

Abstract
Background
Acute systemic inflammatory response syndrome arising from infection can lead to multiple organ failure and death, with greater susceptibility occurring in immunocompromised individuals. Moreover, sub-acute chronic inflammation is a contributor to the pathology of diverse degenerative diseases (Parkinson’s disease, Alzheimer’s disease and arthritis). Given the known limitations in Western medicine to treat a broad range of inflammatory related illness as well as the emergence of antibiotic resistance, there is a renewed interest in complementary and alternative medicines (CAMs) to achieve these means.
Methods
A high throughput (HTP) screening of >1400 commonly sold natural products (bulk herbs, cooking spices, teas, leaves, supplement components, nutraceutical food components, fruit and vegetables, rinds, seeds, polyphenolics etc.) was conducted to elucidate anti-inflammatory substances in lipopolysaccharide (LPS) (E. coli serotype O111:B4) monocytes: RAW 264.7 macrophages [peripheral], BV-2 microglia [brain]) relative to hydrocortisone, dexamethasone and L-N6-(1Iminoethyl)lysine (L-NIL). HTP evaluation was also carried out for lethal kill curves against E.coli 0157:H7 1x106 CFU/mL relative to penicillin. Validation studies were performed to assess cytokine profiling using antibody arrays. Findings were corroborated by independent ELISAs and NO2–/iNOS expression quantified using the Griess Reagent and immunocytochemistry, respectively. For robust screening, we developed an in-vitro efficacy paradigm to ensure anti-inflammatory parameters were observed independent of cytotoxicity. This caution was taken given that many plants exert tumoricidal and anti-inflammatory effects at close range through similar signaling pathways, which could lead to false positives.
Results
The data show that activated BV-2 microglia cells (+ LPS 1μg/ml) release >10-fold greater IL-6, MIP1/2, RANTES and nitric oxide (NO2–), where RAW 264.7 macrophages (+ LPS 1μg/ml) produced > 10-fold rise in sTNFR2, MCP-1, IL-6, GCSF, RANTES and NO2–. Data validation studies establish hydrocortisone and dexamethasone as suppressing multiple pro-inflammatory processes, where L-NIL suppressed NO2–, but had no effect on iNOS expression or IL-6. The screening results demonstrate relative few valid hits with anti-inflammatory effects at < 250μg/ml for the following: Bay Leaf (Laurus nobilis), Elecampagne Root (Inula helenium), Tansy (Tanacetum vulgare),Yerba (Eriodictyon californicum) and Centipeda (Centipeda minima), Ashwagandha (Withania somnifera), Feverfew (Tanacetum parthenium), Rosemary (Rosmarinus officinalis), Turmeric Root (Curcuma Longa), Osha Root (Ligusticum porteri), Green Tea (Camellia sinensis) and constituents: cardamonin, apigenin, quercetin, biochanin A, eupatorin, (-)-epigallocatechin gallate (EGCG) and butein. Natural products lethal against [E. coli 0157:H7] where the LC50 < 100 μg/ml included bioactive silver hydrosol-Argentyn 23, green tea (its constituents EGCG > Polyphenon 60 > (-)-Gallocatechin > Epicatechin > (+)-Catechin), Grapeseed Extract (Vitis vinifera), Chinese Gallnut (its constituents gallic acid > caffeic acid) and gallic acid containing plants such as Babul Chall Bark (Acacia Arabica), Arjun (Terminalia Arjuna) and Bayberry Root Bark (Morella Cerifera).
Conclusions
These findings emphasize and validate the previous work of others and identify the most effective CAM anti-inflammatory, antibacterial compounds using these models. Future work will be required to evaluate potential combination strategies for long-term use to prevent chronic inflammation and possibly lower the risk of sepsis in immunocompromised at risk populations.
Background
Global health initiatives are encumbered by a vast majority of the population suffering from non-communicable inflammatory diseases such as cardiovascular disease, neurodegeneration, diabetes, arthritis, ulcerative colitis/bowel disease and cancer. Also, with increased incidence of antibiotic resistance, acute inflammation from sepsis plays a major role in mortalities arising from diverse infectious agents [1, 2]. Given limitations in Western medicine to treat/prevent a broad range of inflammatory related illness, there is a renewed interest in complementary and alternative medicines (CAMs) to achieve these means [3–9].
While there exist a plethora of scientific publications on the efficacy of individual CAMs in specific inflammatory models, there lacks a relative comparative potency rank of the most commonly marketed CAMs in a single study, conducted under uniform conditions. Our high throughput (HTP) screening library houses over 1400 products most which are available and sold to consumers throughout the world in the form of bulk herbs, cooking spices, teas, leaves, supplement components, nutraceutical food components, fruit and vegetables, roots, rinds, seeds, polyphenolics etc. The purpose of the current study is to screen commonly utilized CAMs for anti-inflammatory efficacy under uniform standard conditions to elucidate the most potent at non-toxic/low therapeutic concentrations (<250 μg/mL), and further to compare these to steroidal and NSAID drugs.
The in-vitro model employed was that of monocytes (peripheral and central nervous system) stimulated by lipopolysaccharide (LPS) derived from E. coli O111:B4. LPS is a cell wall endotoxic component from gram negative bacteria which evokes a deadly cytokine storm associated with septicemia, septic shock and multi organ failure. Known biologic consequences of LPS include the colossal release of chemotactic cytokines, IL-3 IL-12, TNF-alpha, IL-6, IL-1 beta, inducible nitric oxide (iNOS) NO3 -/NO2 -, P-selectin, CD 11b/CD18 (Mac-1) ICAM-1, PGE2 which enable massive neutrophil infiltration and hemolytic [10–13]. While many of these inflammatory molecules at high concentrations are lethal, sub-chronic rises of the same are associated with age related inflammatory degenerative diseases such as Parkinson’s disease, Alzheimer’s disease and arthritis [14–17]. Therefore, the use of LPS in this model and subsequent elucidation of the most effective CAMs against inflammatory parameters, can provide information on potential therapeutics for both chronic and acute inflammatory processes.
In this study, we conduct a HTP screening of CAMs to assess both capacity to kill a pathogenic strain of E.coli 0157:H7 as well as to mitigate the pro-inflammatory effects from E.Coli derived endotoxin cell wall component; LPS.
Methods
Hanks Balanced Salt Solution, (4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid) (HEPES), ethanol, sulfanilamide, 96 well plates, general reagents and supplies, were all purchased from Sigma-Aldrich, (St Louis, MO, USA) or VWR (Radnor, PA, USA). Imaging probes were purchased from Life Technologies (Grand Island, NY, USA). Natural products were purchased from Frontier Natural Products Co-op (Norway, IA, USA), Monterey Bay Spice Company (Watsonville, CA, USA), Mountain Rose Herbs (Eugene, OR, USA), Mayway Traditional Chinese Herbs (Oakland, CA, USA), Kalyx Natural Marketplace (Camden, NY, USA), Futureceuticals (Momence, IL, USA), organic fruit vegetable market: New Leaf (Tallahassee, FL, USA), Florida Food Products Inc. (Eustis, FL, USA), Patel Brothers Indian Grocery (Tampa, FL, USA), Opil Gold from Aging Sciences LLC (Wayland, MA, USA) and Colloidal Silver - Argentyn 23® Natural Immunogenics (Sarasota, FL, USA). Elisa kits and cytokine antibody arrays were purchased from Assay Biotech (Sunnyvale, CA) and Raybiotech (Norcross, GA, USA).
Cell culture
BV-2 microglia (BV-2) cells were provided by Elizabeta Blasi [18], and RAW 264.7 cells were purchased from American Type Culture Collection (Manassas, VA, USA). Cells were cultured in DMEM high glucose media [glucose 4500 mg/L] containing 5% FBS, 4 mM L-glutamine, and penicillin/streptomycin (100 U/0.1 mg/mL). Culture conditions were maintained at 37 °C in 5% CO2/atmosphere and every 2–3 days, the media was replaced and cells sub-cultured. For experiments, plating media consisted of DMEM (minus phenol red) [glucose 4500 mg/L], 2.5% FBS and penicillin/streptomycin (100 U/0.1 mg/mL). LPS O111:B4 was prepared in HBSS at 1 mg/mL and stored at –20 °C. For experiments, LPS was added to the culture media at a working concentration of 1μg/mL.
Bacterial culture
A single colony of E. coli O157:H7 was grown on an agar plate. E. coli was then inoculated into a 20 mL of Luria-Bertani (LB) in a flask, grown at 37 °C with moderate shaking (180 rpm), to an OD 600 nm = 0.6. One mL of the culture suspension was moved into a 1.5 mL Eppendorf tube and centrifuged for 1 min at 10,000 g (4 °C). After discarding the supernatant, the bacterial pellet was re- suspended in 1 mL sterilized water. This centrifugation was repeated twice. The bacteria were stored at 4 °C. The bacterial cell numbers were then determined using colony forming units (CFU) through serial dilution plating on LB plate at 37 °C. The experimental concentration of E. coli was 1 x 106 CFU/mL.
Sample preparation
All natural chemicals and reference drugs were dissolved in DMSO [5–20mg/mL] and crude herbs in absolute ethanol [50 mg/mL] after being diced, macerated and powdered then stored at –20 °C. All plants were cataloged by manufacturer, botanical and common names. All dilutions were prepared in sterile HBSS + 5 mM HEPES, adjusted to a pH of 7.4, ensuring solvent concentration of DMSO or absolute ethanol at less than 0.5%.
Cell and microbial -viability
Cell and microbial viability were assessed using resazurin [7-Hydroxy-3H-phenoxazin-3-one 10-oxide] (Alamar Blue) indicator dye [19]. A working solution of resazurin was prepared in sterile HBSS minus phenol red (0.5 mg/mL), then added (15% v/v) to each sample. Samples were returned to the incubator for 2–4 h, and reduction of the dye by viable cells (to resorufin, a fluorescent compound) was quantitatively analyzed using a Synergy HTX multi-mode reader Bio-Tek Inc. (Winooski, VT, USA) with settings at [550 nm/580 nm], [excitation/emission].
In-Vitro efficacy index
Several methodological concerns were addressed regarding HTP screenings. These included basic controls for pH (neutralized with buffered HBSS) and cell viability. In-vitro, immortal (malignant) immunocompetent cell lines such as glioma cells, macrophages, microglia, lymphocytes or granulocytes are of tumor origin, and many natural compounds simultaneously induce apoptosis in malignant cells and attenuate inflammation via the same pathways (i.e. phosphorylation of extracellular signal-regulated kinase (ERK), c-jun NH2-terminal kinase (JNK) phosphorylation and mitogen-activated protein kinases (MAPK)/NF-κB) [20–26]. For this reason, we constructed and utilized an in-vitro efficacy index (iEI) paradigm, to ensure that anti-inflammatory effects are occurring at non-cytotoxic concentrations. The iEI is defined as the LC50 (toxic concentration)/IC50 (anti-inflammatory concentration) ratio, with higher values reflecting a greater confidence in the anti-inflammatory effects occurring independently of cell death.
