

Abstract
Objective: The aim of this study was to investigate the occurrence and molecular characterization of extended-spectrum β-lactamases (ESBL), plasmid-mediated AmpC β-lactamase (pAmpC) and carbapenemases as well as plasmid-mediated quinolone-resistant (PMQR) among extended-spectrum cephalosporin-resistant (ESC-R) Escherichia coli from dogs in Shaanxi province in China.
Methods: A total of 40 ESC-R Escherichia coli selected from 165 Extraintestinal pathogenic E. coli (ExPEC) isolated from dogs were screened and characterized for the genes encoding for the ESBLs, pAmpC, carbapenemases and PMQR genes by PCR and sequencing. Phylogenetic groups, virulence gene profiles and multilocus sequence typing (MLST) were used to investigate the genetic background of the ESC-R E. coli isolates.
Results: Among 40 ESC-R E. coli, the predominant β-lactamase gene was blaCTX−Ms (n = 35), and followed by blaTEM−1 (n = 31), blaSHV−12 (n = 14), blaOXA−48 (n = 8), blaTEM−30 (n = 4), blaCMY−2 (n = 3) and blaDHA−1 (n = 2). The most common specific blaCTX−M gene subtype was blaCTX−M−15 (n = 31), and followed by blaCTX−M−123 (n = 14), blaCTX−M−1 (n = 10), blaCTX−M−14 (n = 10) and blaCTX−M−9 (n = 7). PMQR genes were detected in 32 (80%) isolates, and the predominant PMQR gene was aac(6′)-Ib-cr (n = 26), followed by qnrS (n = 12), qnrD (n = 9), qnrB (n = 8), qepA (n = 4), and all PMQR genes were detected in co-existence with β-lactamase genes. traT (n = 34) and fimH (n = 32) were the most prevalent virulence genes, and virulence genes fimH, iutA, fyuA, malX, iha, and sat were more prevalent in phylogenetic group B2. The 40 ESC-R isolates analyzed were assigned to 22 sequence types (STs), and the clonal lineages ST131 (n = 10) and ST10 (n = 9) were the predominant STs.
Conclusion: High prevalence of β-lantamases and PMQR genes were detected among ESC-R E. coli from companion animals. This is also the first description of the co-existence of six β-lantamase genes and five PMQR genes in one E. coli isolate. Moreover, 10 ST131 clones harboring CTX-M-15 were detected.
Keywords: Eshcerichia coli, β-lactamases, PMQR, cephalosporin resistance, dog
Introduction
Escherichia coli is one of the most predominant organisms causing infections in human and animals. The β-lactams, particularly the third-generation cephalosporins are important for the treatment of serious infections in companion animals caused by Enterobacteriaceae (Jiang et al., 2012). However, there has been an increasing number of infections worldwide due to the third-generation cephalosporin resistant E. coli isolates worldwide, and companion animals can service as a reservoir of cephalosporin-resistant bacteria as the physical closeness between humans and their pet companions (Costa et al., 2009; Wasyl et al., 2012). One of the currently most important resistance mechanisms in E. coli is based on plasmid mediated production of enzymes, especially the extended-spectrum β-lactamases (ESBL), which inactivate β-lactams by hydrolyzing their β-lactam rings (Zhao and Hu, 2013). Additionally, plasmid-mediated AmpC β-lactamase (pAmpC) blaCMY−2, carbapenemases blaNDM−1, blaOXA−48 and blaKPC−2 are now also increasingly described (Nordmann et al., 2011). β-lactamase genes are usually located on mobile genetic elements, such as plasmids, transposons or integrons. Resistant plasmids are transferred between bacterial isolates or different bacterial species by conjugation mechanism (Cantas et al., 2015). Furthermore, extended-spectrum cephalosporin-resistant (ESC-R) E. coli isolates are often cross-resistant to fluoroquinolones, sulfamethoxazole/trimethoprim or other antimicrobials, and finally expressed multidrug resistance (MDR) (Tian et al., 2009, 2012).
The emergence of plasmid-mediated quinolone resistance (PMQR) indicated that quinolone resistance can also be acquired through horizontal gene transfer, and PMQR genes can co-exist on the same plasmid with genes encoding ESBLs and to be co-transferred to recipients (Briales et al., 2012; Yu et al., 2015). Moreover, PMQR genes can create an environment in E. coli for the rapid selection of high level resistance, although they usually just confer the lower-level resistance (Ishida et al., 2010).
Although, the ESBLs and other β-lactamases in E. coli isolates from humans and food-animals have been characterized in various studies in China (Yuan et al., 2009; Sun et al., 2010; Wang et al., 2012; Xi et al., 2015; Xu et al., 2015; Yu et al., 2015), very little data has been reported on the occurrence of β-lactamases and PMQR as well as the population structure in ExPEC isolates from companion animals in Shaanxi province, even in China. This prompted us to determine and characterize the β-lactamases (ESBLs, pAmpC, and carbapenemases) and PMQR genes in ESC-R E. coli collected from dogs in Shaanxi province, and shed light on the phylogenetic groups, virulence gene profiles and the multilocus sequence typing (MLST).
Materials and methods
Bacterial isolates
A collection of 165 unique ExPEC isolates were isolated from urine, wound, genital tract, anal sac, nasal structure and soft tissue samples of dogs with naturally-occurring infection in the small animal hospitals in Shaanxi province, China from March 2013 to October 2015. Samples were collected from individual dog using a sterile swab and transported to the laboratory within 12 h. Samples were immediately seeded on MacConkey agar at 37°C, and one colony with typical E. coli morphology was selected from each sample. E. coli were identified using API-20E (BioMerieux, Beijing, China). All confirmed E. coli isolates were stored at −80°C in the Tryptic Soy broth medium containing 30% glycerol for further study.
Antimicrobial susceptibility testing
Antimicrobial susceptibility testing was performed for all 165 isolates. The minimum inhibitory concentrations (MICs) of third-generation cephalosporins (ceftiofur, cefotaxime, cefoperazone, cefpodoxime, ceftazidime), fluoroquinolones (enrofloxacin, ciprofloxacin, and pradofloxacin) and other β-lactams (ampicillin, amoxicillin-clavulanic acid, cefoxitin, doxycycline, imipenem, meropenem, sulfamethoxazole/trimethoprim, gentamicin, and amikacin) were determined by a standardized microdilution method followed the CLSI guidelines (CLSI, 2013). All MIC determinations were performed in triplicates and E. coli ATCC 25922 was used for quality control.
Additionally, all the 165 E. coli isolates were screened for ESBL production using microdilution-based Sensititre (TREK diagnostic systems, Cleveland, Ohio) with ESBL Confirmatory MIC plates (ESB1F) as described previously (Aly et al., 2012).
