tall fescue reveals novel insights into plant thermotolerance. BMC Genomics. 2014;15:147. doi: 10.1186/1471-2164-15-147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Kitsios G, Doonan JH. Cyclin dependent protein kinases and stress responses in plants. Plant Signal Behav. 2011;6:204–9. doi: 10.4161/psb.6.2.14835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Larkindale J, Vierling E. Core genome responses involved in acclimation to high temperature. Plant Physiol. 2008;146:748–61. doi: 10.1104/pp.107.112060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Irmler S, Schroder G, St-Pierre B, Crouch NP, Hotze M. Indole alkaloid biosynthesis in Catharanthus roseus: new enzyme activities and identification of cytochrome P450CYP72A1 as secologanin synthase. Plant J. 2000;24:797–804. doi: 10.1046/j.1365-313x.2000.00922.x. [DOI] [PubMed] [Google Scholar]
- 62.Naur P, Petersen BL, Mikkelsen MD, Bak S, Rasmussen H, et al. CYP83A1 and CYP83B1, two nonredundant cytochrome P450 enzymes metabolizing oximes in the biosynthesis of glucosinolates in Arabidopsis. Plant Physiol. 2003;133:63–72. doi: 10.1104/pp.102.019240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Takemoto D, Hayashi M, Doke N, Nishimura M, Kawakita K. Molecular cloning of a defense-response-related cytochrome P450 gene from tobacco. Plant Cell Physiol. 1999;40:1232–42. doi: 10.1093/oxfordjournals.pcp.a029511. [DOI] [PubMed] [Google Scholar]
- 64.Vierling E. The roles of heat shock proteins in plants. Annu Rev Plant Physiol Plant Mol Biol. 1991;42:579–620. doi: 10.1146/annurev.pp.42.060191.003051. [DOI] [Google Scholar]
- 65.Mashaghi A, Kramer G, Lamb DC, Mayer MP, Tans SJ. Chaperone action at the single-molecule level. Chem Rev. 2014;114:660–76. doi: 10.1021/cr400326k. [DOI] [PubMed] [Google Scholar]
- 66.Wang W, Vinocur B, Shoseyov O, Altman A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci. 2004;9:244–52. doi: 10.1016/j.tplants.2004.03.006. [DOI] [PubMed] [Google Scholar]
- 67.Waters ER, Lee GJ, Vierling E. Evolution, structure and function of the small heat shock proteins in plants. J Exp Bot. 1996;47:325–38. doi: 10.1093/jxb/47.3.325. [DOI] [Google Scholar]
- 68.Harndahl U, Hall RB, Osteryoung KW, Vierling E, Bornman JF, et al. The chloroplast small heat shock protein undergoes oxidationdependent conformational changes and may protect plants from oxidative stress. Cell Stress Chap. 1999;4:129–38. doi: 10.1379/1466-1268(1999)004<0129:TCSHSP>2.3.CO;2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Malik MK, Slovin JP, Hwang CH, Zimmerman JL. Modified expression of a carrot small heat shock protein gene hsp17.7 results in increased or decreased thermotolerance. Plant J. 1999;20:89–99. doi: 10.1046/j.1365-313X.1999.00581.x. [DOI] [PubMed] [Google Scholar]
- 70.Queitsch C, Hong SW, Vierling E, Lindquist S. Heat shock protein 101 plays a crucial role in thermotolerance in Arabidopsis. Plant Cell. 2000;12:479–92. doi: 10.1105/tpc.12.4.479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Chauhan H, Khurana N, Nijhavan A, Khurana JP, Khurana P. The wheat chloroplastic small heat shock protein (sHSP26) is involved in seed maturation and germination and imparts tolerance to heat stress. Plant Cell Environ. 2012;35:1912–31. doi: 10.1111/j.1365-3040.2012.02525.x. [DOI] [PubMed] [Google Scholar]
- 72.Wang A, Yu X, Mao Y, Liu Y, Liu G, et al. Overexpression of a small heat-shock-protein gene enhances tolerance to abiotic stresses in rice. Plant Breed. 2015;134:384–93. doi: 10.1111/pbr.12289. [DOI] [Google Scholar]
- 73.Downs CA, Ryan SL, Heckathorn SA. The chloroplast small heat-shock protein: evidence for a general role in protecting photosystem II against oxidative stress and photoinhibition. J Plant Physiol. 1999;155:488–96. doi: 10.1016/S0176-1617(99)80043-1. [DOI] [Google Scholar]
- 74.Blanco-Herrera F, Moreno AA, Tapia R, Reyes F, Araya M, et al. The UDP-glucose: glycoprotein glucosyltransferase (UGGT), a key enzyme in ER quality control, plays a significant role in plant growth as well as biotic and abiotic stress in Arabidopsis thaliana. BMC Plant Biol. 2015;15:127. doi: 10.1186/s12870-015-0525-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Taylor SC, Thibault P, Tessier DC, Bergeron JJ, Thomas DY. Glycopeptide specificity of the secretory protein folding sensor UDP-glucose glycoprotein:-glucosyltransferase. EMBO Rep. 2003;4:405–11. doi: 10.1038/sj.embor.embor797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Cyr DM, Höhfeld J, Patterson C. Protein quality control: U-box-containing E3 ubiquitin ligases join the fold. Trends Biochem Sci. 2002;27:368–75. doi: 10.1016/S0968-0004(02)02125-4. [DOI] [PubMed] [Google Scholar]