Nitrite (NO2–)/iNOS expression
Quantification of nitrite (NO2−) was determined using the Greiss reagent [27]. The Greiss reagent was prepared by mixing an equal volume of 1.0% sulfanilamide in 10% phosphoric acid and 0.1% N-(1-naphthyl)-ethylenediamine in deionized water, which was added directly to the cell supernatant (experimental media consisting of DMEM - phenol red) and incubated at room temperature for 10 min. Controls and blanks were run simultaneously, and subtracted from the final value to eliminate interference. Samples were analyzed at 540 nm on a Synergy HTX multi-mode reader; Bio-Tek (Winooski, VT, USA).
iNOS protein expression was determined using immunocytochemistry. Cells were fixed in 4% paraformaldehyde/permeabilized in 0.2% Triton X 100 in phosphate buffered saline (PBS) and incubated with anti-iNOS, an N-Terminal antibody produced in rabbit for 24 h at 4 °C in a casein blocking buffer. Samples were washed in PBS, then incubated with anti-rabbit Alexa Fluor® 488 conjugate for two hours at RT. Samples were counterstained with propidium iodide and imaged using a fluorescent/inverted microscope, CCD camera and data acquisition using ToupTek View ; ToupTek Photonics Co (Zhejiang, P.R.China).
Mouse cytokine antibody array
Mouse Cytokine Antibody Arrays (Product Code: AAM-CYT-1000) Ray Biotech; (Norcross, GA, USA) were used to profile the effects of LPS (1μg/mL) on BV-2 and RAW 234.7 cell lines. Each experiment was carried out according to the manufacturer’s instructions, and in triplicate. Briefly, antibody-coated array membranes were first incubated for 30 min with 1 mL of blocking buffer. After 30 min, blocking buffer was decanted and replaced with 1 mL supernatant from control (untreated) samples, cells treated with (1ug/mL LPS only) and a media blank. Membranes were incubated overnight at 4 °C with mild shaking. The next day, the medium was decanted; membranes were washed, and subsequently incubated with 1 mL biotin-conjugated antibodies for 6 h. Lastly, biotin-conjugated antibodies were removed and membranes were incubated with HRP-conjugated streptavidin (2h), then evaluated for densitometry using a chemiluminescence substrate monitored on a VersaDoc Imager/Quantity One software from Bio-Rad: (Hercules, CA, USA).
IL-6 (Interleukin-6) ELISA
After experimental treatment, cells supernatants were directly evaluated for concentration of IL-6 using a Murine OmniKine™ IL-6 ELISA (Product Code # OK-0187), Assay Biotechnology Inc. (Sunnyvale, CA, USA), performed according to the manufacturer’s guidelines. Data was quantified by optical density at 450 nm using a Synergy HTX multi-mode reader from Bio-Tek (Winooski, VT, USA).
Data analysis
Statistical analysis was performed using Graph Pad Prism (version 3.0; Graph Pad Software Inc. San Diego, CA, USA) with significance of difference between the groups assessed using a one-way ANOVA then followed by Tukey post hoc means comparison test, or a Student’s t test. IC50s were determined by regression analysis using Origin Software (Origin Lab, Northampton, MA).
Results
Validation
Validation studies were conducted to determine profiled cytokine differentials in LPS activated RAW 264.7 (Fig. 1) and BV-2 cells (Fig. 2), respectively - using semi quantitative antibody microarrays, which were run in triplicate. The representative panel shows both cell lines exposed to LPS prompted the greater release of MCP-1, GCSF, MIP1a, MIP1g and MIP-2, sTNFR1/11, RANTES and IL-6. Quantitative analysis of IL-6 was corroborated by ELISA (Fig. 3), and iNOS protein expression was evaluated by ICC (Fig. 4b) and NO2- release using the Griess Reagent (Fig. 4a), the latter of which was reduced in the presence of iNOS inhibitor (L-NIL).
Fig. 1.
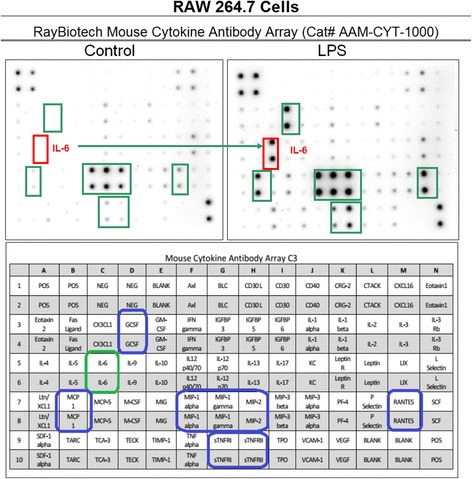
Cytokine release profile in LPS activated RAW 264.7 cells. The data are displayed as the cytokine array blot image and array grid layout with leading changes presented in highlighted boxes
Fig. 2.
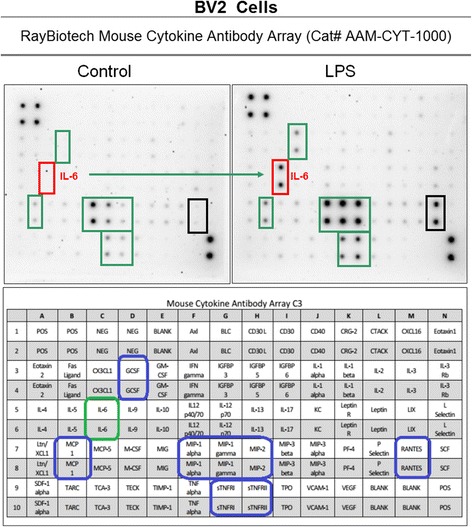
Cytokine release profile in LPS activated BV-2 cells. The data are displayed as the cytokine array blot image and array grid layout with leading changes presented in highlighted boxes
Fig. 3.
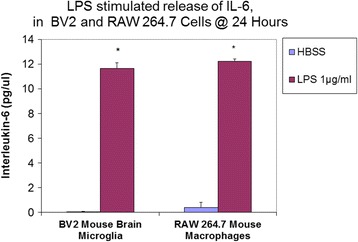
ELISA: Quantification of IL-6 in the supernatant of resting and LPS activated cells (BV-2 cells, RAW 264.7 cells). The data represent IL-6 (pg/μl) and are expressed as the Mean ± S. E. M., n = 3. Differences between resting and LPS activated cells were determined by a student’s T test (*) P < 0.001
Fig. 4.
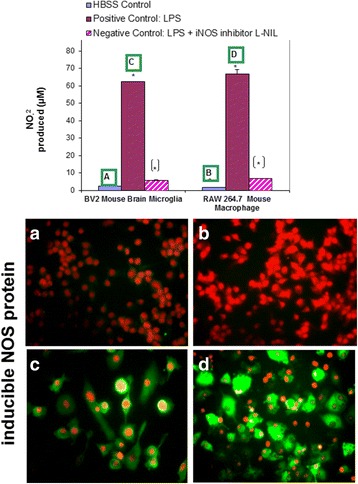
NO2- production in resting and LPS activated cells (BV-2 cells, RAW 264.7 cells) ± selective iNOS inhibitor: L-NIL (12 μg/mL). The data represent NO2- produced (μM) and are expressed as the Mean ± S. E. M., n = 3. Differences between resting and LPS activated cells were determined by a student’s T test (*) P < 0.001. Differences between LPS vs. LPS/L-NIL treated cells were determined by a student’s T test [*] P < 0.001. iNOS expression was analyzed by ICC using rabbit anti-mouse iNOS/goat anti-rabbit Alexafluor 488, in fixed permeabilized, propidium iodide nuclear counterstained cells (a) resting BV-2 cells (b) resting RAW 264.7 cells (c) LPS activated BV-2 cells (d) and RAW 264.7 cells
Screening
The initial HTP screening was conducted using a natural plant library housing over 1400 extracts including: 1) Plants: seeds, fruits, vegetables and herbs (of diverse ethnic nature including Chinese, Egyptian, Indian etc.) 2) Natural derived chemicals/polyphenolics 3) Metabolic Substrates: amino acids, vitamins and energy intermediates such as organic acids, glycolytic intermediates and 4) Reference NSAID and steroidal anti-inflammatory drugs. The preliminary screen was conducted to assess reduction of NO2- in LPS activated cells [equal to or less than 230 μg/mL] for all compounds (Fig. 5a). Substances that attenuated NO2– at less than 50% of the 1st tier starting concentrations, were re-evaluated over a dose range where LC50s (cytotoxicity) and IC50s (NO2-) were simultaneously evaluated (Fig. 5b, c). From the linear regression, LC50, IC50s concentrations were determined and in-vitro efficacy index (iEI) was calculated by the ratio value : LC50/IC50 . The higher the ratio, the greater confidence of true anti-inflammatory effects, not attributable to cell death. All iEI values are presented in Table 1 for RAW 264.7 cells/Table 2 for BV-2 cells, with matching logarithmic scatter-plots (Figs.6 and 7). Figure 8 shows a sample of NO2-/viability dose response data, with corresponding immunochemical imaging for iNOS in RAW 264.7 cells, where supernatant was evaluated for IL–6. The data from these experiments show that L-NIL, while capable of inhibiting the catalytic function of iNOS, was not an anti-inflammatory in the true sense. L-NIL suppressed NO2- but had no effects on cytokine release or expression of iNOS. Most lead compounds that reduced NO2- in both cell lines at sub-lethal concentrations (2 x IC50 for NO2-inhibition) which corresponded to a reduction of IL-6 in sample supernatant (Fig. 9). The antimicrobial effects of natural products on the survival of E.coli 0157:H7 (1x106 CFU/mL) were then evaluated. The data show only a select few have therapeutic potency relative to penicillin/streptomycin (Table 3), colloidal silver being the most effective (Fig. 10). The findings from this study delineate the most potent anti-inflammatory/and antibacterial natural compounds, when conducted in a uniform controlled fashion in these particular models.
Fig. 5.