Phylogenetic typing and virulence genotyping
Genomic DNA were extracted from bacterial preparations using the PreMan® Ultra Preparation Reagent according to manufacturer's protocol. The pathogenicity of E. coli is associated with the presence of virulence genes that can be encoded by chromosomal or plasmid genes, and thus each isolate was examined for the presence of 19 virulence genes (Table S1) known for their association with ExPEC by use of established PCR assays. This panel of genes represent five categories: adhesins genes including fimH, sfa/focDE, afa/draBC, iha, papA, papC, papG, and papG alleles (I, II, III); toxin genes including hlyA, cnf1 and sat; capsule gene including kpsM II; siderophore genes includinge fyuA and iutA; the miscellaneous virulence genes including traT and malX.
Identification of β-lactamase genes and plasmid-mediated quinolone resistance genes
The occurrence of ESBLs (TEM, SHV, CTX-Ms), pAmpC (CMY-2, DHA-1, FOX, ACC, and EBC) and carbapenemase genes (class B, NDM-1; class A, KPC-2; class D, OXA-48) in ESC-R E. coli were determined by PCR using specific primers (Table S1) and conditions previously described (Perez-Perez and Hanson, 2002; Yan et al., 2004; Poirel et al., 2011; Shaheen et al., 2011). Meanwhile, all ESC-R E. coli isolates were characterized by PCR for PMQR genes (qnrA, qnrB, qnrD, qnrS, aac(6′)-Ib-cr, and qepA) as described previously (Liu et al., 2012). All PCR products from ESBL and PMQR genes were confirmed and analyzed by DNA sequencing.
Multilocus sequence typing (MLST)
MLST was performed using seven conserved housekeeping genes of E. coli (adk, fumC, gyrB, icd, mdh, purA, and recA). A detailed scheme of the MLST procedure is available at MLST databases at the at the Warwick University website (http://mlst.warwick.ac.uk/mlst/dbs/Ecoli).
Pulsed-field gel electrophoresis (PFGE) analysis
In order to determine the genetic relatedness of the ESC-R E. coli, the chromosomal DNA of some selected isolates were digested with the restriction enzyme XbaI and then subjected to PFGE analysis according to the PulseNet protocol of the US Centers for Disease Control and Prevention (http://www.cdc.gov/pulsenet/protocols.htm) and previous literature (Tenover et al., 1995). The strain of Salmonella serotype Braenderup restricted with XbaI was used as molecular weight standard.
Conjugation experiments
The transferability of β-lactamase and PMQR genes was performed by mating on 10 randomly selected ESC-R E. coli isolates with azide-resistant E. coli J53 (J53 AZr) as described previously (Loncaric et al., 2013). Transconjugants were selected on Mueller-Hinton (MH) agar supplemented with 150 μg/ml sodium azide and 2 μg/ml cefotaxime. Antimicrobial susceptibility, confirmatory tests for ESBL production, and PCR detection were performed on all transconjugants to confirm transfer of relevant β-lactamase and PMQR genes.
Results
Antimicrobial susceptibility of E. coli isolates
Among the 165 E. coli isolates surveyed, 40 (24.2%) isolates were both ESC-R and ESBL-producing isolates, which exhibited high resistance rate to cefotaxime (100%), ceftazidime (87.5%), ceftoxitin (62.5%), cefoperazone (52.5%), and also exhibited high resistance rate to amoxicillin-clavulanic acid (90%). The frequencies of resistance to non-β-lactam antibiotics were as follows: doxycycline, 95%; enrofloxacin, 92.5%; sulfamethoxazole/trimethoprim, 77.5%; chloramphenicol, 70%; gentamicin, 60%; pradofloxacin, 55%. Additionally, it was noteworthy that 10 (25%) isolates were resistance to imipenem and meropenem, respectively. The resistance rates to other antibacterial agents tested were lower than 40%, and all ESC-R E. coli isolates exhibited the MDR phenotype
Phylogenetic typing and virulence genotyping
Phylogenetic group analysis revealed that the predominant phylogenetic group was A (37.5%) and B2 (35%), followed by phylogenetic groups D (12.5%), E (7.5%), B1 (5%), and F (2.5%) among 40 ESC-R isolates tested (Table 1). Eighteen of 19 investigated virulence genes were detected, and the overall prevalence of the virulence genes ranged from 7.5% (papG) to 85% (traT). fimH, iutA, fyuA, and malX were more prevalent in phylogenetic group B2, but less prevalent in group A isolates (P < 0.001), and iha and sat exclusively present in group B2 (Table 1).
Table 1.
The occurrence of virulence genes in ESC-R E. coli isolates.
| Isolate | PG | fimH | papA | papC | papG | papGIII | afa/draBC | Sfa/focDE | hlyA | cnf1 | iutA | fyuA | traT | ibeA | malX | sat | iha | iroN | kpsM II |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| B1 | A | + | + | ||||||||||||||||
| H2 | A | + | + | + | |||||||||||||||
| W12 | A | + | + | ||||||||||||||||
| B6 | A | + | + | + | + | ||||||||||||||
| Y5 | A | + | + | + | + | + | |||||||||||||
| W4 | A | + | + | + | |||||||||||||||
| X2 | A | + | + | + | + | + | |||||||||||||
| Y15 | A | + | + | + | + | + | + | + | + | + | |||||||||
| B3 | A | + | + | + | + | ||||||||||||||
| W15 | A | + | + | ||||||||||||||||
| W6 | A | + | + | + | |||||||||||||||
| Y13 | A | + | + | + | |||||||||||||||
| X1 | A | + | + | ||||||||||||||||
| B5 | A | + | + | + | + | + | + | ||||||||||||
| H1 | A | + | |||||||||||||||||
| X5 | B1 | + | + | + | |||||||||||||||
| X6 | B1 | + | + | + | + | ||||||||||||||
| Y10 | B2 | + | + | + | |||||||||||||||
| X8 | B2 | + | + | + | + | + | + | + | + | + | + | + | + | ||||||
| W19 | B2 | + | + | + | + | + | + | + | + | ||||||||||
| W9 | B2 | + | + | + | + | + | + | + | + | + | |||||||||
| Y8 | B2 | + | + | + | + | + | + | + | + | + | |||||||||
| X9 | B2 | + | + | + | + | + | + | + | + | + | |||||||||
| X16 | B2 | + | + | + | + | ||||||||||||||
| Y11 | B2 | + | + | + | + | + | + | + | + | + | + | + | |||||||
| Y17 | B2 | + | + | + | + | + | |||||||||||||
| W13 | B2 | + | + | + | + | + | + | + | + | + | |||||||||
| Y4 | B2 | + | + | + | + | + | + | + | + | ||||||||||
| X7 | B2 | + | + | + | + | + | + | + | + | + | + | + | + | + | |||||
| W11 | B2 | + | + | + | + | + | + | + | + | + | + | + | |||||||
| H6 | B2 | + | + | + | + | + | + | + | |||||||||||
| X12 | D | + | + | + | + | + | + | ||||||||||||
| X15 | D | + | + | + | + | + | + | ||||||||||||
| X10 | D | + | + | + | + | + | + | ||||||||||||
| X18 | D | + | + | + | + | + | + | + | + | ||||||||||
| X27 | D | + | + | + | + | + | + | + | + | + | |||||||||
| Y7 | E | + | + | ||||||||||||||||
| W8 | E | + | + | ||||||||||||||||
| W13 | E | + | + | + | + | + | + | + | |||||||||||
| H5 | F | + | + | + |
papGI and papGII were not detected.
