

Significance
Carbon and nitrogen cycles have been altered dramatically by human activities. Methane-producing (methanogenic) and methane-consuming (methanotrophic) microorganisms control the emission of methane, one of the most potent greenhouse gases, to the atmosphere. Earlier studies identified methanotrophic microorganisms that use methane as an electron donor and oxygen, sulfate, nitrite, and nitrate as electron acceptors. Previous research showed that methanotrophy coupled to the reduction of oxidized metals could be important in the environment. In the current paper, we identified archaea of the order Methanosarcinales, related to “Candidatus Methanoperedens nitroreducens,” which couple the reduction of environmentally relevant particulate forms of iron and manganese to methane oxidation, filling one of the remaining lacunas in anaerobic methane oxidation.
Keywords: anaerobic oxidation of methane, archaea, iron reduction, manganese reduction, multiheme proteins
Abstract
Anaerobic oxidation of methane (AOM) is crucial for controlling the emission of this potent greenhouse gas to the atmosphere. Nitrite-, nitrate-, and sulfate-dependent methane oxidation is well-documented, but AOM coupled to the reduction of oxidized metals has so far been demonstrated only in environmental samples. Here, using a freshwater enrichment culture, we show that archaea of the order Methanosarcinales, related to “Candidatus Methanoperedens nitroreducens,” couple the reduction of environmentally relevant forms of Fe3+ and Mn4+ to the oxidation of methane. We obtained an enrichment culture of these archaea under anaerobic, nitrate-reducing conditions with a continuous supply of methane. Via batch incubations using [13C]methane, we demonstrated that soluble ferric iron (Fe3+, as Fe-citrate) and nanoparticulate forms of Fe3+ and Mn4+ supported methane-oxidizing activity. CO2 and ferrous iron (Fe2+) were produced in stoichiometric amounts. Our study connects the previous finding of iron-dependent AOM to microorganisms detected in numerous habitats worldwide. Consequently, it enables a better understanding of the interaction between the biogeochemical cycles of iron and methane.
Methane is of interest to scientists and society for many reasons: It is one of the most important fossil fuels, a significant greenhouse gas, as well as an abundant natural hydrocarbon and substrate for microorganisms. However, due to its high activation energy, it can only be used by a few specialized groups of archaea and bacteria. Although aerobic methane-oxidizing bacteria have been known for 110 years (1), it was only at the turn of the last century that organisms performing anaerobic oxidation of methane (AOM) were first identified (2). These organisms couple AOM to sulfate reduction and belong to distinct clades within the methanogens (phylum Euryarchaeota), microorganisms that normally produce methane under anoxic conditions. These anaerobic methanotrophic (ANME) archaea have vast importance in regulating Earth’s climate, as they prevent the escape of the large quantities of methane contained in the oceans’ sediments and gas hydrates (2). Because CO2 has a 12 times weaker radiative forcing per molecule than methane (3), converting methane-carbon to CO2, assimilating it into biomass, or precipitating it as carbonates altogether reduce global warming effects considerably (2).
Until a decade ago, sulfate, a major constituent of seawater, was the only known electron acceptor for anaerobic oxidation of methane (2, 4). However, a variety of other environmentally important oxidized compounds—most notably Fe3+, Mn4+, nitrate (NO3−) (5), and nitrite (NO2−) (6)—are thermodynamically more favorable electron acceptors than sulfate. The lack of known methanotrophs capable of coupling AOM to metal oxide reduction in particular represents a fundamental gap in our understanding of the dynamics of AOM in the environment.
For nitrogenous electron acceptors, two microbial players and processes have by now been identified: Methylomirabilis oxyfera, a bacterium of the NC10 phylum, is capable of producing its own oxygen from nitrite, and uses this oxygen to activate methane, analogous to aerobic methanotrophs (6). In addition, archaea of a new group within Methanosarcinales [termed AOM-associated archaea (AAA) (2), sometimes also referred to as ANME-2d], originally coenriched together with M. oxyfera (7), were recently found to couple the oxidation of methane to the reduction of nitrate to nitrite (5). In reaching these insights, laboratory enrichment cultures of the responsible microorganisms were of crucial importance.
Even though rock weathering supplies ample amounts of iron, manganese, and other metal oxides suitable as electron acceptors to freshwater bodies (8), and eventually oceans, evidence for the occurrence of iron- and manganese-dependent methane oxidation in nature is scarce. By geochemical profiling or isotope tracer studies, the coupling of methane oxidation to metal reduction has been demonstrated or suggested in lake sediments (9–11), lake water (12), a contaminated aquifer (13), a mud volcano (14), and marine sediments (15–18). The possible role of a cryptic sulfur cycle supplying sulfate via Fe3+-mediated reoxidation of sulfides (19, 20) could often not be excluded in these studies, and none of them identified the microorganisms responsible for the process nor succeeded in obtaining an enrichment culture. Only two studies investigated methane oxidation coupled to Fe3+ or Mn4+ reduction in naturally enriched samples of ANME-2 archaea. Using marine samples, Nauhaus et al. reported that methane oxidation was not coupled to the reduction of ferrihydrite, iron citrate, humic acids, and anthraquinone-2,6-disulfonate (AQDS) (21), whereas recently Scheller et al. showed that marine samples containing ANME-2a, ANME-2c, and other archaea oxidized methane in the presence of chelated iron and could couple methane oxidation activity to AQDS reduction (22). Nevertheless, in nature, metals predominantly occur in particulate forms, which are more difficult to access (15, 16, 19, 23). In short, the microorganisms that can couple the oxidation of methane to the reduction of environmentally relevant species of oxidized metals remain unknown.