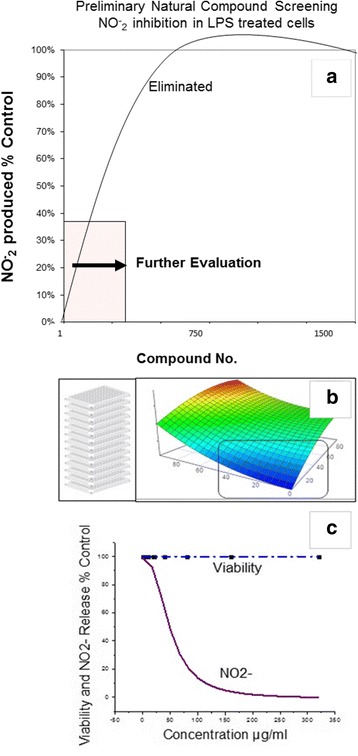
High-through-put study design. The basic study layout consisted of a primary first level tier 1 screening by which all CAMs were tested to reduce NO2- in LPS treated BV-2 and RAW 264.7 cells [maximum working concentrations were : 230μg/mL (plant based) and 92μg/ml (metabolites, drugs and polyphenolics)]. a, b Compounds displaying an IC50 below the 1st tier concentrations were further evaluated as per the template (c). All compounds were simultaneously evaluated for toxicity/anti-inflammatory effects and an iEI differential was established (LC50/IC50) to prevent false positives incurred by cytotoxic effects
Table 1.
Efficacy of anti-inflammatory CAMS relative to cellular toxicity in LPS activated RAW 264.7 cells
| RAW 264.7 cells + 1μ/mL LPS: anti-inflammatory potency | |||
|---|---|---|---|
| Substance | Anti-inflammatory IC50 (μg/mL) | Toxicity LC50 (μg/mL) | iEI (LC50/IC50) |
| L-N-lysine dihydrochloride | 4.4 | 250.0 | >57.4 |
| Cardamonin | 6.1 | 250.0 | >40.8 |
| Dexamethasone | 1.6 | 260.0 | >22.4 |
| Hydrocortisone | 45.6 | 250.0 | >5.4 |
| Bay Leaf/Laurus nobilis | 92.6 | 537.0 | >5.8 |
| Tansy Herb/Tanacetum vulgare | 76.7 | 934.9 | 12.2 |
| Apicidin | 0.2 | 2.4 | 11.7 |
| Apigenin | 30.8 | 252.0 | 8.2 |
| Yerba Santa Lf/E. californicum | 194.0 | 1413.4 | 7.3 |
| Butein | 8.0 | 57.9 | 7.3 |
| Ashwagandha/Withania somnifera | 457.2 | 3306.0 | 7.2 |
| Centipeda Herb/Centipeda minima | 213.7 | 1260.5 | 5.9 |
| Rosemary Lf/Rosmarinus officinalis | 132.4 | 754.8 | 5.7 |
| Feverfew/Tanacetum parthenium | 48.4 | 264.2 | 5.5 |
| Green Tea Std sigma T5550 | 45.5 | 228.7 | 5.0 |
| Elecampane Root/Inula helenium | 257.8 | 1276.0 | 4.9 |
| Quercetin | 14.3 | 63.8 | 4.5 |
| Commiphora myrrha resin | 127.1 | 5221.1 | 4.1 |
| Amla/Phyllanthus emblica | 156.7 | 641.0 | 4.1 |
| Herb de province | 203.4 | 793.2 | 3.9 |
| Turmeric Root/Curcuma longa | 87.3 | 274.3 | 3.1 |
| Biochanin A | 119.0 | 345.1 | 2.9 |
| Trifala | 195.3 | 559.5 | 2.9 |
| Cinnamon/Cinnamomum burmannii | 344.9 | 923.0 | >2.6 |
| EGCG | 20.0 | 50.8 | 2.5 |
| Bergamottin | 67.1 | 161.7 | 2.4 |
| Osha Root/Ligusticum porteri | 43.4 | 104.1 | 2.4 |
| Kalijiri Purple Fleablame | 58.0 | 130.0 | 2.2 |
| Curcumin | 12.6 | 28.1 | 2.2 |
| Rabdosia rubescens Herb | 104.9 | 220.3 | 2.1 |
| White Sage/Salvia apiana | 62.0 | 129.5 | 2.1 |
| Blood Root/Sanguinaria canadensis | 23.4 | 47.4 | 2.0 |
The data represent LC50 values for toxicity and IC50 values for NO2- reduction both determined by regression analysis on a minimum of 6 concentrations,(n = 4). The ratio of LC50/IC50 μg/mL is the iEI (in- vitro efficacy index), where the greater the value the greater the confidence in the anti-inflammatory effects. The symbol [>] denotes an iEI value acquired on a maximum upper limit concentration being tested
Table 2.
Efficacy of natural anti-inflammatory compounds relative to cellular toxicity in LPS activated BV-2 cells
| BV-2 microglia cells + 1μg/mL LPS: anti-inflammatory potency | |||
|---|---|---|---|
| Substance | Anti-inflammatory IC50 (μg/mL) | Toxicity LC50 (μg/mL) | iEI (LC50/IC50) |
| Cardamonin | 1.6 | 265.0 | >169.4 |
| Dexamethasone | 1.9 | 260.0 | >136.8 |
| Bay Leaf/Laurus nobilis | 34.2 | 537.0 | >15.7 |
| Quercetin | 27.8 | 250.0 | >8.9 |
| Apicidin | 0.0 | 0.6 | 65.5 |
| L-N-lysine dihydrochloride | 4.2 | 247.2 | 58.8 |
| Elecampane Root/Inula helenium | 154.4 | 1486.0 | 35.7 |
| Ashwagandha/Withania somnifera | 166.4 | 2848.3 | 17.1 |
| Hydrocortisone | 13.0 | 219.2 | 16.9 |
| Apigenin | 25.2 | 337.0 | 13.4 |
| Optilgold | 9.4 | 113.6 | 12.1 |
| Biochanin A | 33.7 | 369.2 | 10.9 |
| Tansy Herb/Tanacetum vulgare | 143.0 | 1302.1 | 9.1 |
| Feverfew/Tanacetum parthenium | 28.2 | 230.2 | 8.2 |
| Centipeda/Centipeda minima | 258.4 | 2105.2 | 8.1 |
| Osha Root/Ligusticum porteri | 29.5 | 203.4 | 6.9 |
| Eupatorin | 39.3 | 265.2 | 6.7 |
| Turmeric Root/Curcuma longa | 74.4 | 498.9 | 6.7 |
| Herb de province | 167.2 | 1115.3 | 6.7 |
| Granati peel/Punica granatum rind | 75.7 | 439.1 | 5.8 |
| Rabdosia rubescens Herb | 34.5 | 192.7 | 5.6 |
| Rosemary Lf/Rosmarinus officinali | 43.0 | 214.8 | 5.0 |
| Trifala | 88.3 | 408.8 | 4.6 |
| Green Tea Std Sigma T5550 | 38.0 | 169.7 | 4.5 |
| Curcumin | 10.2 | 43.2 | 4.2 |
| Myrrh/Commiphora myrrha | 31.8 | 132.8 | 4.2 |
| Clove/Syzygium aromaticum | 149.0 | 615.1 | 4.1 |
| Indomethacin | 17.9 | 67.2 | 3.7 |
| Sage leaf/Salvia officinalis | 80.3 | 298.4 | 3.7 |
| Amla/Phyllanthus emblica | 206.1 | 736.1 | 3.6 |
| White Sage/Salvia apiana | 79.2 | 282.0 | 3.6 |
| Ganthoda | 168.3 | 563.7 | 3.3 |
| Succinum Resin | 20.1 | 56.3 | 2.8 |
| Genistein | 5.6 | 14.7 | 2.6 |
| Baicalein | 7.8 | 20.1 | 2.6 |
| Butein | 1.2 | 2.9 | 2.3 |
| Maddar root/Rubia tinctorum | 59.2 | 135.0 | 2.3 |
| Yerba Santa Lf/E. californicum | 18.3 | 41.4 | 2.3 |
| EGCG | 11.3 | 24.8 | 2.2 |
| Phloretin | 17.2 | 37.8 | 2.2 |
| Frankincense/Boswellia carterii | 15.4 | 32.5 | 2.1 |
| Fisetin | 2.3 | 4.7 | 2.1 |
| Piperine | 36.2 | 75.1 | 2.1 |
| Javentri Powder | 16.5 | 34.1 | 2.1 |
| Bergamottin | 111.4 | 227.5 | 2.0 |
| Cinnamon/Cinnamomum burmann | 135.0 | 260.0 | <1.92 |
The data represent LC50 values for toxicity and IC50 values for NO2- reduction both determined by regression analysis on a minimum of 6 concentrations,(n = 4). The ratio of LC50/IC50 μg/mL is the iEI (in- vitro efficacy index), where the greater the value the greater the confidence in the anti-inflammatory effects. The symbol [>] denotes an iEI value acquired on a maximum upper limit concentration being tested
Fig. 6.
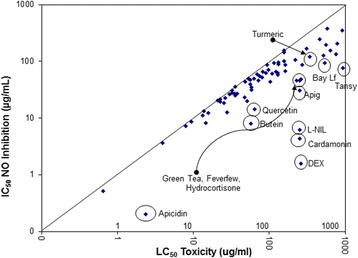
In-vitro efficacy scatter-plot for NO2–, inhibition vs. cell toxicity in RAW 264.7 cells. The data are presented as LC50 (toxicity) vs. IC50 (iNOS inhibition) determined from a regression analysis on a minimum of 6 concentrations
Fig. 7.
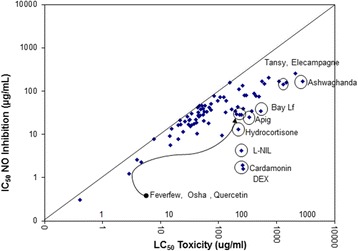
In-vitro efficacy scatter-plot for NO2–, inhibition vs. cell toxicity in BV-2 cells. The data are presented as LC50 (toxicity) vs. IC50 (iNOS inhibition) determined from a regression analysis on a minimum of 6 concentrations
Fig. 8.