Characterization of β-lactamases and PMQR determinants
All ESC-R E. coli tested harbored one or more β-lactamase genes. blaTEM, blaSHV, blaCTX−M, blaCMY−2, and blaDHA−1 were determined in 35 (87.5%), 14 (35%), 35 (87.5%), 3 (7.5%), and 2 (5%) of ESC-R E. coli isolates, respectively (Table 2). Moreover, eight blaOXA−48 genes associated with carbapenems (imipenem and meropenem) resistance were detected. For the blaCTX−M positive isolates, blaCTX−M−15 (n = 35) was the predominant specific subtype, and followed by blaCTX−M−123 (n = 14), blaCTX−M−1 (n = 10), blaCTX−M−14 (n = 10), and blaCTX−M−9 (n = 7). Sequencing of blaTEM gene revealed 31 blaTEM−1 and four blaTEM−30, whereas sequencing of blaSHV gene revealed 14 blaSHV−12. Eighty percent (32/40) of tested isolates harbored at least one PMQR gene, and 16 isolates harbored more than one PMQR genes. qnrB, qnrD, qnrS, qepA, and aac(6′)-Ib-cr were detected alone or incombination in 20, 22.5, 30, 10, and 65% of 40 ESC-R isolates, respectively, while qnrA was not detected (Table 2). Furthermore, all PMQR genes were detected in co-existence with β-lactamase genes, and one isolate harbored simultaneously β-lactamase genes blaTEM−1, blaSHV−12, blaCTX−M−15, blaCTX−M−9, blaCTX−M−14, blaDHA−1, and blaOXA−48 as well as PMQR genes qnrB, qnrD, qnrS, qepA, and aac(6′)-Ib-cr.
Table 2.
The occurrence of β-lactamases and PMQR genes in ESC-R E. coli isolates.
| Isolates ID | PG | MLST | β-lactamase genes | PMQR genes | Resistance profiles | |||
|---|---|---|---|---|---|---|---|---|
| Non-ESBL | ESBL | pAmpC | Carbapenemase | |||||
| B1 | A | ST1820 | TEM-30 | qnrS, aac(6′)-Ib-cr | CFZ, CTX, CPO, ENR, DOX, GEN | |||
| H2 | A | ST746 | TEM-1 | CTX-M-14 | AMC, CFZ, CTX, CAZ, ENR, DOX, SXT | |||
| W12 | A | ST44 | TEM-1 | CTX-M-15 | AMC, CFZ, CTX, CAZ, ENR, DOX, CHL | |||
| B6 | A | ST1700 | TEM-1 | CTX-M-1, CTX-M-15, CTX-M-123 | aac(6′)-Ib-cr | AMC, CFZ, CTX, CAZ, CPD, CRO, FOX, ENR, PRA, DOX, AMK, SXT | ||
| Y5 | A | ST167 | TEM-1 | SHV-12 | qnrD, qnrS | AMC, CTX, ENR, DOX, CHL, GEN | ||
| W4 | A | ST167 | TEM-1 | CTX-M-9, CTX-M-14, CTX-M-123 | OXA-48 | qnrD, qnrS | AMC, CFZ, CTX, CAZ, CPD, CRO, CPO, FOX, MEM, ENR, PRA, DOX, CHL, GEN, AMK, SXT | |
| X2 | A | ST10 | TEM-1 | CTX-M-15 | qnrD, qnrS | AMC, CFZ, CTX, CAZ, ENR, DOX, SXT | ||
| Y15 | A | ST10 | SHV-12, CTX-M-15 | qnrB, qnrD | AMC, CTX, CAZ, CPD, CPO, FOX, ENR, DOX, CHL, GEN, SXT | |||
| B3 | A | ST10 | SHV-12, CTX-M-15 | qnrB, aac(6′)-Ib-cr | AMC, CTX, CAZ, CPD, CRO, CPO, ENR, PRA, DOX, GEN, AMK, SXT | |||
| W15 | A | ST10 | TEM-1 | SHV-12, CTX-M-15 | OXA-48 | CFZ, CTX, CAZ, CPD, CRO, FOX, IPM, DOX, CHL, GEN, SXT | ||
| W6 | A | ST10 | SHV-12, CTX-M-15, CTX-M-14 | qnrB, qepA, aac(6′)-Ib-cr | AMC, CFZ, CTX, CAZ, CRO, CPO, FOX, ENR, PRA, DOX, CHL, SXT | |||
| Y13 | A | ST10 | TEM-1 | CTX-M-15, CTX-M-9, CTX-M-14 | qnrB, qnrD, qnrS, aac(6′)-Ib-cr | AMC, CFZ, CTX, CAZ, CPD, CRO, CPO, FOX, ENR, PRA, DOX, CHL, GEN, AMK, SXT | ||
| X1 | A | ST10 | TEM-1 | CTX-M-1, CTX-M-15, CTX-M-123 | qnrS, aac(6′)-Ib-cr | AMC, CFZ, CTX, CAZ, CPD, CRO, CPO, FOX, ENR, PRA, DOX, CHL, SXT | ||
| B5 | A | ST10 | TEM-1 | CTX-M-1, CTX-M-15, CTX-M-123 | qnrD, qnrS, aac(6′)-Ib-cr | AMC, CFZ, CTX, CAZ, CPD, CRO, CPO, FOX, ENR, PRA, DOX, CHL, SXT | ||
| H1 | A | ST10 | TEM-1 | CTX-M-1, CTX-M-15, CTX-M-123 | CMY-2 | qnrS, aac(6′)-Ib-cr | AMC, CTX, CAZ, CRO, CPO, FOX, IPM, ENR, PRA, DOX, CHL, GEN, AMK, SXT | |