Here we demonstrate Fe3+- and Mn4+-dependent methane oxidation in a freshwater enrichment culture of archaeal methanotrophs enriched from the sediment of a Dutch canal (Twentekanaal) with a continuous supply of methane, nitrite, and nitrate (7). A subculture with nitrate as the only electron acceptor in the mineral medium resulted in a microbial community dominated by archaea [ca. 40–50%, belonging to the AAA cluster, 95% similar to Methanoperedens nitroreducens (3), and previously referred to as M. nitroreducens MPEBLZ (24)] and a microorganism closely related to M. oxyfera (ca. 40%) as judged by FISH microscopy (Fig. 1). The enrichment culture coupled methane oxidation [assessed as production of 13CO2, 180 nmol mg(protein)−1 h−1] to the reduction of nitrate [360 nmol mg(protein)−1 h−1], with dinitrogen gas (N2) as the main end product [153 nmol mg(protein)−1 h−1] (Fig. 2A), similar to the theoretical stoichiometry:
| [1] |
The discrepancy (5:10 instead of 5:8) is most likely due to carbon assimilation into biomass and the conversion of about 10% of nitrate to ammonium [2] (Fig. 2B):
| [2] |
This observation adds a new aspect to the current understanding of AAA physiology (5): In the absence of ammonium-oxidizing bacteria, ammonium production in our enrichment culture could be measured. Despite M. oxyfera-like bacteria reducing nitrite to N2 a transient accumulation of nitrite was also often observed, although it was consumed when nitrate became limiting (Fig. 2B). Nitrate reduction in the enrichment culture was largely (>95%) dependent on methane. All genes necessary for nitrate reduction to nitrite and nitrite reduction to ammonium are present in the genome of AAA (5, 24). Together, these results suggest that AAA most likely couples methane oxidation to dissimilatory nitrate reduction to ammonium via nitrite, a trait rarely observed in archaea (25).
Fig. 1.
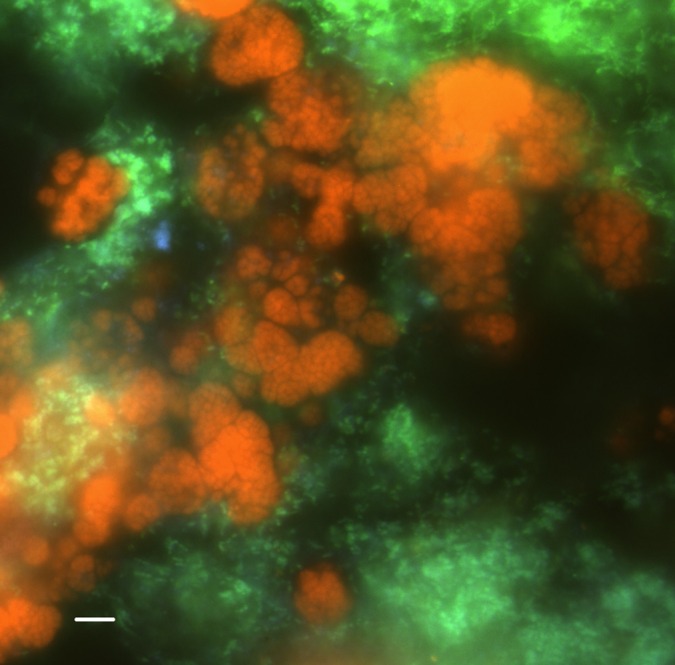
Fluorescence in situ hybridization of biomass from the enrichment culture of AAA and M. oxyfera-like bacteria. AAA were hybridized with Cy3-labeled AAA-specific probe S-*-AAA-FW-641 (red) (33); M. oxyfera-like bacteria were hybridized with Fluos-labeled NC10 bacteria-specific probe S-*-DBACT-1027-a-A-18 (yellow) and a Cy5-labeled mixture of EUB I to III and V (blue; overlay appears green). Other bacteria only hybridizing with EUB I to III and V appear blue. (Scale bar, 5 µm.)
Fig. 2.

(A) Stoichiometric coupling of [15N]nitrate reduction (open squares) to 30N2 (open triangles) and 13CO2 (filled circles) production in the presence of 13CH4 in batch incubations. Data of CO2 and N2 are the average of four replicate incubations measured twice each; nitrate values represent two replicates. Each incubation contained 77.0 ± 3.6 μg protein. Error bars indicate SD. (B) Transient formation of nitrite (open diamonds) and accumulation of ammonium (open circles) after repeated pulse feeding (arrows) of nitrate (open squares) in the continuous culture reactor with constant methane supply. Note the different scale for nitrite.
We incubated this enrichment culture with AAA as the only archaea with a range of oxidized metal compounds. In the absence of an electron acceptor in batch incubations, methane oxidation did not occur, but activity began immediately after addition of ferric citrate (Fig. 3A). Recently, using a sample that contained ANME-2a and ANME-2c, Scheller et al. reported 13CO2 production from 13CH4 oxidation in the presence of ferric citrate (22). Due to the notorious difficulties associated with measuring dissolved ferrous (Fe2+) iron in complex media or natural samples, the stoichiometric relationship between the reduction of Fe3+ and oxidation of CH4 was not shown (22). Here, in a setup with destructive sampling, where total CO2 and Fe2+ could both be measured, we observed a near-perfect fit to the theoretically expected stoichiometry of 1:8 (Fig. 3B):
| [3] |
In nature, iron reduction is often limited by the bioavailability of iron oxides, and soluble or complexed Fe3+ is scarce (19, 23). Therefore, we also incubated the enrichment culture with more environmentally relevant forms of oxidized metals: nanoparticulate ferrihydrite (Fe3+) and birnessite (Mn4+). These were suitable as electron acceptors for the oxidation of methane to CO2 by the enrichment culture, albeit at lower rates (Fig. 3C): 6 and 21 nmol CO2 mg(protein)−1 h−1, compared with 80 and 180 nmol CO2 mg(protein)−1 h−1, with soluble Fe3+ (ferric citrate) and nitrate as electron acceptors, respectively.
Fig. 3.