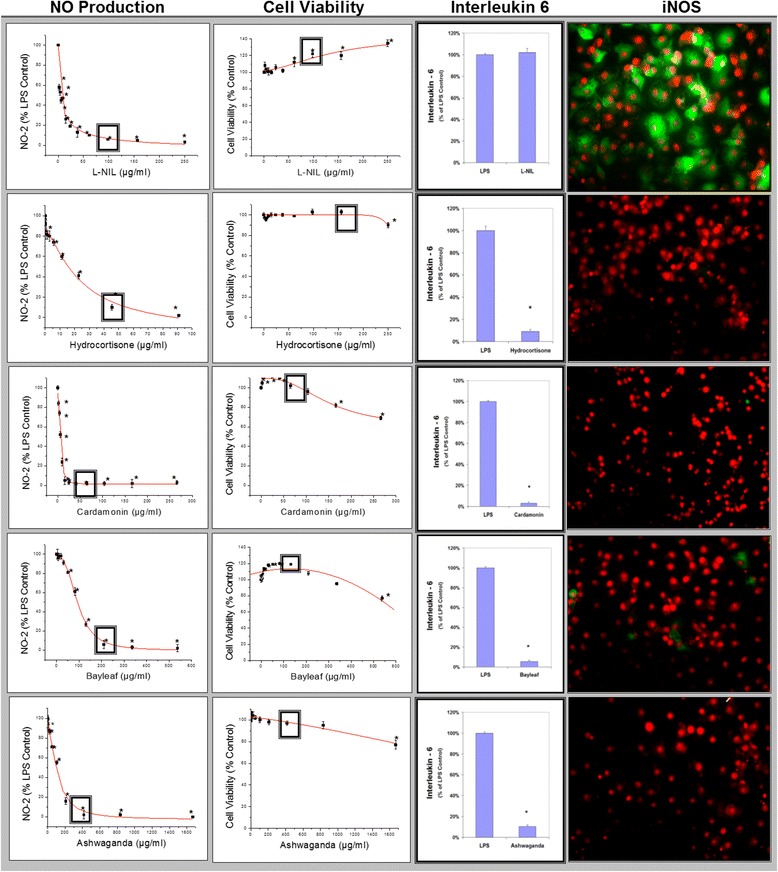
Linear regression profiles for NO2- inhibition and toxicity in RAW 264.7 cells, by which LC50s and IC50s were calculated for all compounds, presented along with corresponding iNOS ICC images and IL-6 release measured at concentrations reflected by a square (◘). The data represent NO2- and viability (% LPS Control), presented as the Mean ± S.E.M, n = 4. Statistical difference from the Controls were determined by a one-way ANOVA, followed by a Tukey post – hoc test * P < 0.05 and IL-6 from controls by a student’s t-test* P < 0.05
Fig. 9.
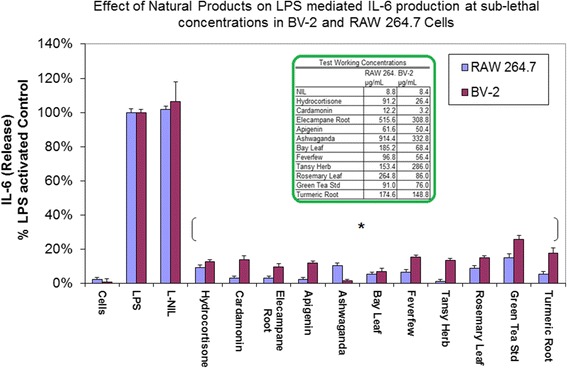
Effects of lead anti-inflammatory products on IL-6 release in LPS treated BV-2 and RAW 264.7 cells. The data represents IL-6 (as % LPS Control) and expressed as the Mean ± S.E.M., n = 3. Differences between activated cells ± natural compounds at sub-lethal dose were determined by a student’s T test (*) P < 0.001
Table 3.
Efficacy of CAM antibacterial compounds on survival of E.coli 0157:H7 (1x106 CFU/ml) @ 8 h in 31 °C
| Anti-bacterial effects of natural compounds on E. coli 0157:H7 survival | ||
|---|---|---|
| Compound | LC50 | Units |
| Penicillin/Streptomycin | 0.01 | Units/μg.ml |
| Cholloidal Silver-Argentym 23® | 0.02 | μg/ml |
| (+) Gossypol | 1.5 | μg/ml |
| Grapeseed Extract/Vitis Vinifera | 1.8 | μg/ml |
| Green Tea Extract/Camellia Sinensis | 2.0 | μg/ml |
| (-)-Epigallocatechin gallate | 3.7 | μg/ml |
| Doxorubin hydrochloride | 5.4 | μg/ml |
| Gallic Acid | 8.4 | μg/ml |
| Caffeic Acid | 10.1 | μg/ml |
| Chinese Gallnut | 10.4 | μg/ml |
| Babul Chall Bark/Acacia arabica | 13.4 | μg/ml |
| Polyphenon 60 | 14.1 | μg/ml |
| Uva Ursi/Arctostaphylos uva ursi | 19.1 | μg/ml |
| Arjun/Terminalia arjuna | 23.8 | μg/ml |
| Balm of Gilead Bud/Populus candicans | 26.7 | μg/ml |
| Bayberry Root/Morella cerifera | 28.5 | μg/ml |
| Blood Root/Sanguinaria canadensis/ | 33.5 | μg/ml |
| (-)-Gallocatechin | 35.8 | μg/ml |
| 2-D08 | 37.0 | μg/ml |
| Glyoxal Acid | 39.6 | μg/ml |
| Scutellarian | 42.0 | μg/ml |
| Kokum Black/Garcinia Indica | 47.5 | μg/ml |
| Indole | 47.5 | μg/ml |
| Trifala | 47.7 | μg/ml |
| CraneSbill Root/Geranium maculatum | 51.6 | μg/ml |
| Shi Liu Pi (Granati Peel) | 60.4 | μg/ml |
| Catuaba Bark/Trichilia Catigua | 63.4 | μg/ml |
| Chapparal/Larrea tridentata | 73.6 | μg/ml |
| Thymol | 92.0 | μg/ml |
| Esculetin | 93.3 | μg/ml |
| Epicatechin | 114.0 | μg/ml |
| Piperonal | 115.1 | μg/ml |
| (+)-Catechin | 173.0 | μg/ml |
The data represents LC50 values determined by regression analysis, on a minimum of 6 concentrations, (n = 4)
Fig. 10.
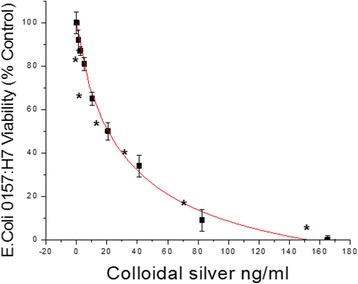
The effect of colloidal Silver-Argentyn 23® on the survival of E.coli 0157:H7 (1x106 CFU/ml) @ 8 h in 31°C. The data represent viability (% Control), presented as the Mean ± S.E.M, n = 4. Statistical differences from the Control were determined by a one-way ANOVA, using the Tukey post – hoc test. * P < 0.05
Discussion
The data from this study establish several findings including [A] uncorroborated anti-inflammatory/antimicrobial effects for over a thousand natural compounds at concentrations less than 230 μg/mL using this model; [B] corroborating data of existing work by other research groups regarding anti-inflammatory effects of green tea, curcumin, turmeric and rosemary ; and antimicrobial properties of green tea, its catechins, Chinese gallnut, gallic acid plant derived anti-fungal agents (cotton/gossypol) or silver nanoparticle dispersions [28–37]. Lastly; [C] this work provides new evidence on some lesser acknowledged herbs to which historical medicinal value has been attributed, but little research has been documented. Some of these include the following.
Elecampane ( Inula helenium) (IH) has extensive historical medicinal value, where its use dates back to the Iron Age (c. 800–450 B.C.) throughout the third century B.C. to 79 A.D. also mentioned by Pliny and further corroborated in the Chilandar Medical Codex (13th or 14th centuries A.D.) [38]. Within the last century, scientific documentation is somewhat sparse on this herb having primarily focused on its ability to cause allergic dermatitis or act as an anti-cancer agent attributable to the content of alantolactone and isoalantolactone [39–44]. Meager work has been performed investigating the effect of IH on sepsis or age relate chronic inflammatory conditions. Although meager research has been conducted in IH, the findings presented here are in alignment with existing researchers who have reported its capacity to attenuate iNOS/NO2-, COX-2/PGE2, HMGB1 release and NF-κB in LPS-activated RAW 264.7 cells or phorbol activated T cells [45–48]. Interestingly, although we did not find IH to have significant antimicrobial effects on E.coli 0157:H7 (1x106 CFU/mL) at the low concentrations criteria used in this study, others have reported its capacity to destroy invasive pathogens such as Staphylococcus aureus/methicillin-resistant (MRSA) gram-positive bacteria, yeasts parasites and Mycobacterium tuberculosis [42, 49–53]. These studies suggest IH as being somewhat promising for attenuating inflammation arising from diverse infective or inflammatory insults.
The data from this work also show that fresh dried ethanol extracts of Bay leaf (BL) ( Laurus nobilis ) contains anti-inflammatory properties [54, 55]. Previous work by others demonstrates the oil (not aqueous) extracts to contain antimicrobial/food preserving properties due to cineole, eugenol, pinene, eucalyptol, linalool, carvacrol and α-terpinenyl acetate all evidentially toxic to Gram-positive bacteria (Staphylococcus aureus/pyogenes) and fungi (Candida albicans, Aspergillus fumigatus) [56–59]. Again, regarding the aqueous extract of BL, our work corroborates the work of others having reported the capacity to attenuate LPS mediated microglia/macrophage activation thought to be attributable to its sesquiterpene content [60, 61]. These type of substances are thought to be beneficial in chronic age related degeneration, by not only reducing inflammation but also blocking neurotoxicity of AD pathological Aβ (25-35)-component fragments [62].
Another herbal extract used in the current study to which little data exists is Centipeda minima (CM). CM has previously been reported to contain high levels of helenalin with the capacity to LPS mediated elevation of NO2–,TNF-α, IL-1b, iNOS and cyclooxygenase-2 in macrophages [63]. CM also demonstrates the capacity to attenuate tissue injury in-vivo involving inflammation such as carrageen paw edema and liver fibrosis [63–65]. Although we did not find CM to have significant antimicrobial effects on E.coli 0157:H7 (1 x 106 CFU/mL) at the low concentration criteria used in this study, others have reported its capacity to kill Enterobacter aerogenes, Staphylococcus aureus, Yersinia enterocolitica and Bacillus subtilis [66, 67].
Feverfew (Tanacetum parthenium) (TP) is another rarely evaluated herb which long been reported to treat inflammatory conditions including psoriasis, allergies, arthritis, asthma and particularly migraines [68]. TP derived sesquiterpene lactones such as parthenolide are believed to be responsible for observed anti-inflammatory effects in animal models of carrageenan-induced edema, osteoarthritis, colitis cystic fibrosis and phorbol triggered mouse-ear edema [69–73]. TP constituents also antagonize toll-like receptors, Akt/mTOR and NF-κB pathways and block the downstream release of cytokines [74, 75]. Like the present study, previous research reports also corroborate capacity to reduce LPS activation of BV-2 cells and RAW 264.7 cells alike [76, 77]. There is also an antinociceptive aspect to feverfew commonly reported, having benefit to ameliorate pain associated with diabetic peripheral neuropathy [78].