| X5 | B1 | ST75 | SHV-12, CTX-M-15 | aac(6′)-Ib-cr | AMC, CTX, CAZ, CRO, ENR, CHL, SXT | |||
| X6 | B1 | ST1177 | TEM-1 | CTX-M-15, CTX-M-123 | AMC, CFZ, CTX, CAZ, CPD, CRO, CPO, DOX, CHL, SXT | |||
| Y10 | B2 | ST375 | TEM-30 | aac(6′)-Ib-cr | AMC, CTX, CAZ, ENR | |||
| X8 | B2 | ST302 | TEM-1 | CTX-M-15 | aac(6′)-Ib-cr | AMC, CFZ, CTX, CAZ, ENR, DOX, GEN | ||
| W19 | B2 | ST73 | TEM-1 | CTX-M-1, CTX-M-15, CTX-M-123 | aac(6′)-Ib-cr | AMC, CFZ, CTX, CAZ, CPD, CRO, FOX, ENR, DOX, CHL | ||
| W9 | B2 | ST104 | TEM-1 | CTX-M-1, CTX-M-9, CTX-M-14 | OXA-48 | AMC, CFZ, CTX, CAZ, CPD, CRO, CPO, FOX, MEM, ENR, DOX, CHL, GEN, SXT | ||
| Y8 | B2 | ST131 | TEM-1 | CTX-M-15 | aac(6′)-Ib-cr | AMC, CTX, CAZ, CRO, ENR, PRA, DOX | ||
| X9 | B2 | ST131 | TEM-1 | CTX-M-15 | DHA-1 | qnrD, qnrS | AMC, CFZ, CTX, CAZ, CPD, CRO, CPO, FOX, MEM, ENR, PRA, DOX, CHL, SXT | |
| X16 | B2 | ST131 | TEM-1 | CTX-M-15 | OXA-48 | qnrS, qepA, aac(6′)-Ib-cr | AMC, CFZ, CTX, CAZ, CPD, CRO, CPO, FOX, IPM, ENR, PRA, DOX, CHL, GEN, AMK, SXT | |
| Y11 | B2 | ST131 | TEM-1 | SHV-12, CTX-M-15 | qnrB | AMC, CFZ, CTX, ENR, DOX, SXT | ||
| Y17 | B2 | ST131 | TEM-1 | SHV-12, CTX-M-15 | aac(6′)-Ib-cr | AMC, CFZ, CTX, CAZ, CPO, FOX, ENR, PRA, DOX, SXT | ||
| W13 | B2 | ST131 | TEM-1 | CTX-M-1, CTX-M-15 | aac(6′)-Ib-cr | AMC, CFZ, CTX, CAZ, CPD, CRO, FOX, ENR, DOX, SXT | ||
| Y4 | B2 | ST131 | TEM-1 | CTX-M-15, CTX-M-123 | qnrS, aac(6′)-Ib-cr | CFZ, CTX, CAZ, CPD, CRO, CPO, FOX, ENR, PRA, DOX, CHL, GEN, AMK, SXT | ||
| X7 | B2 | ST131 | TEM-1 | SHV-12, CTX-M-15, CTX-M-123 | CMY-2 | qnrB, qnrD, aac(6′)-Ib-cr | AMC, CFZ, CTX, CAZ, CPD, CRO, CPO, FOX, ENR, PRA, DOX, CHL, GEN, AMK, SXT | |
| W11 | B2 | ST131 | TEM-1 | SHV-12, CTX-M-1, CTX-M-15, CTX-M-123 | CMY-2 | aac(6′)-Ib-cr | AMC, CFZ, CTX, CAZ, CPD, CRO, CPO, FOX, ENR, PRA, DOX, CHL, GEN, AMK, SXT | |
| H6 | B2 | ST131 | TEM-1 | CTX-M-1, CTX-M-15, CTX-M-9, CTX-M-14, CTX-M-123 | aac(6′)-Ib-cr | AMC, CFZ, CTX, CAZ, CPD, CRO, FOX, ENR, PRA, DOX, CHL, GEN, AMK, SXT | ||
| X12 | D | ST69 | TEM-1 | CTX-M-15, CTX-M-123 | OXA-48 | aac(6′)-Ib-cr | AMC, CFZ, CTX, CAZ, CPD, CRO, CPO, FOX, MEM, ENR, PRA, DOX, CHL, GEN, AMK, SXT | |
| X15 | D | ST38 | TEM-1 | CTX-M-15, CTX-M-9, CTX-M-14 | AMC, CFZ, CTX, CAZ, CPD, CRO, DOX, CHL, SXT | |||
| X10 | D | ST405 | TEM-1 | SHV-12, CTX-M-15, CTX-M-14, CTX-M-123 | OXA-48 | aac(6′)-Ib-cr | AMC, CFZ, CTX, CAZ, CPD, CRO, CPO, FOX, IPM, ENR, PRA, DOX, CHL, GEN, AMK, SXT | |
| X18 | D | ST648 | TEM-1 | SHV-12, CTX-M-15, CTX-M-9, CTX-M-14 | DHA-1 | OXA-48 | qnrB, qnrD, qnrS, qepA, aac(6′)-Ib-cr | AMC, CFZ, CTX, CAZ, CPD, CRO, CPO, FOX, IPM, ENR, PRA, DOX, CHL, GEN, AMK, SXT |
| X27 | D | ST68 | TEM-30 | CTX, ENR, DOX | ||||
| Y7 | E | ST1421 | TEM-30 | qnrB, aac(6′)-Ib-cr | AMC, CFZ, CTX, CPD, ENR, PRA, DOX | |||
| W8 | E | ST2375 | SHV-12, CTX-M-15 | AMC, CFZ, CTX, CAZ, CPD, DOX, CHL, SXT | ||||
| W13 | E | ST3058 | TEM-1 | SHV-12, CTX-M-15 | OXA-48 | aac(6′)-Ib-cr | AMC, CFZ, CTX, CAZ, CPD, CRO, CPO, FOX, MEM, ENR, DOX, CHL, GEN, AMK, SXT | |
| H5 | F | ST3630 | TEM-1 | CTX-M-1, CTX-M-9, CTX-M-14, CTX-M-123 | qepA, aac(6′)-Ib-cr | AMC, CFZ, CTX, CAZ, CPD, CRO, FOX, ENR, PRA, DOX, CHL, SXT | ||
AMC, amoxicillin-clavulanic acid; CFZ, cefazolin; CTX, cefotaxime; CAZ, ceftazidime; CPD, cefpodoxime; CRO, ceftriaxone; CPO, cefoperazone; FOX, cefoxitin; IPM, imipenem; MEM, meropenem; ENR, enrofloxacin; PRA, pradofloxacin; DOX, doxycycline; CHL, chloramphenicol; GEN, gentamicin; AMK, amikacin; SXT, sulfamethoxazole-trimethoprim.