Metal-dependent oxidation of 13C-labeled CH4 to 13CO2 by an enrichment culture of AAA. (A) Fe3+-citrate addition at the start of the experiment and after 150 h (arrow) stimulates methane oxidation. Control incubations of the enrichment culture without any addition, sulfate or citrate only, as well as an archaea-free enrichment culture of M. oxyfera (34) incubated with Fe3+-citrate were not active. Data are the average of two incubations containing 1 ± 0.1 mg protein each. (B) Stoichiometric coupling of CH4 oxidation to Fe2+ production. Methane oxidation was deduced from 13CO2 production; data points represent the average of four incubations of 77 ± 3.6 µg protein each. (C) Stimulation of 13CO2 production from 13CH4 by the addition of nanoparticulate suspensions of ferrihydrite (Fe3+) and birnessite (Mn4+). Protein content in these experiments was 77.0 ± 3.6 µg per vial. Error bars indicate SD.
Negative controls with ambient sulfate concentrations (∼15 µM) or added sulfate (1 mM) (Fig. 3A) excluded the possibility of a cryptic sulfur cycle (19, 20) as an indirect route to (re)generate electron acceptors (sulfate). Such a mechanism would also not fit the observed stoichiometry [3]. In addition, iron reduction was dependent on the presence of methane: In controls without methane (<15 ppm), the enrichment culture did not produce any Fe2+ (above the background of ∼2 µM remaining from 3.75 µM FeSO4 supplied as a micronutrient), the end product of Fe3+ reduction in incubations with methane. Finally, an enrichment culture containing M. oxyfera as the only methanotroph (26) did not oxidize methane coupled to the reduction of Fe3+. Taken together, these findings strongly indicated that the AAA in the enrichment culture directly coupled methane oxidation to Fe3+ and Mn4+ reduction.
AAA and an M. oxyfera-like microorganism dominated our enrichment culture (Fig. 1). The analyses of the 16S rRNA gene reads of the metagenome (DDBJ/EMBL/GenBank accession no. LKCM01000000) revealed that other archaea or known iron-reducing microorganisms [Geobacter or Shewanella (27)] were absent from the enrichment culture; this was expected, as the culture had never been exposed to iron exceeding the above-mentioned 3.75 µM FeSO4 in the medium. Furthermore, the immediate onset of Fe3+ reduction in batch incubations and its linear course excluded the growth of an iron-reducing subpopulation interacting with a methanotroph being responsible for AOM in our culture.
Rather, the analysis of the partially reconstructed AAA genome (24) supported that the AAA alone were able to couple methane oxidation to the reduction of different electron acceptors. All genes of the reverse methanogenesis pathway (28, 29) were present in the enriched strain of AAA, in complete agreement with its closest relative, M. nitroreducens (5, 24). Nitrate reduction could be accomplished by two candidate enzyme complexes, one encoded by the narGH genes (MPEBLZ_02035 and 2036), closely related to the ones of M. nitroreducens (81 and 83% identity, respectively), and the other by an nxrAB-related operon (MPEBLZ_01127 and 1129) absent from M. nitroreducens and similar to that in nitrite-oxidizing bacteria (68/78% similarity to Nitrolancea hollandica and 68/69% to Nitrobacter winogradskyi). The further reduction of nitrite to N2 gas could be accomplished by the M. oxyfera-like microorganism as previously demonstrated (6) or, alternatively, ammonium could be formed by a NrfAH-like protein complex (MPEBLZ_01114 and 1115) transcribed by the AAA (24). As the likely genetic basis for Fe3+ reduction, we identified a wealth of genes encoding heme c-containing proteins (c-type cytochromes), which are generally recognized to function as electron shuttles from the cell to soluble or solid electron acceptors such as iron minerals (27, 30). The AAA in our enrichment culture encode numerous (41 species) multiheme c-type cytochromes (Table S1). This number is much higher than for methanogens, of which only Methanosarcinales encodes few multiheme cytochromes, but similar to M. nitroreducens (38 species) (5), the sulfate-dependent ANME-1 (11 species) (28), ANME-2a (16 species) (29, 31), as well as the iron-reducing archaea Ferroglobus placidus (22 species) and Geoglobus (15 species) (32).
Table S1.
Multi(>2)-heme c-type cytochromes predicted from the contigs assigned to AAA in the metagenome of the enrichment culture