The use of tansy (Tanacetum vulgare) (TV) as a medicinal plant was reported dating back to the 8th century A.D., when the Benedictine monks used it to treat intestinal worms, ameliorate digestive problems, fevers and sores. Interestingly, both feverfew and tansy have in common hyper allergenic potential due to parthenolide [79, 80]. Tansy is also rich in flavonoid glycosides, 7-O-glucosides of apigenin, luteolin, scutellarein and 6- hydroxyluteolin, chrysoeriol and eriodictyol as well as aglycones, hispidulin, nepetin, eupatilin, jaceosidin, pectolinarigenin and axillarin [81]. The oil contains 1,8-cineole and β-thujone as a major constituent along with carveol, umbellulone, davanone, dihydrocarvone, chrysanthenol, borneol and myrtenol [82–85].
Ashwagandha (Withania somnifera) (WS) is a highly studied herb with a plethora of known health benefits, in particular for prevention of cardiovascular disease. Its primary bioactive compound (Withaferin A) directly inhibits β1-adrenergic receptors, HMG-CoA, angiotensinogen-converting enzyme, total cholesterol, triglycerides, low density lipoprotein and elevation of protective high density lipoproteins and endogenous antioxidant systems [86, 87]. In animal models, WS prevents isoproterenol induced myocardial infarction, stroke distal middle cerebral artery occlusion and monocrotaline induced pulmonary hypertension in rats [88–91]. With respect to the immune system, WS can attenuate mitogen induced T/B-cell activation, secretion of Th1 and Th2 cytokines and inhibit NF-κB nuclear translocation in lymphocytes [92]. These immunomodulating effects of WS are also reported in-vitro for systemic LPS or E. coli administration in mammals, where there is a reduction in neutrophil tissue infiltration [93, 94] as well as tissue damage and pain associated with rheumatoid arthritis [95–97]. Ashwagandha is also an anti-infective agent lethal to gram-positive bacteria/cocci such as methicillin resistant Staphylococcus aureus and Enterococcus, respectively [98]. The data in this study ranks, validates and confirms pre-existing research showing significant antimicrobial effects of green tea EGCG polyphenon-60 (PP-60) Acacia arabica,grapeseed extract, caffeic, gallic acid, chapparal (Larrea tridentata) [99–109], where little has been investigated on antimicrobial herbs such as balm of Gilead Bud (Populus candicans), an herb of great historical significance with observed antibacterial and anti-inflammatory properties.
Conclusion
In conclusion, the data obtained in this work affords general information on validated CAM anti-inflammatory and antimicrobial compounds and relative potency at sub lethal concentrations in LPS activated BV-2 and RAW 264.7 cells. Moreover, the data obtained also provide relative lethal potency of CAMs against the growth of E.coli 0157:H7. These findings can serve as a guide for future examination of specific CAM based herbal/nutraceutical anti-inflammatory/antimicrobial modalities for use in prevention or treatment of disease.
Acknowledgments
We wish to thank undergraduate students for sample prep and diverse project tasks: Ms. Swan and Marquis Cromartie, as well as the STEM Summer Students- Leandria Harvey and Gabrielle Bradley and their mentor Dr. Tiffany Ardley.
Funding
This research was supported by the National Institute of Minority Health and Health Disparities of the National Institutes of Health through Grant Number 8 G12MD007582-28 and Grant Number 1P20 MD006738-01.
Availability of data and materials
Materials and data of this study are available to other researchers upon request.
Authors’ contributions
EAM was responsible for overview, planning, carrying out basic natural product screenings, confirmation of ELISAS, imaging and manuscript preparation. NL, MD, HW and LT were responsible for planning, overseeing and carrying out the microbial HTP screenings, DB, PM and ET were responsible for cytokine profiling and KFAS was in charge of project planning, overview and manuscript preparation. All authors read and approved the final manuscript.
Competing interest
The authors declare that they have no competing interests.
Consent for publication
NA.
Ethics approval and consent to participate
NA.
Abbreviations
- ANOVA
One-way analysis of variance
- CAMs
Complementary and alternative medicines
- CM
Centipeda minima
- CNS
Central nervous system
- COX2
Cyclooxygenase-2
- DMEM
Dulbecco’s modified eagle medium
- EGCG
(-)-epigallocatechin gallate
- ELISA
Enzyme-linked immunosorbent assay
- GCSF
Granulocyte-colony stimulating factor
- HMGB1
High mobility group box 1 protein
- HTP
High throughput
- iEI
in-vitro efficacy index
- IH
Inula helenium
- INOS
Inducible nitric oxide
- LPS
Lipopolysaccharide
- MSRA
Methicillin-resistant
- NO2-
Nitrite
- NSAID
Non-steroidal anti-inflammatory drugs
- PBS
phosphate buffered saline
- RANTES
Regulated on activation, normal T cell expressed and secreted
- TP
Tanacetum parthenium (Feverfew)
- TV
Tanacetum vulgare (Tansy)
- WS
Withania somnifera (Ashwagandha)
References
- 1.Hostinar CE, Ross KM, Chen E, Miller GE. Modeling the association between lifecourse socioeconomic disadvantage and systemic inflammation in healthy adults: The role of self-control. Health Psychol. 2015;34(6):580–590. doi: 10.1037/hea0000130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Hebert JR, Braun KL, Kaholokula JK, Armstead CA, Burch JB, Thompson B. Considering the Role of Stress in Populations of High-Risk, Underserved Community Networks Program Centers. Prog Community Health Partnersh. 2015;9(Suppl):71–82. doi: 10.1353/cpr.2015.0028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Yang YH, Rajaiah R, Lee DY, Ma Z, Yu H, Fong HH, Lao L, Berman BM, Moudgil KD. Suppression of ongoing experimental arthritis by a chinese herbal formula (huo-luo-xiao-ling dan) involves changes in antigen-induced immunological and biochemical mediators of inflammation. Evid Based Complement Alternat Med. 2011;2011:642027. doi: 10.1155/2011/642027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Vojdani A, Lambert J, Kellermann G. The Role of Th17 in Neuroimmune Disorders: A Target for CAM Therapy. Part III. Evid Based Complement Alternat Med. 2011;2011:548086. doi: 10.1093/ecam/nep064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Kes VB, Cesarik M, Matovina LZ, Zavoreo I, Coric L, Drnasin S, Demarin V. The role of complementary and alternative medicine in therapy of multiple sclerosis. Acta Clin Croat. 2013;52(4):464–471. [PubMed] [Google Scholar]
- 6.Lissat A, Joerschke M, Shinde DA, Braunschweig T, Meier A, Makowska A, Bortnick R, Henneke P, Herget G, Gorr TA, et al. IL6 secreted by Ewing sarcoma tumor microenvironment confers anti-apoptotic and cell-disseminating paracrine responses in Ewing sarcoma cells. BMC Cancer. 2015;15:552. doi: 10.1186/s12885-015-1564-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Yao H, Chen Y, Zhang L, He X, Lian L, Wu X, Lan P. Carnosol inhibits cell adhesion molecules and chemokine expression by tumor necrosis factor-alpha in human umbilical vein endothelial cells through the nuclear factor-kappaB and mitogen-activated protein kinase pathways. Mol Med Rep. 2014;9(2):476–480. doi: 10.3892/mmr.2013.1839. [DOI] [PubMed] [Google Scholar]
- 8.Salaga M, Zatorski H, Sobczak M, Chen C, Fichna J. Chinese herbal medicines in the treatment of IBD and colorectal cancer: a review. Curr Treat Options Oncol. 2014;15(3):405–420. doi: 10.1007/s11864-014-0288-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kwak S, Ku SK, Bae JS. Fisetin inhibits high-glucose-induced vascular inflammation in vitro and in vivo. Inflamm Res. 2014;63(9):779–787. doi: 10.1007/s00011-014-0750-4. [DOI] [PubMed] [Google Scholar]
- 10.Weber GF, Chousterman BG, He S, Fenn AM, Nairz M, Anzai A, Brenner T, Uhle F, Iwamoto Y, Robbins CS, et al. Interleukin-3 amplifies acute inflammation and is a potential therapeutic target in sepsis. Science. 2015;347(6227):1260–1265. doi: 10.1126/science.aaa4268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Martin I, Caban-Hernandez K, Figueroa-Santiago O, Espino AM. Fasciola hepatica fatty acid binding protein inhibits TLR4 activation and suppresses the inflammatory cytokines induced by lipopolysaccharide in vitro and in vivo. J Immunol. 2015;194(8):3924–3936. doi: 10.4049/jimmunol.1401182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lee SK, Park YJ, Ko MJ, Wang Z, Lee HY, Choi YW, Bae YS. A novel natural compound from garlic (Allium sativum L.) with therapeutic effects against experimental polymicrobial sepsis. Biochem Biophys Res Commun. 2015;464(3):774–779. doi: 10.1016/j.bbrc.2015.07.031. [DOI] [PubMed] [Google Scholar]