MLST profiles
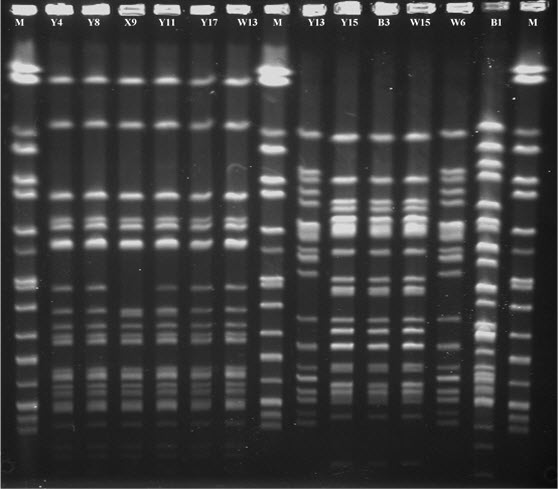
The 40 ESC-R E. coli isolates tested were assigned to 22 STs (Table 2), and ST131 (n = 10) of phylogenetic group B2 and ST10 (n = 9) of phylogenetic group A accounted for almost 50% of isolates. Two isolates were assigned to ST167. The remaining isolates exhibited diverse ST types. All ST131 and ST10 isolates expressed resistance to ESC-R E. coli tested, and they were also ST10 positively associated with blaCTX−M−15. Furthermore, the ST131 and ST10 isolates were analyzed by PFGE in order to determine the genetic relatedness, PFGE indicated that some ST131 and ST10 isolates displayed the same PFGE profile (Figure S1).
Conjugation experiments
Five transconjugants were obtained finally, the parental isolates successfully transferred the bla genes to the E. coli J53 AZr recipient strain. Moreover, qnrS and aac-(6′)-Ib-cr were co-transferred with blaCTX−M, whereas other PMQR genes were not observed among transconjugants. The transconjugants had resistance profiles similar to those of their parental isolates. All transconjugants showed high-level resistance to β-lactam antibiotics at the same level as the donor strain. For fluoroquinolones, the transconjugants showed 4–8-fold increases in the MICs of ciprofloxacin when compared with the donor strain E. coli J53 AZr. However, the transconjugants remained susceptible to gentamicin, doxycycline and trimethoprim-sulfamethoxazole.
Discussion
Cephalosporin-resistant E. coli isolates seem to be the emergent cause of serious infections of humans and animals worldwide. The present study firstly demonstrated the widespread occurrence and molecular characterization of β-lactamases and PMQR genes among ESC-R E. coli isolates from dogs in Shaanxi province, China. The previous studies showed that the occurrence of ESBL carriage in companion animals varies considerably between countries from 1 to 55% (O'keefe et al., 2010; Hordijk et al., 2013), and our data showed that 24.2% of isolates from dogs were ESC-R E. coli, which is close to the prevalence (22.3%) reported in the retail meat in Shaanxi province (Xi et al., 2015). While it is significantly higher than the prevalence in the United States (3%; Shaheen et al., 2011) and Switzerland (7.5%; Huber et al., 2013; P < 0.0001), and much lower than that in The Netherlands (45–55%) and Guangdong province of China (54.5%) (Sun et al., 2010; Hordijk et al., 2013). The possible reason for this obvious difference is that the difference geographical origin of the isolates, the differences in study population and associated antimicrobial selective pressure. The fluoroquinolones and the third-generation cephalosporins are important for the treatment of serious infections in humans and companion animals caused by Enterobacteriaceae (Jiang et al., 2012), while ESBL-producing E. coli isolates associated with MDR phenotype continuously increased with the wide use of cephalosporins, fluoroquinolones and other antimicrobials.
All isolates in the current study expressed MDR phenotype, and exhibited high resistance rates to cefotaxime (100%) and enrofloxacin (95%). Furthermore, it was noteworthy that 10 ESC-R E. coli isolates were resistance to meropenem and imipenem agreeing with a previous study in the United States (Liu et al., 2016). It is worrisome although they were still the most potent and effective antibiotics in our study. Owing to the relevance of imipenem and meropenem in the treatment of infections caused by ESBL-positive or MDR E. coli, the surveillance of carbapenems resistance and the implementation of guidelines for the rational use of carbapenems will be urgently needed to prevent the progress of antimicrobial resistance in China. Our results showed that ESC-R E. coli isolates from dogs belonged mainly to phylogroup A and B2, and to a lesser extent, to phylogenetic D, whereas groups B1, F, and E isolates were very little. The previous studies showed that E. coli from pigs or duck in China also mainly felled into phylogenetic groups A (Wang et al., 2010; Ma et al., 2012). While another study has reported that the E. coli from cats in the United States belonged predominantly to phyogenetic group B2 (Liu et al., 2015). In regards to the linkage of phylogenetic group and the PMQR genes and β-lactamases, isolates of group A harbored more PMQR genes, and the isolates of group B2 harbored more β-lactamases.
The blaCTX−M was the dominating ESBL gene in the E. coli tested despite the specific genotype of blaCTX−M are undergoing great changes. blaCTX−M was detected in 87.5% of ESC-R E. coli isolates, and blaCTX−M−15 was the most common specific CTX-M subtype in the present study. While blaCTX−M−14 subtype was the most common CTX-M enzyme in the E. coli from food animals in China according to the previous study (Yu et al., 2015). It was worth noting that blaOXA−48 genes were detected in eight of the 40 (20%) ESC-R E. coli isolates, and they were also associated with imipenem or meropenem resistance as a previous study described (Liu et al., 2016). blaOXA−48 was firstly discovered in E. coli from dogs in Germany in 2013, and then it was reported in E. coli from companion animals in the United States in 2016 (Stolle et al., 2013; Liu et al., 2016). To our knowledge, this is the first report of blaOXA−48 in E. coli from dogs in China. Although carbapenemase is not an ESBL, blaOXA−48 has emerged as a major carbapenemase associated with the Enterobacteriaceae, and it can also hydrolyze carbapenems and hydrolyzes β-lactamase inhibitors (Mathers et al., 2013). Emergence of PMQR genes have been reported worldwide and is being documented in ESBL-producing E. coli (Xu et al., 2015). In the present study, PMQR genes were present in 80% of ESC-R E. coli isolates, and aac(6′)-Ib-cr was detected alone or in combination with qnr and qepA in present in 65% of isolates, and it occurred in ESBL producers with a higher prevalence than qnr genes, especially in E. coli isolates carrying the blaCTX−M−15 gene. It was consistent with a previous study that aac(6′)-Ib-cr gene was the predominant PMQR gene among Enterobacteriaceae isolates from companion animals in Guangdong province in China (Ma et al., 2009). While oqxAB was the predominant PMQR gene in E. coli from food animals in China (Liu et al., 2013; Xu et al., 2015). In addition, qepA gene was detected in combination with other PMQR genes and ESBL genes in four isolates (10%). The presence of PMQRs is of great importance because they are not merely able to confer resistance against fluoroquinolones but as well are often related to ESBLs and/or AmpC β-lactamases. Emergence of PMQR gene qnr, aac(6′)-Ib-cr and qepA has been reported worldwide and were often found to be strongly associated with ESBL genes even located on the same plasmid (Strahilevitz et al., 2009; Poirel et al., 2012). Our results showed that qnrS and aac-(6′)-Ib-cr were co-transferred with blaCTX−M in the conjugation experiments, and more studies should be carried out in the future in order to ensure that the six PMQR genes were located on the same plasmid or not. However, we cannot find out the linkage among the virulence gene profiles, the PMQR genes and the β-lactamases as the limited number of isolates.