- 77.Cho SK, Ryu MY, Song C, Kwak JM, Kim WT. Arabidopsis PUB22 and PUB23 are homologous U-box E3 ubiquitin ligases that play combinatory roles in response to drought stress. Plant Cell. 2008;20:1899–1914. doi: 10.1105/tpc.108.060699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Seo DH, Ryu MY, Jammes F, Hwang JH, Turek M, et al. Roles of four Arabidopsis U-box E3 ubiquitin ligases in negative regulation of abscisic acid-mediated drought stress responses. Plant Physiol. 2012;160:556–68. doi: 10.1104/pp.112.202143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Forrest KL, Bhave M. The PIP and TIP aquaporins in wheat form a large and diverse family with unique gene structures and functionally important features. Funct Integr Genomics. 2008;8:115–33. doi: 10.1007/s10142-007-0065-4. [DOI] [PubMed] [Google Scholar]
- 80.Li J, Ban L, Wen H, Wang Z, Dzyubenko N, et al. An aquaporin protein is associated with drought stress tolerance. Biochem Biophys Res Commun. 2015;459:208–13. doi: 10.1016/j.bbrc.2015.02.052. [DOI] [PubMed] [Google Scholar]
- 81.Christou A, Filippou P, Manganaris GA, Fotopoulos V. Sodium hydrosulfide induces systemic thermotolerance to strawberry plants through transcriptional regulation of heat shock proteins and aquaporin. BMC Plant Biol. 2014;14:42. doi: 10.1186/1471-2229-14-42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Zhao CX, Shao HB, Chu LY. Aquaporin structure-function relationships: water flow through plant living cells. Colloid Surf B: Biointerfaces. 2008;62:163–72. doi: 10.1016/j.colsurfb.2007.10.015. [DOI] [PubMed] [Google Scholar]
- 83.Debeaujon I, Peeters AJM, Léon-Kloosterziel KM, Koornneef M. The TRANSPARENT TESTA12 gene of Arabidopsis encodes a multidrug secondary transporter-like protein required for flavonoid sequestration in vacuoles of the seed coat endothelium. Plant Cell. 2001;13:853–71. doi: 10.1105/tpc.13.4.853. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Pirkkala L, Nykaenen P, Sistonen L. Roles of the heat shock transcription factors in regulation of the heat shock response and beyond. FASEB J. 2001;15:1118–31. doi: 10.1096/fj00-0294rev. [DOI] [PubMed] [Google Scholar]
- 85.Nover L, Bharti K, Doring P, Mishra SK, Ganguli A, et al. Arabidopsis and the heat stress transcription factor world: how many heat stress transcription factors do we need? Cell Stress Chap. 2001;6:177–89. doi: 10.1379/1466-1268(2001)006<0177:AATHST>2.0.CO;2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Kotak S, Port M, Ganguli A, Bicker F, von Koskull-Doring P. Characterization of C-terminal domains of Arabidopsis heat stress transcription factors (Hsfs) and identification of a new signature combination of plant class A Hsfs with AHA and NES motifs essential for activator function and intracellular localization. Plant J. 2004;39:98–112. doi: 10.1111/j.1365-313X.2004.02111.x. [DOI] [PubMed] [Google Scholar]
- 87.Liu HC, Liao HT, Charng YY. The role of class A1 heat shock factors (HSFA1s) in response to heat and other stresses in Arabidopsis. Plant Cell Environ. 2011;34:738–51. doi: 10.1111/j.1365-3040.2011.02278.x. [DOI] [PubMed] [Google Scholar]
- 88.Mishra SK, Tripp J, Winkelhaus S, Tschiersch B, Theres K, et al. In the complex family of heat stress transcription factors, HsfA1 has a unique role as master regulator of thermotolerance in tomato. Genes Dev. 2002;16:1555–67. doi: 10.1101/gad.228802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Lohmann C, Eggers-Schumacher G, Wunderlich M, Schoffl F. Two different heat shock transcription factors regulate immediate early expression of stress genes in Arabidopsis. Mol Genet Genomics. 2004;271:11–21. doi: 10.1007/s00438-003-0954-8. [DOI] [PubMed] [Google Scholar]