- 13.Barichello T, Generoso JS, Silvestre C, Costa CS, Carrodore MM, Cipriano AL, Michelon CM, Petronilho F, Dal-Pizzol F, Vilela MC, et al. Circulating concentrations, cerebral output of the CINC-1 and blood-brain barrier disruption in Wistar rats after pneumococcal meningitis induction. Eur J Clin Microbiol Infect Dis. 2012;31(8):2005–2009. doi: 10.1007/s10096-011-1533-2. [DOI] [PubMed] [Google Scholar]
- 14.Bessler H, Djaldetti R, Salman H, Bergman M, Djaldetti M. IL-1 beta, IL-2, IL-6 and TNF-alpha production by peripheral blood mononuclear cells from patients with Parkinson’s disease. Biomed Pharmacother. 1999;53(3):141–145. doi: 10.1016/S0753-3322(99)80079-1. [DOI] [PubMed] [Google Scholar]
- 15.Blum-Degen D, Muller T, Kuhn W, Gerlach M, Przuntek H, Riederer P. Interleukin-1 beta and interleukin-6 are elevated in the cerebrospinal fluid of Alzheimer’s and de novo Parkinson’s disease patients. Neurosci Lett. 1995;202(1-2):17–20. doi: 10.1016/0304-3940(95)12192-7. [DOI] [PubMed] [Google Scholar]
- 16.Brodacki B, Staszewski J, Toczylowska B, Kozlowska E, Drela N, Chalimoniuk M, Stepien A. Serum interleukin (IL-2, IL-10, IL-6, IL-4), TNFalpha, and INFgamma concentrations are elevated in patients with atypical and idiopathic parkinsonism. Neurosci Lett. 2008;441(2):158–162. doi: 10.1016/j.neulet.2008.06.040. [DOI] [PubMed] [Google Scholar]
- 17.Zhu L, Wang J, Wei T, Gao J, He H, Chang X, Yan T. Effects of Naringenin on inflammation in complete freund’s adjuvant-induced arthritis by regulating Bax/Bcl-2 balance. Inflammation. 2015;38(1):245–251. doi: 10.1007/s10753-014-0027-7. [DOI] [PubMed] [Google Scholar]
- 18.Blasi E, Barluzzi R, Bocchini V, Mazzolla R, Bistoni F. Immortalization of murine microglial cells by a v-raf/v-myc carrying retrovirus. J Neuroimmunol. 1990;27(2-3):229–237. doi: 10.1016/0165-5728(90)90073-V. [DOI] [PubMed] [Google Scholar]
- 19.Evans SM, Casartelli A, Herreros E, Minnick DT, Day C, George E, Westmoreland C. Development of a high throughput in vitro toxicity screen predictive of high acute in vivo toxic potential. Toxicol In Vitro. 2001;15(4-5):579–584. doi: 10.1016/S0887-2333(01)00064-9. [DOI] [PubMed] [Google Scholar]
- 20.Zhang X, Luo W, Zhao W, Lu J, Chen X. Isocryptotanshinone Induced Apoptosis and Activated MAPK Signaling in Human Breast Cancer MCF-7 Cells. J Breast Cancer. 2015;18(2):112–118. doi: 10.4048/jbc.2015.18.2.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Huang WC, Hsu RM, Chi LM, Leu YL, Chang YS, Yu JS. Selective downregulation of EGF receptor and downstream MAPK pathway in human cancer cell lines by active components partially purified from the seeds of Livistona chinensis R. Brown Cancer Lett. 2007;248(1):137–146. doi: 10.1016/j.canlet.2006.06.010. [DOI] [PubMed] [Google Scholar]
- 22.Amirghofran Z. Herbal medicines for immunosuppression. Iran J Allergy Asthma Immunol. 2012;11(2):111–119. [PubMed] [Google Scholar]
- 23.Pan HC, Jiang Q, Yu Y, Mei JP, Cui YK, Zhao WJ. Quercetin promotes cell apoptosis and inhibits the expression of MMP-9 and fibronectin via the AKT and ERK signalling pathways in human glioma cells. Neurochem Int. 2015;80:60–71. doi: 10.1016/j.neuint.2014.12.001. [DOI] [PubMed] [Google Scholar]
- 24.Cheng YW, Chang CY, Lin KL, Hu CM, Lin CH, Kang JJ. Shikonin derivatives inhibited LPS-induced NOS in RAW 264.7 cells via downregulation of MAPK/NF-kappaB signaling. J Ethnopharmacol. 2008;120(2):264–271. doi: 10.1016/j.jep.2008.09.002. [DOI] [PubMed] [Google Scholar]
- 25.Chen CC, Tsai PC, Wei BL, Chiou WF. 8-Prenylkaempferol suppresses inducible nitric oxide synthase expression through interfering with JNK-mediated AP-1 pathway in murine macrophages. Eur J Pharmacol. 2008;590(1-3):430–436. doi: 10.1016/j.ejphar.2008.05.018. [DOI] [PubMed] [Google Scholar]
- 26.Ci X, Liang X, Luo G, Yu Q, Li H, Wang D, Li R, Deng X. Regulation of inflammatory mediators in lipopolysaccharide-stimulated RAW 264.7 cells by 2”-hydroxy-3”-en-anhydroicaritin involves down-regulation of NF-kappaB and MAPK expression. Int Immunopharmacol. 2010;10(9):995–1002. doi: 10.1016/j.intimp.2010.04.015. [DOI] [PubMed] [Google Scholar]
- 27.Cendan JC, Topping DL, Pruitt J, Snowdy S, Copeland EM, 3rd, Lind DS. Inflammatory mediators stimulate arginine transport and arginine-derived nitric oxide production in a murine breast cancer cell line. J Surg Res. 1996;60(2):284–288. doi: 10.1006/jsre.1996.0044. [DOI] [PubMed] [Google Scholar]
- 28.Yang F, de Villiers WJ, McClain CJ, Varilek GW. Green tea polyphenols block endotoxin-induced tumor necrosis factor-production and lethality in a murine model. J Nutr. 1998;128(12):2334–2340. doi: 10.1093/jn/128.12.2334. [DOI] [PubMed] [Google Scholar]
- 29.Chan MM, Fong D, Ho CT, Huang HI. Inhibition of inducible nitric oxide synthase gene expression and enzyme activity by epigallocatechin gallate, a natural product from green tea. Biochem Pharmacol. 1997;54(12):1281–1286. doi: 10.1016/S0006-2952(97)00504-2. [DOI] [PubMed] [Google Scholar]
- 30.Senthil Kumaran V, Arulmathi K, Sundarapandiyan R, Kalaiselvi P. Attenuation of the inflammatory changes and lipid anomalies by epigallocatechin-3-gallate in hypercholesterolemic diet fed aged rats. Exp Gerontol. 2009;44(12):745–751. doi: 10.1016/j.exger.2009.08.010. [DOI] [PubMed] [Google Scholar]
- 31.Bengmark S, Mesa MD, Gil A. Plant-derived health: the effects of turmeric and curcuminoids. Nutr Hosp. 2009;24(3):273–281. [PubMed] [Google Scholar]
- 32.Al-Okbi SY. Nutraceuticals of anti-inflammatory activity as complementary therapy for rheumatoid arthritis. Toxicol Ind Health. 2014;30(8):738–749. doi: 10.1177/0748233712462468. [DOI] [PubMed] [Google Scholar]
- 33.Liang W, Fernandes AP, Holmgren A, Li X, Zhong L. Bacterial thioredoxin and thioredoxin reductase as mediators for epigallocatechin 3-gallate-induced antimicrobial action. FEBS J. 2016;283(3):446–458. doi: 10.1111/febs.13587. [DOI] [PubMed] [Google Scholar]
- 34.Noormandi A, Dabaghzadeh F. Effects of green tea on Escherichia coli as a uropathogen. J Tradit Complement Med. 2015;5(1):15–20. doi: 10.1016/j.jtcme.2014.10.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Shao D, Li J, Li J, Tang R, Liu L, Shi J, Huang Q, Yang H. Inhibition of Gallic Acid on the Growth and Biofilm Formation of Escherichia coli and Streptococcus mutans. J Food Sci. 2015;80(6):M1299–M1305. doi: 10.1111/1750-3841.12902. [DOI] [PubMed] [Google Scholar]
- 36.Chen XY, Chen Y, Heinstein P, Davisson VJ. Cloning, expression, and characterization of (+)-delta-cadinene synthase: a catalyst for cotton phytoalexin biosynthesis. Arch Biochem Biophys. 1995;324(2):255–266. doi: 10.1006/abbi.1995.0038. [DOI] [PubMed] [Google Scholar]
- 37.He M, Lu L, Zhang J, Li D. Immobilized Silver Nanoparticles on Chitosan with Special Surface State-Enhanced Antimicrobial Efficacy and Reduced Cytotoxicity. J Nanosci Nanotechnol. 2015;15(9):6435–6443. doi: 10.1166/jnn.2015.10782. [DOI] [PubMed] [Google Scholar]
- 38.Al-Gammal SY. Elecampane and Job’s disease. Bull Indian Inst Hist Med Hyderabad. 1998;28(1):7–11. [PubMed] [Google Scholar]
- 39.P’Iankova ZP. Nugmanova ML: [Dermatitis caused by elecampane] Vestn Dermatol Venerol. 1975;12:52–54. [PubMed] [Google Scholar]
- 40.Wang J, Zhao YM, Zhang B, Guo CY. Protective Effect of Total Phenolic Compounds from Inula helenium on Hydrogen Peroxide-induced Oxidative Stress in SH-SY5Y Cells. Indian J Pharm Sci. 2015;77(2):163–169. doi: 10.4103/0250-474X.156553. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Zhao YM, Wang J, Liu HB, Guo CY, Zhang WM. Microwave-assisted Extraction of Alantolactone and Isoalantolactone from Inula helenium. Indian J Pharm Sci. 2015;77(1):116–120. doi: 10.4103/0250-474X.151594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Cantrell CL, Abate L, Fronczek FR, Franzblau SG, Quijano L, Fischer NH. Antimycobacterial eudesmanolides from Inula helenium and Rudbeckia subtomentosa. Planta Med. 1999;65(4):351–355. doi: 10.1055/s-1999-14001. [DOI] [PubMed] [Google Scholar]
- 43.Rasul A, Khan M, Ali M, Li J, Li X. Targeting apoptosis pathways in cancer with alantolactone and isoalantolactone. ScientificWorldJournal. 2013;2013:248532. doi: 10.1155/2013/248532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Yang C, Yang J, Sun M, Yan J, Meng X, Ma T. Alantolactone inhibits growth of K562/adriamycin cells by downregulating Bcr/Abl and P-glycoprotein expression. IUBMB Life. 2013;65(5):435–444. doi: 10.1002/iub.1141. [DOI] [PubMed] [Google Scholar]
- 45.Cabronero MC, Campos E, Picazo JJ, Romero J. Evaluation of an agglutination method of latex particles sensitized for the detection of vaginal candidiasis. Med Clin (Barc) 1990;94(9):329–332. [PubMed] [Google Scholar]
- 46.Nesterova Iu V, Zelenskaia KL, Vetoshkina TV, Aksinenko SG, Gorbacheva AV, Gorbatykh NA. Mechanisms of antistressor activity of Inula helenium preparations. Eksp Klin Farmakol. 2003;66(4):63–65. [PubMed] [Google Scholar]