MLST investigation showed that ST131 (n = 10) and ST10 (n = 9) accounted for 47.5% of tested isolates, all ST131 and ST10 isolates were associated with the blaCTX−M−15 enzyme. ST131 clone has high virulence potential all over the world and represents a major public health problem, and it has emerged and disseminated in E. coli from dogs in different continents (Ewers et al., 2010; Platell et al., 2011; Harada et al., 2012; Dahmen et al., 2013; Liu et al., 2016). Other STs, such as ST10, ST38, ST69, and ST167 identified in this study were also reported in human and dogs according to the present data from the MLST database, and ST38 clone can play an important role in the worldwide distribution of CTX-M-producing E. coli (Pitout, 2012). PFGE is considered to be a highly discriminative subtyping method in epidemiological investigation, our results suggested that some ST131 isolates and ST10 isolates were closely related, and displayed related restriction patterns, respectively. We should pay attention to these isolates in the future study. However, we cannot find the relationship between the virulence gene profiles and the prevalence of β-lactamases and the STs as the relatively small sample size (40 isolates). Alarmingly, one ST648 clone of phylogenetic group D carried simultaneously blaTEM−1, blaTSHV−12, blaCTX−M−15, blaCTX−M−9, blaCTX−M−14, blaOXA−48, qnrB, qnrD qnrS, qepA, and aac(6′)-1b-cr. Two recent studies in Europe suggested that ST648 clone may represent a novel genotype that combines MDR phenotype, extraintestinal virulence and zoonotic potential in companion animals (Huber et al., 2013; Ewers et al., 2014). To our knowledge, this is the first report of co-existence of 11 β-lactamase and PMQR genes in one E. coli isolate, which will obviously improve the resistance to cephalosporins, β-lactamase inhibitors and fluoroquinolones. It is an extremely worrisome sign of development of untreatable infections as the use of β-lactams for the treatment of infections caused by E. coli has been and will continue to be the main line of defense against these bacterial agents (Silva-Sanchez et al., 2013).
Conclusion
In summary, all investigated ESC-R E. coli isolates from dogs in Shaanxi province expressed MDR phenotype, exhibited high resistance rate to extended-spectrum cephalosporins and fluoroquinolones, showed high prevalence of β-lactamases and PMQR genes, and all PMQR genes were detected in co-existence with at least one β-lantamase gene. The clonal group ST131, blaCTX−M−15 gene and aac(6′)-Ib-cr gene were the dominant ST, β-lactamase gene and PMQR gene among the tested isolates, respectively. This is also the first description of the co-existence of blaTEM−1, blaSHV−12, blaCTX−M−15, blaCTX−M−9, blaCTX−M−14, blaOXA−48, qnrB, qnrD qnrS, qepA, and aac(6′)-Ib-cr genes in one E. coli isolate. High prevalence and combination of β-lactamases and PMQR genes is an extremely worrisome sign of treatment of the infections.
Author contributions
XL conceived and designed the experiments. HL designed the experiment and drafted the manuscript. XL, HL and CH performed the experiments. XL, YL analyzed and explained the data for the work. All authors critically revised and approved the final manuscript.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This study was supported by the Fundamental Research Funds for the Central Universities (No. 2452016044) and Scientific Research Foundation for PhD of Northwest A&F University (No. 2013BSJJ012).
Supplementary material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2016.01843/full#supplementary-material
XbaI-generated pulse-field gel electrophoresis (PFGE) patterns of six ST131 (Y4, Y8, X9, Y11, Y17, and W13 from lane 2–7) and six ST10 (Y13, Y15, B3, W15, W6, and B1 from lane 9–14) isolates. M, Salmonella marker (lane 1, 8, 15).
{kind=link}
The Oligonucleotide primers used in this study.
References
- Aly S. A., Debavalya N., Suh S. J., Oryazabal O. A., Boothe D. M. (2012). Molecular mechanisms of antimicrobial resistance in fecal Escherichia coli of healthy dogs after enrofloxacin or amoxicillin administration. Can. J. Microbiol. 58, 1288–1294. 10.1139/w2012-105 [DOI] [PubMed] [Google Scholar]
- Briales A., Rodriguez-Martinez J. M., Velasco C., De Alba P. D., Rodriguez-Bano J., Martinez-Martinez L., et al. (2012). Prevalence of plasmid-mediated quinolone resistance determinants qnr and aac(6′)-Ib-cr in Escherichia coli and Klebsiella pneumoniae producing extended-spectrum β-lactamases in Spain. Int. J. Antimicrob. Agents 39, 431–434. 10.1016/j.ijantimicag.2011.12.009 [DOI] [PubMed] [Google Scholar]
- Cantas L., Suer K., Guler E., Imir T. (2015). High emergence of ESBL-producing E. coli cystitis: time to get smarter in Cyprus. Front. Microbiol. 6:1446. 10.3389/fmicb.2015.01446 [DOI] [PMC free article] [PubMed] [Google Scholar]
- CLSI (2013). Performance standards for antimicrobial disk and dilution susceptibility tests for bacterial isolated from animals; approved standard. fourth edition and supplement, CLSI document VET01-A4 (standard) and VET01-S2 (supplement), in Clinical and Laboratory Standards Institute (Wayne, PA: ). [Google Scholar]
- Costa D., Vinue L., Poeta P., Coelho A. C., Matos M., Saenz Y., et al. (2009). Prevalence of extended-spectrum β-lactamase-producing Escherichia coli isolates in faecal samples of broilers. Vet. Microbiol. 138, 339–344. 10.1016/j.vetmic.2009.03.029 [DOI] [PubMed] [Google Scholar]
- Dahmen S., Haenni M., Châtre P., Madec J. Y. (2013). Characterization of blaCTX−M IncFII plasmids and clones of Escherichia coli from pets in France. J. Antimicrob. Chemother. 68, 2797–2801. 10.1093/jac/dkt291 [DOI] [PubMed] [Google Scholar]
- Ewers C., Bethe A., Stamm I., Grobbel M., Kopp P. A., Guerra B., et al. (2014). CTX-M-15-D-ST648 Escherichia coli from companion animals and horses: another pandemic clone combining multiresistance and extraintestinal virulence? J. Antimicrob. Chemother. 69, 1224–1230. 10.1093/jac/dkt516 [DOI] [PubMed] [Google Scholar]