| Locus tag | Protein length, no. of amino acids | CxxCH and other motifs | No. of transmembrane helices | Best hit identifier | Best hit organism | Best hit identity |
| MPEBLZ_00008 | 358 | 4 | 0 | gi|851163073|ref|WP_048092705.1| | Candidatus Methanoperedens nitroreducens | 44.5 |
| MPEBLZ_00010 | 190 | 6 | 1 | gi|851163076|ref|WP_048092707.1| | Ca. M. nitroreducens | 60.5 |
| MPEBLZ_00012 | 319 | 4 | 1 | gi|851163096|ref|WP_048092715.1| | Ca. M. nitroreducens | 61.2 |
| MPEBLZ_00015 | 253 | 7 | 1 | gi|851163104|ref|WP_048092719.1| | Ca. M. nitroreducens | 60.8 |
| MPEBLZ_00016 | 343 | 4 | 0 | gi|851163073|ref|WP_048092705.1| | Ca. M. nitroreducens | 45.3 |
| MPEBLZ_00238 | 317 | 11 | 2 | gi|630830727|gb|KCZ72437.1| | Ca. M. nitroreducens | 60.6 |
| MPEBLZ_00452 | 577 | 9 | 0 | gi|851163118|ref|WP_048092727.1| | Ca. M. nitroreducens | 50.4 |
| MPEBLZ_00526 | 864 | 16 | 1 | gi|851162463|ref|WP_048092373.1| | Ca. M. nitroreducens | 37.9 |
| MPEBLZ_00803 | 1,846 | 7* | 1 | gi|851163104|ref|WP_048092719.1| | Ca. M. nitroreducens | 54.5 |
| MPEBLZ_00816 | 316 | 6† | 0 | gi|851164534|ref|WP_048093459.1| | Ca. M. nitroreducens | 71.9 |
| MPEBLZ_00817 | 309 | 6† | 2 | gi|851164537|ref|WP_048093461.1| | Ca. M. nitroreducens | 63.2 |
| MPEBLZ_01070 | 678 | 9‡ | 0 | gi|851160641|ref|WP_048091456.1| | Ca. M. nitroreducens | 60.6 |
| MPEBLZ_01114 | 437 | 5* | 1 | gi|851165424|ref|WP_048093932.1| | Ca. M. nitroreducens | 74.5 |
| MPEBLZ_01115 | 167 | 4 | 0 | gi|630828680|gb|KCZ70396.1| | Ca. M. nitroreducens | 61.1 |
| MPEBLZ_01126 | 288 | 2 | 2 | gi|490648195|ref|WP_004513190.1| | Geobacter metallireducens | 43.4 |
| MPEBLZ_01329 | 202 | 5 | 1 | gi|851154723|ref|WP_048088734.1| | Ca. M. nitroreducens | 57.4 |
| MPEBLZ_01361 | 1,137 | 16 | 1 | gi|851162463|ref|WP_048092373.1| | Ca. M. nitroreducens | 40.3 |
| MPEBLZ_01741 | 696 | 21 | 0 | gi|851163401|ref|WP_048092876.1| | Geoglobus acetivorans | 28.6 |
| MPEBLZ_01742 | 561 | 11 | 0 | gi|851155354|ref|WP_048089050.1| | Ca. M. nitroreducens | 31.9 |
| MPEBLZ_01743 | 914 | 18 | 1 | gi|851149563|ref|WP_048086421.1| | Ferroglobus placidus | 30.0 |
| MPEBLZ_01744 | 606 | 6†,‡ | 1 | gi|851163118|ref|WP_048092727.1| | Ca. M. nitroreducens | 59.3 |
| MPEBLZ_01877 | 828 | 16‡ | 1 | gi|851163401|ref|WP_048092876.1| | G. acetivorans | 38.2 |
| MPEBLZ_01950 | 432 | 8 | 0 | gi|851165403|ref|WP_048093919.1| | Ca. M. nitroreducens | 69.1 |
| MPEBLZ_02010 | 573 | 6 | 0 | gi|851163118|ref|WP_048092727.1| | Ca. M. nitroreducens | 51.1 |
| MPEBLZ_02042 | 403 | 5 | 2 | gi|851156284|ref|WP_048089356.1| | Ca. M. nitroreducens | 67.5 |
| MPEBLZ_02500 | 373 | 7 | 1 | gi|851153793|ref|WP_048088239.1| | Ca. M. nitroreducens | 74.2 |
| MPEBLZ_02501 | 179 | 5 | 0 | gi|851153793|ref|WP_048088239.1| | Ca. M. nitroreducens | 67.1 |
| MPEBLZ_02503 | 469 | 8 | 0 | gi|851153795|ref|WP_048088240.1| | Ca. M. nitroreducens | 69.1 |
| MPEBLZ_02608 | 209 | 5 | 0 | gi|851160641|ref|WP_048091456.1| | Ca. M. nitroreducens | 51.5 |
| MPEBLZ_02874 | 305 | 7‡ | 2 | gi|630831567|gb|KCZ73276.1| | Ca. M. nitroreducens | 41.3 |
| MPEBLZ_02879 | 267 | 7 | 2 | gi|851162380|ref|WP_048092326.1| | Ca. M. nitroreducens | 54.6 |
| MPEBLZ_03194 | 318 | 6† | 1 | gi|630831820|gb|KCZ73529.1| | Ca. M. nitroreducens | 71.2 |
| MPEBLZ_03195 | 149 | 4‡ | 1 | gi|630831821|gb|KCZ73530.1| | Ca. M. nitroreducens | 70.5 |
| MPEBLZ_03279 | 655 | 2 | 1 | gi|635652775|ref|WP_024299646.1| | Methylosarcina lacus | 68.3 |
| MPEBLZ_03539 | 237 | 5‡ | 1 | gi|630831567|gb|KCZ73276.1| | Ca. M. nitroreducens | 78.1 |
| MPEBLZ_04272 | 521 | 12* | 1 | gi|851149560|ref|WP_048086419.1| | F. placidus | 27.5 |
| MPEBLZ_04274 | 227 | 4 | 1 | gi|851166986|ref|WP_048094540.1| | Geoglobus ahangari | 35.6 |
| MPEBLZ_04276 | 224 | 5 | 1 | gi|851163417|ref|WP_048092885.1| | G. acetivorans | 27.7 |
| MPEBLZ_04299 | 324 | 6† | 2 | gi|851155273|ref|WP_048089031.1| | Ca. M. nitroreducens | 61.9 |
| MPEBLZ_04300 | 364 | 13† | 1 | gi|851155276|ref|WP_048089032.1| | Ca. M. nitroreducens | 73.0 |
| MPEBLZ_04301 | 246 | 2 | 1 | gi|258591543|emb|CBE67844.1| | Ca. Methylomirabilis oxyfera | 26.2 |
Prediction of multiheme cytochrome c proteins was done using a perl script (prot_motif_search.pl) available at https://github.com/dspeth followed by BLAST analysis of conservation of the heme-binding motifs in related sequences. Prediction of transmembrane helices was performed using TMHMM (www.cbs.dtu.dk/services/TMHMM/) (49). Each superscript denotes one divergent heme binding motif.
CxxCK.
CxxxCH.
CxxxxCH.