- 47.Danilets MG, Belska NV, Bel’sky YP, Uchasova EG, Trophimova ES, Ligatcheva AA, Guriev AM, Belousov MV, Ahmedganov RR, Usubov MS, et al. Effects of plant water-soluble polysaccharides on the production of immunoglobulins E and G1 by lymphocytes of mice sensitized with ovalbumin. Bull Exp Biol Med. 2008;146(5):585–587. doi: 10.1007/s10517-009-0351-3. [DOI] [PubMed] [Google Scholar]
- 48.Na BR, Kim HR, Kwon MS, Lee HS, Piragyte I, Choi EJ, Choi HK, Han WC, Lee SH, Jun CD. Aplotaxene blocks T cell activation by modulation of protein kinase C-theta-dependent pathway. Food Chem Toxicol. 2013;62:23–31. doi: 10.1016/j.fct.2013.08.016. [DOI] [PubMed] [Google Scholar]
- 49.Radulovic NS, Denic MS, Stojanovic-Radic ZZ. Synthesis of small combinatorial libraries of natural products: identification and quantification of new long-chain 3-methyl-2-alkanones from the root essential oil of Inula helenium L. (Asteraceae) Phytochem Anal. 2014;25(1):75–80. doi: 10.1002/pca.2466. [DOI] [PubMed] [Google Scholar]
- 50.Gokbulut A, Ozhan O, Satilmis B, Batcioglu K, Gunal S, Sarer E. Antioxidant and antimicrobial activities, and phenolic compounds of selected Inula species from Turkey. Nat Prod Commun. 2013;8(4):475–478. [PubMed] [Google Scholar]
- 51.O’Shea S, Lucey B, Cotter L. In vitro activity of Inula helenium against clinical Staphylococcus aureus strains including MRSA. Br J Biomed Sci. 2009;66(4):186–189. doi: 10.1080/09674845.2009.11730271. [DOI] [PubMed] [Google Scholar]
- 52.El Garhy MF, Mahmoud LH. Anthelminthic efficacy of traditional herbs on Ascaris lumbricoides. J Egypt Soc Parasitol. 2002;32(3):893–900. [PubMed] [Google Scholar]
- 53.Cantrell CL, Fischer NH, Urbatsch L, McGuire MS, Franzblau SG. Antimycobacterial crude plant extracts from South, Central, and North America. Phytomedicine. 1998;5(2):137–145. doi: 10.1016/S0944-7113(98)80011-1. [DOI] [PubMed] [Google Scholar]
- 54.Kilic A, Hafizoglu H, Kollmannsberger H, Nitz S. Volatile constituents and key odorants in leaves, buds, flowers, and fruits of Laurus nobilis L. J Agric Food Chem. 2004;52(6):1601–1606. doi: 10.1021/jf0306237. [DOI] [PubMed] [Google Scholar]
- 55.Yahyaa M, Matsuba Y, Brandt W, Doron-Faigenboim A, Bar E, McClain A, Davidovich-Rikanati R, Lewinsohn E, Pichersky E, Ibdah M. Identification, Functional Characterization, and Evolution of Terpene Synthases from a Basal Dicot. Plant Physiol. 2015;169(3):1683–1697. doi: 10.1104/pp.15.00930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Marzouki H, Khaldi A, Marongiu B, Piras A, Harzallah-Skhiri F. Chemical polymorphism of essential oils from populations of Laurus nobilis grown on Tunisia. Algeria and France Nat Prod Commun. 2011;6(10):1483–1486. [PubMed] [Google Scholar]
- 57.Ramos C, Teixeira B, Batista I, Matos O, Serrano C, Neng NR, Nogueira JM, Nunes ML, Marques A. Antioxidant and antibacterial activity of essential oil and extracts of bay laurel Laurus nobilis Linnaeus (Lauraceae) from Portugal. Nat Prod Res. 2012;26(6):518–529. doi: 10.1080/14786419.2010.531478. [DOI] [PubMed] [Google Scholar]
- 58.Fukuyama N, Ino C, Suzuki Y, Kobayashi N, Hamamoto H, Sekimizu K, Orihara Y. Antimicrobial sesquiterpenoids from Laurus nobilis L. Nat Prod Res. 2011;25(14):1295–1303. doi: 10.1080/14786419.2010.502532. [DOI] [PubMed] [Google Scholar]
- 59.Liu MH, Otsuka N, Noyori K, Shiota S, Ogawa W, Kuroda T, Hatano T, Tsuchiya T. Synergistic effect of kaempferol glycosides purified from Laurus nobilis and fluoroquinolones on methicillin-resistant Staphylococcus aureus. Biol Pharm Bull. 2009;32(3):489–492. doi: 10.1248/bpb.32.489. [DOI] [PubMed] [Google Scholar]
- 60.Chen H, Xie C, Wang H, Jin DQ, Li S, Wang M, Ren Q, Xu J, Ohizumi Y, Guo Y. Sesquiterpenes inhibiting the microglial activation from Laurus nobilis. J Agric Food Chem. 2014;62(20):4784–4788. doi: 10.1021/jf501515v. [DOI] [PubMed] [Google Scholar]
- 61.De Marino S, Borbone N, Zollo F, Ianaro A, Di Meglio P, Iorizzi M. New sesquiterpene lactones from Laurus nobilis leaves as inhibitors of nitric oxide production. Planta Med. 2005;71(8):706–710. doi: 10.1055/s-2005-864191. [DOI] [PubMed] [Google Scholar]
- 62.Pacifico S, Gallicchio M, Lorenz P, Duckstein SM, Potenza N, Galasso S, Marciano S, Fiorentino A, Stintzing FC, Monaco P. Neuroprotective potential of Laurus nobilis antioxidant polyphenol-enriched leaf extracts. Chem Res Toxicol. 2014;27(4):611–626. doi: 10.1021/tx5000415. [DOI] [PubMed] [Google Scholar]
- 63.Huang SS, Chiu CS, Lin TH, Lee MM, Lee CY, Chang SJ, Hou WC, Huang GJ, Deng JS. Antioxidant and anti-inflammatory activities of aqueous extract of Centipeda minima. J Ethnopharmacol. 2013;147(2):395–405. doi: 10.1016/j.jep.2013.03.025. [DOI] [PubMed] [Google Scholar]
- 64.Qin RA, Mei X, Wan L, Shi JL, Shen YJ. Effects of the volatile oil of Centipeda minima on acute pleural effusion in rats induced by an intrapleural injection of car. Zhongguo Zhong Yao Za Zhi. 2005;30(15):1192–1194. [PubMed] [Google Scholar]
- 65.Lin X, Zhang S, Huang R, Wei L, Tan S, Liang S, Tian Y, Wu X, Lu Z, Huang Q. Helenalin attenuates alcohol-induced hepatic fibrosis by enhancing ethanol metabolism, inhibiting oxidative stress and suppressing HSC activation. Fitoterapia. 2014;95:203–213. doi: 10.1016/j.fitote.2014.03.020. [DOI] [PubMed] [Google Scholar]
- 66.Soetardjo S, Jr, Jong PC, Ahmad MN, Lachimanan YL, Sreenivasan S. Chemical Composition and Biological Activity of the Centipeda minima (Asteraceae) Malays J Nutr. 2007;13(1):81–87. [PubMed] [Google Scholar]
- 67.Taylor RS, Towers GH. Antibacterial constituents of the Nepalese medicinal herb, Centipeda minima. Phytochemistry. 1998;47(4):631–634. doi: 10.1016/S0031-9422(97)00534-7. [DOI] [PubMed] [Google Scholar]
- 68.Murphy JJ, Heptinstall S, Mitchell JR. Randomised double-blind placebo-controlled trial of feverfew in migraine prevention. Lancet. 1988;2(8604):189–192. doi: 10.1016/S0140-6736(88)92289-1. [DOI] [PubMed] [Google Scholar]
- 69.Zhao Y, Chen SJ, Wang JC, Niu HX, Jia QQ, Chen XW, Du XY, Lu L, Huang B, Zhang Q, et al. Sesquiterpene lactones inhibit advanced oxidation protein product-induced MCP-1 expression in podocytes via an IKK/NF-kappaB-dependent mechanism. Oxid Med Cell Longev. 2015;2015:934058. doi: 10.1155/2015/934058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Di Cesare ML, Tenci B, Zanardelli M, Maidecchi A, Lugli A, Mattoli L, Ghelardini C. Widespread pain reliever profile of a flower extract of Tanacetum parthenium. Phytomedicine. 2015;22(7-8):752–758. doi: 10.1016/j.phymed.2015.05.006. [DOI] [PubMed] [Google Scholar]
- 71.Zhao ZJ, Xiang JY, Liu L, Huang XL, Gan HT. Parthenolide, an inhibitor of the nuclear factor-kappaB pathway, ameliorates dextran sulfate sodium-induced colitis in mice. Int Immunopharmacol. 2012;12(1):169–174. doi: 10.1016/j.intimp.2011.11.007. [DOI] [PubMed] [Google Scholar]
- 72.Saadane A, Masters S, DiDonato J, Li J, Berger M. Parthenolide inhibits IkappaB kinase, NF-kappaB activation, and inflammatory response in cystic fibrosis cells and mice. Am J Respir Cell Mol Biol. 2007;36(6):728–736. doi: 10.1165/rcmb.2006-0323OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Schinella GR, Giner RM, Recio MC. Mordujovich de Buschiazzo P, Rios JL, Manez S: Anti-inflammatory effects of South American Tanacetum vulgare. J Pharm Pharmacol. 1998;50(9):1069–1074. doi: 10.1111/j.2042-7158.1998.tb06924.x. [DOI] [PubMed] [Google Scholar]
- 74.Nam YJ, da Lee H, Lee MS, Lee CS. Sesquiterpene lactone parthenolide attenuates production of inflammatory mediators by suppressing the Toll-like receptor-4-mediated activation of the Akt, mTOR, and NF-kappaB pathways. Naunyn Schmiedebergs Arch Pharmacol. 2015;388(9):921–930. doi: 10.1007/s00210-015-1132-3. [DOI] [PubMed] [Google Scholar]
- 75.Park SJ, Shin HJ, Youn HS. Parthenolide inhibits TRIF-dependent signaling pathway of Toll-like receptors in RAW264.7 macrophages. Mol Cells. 2011;31(3):261–265. doi: 10.1007/s10059-011-0032-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Magni P, Ruscica M, Dozio E, Rizzi E, Beretta G, Maffei Facino R. Parthenolide inhibits the LPS-induced secretion of IL-6 and TNF-alpha and NF-kappaB nuclear translocation in BV-2 microglia. Phytother Res. 2012;26(9):1405–1409. doi: 10.1002/ptr.3732. [DOI] [PubMed] [Google Scholar]
- 77.Hwang D, Fischer NH, Jang BC, Tak H, Kim JK, Lee W. Inhibition of the expression of inducible cyclooxygenase and proinflammatory cytokines by sesquiterpene lactones in macrophages correlates with the inhibition of MAP kinases. Biochem Biophys Res Commun. 1996;226(3):810–818. doi: 10.1006/bbrc.1996.1433. [DOI] [PubMed] [Google Scholar]