- Ewers C., Grobbel M., Stamm I., Kopp P. A., Diehl I., Semmler T., et al. (2010). Emergence of human pandemic O25:H4-ST131 CTX-M-15 extended-spectrum-β-lactamase-producing Escherichia coli among companion animals. J. Antimicrob. Chemother. 65, 651–660. 10.1093/jac/dkq004 [DOI] [PubMed] [Google Scholar]
- Harada K., Nakai Y., Kataoka Y. (2012). Mechanisms of resistance to cephalosporin and emergence of O25b-ST131 clone harboring CTX-M-27 β-lactamase in extraintestinal pathogenic Escherichia coli from dogs and cats in Japan. Microbiol. Immunol. 56, 480–485. 10.1111/j.1348-0421.2012.00463.x [DOI] [PubMed] [Google Scholar]
- Hordijk J., Schoormans A., Kwakernaak M., Duim B., Broens E., Dierikx C., et al. (2013). High prevalence of fecal carriage of extended spectrum β-lactamase/AmpC-producing Enterobacteriaceae in cats and dogs. Front. Microbiol. 4:242. 10.3389/fmicb.2013.00242 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huber H., Zweifel C., Wittenbrink M. M., Stephan R. (2013). ESBL-producing uropathogenic Escherichia coli isolated from dogs and cats in Switzerland. Vet. Microbiol. 162, 992–996. 10.1016/j.vetmic.2012.10.029 [DOI] [PubMed] [Google Scholar]
- Ishida Y., Ahmed A. M., Mahfouz N. B., Kimura T., El-Khodery S. A., Moawad A. A., et al. (2010). Molecular analysis of antimicrobial resistance in gram-negative bacteria isolated from fish farms in Egypt. J. Vet. Med. Sci. 72, 727–734. 10.1292/jvms.09-0538 [DOI] [PubMed] [Google Scholar]
- Jiang H. X., Tang D., Liu Y. H., Zhang X. H., Zeng Z. L., Xu L., et al. (2012). Prevalence and characteristics of β-lactamase and plasmid-mediated quinolone resistance genes in Escherichia coli isolated from farmed fish in China. J. Antimicrob. Chemother. 67, 2350–2353. 10.1093/jac/dks250 [DOI] [PubMed] [Google Scholar]
- Liu B. T., Yang Q. E., Li L., Sun J., Liao X. P., Fang L. X., et al. (2013). Dissemination and characterization of plasmids carrying oqxAB-blaCTX−M genes in Escherichia coli isolates from food-producing animals. PLoS ONE 8:e73947. 10.1371/journal.pone.0073947 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu X., Boothe D. M., Thungrat K., Aly S. (2012). Mechanisms accounting for fluoroquinolone multidrug resistance Escherichia coli isolated from companion animals. Vet. Microbiol. 161, 159–168. 10.1016/j.vetmic.2012.07.019 [DOI] [PubMed] [Google Scholar]
- Liu X., Thungrat K., Boothe D. M. (2015). Multilocus sequence typing and virulence profiles in uropathogenic Escherichia coli isolated from cats in the United States. PLoS ONE 10:e0143335. 10.1145/2818302 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu X., Thungrat K., Boothe D. M. (2016). Occurrence of OXA-48 carbapenemase and other β-lactamase genes in ESBL-producing multidrug resistant Escherichia coli from dogs and cats in the United States, 2009-2013. Front. Microbiol. 7:1057. 10.3389/fmicb.2016.01057 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loncaric I., Stalder G. L., Mehinagic K., Rosengarten R., Hoelzl F., Knauer F., et al. (2013). Comparison of ESBL–and AmpC producing Enterobacteriaceae and methicillin-resistant Staphylococcus aureus (MRSA) isolated from migratory and resident population of rooks (Corvus frugilegus) in Austria. PLoS ONE 8:e84048. 10.1371/journal.pone.0084048 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma J., Liu J. H., Lv L., Zong Z., Sun Y., Zheng H., et al. (2012). Characterization of extended-spectrum β-lactamase genes found among Escherichia coli isolates from duck and environmental samples obtained on a duck farm. Appl. Environ. Microbiol. 78, 3668–3673. 10.1128/AEM.07507-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma J., Zeng Z., Chen Z., Xu X., Wang X., Deng Y., et al. (2009). High prevalence of plasmid-mediated quinolone resistance determinants qnr, aac(6′)-Ib-cr, and qepA among ceftiofur-resistant Enterobacteriaceae isolates from companion and food-producing animals. Antimicrob. Agents Chemother. 53, 519–524. 10.1128/AAC.00886-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mathers A. J., Hazen K. C., Carroll J., Yeh A. J., Cox H. L., Bonomo R. A., et al. (2013). First clinical cases of OXA-48-producing carbapenem-resistant Klebsiella pneumoniae in the United States: the “menace” arrives in the new world. J. Clin. Microbiol. 51, 680–683. 10.1128/JCM.02580-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nordmann P., Naas T., Poirel L. (2011). Global spread of Carbapenemase-producing Enterobacteriaceae. Emerging Infect. Dis. 17, 1791–1798. 10.3201/eid1710.110655 [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'keefe A., Hutton T. A., Schifferli D. M., Rankin S. C. (2010). First detection of CTX-M and SHV extended-spectrum β-lactamases in Escherichia coli urinary tract isolates from dogs and cats in the United States. Antimicrob. Agents Chemother. 54, 3489–3492. 10.1128/AAC.01701-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perez-Perez F. J., Hanson N. D. (2002). Detection of plasmid-mediated AmpC β-lactamase genes in clinical isolates by using multiplex PCR. J. Clin. Microbiol. 40, 2153–2162. 10.1128/JCM.40.6.2153-2162.2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pitout J. D. (2012). Extraintestinal pathogenic Escherichia coli: a combination of virulence with antibiotic resistance. Front. Microbiol. 3:9. 10.3389/fmicb.2012.00009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Platell J. L., Cobbold R. N., Johnson J. R., Heisig A., Heisig P., Clabots C., et al. (2011). Commonality among Fluoroquinolone-Resistant Sequence Type ST131 Extraintestinal Escherichia coli Isolates from humans and companion animals in Australia. Antimicrob. Agents Chemother. 55, 3782–3787. 10.1128/AAC.00306-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poirel L., Cattoir V., Nordmann P. (2012). Plasmid-Mediated quinolone resistance; interactions between human, animal, and environmental ecologies. Front. Microbiol. 3:24. 10.3389/fmicb.2012.00024 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poirel L., Walsh T. R., Cuvillier V., Nordmann P. (2011). Multiplex PCR for detection of acquired carbapenemase genes. Diagn. Microbiol. Infect. Dis. 70, 119–123. 10.1016/j.diagmicrobio.2010.12.002 [DOI] [PubMed] [Google Scholar]