This genomic blueprint suggests that AAA, and possibly also the cytochrome-rich ANME-1 (28) and ANME-2a (29, 31) archaea, known for sulfate-dependent methane oxidation, could be versatile methanotrophs that are able to switch between different electron acceptors, depending on environmental conditions (17). It is also conceivable that ANME-2a and ANME-2c archaea, which were shown to couple the reduction of solubilized iron species (iron citrate) to methane oxidation recently (22), could also be capable of converting particulate Fe3+ or Mn4+. Compared with M. oxyfera, the ecological niche of AAA probably extends into more reduced environments, as these microorganisms can also use oxidized metals. However, AAA can form stable cocultures with microorganisms related to the nitrite-dependent methanotroph M. oxyfera, as previously reported (7). While AAA supply nitrite, which is often a limiting electron acceptor in nature, to M. oxyfera-like bacteria, or as in the culture described by ref. 5, to anaerobic ammonium-oxidizing (anammox) bacteria, the bacterial partner in these cocultures keeps the concentration of the potentially toxic nitrite low. A bioreactor containing all three microorganisms could be applied in wastewater treatment to simultaneously convert ammonium, methane, and oxidized nitrogen species to harmless N2 and CO2.
Molecular surveys of 16S rRNA or mcrA genes in the environment have often retrieved AAA-affiliated sequences in ecosystems poor in nitrate but rich in oxidized metals (14, 33) (Fig. S1), so far without an ecophysiological explanation. Our results show that the AAA directly coupled the reduction of these environmentally relevant, particulate oxidized metal species to methane oxidation and could play an as-yet largely overlooked role in decreasing methane emissions from anoxic environments to the atmosphere. Given the 8:1 stoichiometry of Fe3+ reduced to methane oxidized [3], their contribution to iron cycling, thereby impacting also the geochemistry of phosphorus and sulfur, may be even more important than their impact on the carbon cycle (11, 16, 18, 20). The enrichment of at least one of the microbial players not only now enables a targeted investigation of iron-dependent methane oxidation in physiological and molecular detail, but may also shed light on the long-standing discussion about Fe2+-producing processes on early Earth, when AAA-related organisms may have thrived under the methane-rich atmosphere in the ferruginous Archean oceans (12, 18, 30).
Fig. S1.

(A) Phylogenetic position of AOM-associated archaea (AAA) (red) among other archaeal methanotrophs (blue) and methanogens (gray) based on sequences of 16S rRNA gene sequences obtained from GenBank. (B) Detailed phylogenetic tree of 16S rRNA genes of the AAA group within Methanosarcinales. Sequences from enrichment cultures are highlighted in bold letters; sequences from sites or cultures with demonstrated AOM activity are shown in blue; those from particularly metal-rich or iron-reducing environments are shown in orange. The evolutionary analysis was conducted with the maximum-likelihood algorithm implemented in MEGA6 (39). There were a total of 949 positions in the final dataset. Bootstrap support >90 is shown at the branches. (Scale bar, number of substitutions per site.)
Methods
Enrichment of the Coculture of AAA and the M. oxyfera-like Bacteria.
The enrichment of the biomass used here was described in brief previously (24). Biomass from the settler of an M. oxyfera-dominated reactor (26) containing about 10% (of the total microbial community) AAA was used to inoculate a continuous culture reactor (2-L working volume) to further enrich AAA. The reactor was run at room temperature with a pH of 7.5 (±0.15) and stirred gently with a magnetic bar (100 rpm). Initially, the reactor was coupled to the effluent of an M. oxyfera reactor, receiving once per day the methane-saturated effluent containing low nitrite (0 to 50 µM) and high nitrate (2 to 3 mM) concentrations. Along with the growth of biomass in the reactor, nitrite and nitrate consumption rates increased. Therefore, increasing amounts of nitrate were manually injected into the reactor (from 100 to 1,000 µM final concentration) when nitrite and nitrate were below 100 µM. After about 1 year of incubation, the enrichment culture was uncoupled from the M. oxyfera reactor. Methane was provided by constant sparging with CH4:CO2 (95:5, vol:vol; ca. 10 mL⋅min−1). Nitrate (0.5 to 1.5 mM final concentration) as the sole electron acceptor was injected daily into the reactor for over 2 years. Supernatant (ca. 0.8 L) was removed and replenished with fresh anoxic mineral medium (as previously described by ref. 34, omitting nitrate and nitrite) after settling of biomass every 2 weeks.
Nitrate Reduction and Nitrite and Ammonium Production by the Enrichment Culture.
Conversion of nitrogenous compounds was measured in the continuous culture while sparging the reactor with CH4:CO2 (95:5, vol:vol; ca. 10 mL⋅min−1). After nitrate depletion (to <10 µM), fresh nitrate was added to a final concentration of ∼1 mM on 3 consecutive days. Liquid samples (1.5 mL) were taken every 3 to 4 h from the reactor and centrifuged at 14,000 × g, and supernatants were stored at 4 °C before nitrate, nitrite, and ammonium analysis.
Activity Tests with Different Electron Acceptors.
To test the effect of various electron acceptors, the enrichment culture was left under continuous CH4:CO2 (95:5, vol:vol) sparging but without nitrate supply until residual nitrate was depleted. Twenty milliliters of culture liquid (protein content ca. 1.0 mg) was taken and anaerobically transferred to a sealed, evacuated serum bottle (50 mL). After removal of unlabeled methane by flushing with Ar:CO2 (90:10, vol:vol), 10 mL 13CH4 (>99.99% 13C; Sigma-Aldrich) was injected into the 1-bar Ar:CO2 headspace. Ferric citrate (pH 6.8) and sodium sulfate were added from anoxic stock solutions to final concentrations of 2 and 1 mM, respectively. Particulate iron and manganese were added as ferrihydrite or birnessite nanoparticles from anoxic suspensions. The incubation of nitrite-depleted M. oxyfera biomass (34) with ferric citrate was prepared in the same manner as described above. To control for the effect of electron acceptor addition, additional incubations were prepared accordingly, but either without any additions or with sodium citrate only (2 mM final concentration). All serum bottles were incubated at room temperature and shaken gently at 50 rpm. Intervals between headspace sampling for concentration and isotopic composition of CH4 and CO2 were chosen according to the observed activity.
Stoichiometry Determination in Activity Tests with Destructive Sampling.