- 90.Charng YY, Liu HC, Liu NY, Chi WT, Wang CN, et al. A Heat-inducible transcription factor, HsfA2, is required for extension of acquired thermotolerance in Arabidopsis. Plant Physiol. 2007;143:251–62. doi: 10.1104/pp.106.091322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Li C, Chen Q, Gao X, Qi B, Chen N, et al. AtHsfA2 modulates expression of stress responsive genes and enhances tolerance to heat and oxidative stress in Arabidopsis. Sci China C Life Sci. 2005;4:540–50. doi: 10.1360/062005-119. [DOI] [PubMed] [Google Scholar]
- 92.Yoshida T, Sakuma Y, Todaka D, Maruyama K, Qin F, et al. Functional analysis of an Arabidopsis heat-shock transcription factor HsfA3 in the transcriptional cascade downstream of the DREB2A stress-regulatory system. Biochem Biophys Res Commun. 2008;368:515–21. doi: 10.1016/j.bbrc.2008.01.134. [DOI] [PubMed] [Google Scholar]
- 93.Schramm F, Larkindale J, Kiehlmann E, Ganguli A, Englich G, et al. A cascade of transcription factor DREB2A and heat stress transcription factor HsfA3 regulates the heat stress response of Arabidopsis. Plant J. 2008;53:264–74. doi: 10.1111/j.1365-313X.2007.03334.x. [DOI] [PubMed] [Google Scholar]
- 94.Pérez-Salamó I, Papdi C, Rigó G, Zsigmond L, Vilela B, et al. The heat shock factor A4A confers salt tolerance and is regulated by oxidative stress and the mitogen-activated protein kinases MPK3 and MPK6. Plant Physiol. 2014;165:319–34. doi: 10.1104/pp.114.237891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Xu Z-S, Chen M, Li L-C, Ma Y-Z. Functions and application of the AP2/ERF transcription factor family in crop improvement. J Integr Plant Biol. 2011;53:570–85. doi: 10.1111/j.1744-7909.2011.01062.x. [DOI] [PubMed] [Google Scholar]
- 96.Nakano T, Suzuki K, Fujimura T, Shinshi H. Genome-wide analysis of the ERF gene family in Arabidopsis and rice. Plant Physiol. 2006;140:411–32. doi: 10.1104/pp.105.073783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Licausi F, Ohme-Takagi M, Perata P. APETALA2/ethylene responsive factor (AP2/ERF) transcription factors: mediators of stress responses and developmental programs. New Phytol. 2013;199:639–49. doi: 10.1111/nph.12291. [DOI] [PubMed] [Google Scholar]
- 98.Sakuma Y, Maruyama K, Qin F, Osakabe Y, Shinozaki K, et al. Dual function of an Arabidopsis transcription factor DREB2A in water-stress-responsive and heat-stress-responsive gene expression. Proc Natl Acad Sci U S A. 2006;103:18822–27. doi: 10.1073/pnas.0605639103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Mukhtar MS, Deslandes L, Auriac MC, Marco Y, Somssich IE. The Arabidopsis transcription factor WRKY27 influences wilt disease symptom development caused by Ralstonia solanacearum. Plant J. 2008;56:935–47. doi: 10.1111/j.1365-313X.2008.03651.x. [DOI] [PubMed] [Google Scholar]
- 100.Li Y-F, Wang Y, Tang Y, Kakani VG, Mahalingam R. Transcriptome analysis of heat stress response in switchgrass (Panicum virgatum L.) BMC Plant Biol. 2013;13:153. doi: 10.1186/1471-2229-13-153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Li S, Li F, Wang J, Zhang W, Meng Q, et al. Glycinebetaine enhances the tolerance of tomato plants to high temperature during germination of seeds and growth of seedlings. Plant Cell Environ. 2011;34:1931–43. doi: 10.1111/j.1365-3040.2011.02389.x. [DOI] [PubMed] [Google Scholar]
- 102.Li S, Zhou X, Chen L, Huang W, Yu D. Functional characterization of Arabidopsis thaliana WRKY39 in heat stress. Mol Cells. 2010;29:475–83. doi: 10.1007/s10059-010-0059-2. [DOI] [PubMed] [Google Scholar]
- 103.Sabir JSM, Jansen RK, Arasappan D, Calderon V, Noutahi, et al. The nuclear genome of Rhazya stricta and the evolution of alkaloid diversity in a medically relevant clade of Apocynaceae. Sci Rep. 2016;6:33782. [DOI] [PMC free article] [PubMed]
- 104.Langmead B, Trapnell C, Pop M, Salzberg SL. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009;10:R25. doi: 10.1186/gb-2009-10-3-r25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Conesa A, Götz S, García-Gómez JM, Terol J, Talón M, et al. Blast2GO: a universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics. 2005;21:3674–6. doi: 10.1093/bioinformatics/bti610. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The raw reads from the RNA-Seq data were deposited in the short read archive (SRA) of the NCBI (study SRP028238). Permission to collect Rhazya stricta plant material and to perform fieldwork at the site was granted by the Governor of Makkah Province, Prince Khalid Al- Faisal.