- Shaheen B. W., Nayak R., Foley S. L., Kweon O., Deck J., Park M., et al. (2011). Molecular characterization of resistance to extended-spectrum cephalosporins in clinical Escherichia coli isolates from companion animals in the United States. Antimicrob. Agents Chemother. 55, 5666–5675. 10.1128/AAC.00656-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silva-Sanchez J., Cruz-Trujillo E., Barrios H., Reyna-Flores F., Sanchez-Perez A., Bacterial Resistance C., et al. (2013). Characterization of plasmid-mediated quinolone resistance (PMQR) genes in extended-spectrum β-lactamase-producing Enterobacteriaceae pediatric clinical isolates in Mexico. PLoS ONE 8:e77968. 10.1371/journal.pone.0077968 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stolle I., Prenger-Berninghoff E., Stamm I., Scheufen S., Hassdenteufel E., Guenther S., et al. (2013). Emergence of OXA-48 carbapenemase-producing Escherichia coli and Klebsiella pneumoniae in dogs. J. Antimicrob. Chemother. 68, 2802–2808. 10.1093/jac/dkt259 [DOI] [PubMed] [Google Scholar]
- Strahilevitz J., Jacoby G. A., Hooper D. C., Robicsek A. (2009). Plasmid-mediated quinolone resistance: a multifaceted threat. Clin. Microbiol. Rev. 22, 664–689. 10.1128/CMR.00016-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun Y., Zeng Z., Chen S., Ma J., He L., Liu Y., et al. (2010). High prevalence of bla(CTX-M) extended-spectrum β-lactamase genes in Escherichia coli isolates from pets and emergence of CTX-M-64 in China. Clin. Microbiol. Infect. 16, 1475–1481. 10.1111/j.1469-0691.2010.03127.x [DOI] [PubMed] [Google Scholar]
- Tenover F. C., Arbeit R. D., Goering R. V., Mickelsen P. A., Murray B. E., Persing D. H., et al. (1995). Interpreting chromosomal DNA restriction patterns produced by Pulsed-Field Gel-Electrophoresis - Criteria for bacterial strain typing. J. Clin. Microbiol. 33, 2233–2239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tian G. B., Wang H. N., Zhang A. Y., Zhang Y., Fan W. Q., Xu C. W., et al. (2012). Detection of clinically important β-lactamases in commensal Escherichia coli of human and swine origin in western China. J. Med. Microbiol. 61, 233–238. 10.1099/jmm.0.036806-0 [DOI] [PubMed] [Google Scholar]
- Tian G. B., Wang H. N., Zou L. K., Tang J. N., Zhao Y. W., Ye M. Y., et al. (2009). Detection of CTX-M-15, CTX-M-22, and SHV-2 extended-spectrum β-lactamases (ESBLs) in Escherichia coli fecal-sample isolates from pig farms in China. Foodborne Pathog. Dis. 6, 297–304. 10.1089/fpd.2008.0164 [DOI] [PubMed] [Google Scholar]
- Wang X. M., Jiang H. X., Liao X. P., Liu J. H., Zhang W. J., Zhang H., et al. (2010). Antimicrobial resistance, virulence genes, and phylogenetic background in Escherichia coli isolates from diseased pigs. FEMS Microbiol. Lett. 306, 15–21. 10.1111/j.1574-6968.2010.01917.x [DOI] [PubMed] [Google Scholar]
- Wang Y., He T., Han J., Wang J., Foley S. L., Yang G., et al. (2012). Prevalence of ESBLs and PMQR genes in fecal Escherichia coli isolated from the non-human primates in six zoos in China. Vet. Microbiol. 159, 53–59. 10.1016/j.vetmic.2012.03.009 [DOI] [PubMed] [Google Scholar]
- Wasyl D., Hasman H., Cavaco L. M., Aarestrup F. M. (2012). Prevalence and characterization of cephalosporin resistance in nonpathogenic Escherichia coli from food-producing animals slaughtered in Poland. Microb. Drug Resist. 18, 79–82. 10.1089/mdr.2011.0033 [DOI] [PubMed] [Google Scholar]
- Xi M., Wu Q., Wang X., Yang B., Xia X., Li D. (2015). Characterization of Extended-Spectrum β-Lactamase-Producing Escherichia coli strains isolated from retail foods in Shaanxi Province, China. J. Food Prot. 78, 1018–1023. 10.4315/0362-028X.JFP-14-490 [DOI] [PubMed] [Google Scholar]
- Xu G., An W., Wang H., Zhang X. (2015). Prevalence and characteristics of extended-spectrum β-lactamase genes in Escherichia coli isolated from piglets with post-weaning diarrhea in Heilongjiang province, China. Front. Microbiol. 6:1103. 10.3389/fmicb.2015.01103 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yan J. J., Hong C. Y., Ko W. C., Chen Y. J., Tsai S. H., Chuang C. L., et al. (2004). Dissemination of blaCMY−2 among Escherichia coli isolates from food animals, retail ground meats, and humans in southern Taiwan. Antimicrob. Agents Chemother. 48, 1353–1356. 10.1128/AAC.48.4.1353-1356.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu T., Jiang X., Fu K., Liu B., Xu D., Ji S., et al. (2015). Detection of Extended-Spectrum β-Lactamase and Plasmid-Mediated quinolone resistance determinants in Escherichia coli Isolates from retail meat in China. J. Food Sci. 80, M1039–M1043. 10.1111/1750-3841.12870 [DOI] [PubMed] [Google Scholar]
- Yuan L., Liu J. H., Hu G. Z., Pan Y. S., Liu Z. M., Mo J., et al. (2009). Molecular characterization of extended-spectrum β-lactamase-producing Escherichia coli isolates from chickens in Henan Province, China. J. Med. Microbiol. 58, 1449–1453. 10.1099/jmm.0.012229-0 [DOI] [PubMed] [Google Scholar]
- Zhao W. H., Hu Z. Q. (2013). Epidemiology and genetics of CTX-M extended-spectrum β-lactamases in Gram-negative bacteria. Crit. Rev. Microbiol. 39, 79–101. 10.3109/1040841X.2012.691460 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
XbaI-generated pulse-field gel electrophoresis (PFGE) patterns of six ST131 (Y4, Y8, X9, Y11, Y17, and W13 from lane 2–7) and six ST10 (Y13, Y15, B3, W15, W6, and B1 from lane 9–14) isolates. M, Salmonella marker (lane 1, 8, 15).
The Oligonucleotide primers used in this study.