To be able to precisely determine the stoichiometry of methane oxidation coupled to nitrate or Fe3+ reduction and minimize the biomass needed per experiment, the experimental setup was miniaturized and incubations were performed in 3-mL Exetainer vials (Labco). Before distribution of biomass over the vials, residual methane was stripped from nitrate-depleted enrichment culture by stirring and concomitant flushing with He:CO2 (90:10, vol:vol) in a previously evacuated and sealed serum bottle. The biomass was then transferred to an anaerobic chamber with an Ar:H2 (95:5, vol:vol) atmosphere (O2 in the anaerobic chamber was removed by passing of the gas over a Pd catalyst resulting in a residual O2 concentration of ≤0.2 ppm) and amended with an electron acceptor (nitrate or ferric citrate). The biomass was then equally distributed to aliquots of 1.5 mL in Exetainers, resulting in a protein content of 77.0 ± 3.6 µg per vial. Exetainers were sealed and 1 mL of 13CH4:CO2 (80:20, vol:vol) was injected into the headspace to provide an electron donor, to buffer pH, and create overpressure. The vials were incubated at room temperature in an overhead shaker inside the anaerobic cabinet. For incubations with ferric citrate, three vials were harvested every 3 h and acidified with 0.5 mL anaerobic HCl solution (final concentration 0.5 mM) and kept in the anaerobic tent for at least 0.5 h before GC/MS measurement to allow CO2 to equilibrate with the headspace. After gas measurements, biomass from each vial was transferred to a 1.5-mL tube and centrifuged, and the supernatant was stored at 4 °C until Fe2+ determination. For incubations with nitrate, four vials were harvested every 2 h, two of which were acidified with 0.5 mL anaerobic HCl solution. CO2 production in all four vials’ headspace was measured. The biomass from the nonacidified vials was transferred to a 1.5-mL cup and centrifuged, and supernatant was stored at 4 °C until nitrate determination by HPLC.
Analytical Procedures and Synthesis of Reagents.
Quantity and isotopic composition of CH4, CO2, and N2 in the headspace were measured by GC/MS (GC 6890 coupled to MSD 5975C; Agilent) as previously described (34). Total amounts indicated in the incubations take into account the dissolved fraction of the respective gases. Nitrate and nitrite were measured after a reduction step to NO with a Sievers Nitric Oxide Analyzer (NOA 280i; Analytix). Nitrate and nitrite were both reduced with a saturated solution of vanadium (III) chloride in hydrochloric acid (1 M) at 90 °C. Nitrite was colorimetrically measured by the Griess reaction as previously described (35). Nitrate concentrations were obtained by subtracting the nitrite concentration from the sum of nitrate and nitrite. Ammonium was measured colorimetrically at 623 nm after a 30-min reaction of 100-µL samples with alkaline phenol catalyzed by sodium nitroprusside at 37 °C (36). For the miniaturized setup (Exetainer incubations), nitrate was quantified with HPLC using a conductivity detector (26). Fe2+ was measured using the ferrozine method (37).
Ferrihydrite and birnessite were synthesized according to protocols described previously (23, 38). Briefly, for ferrihydrite, 21 g of ferric citrate (Sigma) was dissolved in 300 mL MilliQ water by vigorous stirring while being kept at 80 °C. After cooling the ferric citrate solution to room temperature, the pH was quickly adjusted to 8.0 with 10 M NaOH while stirring rigorously for 30 min. The supernatant of the resulting suspension was discarded after centrifuging at 8,000 × g at 4 °C for 1 h, and the pellet was resuspended in 500 mL MilliQ water. After four washing cycles, the pellet was finally resuspended in a small volume of MilliQ water. For birnessite, concentrated HCl (46%) was slowly added to a boiling and rigorously stirring 0.4 M potassium permanganate solution to a final concentration of 0.8 M in a fume hood. After further boiling and stirring for 10 min, the suspension was cooled to room temperature and centrifuged. Washing and resuspension were performed as described for ferrihydrite. Approximate iron and manganese concentrations in the suspensions were determined by measuring the dry weight of an aliquot.
Fluorescence in Situ Hybridization.
Biomass from the enrichment culture (2 mL) was harvested by centrifugation, fixed, hybridized, and visualized as previously described (26) with additional use of the probe S-*-AAA-FW-641 (33), specifically targeting AAA.
Acknowledgments
We thank Francisca A. Luesken for starting and initially maintaining the AAA enrichment culture. K.F.E. was supported by a VENI grant from The Netherlands Organization for Scientific Research (863.13.007), B.Z. by the joint program of the Chinese Academy of Science/Royal Dutch Academy of Science (09PhD02), D.S. by the BE-Basic Foundation (fp07.002), M.S.M.J. by the European Research Council (ERC 339880) and the Spinoza Prize, and B.K. by a VENI grant from The Netherlands Organization for Scientific Research (863.11.003) and the European Research Council (ERC 640422).
Footnotes
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
Data deposition: The sequence reported in this paper has been deposited in the DDBJ/EMBL/GenBank database (metagenome accession no. LKCM01000000).
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1609534113/-/DCSupplemental.