- 78.Galeotti N, Maidecchi A, Mattoli L, Burico M, Ghelardini C. St. John’s Wort seed and feverfew flower extracts relieve painful diabetic neuropathy in a rat model of diabetes. Fitoterapia. 2014;92:23–33. doi: 10.1016/j.fitote.2013.10.003. [DOI] [PubMed] [Google Scholar]
- 79.Salapovic H, Geier J, Reznicek G. Quantification of Sesquiterpene Lactones in Asteraceae Plant Extracts: Evaluation of their Allergenic Potential. Sci Pharm. 2013;81(3):807–818. doi: 10.3797/scipharm.1306-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Jovanovic M, Poljacki M, Duran V, Vujanovic L, Sente R, Stojanovic S. Contact allergy to Compositae plants in patients with atopic dermatitis. Med Pregl. 2004;57(5-6):209–218. doi: 10.2298/MPNS0406209J. [DOI] [PubMed] [Google Scholar]
- 81.Uehara A, Akiyama S, Iwashina T. Foliar flavonoids from Tanacetum vulgare var. boreale and their geographical variation. Nat Prod Commun. 2015;10(3):403–405. [PubMed] [Google Scholar]
- 82.Piras A, Falconieri D, Bagdonaite E, Maxia A, Goncalves MJ, Cavaleiro C, Salgueiro L, Porcedda S. Chemical composition and antifungal activity of supercritical extract and essential oil of Tanacetum vulgare growing wild in Lithuania. Nat Prod Res. 2014;28(21):1906–1909. doi: 10.1080/14786419.2014.939085. [DOI] [PubMed] [Google Scholar]
- 83.Godinho LS, de Carvalho LS A, de Castro CC B, Dias MM, Pinto Pde F, Crotti AE, Pinto PL, de Moraes J, Da Silva Filho AA. Anthelmintic activity of crude extract and essential oil of Tanacetum vulgare (Asteraceae) against adult worms of Schistosoma mansoni. Sci World J. 2014;2014:460342. doi: 10.1155/2014/460342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Formisano C, Senatore F, Bruno M, Rosselli S, Bellone G, Spadaro V. Essential oil composition of Tanacetum vulgare subsp. siculum (Guss.) Raimondo et Spadaro (Asteraceae) from Sicily. Nat Prod Commun. 2009;4(4):567–570. [PubMed] [Google Scholar]
- 85.Hethelyi E, Tetenyi P, Dabi E, Danos B. The role of mass spectrometry in medicinal plant research. Biomed Environ Mass Spectrom. 1987;14(11):627–632. doi: 10.1002/bms.1200141110. [DOI] [PubMed] [Google Scholar]
- 86.Ravindran R, Sharma N, Roy S, Thakur AR, Subhadra G, Sriram K, Devi J, Rajkumar J. Interaction studies of Withania somnifera’s key metabolite Withaferin A with different receptors associated with cardiovascular disease. Curr Comput Aided Drug Des. 2015. [DOI] [PubMed]
- 87.Shukla K, Dikshit P, Shukla R, Sharma S, Gambhir JK. Hypolipidemic and antioxidant activity of aqueous extract of fruit of Withania coagulans (Stocks) Dunal in cholesterol-fed hyperlipidemic rabbit model. Indian J Exp Biol. 2014;52(9):870–875. [PubMed] [Google Scholar]
- 88.Khalil MI, Ahmmed I, Ahmed R, Tanvir EM, Afroz R, Paul S, Gan SH, Alam N. Amelioration of Isoproterenol-Induced Oxidative Damage in Rat Myocardium by Withania somnifera Leaf Extract. Biomed Res Int. 2015;2015:624159. doi: 10.1155/2015/624159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Ahmad H, Khandelwal K, Samuel SS, Tripathi S, Mitra K, Sangwan RS, Shukla R, Dwivedi AK. Neuro-protective potential of a vesicular system of a standardized extract of a new chemotype of Withania somnifera Dunal (NMITLI118RT+) against cerebral stroke in rats. Drug Deliv. 2015;1–12. [DOI] [PubMed]
- 90.Raghavan A, Shah ZA. Withania somnifera Improves Ischemic Stroke Outcomes by Attenuating PARP1-AIF-Mediated Caspase-Independent Apoptosis. Mol Neurobiol. 2015;52(3):1093–1105. doi: 10.1007/s12035-014-8907-2. [DOI] [PubMed] [Google Scholar]
- 91.Kaur G, Singh N, Samuel SS, Bora HK, Sharma S, Pachauri SD, Dwivedi AK, Siddiqui HH, Hanif K. Withania somnifera shows a protective effect in monocrotaline-induced pulmonary hypertension. Pharm Biol. 2015;53(1):147–157. doi: 10.3109/13880209.2014.912240. [DOI] [PubMed] [Google Scholar]
- 92.Gambhir L, Checker R, Sharma D, Thoh M, Patil A, Degani M, Gota V, Sandur SK. Thiol dependent NF-kappaB suppression and inhibition of T-cell mediated adaptive immune responses by a naturally occurring steroidal lactone Withaferin A. Toxicol Appl Pharmacol. 2015;289(2):297–312. doi: 10.1016/j.taap.2015.09.014. [DOI] [PubMed] [Google Scholar]
- 93.Gao S, Li H, Zhou XQ, You JB, Tu DN, Xia G, Jiang JX, Xin C. Withaferin A attenuates lipopolysaccharide-induced acute lung injury in neonatal rats. Cell Mol Biol (Noisy-le-grand) 2015;61(3):102–106. [PubMed] [Google Scholar]
- 94.El-Boshy Mel S, Abdalla OM, Risha A, Moustafa F. Effect of Withania somnifera Extracts on Some Selective Biochemical, Hematological, and Immunological Parameters in Guinea Pigs Experimental Infected with E. coli. ISRN Vet Sci. 2013;2013:153427. doi: 10.1155/2013/153427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Kumar G, Srivastava A, Sharma SK, Rao TD, Gupta YK. Efficacy & safety evaluation of Ayurvedic treatment (Ashwagandha powder & Sidh Makardhwaj) in rheumatoid arthritis patients: a pilot prospective study. Indian J Med Res. 2015;141(1):100–106. doi: 10.4103/0971-5916.154476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Khan MA, Subramaneyaan M, Arora VK, Banerjee BD, Ahmed RS. Effect of Withania somnifera (Ashwagandha) root extract on amelioration of oxidative stress and autoantibodies production in collagen-induced arthritic rats. J Complement Integr Med. 2015;12(2):117–125. doi: 10.1515/jcim-2014-0075. [DOI] [PubMed] [Google Scholar]
- 97.Gupta A, Singh S. Evaluation of anti-inflammatory effect of Withania somnifera root on collagen-induced arthritis in rats. Pharm Biol. 2014;52(3):308–320. doi: 10.3109/13880209.2013.835325. [DOI] [PubMed] [Google Scholar]
- 98.Bisht P, Rawat V. Antibacterial activity of Withania somnifera against Gram-positive isolates from pus samples. Ayu. 2014;35(3):330–332. doi: 10.4103/0974-8520.153757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Zakir M, Sultan KB, Khan H, Ihsaanullah, Khan MA, Fazal H, Rauf A. Antimicrobial activity of different tea varieties available in Pakistan. Pak J Pharm Sci. 2015;28(6):2091–2094. [PubMed] [Google Scholar]
- 100.Klancnik A, Piskernik S, Bucar F, Vuckovic D, Mozina SS, Jersek B. Reduction of microbiological risk in minced meat by a combination of natural antimicrobials. J Sci Food Agric. 2014;94(13):2758–2765. doi: 10.1002/jsfa.6621. [DOI] [PubMed] [Google Scholar]
- 101.Gupta S, Bansal R, Ali J, Gabrani R, Dang S. Development and characterization of polyphenon 60 and caffeine microemulsion for enhanced antibacterial activity. Biomed Res Int. 2014;2014:932017. doi: 10.1155/2014/932017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Tambekar DH, Khante BS, Chandak BR, Titare AS, Boralkar SS, Aghadte SN. Screening of antibacterial potentials of some medicinal plants from Melghat forest in India. Afr J Tradit Complement Altern Med. 2009;6(3):228–232. doi: 10.4314/ajtcam.v6i3.57158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Ahn J, Grun IU, Mustapha A. Antimicrobial and antioxidant activities of natural extracts in vitro and in ground beef. J Food Prot. 2004;67(1):148–155. doi: 10.4315/0362-028x-67.1.148. [DOI] [PubMed] [Google Scholar]
- 104.Vandal J, Abou-Zaid MM, Ferroni G, Leduc LG. Antimicrobial activity of natural products from the flora of Northern Ontario. Canada Pharm Biol. 2015;53(6):800–806. doi: 10.3109/13880209.2014.942867. [DOI] [PubMed] [Google Scholar]
- 105.Favela-Hernandez JM, Garcia A, Garza-Gonzalez E, Rivas-Galindo VM, Camacho-Corona MR. Antibacterial and antimycobacterial lignans and flavonoids from Larrea tridentata. Phytother Res. 2012;26(12):1957–1960. doi: 10.1002/ptr.4660. [DOI] [PubMed] [Google Scholar]
- 106.Pinho E, Ferreira IC, Barros L, Carvalho AM, Soares G, Henriques M. Antibacterial potential of northeastern Portugal wild plant extracts and respective phenolic compounds. Biomed Res Int. 2014;2014:814590. doi: 10.1155/2014/814590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Lopez-Nicolas R, Gonzalez-Bermudez CA, Ros-Berruezo G, Frontela-Saseta C. Influence of in vitro gastrointestinal digestion of fruit juices enriched with pine bark extract on intestinal microflora. Food Chem. 2014;157:14–19. doi: 10.1016/j.foodchem.2014.01.126. [DOI] [PubMed] [Google Scholar]
- 108.Li Z, Summanen PH, Komoriya T, Henning SM, Lee RP, Carlson E, Heber D, Finegold SM. Pomegranate ellagitannins stimulate growth of gut bacteria in vitro: Implications for prebiotic and metabolic effects. Anaerobe. 2015;34:164–168. doi: 10.1016/j.anaerobe.2015.05.012. [DOI] [PubMed] [Google Scholar]
- 109.Valéria NL, Oliveira-Tintino CD, Santos ES, et al. Antimicrobial and enhancement of the antibiotic activity by phenolic compounds: Gallic acid, caffeic acid and pyrogallol. Microb Pathog. 2016;99:56–61. [DOI] [PubMed]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Materials and data of this study are available to other researchers upon request.