References
- 1.Trotsenko YA, Murrell JC. Metabolic aspects of aerobic obligate methanotrophy. In: Laskin AI, Sariaslani S, Gadd GM, editors. Advances in Applied Microbiology. Vol 63. Academic; Cambridge, MA: 2008. pp. 183–229. [DOI] [PubMed] [Google Scholar]
- 2.Knittel K, Boetius A. Anaerobic oxidation of methane: Progress with an unknown process. Annu Rev Microbiol. 2009;63:311–334. doi: 10.1146/annurev.micro.61.080706.093130. [DOI] [PubMed] [Google Scholar]
- 3.Myhre G, et al. Anthropogenic and natural radiative forcing. In: Stocker TF, et al., editors. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge Univ Press; Cambridge, UK: 2014. pp. 659–740. [Google Scholar]
- 4.Milucka J, et al. Zero-valent sulphur is a key intermediate in marine methane oxidation. Nature. 2012;491(7425):541–546. doi: 10.1038/nature11656. [DOI] [PubMed] [Google Scholar]
- 5.Haroon MF, et al. Anaerobic oxidation of methane coupled to nitrate reduction in a novel archaeal lineage. Nature. 2013;500(7464):567–570. doi: 10.1038/nature12375. [DOI] [PubMed] [Google Scholar]
- 6.Ettwig KF, et al. Nitrite-driven anaerobic methane oxidation by oxygenic bacteria. Nature. 2010;464(7288):543–548. doi: 10.1038/nature08883. [DOI] [PubMed] [Google Scholar]
- 7.Raghoebarsing AA, et al. A microbial consortium couples anaerobic methane oxidation to denitrification. Nature. 2006;440(7086):918–921. doi: 10.1038/nature04617. [DOI] [PubMed] [Google Scholar]
- 8.Martin J-M, Meybeck M. Elemental mass-balance of material carried by major world rivers. Mar Chem. 1979;7(3):173–206. [Google Scholar]
- 9.Norði Kà, Thamdrup B, Schubert CJ. Anaerobic oxidation of methane in an iron-rich Danish freshwater lake sediment. Limnol Oceanogr. 2013;58(2):546–554. [Google Scholar]
- 10.Sivan O, et al. Geochemical evidence for iron-mediated anaerobic oxidation of methane. Limnol Oceanogr. 2011;56(4):1536–1544. [Google Scholar]
- 11.Torres NT, et al. Early diagenetic processes generate iron and manganese oxide layers in the sediments of Lake Baikal, Siberia. Environ Sci Process Impacts. 2014;16(4):879–889. doi: 10.1039/c3em00676j. [DOI] [PubMed] [Google Scholar]
- 12.Crowe SA, et al. The methane cycle in ferruginous Lake Matano. Geobiology. 2011;9(1):61–78. doi: 10.1111/j.1472-4669.2010.00257.x. [DOI] [PubMed] [Google Scholar]
- 13.Amos RT, et al. Evidence for iron-mediated anaerobic methane oxidation in a crude oil-contaminated aquifer. Geobiology. 2012;10(6):506–517. doi: 10.1111/j.1472-4669.2012.00341.x. [DOI] [PubMed] [Google Scholar]
- 14.Chang YH, et al. Microbial methane cycling in a terrestrial mud volcano in eastern Taiwan. Environ Microbiol. 2012;14(4):895–908. doi: 10.1111/j.1462-2920.2011.02658.x. [DOI] [PubMed] [Google Scholar]
- 15.Beal EJ, House CH, Orphan VJ. Manganese- and iron-dependent marine methane oxidation. Science. 2009;325(5937):184–187. doi: 10.1126/science.1169984. [DOI] [PubMed] [Google Scholar]
- 16.Egger M, et al. Iron-mediated anaerobic oxidation of methane in brackish coastal sediments. Environ Sci Technol. 2015;49(1):277–283. doi: 10.1021/es503663z. [DOI] [PubMed] [Google Scholar]
- 17.Wankel SD, et al. Anaerobic methane oxidation in metalliferous hydrothermal sediments: Influence on carbon flux and decoupling from sulfate reduction. Environ Microbiol. 2012;14(10):2726–2740. doi: 10.1111/j.1462-2920.2012.02825.x. [DOI] [PubMed] [Google Scholar]
- 18.Riedinger N, et al. An inorganic geochemical argument for coupled anaerobic oxidation of methane and iron reduction in marine sediments. Geobiology. 2014;12(2):172–181. doi: 10.1111/gbi.12077. [DOI] [PubMed] [Google Scholar]
- 19.Hansel CM, et al. Dominance of sulfur-fueled iron oxide reduction in low-sulfate freshwater sediments. ISME J. 2015;9(11):2400–2412. doi: 10.1038/ismej.2015.50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Holmkvist L, Ferdelman TG, Jørgensen BB. A cryptic sulfur cycle driven by iron in the methane zone of marine sediment (Aarhus Bay, Denmark) Geochim Cosmochim Acta. 2011;75(12):3581–3599. [Google Scholar]
- 21.Nauhaus K, Treude T, Boetius A, Krüger M. Environmental regulation of the anaerobic oxidation of methane: A comparison of ANME-I and ANME-II communities. Environ Microbiol. 2005;7(1):98–106. doi: 10.1111/j.1462-2920.2004.00669.x. [DOI] [PubMed] [Google Scholar]
- 22.Scheller S, Yu H, Chadwick GL, McGlynn SE, Orphan VJ. Artificial electron acceptors decouple archaeal methane oxidation from sulfate reduction. Science. 2016;351(6274):703–707. doi: 10.1126/science.aad7154. [DOI] [PubMed] [Google Scholar]
- 23.Bosch J, Heister K, Hofmann T, Meckenstock RU. Nanosized iron oxide colloids strongly enhance microbial iron reduction. Appl Environ Microbiol. 2010;76(1):184–189. doi: 10.1128/AEM.00417-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Arshad A, et al. A metagenomics-based metabolic model of nitrate-dependent anaerobic oxidation of methane by Methanoperedens-like archaea. Front Microbiol. 2015;6:1423. doi: 10.3389/fmicb.2015.01423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Simon J, Klotz MG. Diversity and evolution of bioenergetic systems involved in microbial nitrogen compound transformations. Biochim Biophys Acta. 2013;1827(2):114–135. doi: 10.1016/j.bbabio.2012.07.005. [DOI] [PubMed] [Google Scholar]
- 26.Ettwig KF, et al. Denitrifying bacteria anaerobically oxidize methane in the absence of Archaea. Environ Microbiol. 2008;10(11):3164–3173. doi: 10.1111/j.1462-2920.2008.01724.x. [DOI] [PubMed] [Google Scholar]
- 27.Shi L, Squier TC, Zachara JM, Fredrickson JK. Respiration of metal (hydr)oxides by Shewanella and Geobacter: A key role for multihaem c-type cytochromes. Mol Microbiol. 2007;65(1):12–20. doi: 10.1111/j.1365-2958.2007.05783.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Meyerdierks A, et al. Metagenome and mRNA expression analyses of anaerobic methanotrophic archaea of the ANME-1 group. Environ Microbiol. 2010;12(2):422–439. doi: 10.1111/j.1462-2920.2009.02083.x. [DOI] [PubMed] [Google Scholar]
- 29.Wang F-P, et al. Methanotrophic archaea possessing diverging methane-oxidizing and electron-transporting pathways. ISME J. 2014;8(5):1069–1078. doi: 10.1038/ismej.2013.212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Weber KA, Achenbach LA, Coates JD. Microorganisms pumping iron: Anaerobic microbial iron oxidation and reduction. Nat Rev Microbiol. 2006;4(10):752–764. doi: 10.1038/nrmicro1490. [DOI] [PubMed] [Google Scholar]
- 31.McGlynn SE, Chadwick GL, Kempes CP, Orphan VJ. Single cell activity reveals direct electron transfer in methanotrophic consortia. Nature. 2015;526(7574):531–535. doi: 10.1038/nature15512. [DOI] [PubMed] [Google Scholar]
- 32.Kletzin A, et al. Cytochromes c in Archaea: Distribution, maturation, cell architecture, and the special case of Ignicoccus hospitalis. Front Microbiol. 2015;6:439. doi: 10.3389/fmicb.2015.00439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Schubert CJ, et al. Evidence for anaerobic oxidation of methane in sediments of a freshwater system (Lago di Cadagno) FEMS Microbiol Ecol. 2011;76(1):26–38. doi: 10.1111/j.1574-6941.2010.01036.x. [DOI] [PubMed] [Google Scholar]
- 34.Ettwig KF, van Alen T, van de Pas-Schoonen KT, Jetten MSM, Strous M. Enrichment and molecular detection of denitrifying methanotrophic bacteria of the NC10 phylum. Appl Environ Microbiol. 2009;75(11):3656–3662. doi: 10.1128/AEM.00067-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Kartal B, et al. Adaptation of a freshwater anammox population to high salinity wastewater. J Biotechnol. 2006;126(4):546–553. doi: 10.1016/j.jbiotec.2006.05.012. [DOI] [PubMed] [Google Scholar]
- 36.Weatherburn M. Phenol-hypochlorite reaction for determination of ammonia. Anal Chem. 1967;39(8):971–974. [Google Scholar]
- 37.Viollier E, Inglett PW, Hunter K, Roychoudhury AN, Van Cappellen P. The ferrozine method revisited: Fe(II)/Fe(III) determination in natural waters. Appl Geochem. 2000;15(6):785–790. [Google Scholar]
- 38.McKenzie R. The synthesis of birnessite, cryptomelane, and some other oxides and hydroxides of manganese. Mineral Mag. 1971;38(296):493–502. [Google Scholar]
- 39.Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol Biol Evol. 2013;30(12):2725–2729. doi: 10.1093/molbev/mst197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Kato S, et al. Prokaryotic abundance and community composition in a freshwater iron-rich microbial mat at circumneutral pH. Geomicrobiol J. 2012;29(10):896–905. [Google Scholar]
- 41.MacLean L, et al. Mineralogical, chemical and biological characterization of an anaerobic biofilm collected from a borehole in a deep gold mine in South Africa. Geomicrobiol J. 2007;24(6):491–504. [Google Scholar]
- 42.Niemann H, et al. Biogeochemistry of a low-activity cold seep in the Larsen B area, western Weddell Sea, Antarctica. Biogeosciences. 2009;6(11):2383–2395. [Google Scholar]
- 43.Rastogi G, Sani RK, Peyton BM, Moberly JG, Ginn TR. Molecular studies on the microbial diversity associated with mining-impacted Coeur d’Alene River sediments. Microb Ecol. 2009;58(1):129–139. doi: 10.1007/s00248-008-9445-0. [DOI] [PubMed] [Google Scholar]
- 44.Schauer R, et al. Bacterial sulfur cycling shapes microbial communities in surface sediments of an ultramafic hydrothermal vent field. Environ Microbiol. 2011;13(10):2633–2648. doi: 10.1111/j.1462-2920.2011.02530.x. [DOI] [PubMed] [Google Scholar]
- 45.Stein LY, La Duc MT, Grundl TJ, Nealson KH. Bacterial and archaeal populations associated with freshwater ferromanganous micronodules and sediments. Environ Microbiol. 2001;3(1):10–18. doi: 10.1046/j.1462-2920.2001.00154.x. [DOI] [PubMed] [Google Scholar]
- 46.Zhang G, et al. Methanogen community in Zoige wetland of Tibetan plateau and phenotypic characterization of a dominant uncultured methanogen cluster ZC-I. Environ Microbiol. 2008;10(7):1850–1860. doi: 10.1111/j.1462-2920.2008.01606.x. [DOI] [PubMed] [Google Scholar]
- 47.Ding Z-W, Ding J, Fu L, Zhang F, Zeng RJ. Simultaneous enrichment of denitrifying methanotrophs and anammox bacteria. Appl Microbiol Biotechnol. 2014;98(24):10211–10221. doi: 10.1007/s00253-014-5936-8. [DOI] [PubMed] [Google Scholar]
- 48.Kasai Y, Takahata Y, Hoaki T, Watanabe K. Physiological and molecular characterization of a microbial community established in unsaturated, petroleum-contaminated soil. Environ Microbiol. 2005;7(6):806–818. doi: 10.1111/j.1462-2920.2005.00754.x. [DOI] [PubMed] [Google Scholar]
- 49.Krogh A, Larsson B, von Heijne G, Sonnhammer EL. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J Mol Biol. 2001;305(3):567–580. doi: 10.1006/jmbi.2000.4315. [DOI] [PubMed] [Google Scholar]