

Abstract
The present study aimed to investigate the in vitro mutagenic activity of Origanum majorana essential oil. The most abundant compounds identified by GC-MS were γ-terpinene (25.73%), α-terpinene (17.35%), terpinen-4-ol (17.24%), and sabinene (10.8%). Mutagenicity was evaluated by the Salmonella/microsome test using the preincubation procedure on TA98, TA97a, TA100, TA102, and TA1535 Salmonella typhimurium strains, in the absence or in the presence of metabolic activation. Cytotoxicity was detected at concentrations higher than 0.04 μL/plate in the absence of S9 mix and higher than 0.08 μL/plate in the presence of S9 mix and no gene mutation increase was observed. For the in vitro mammalian cell micronucleus test, V79 Chinese hamster lung fibroblasts were used. Cytotoxicity was only observed at concentrations higher than or equal to 0.05 μg/mL. Moreover, when tested in noncytotoxic concentrations, O. majorana essential oil was not able to induce chromosome mutation. The results from this study therefore suggest that O. majorana essential oil is not mutagenic at the concentrations tested in the Salmonella/microsome and micronucleus assays.
1. Introduction
The Origanum genus belongs to the Lamiaceae family and includes species with interesting pharmacological effects [1]. Origanum majorana L. or Majorana hortensis Moench is an aromatic plant rich in essential oils and commercially grown in southern Europe and in the Mediterranean region [2, 3]. It is popularly known as marjoram and has been used in the form of herbal infusions in folk medicine for asthma, cold, coughs, cramps, depression, dizziness, gastrointestinal disorders, hay fever, headache, toothache, and sinus congestion and as a diuretic and to promote menstruation [2, 3].
O. majorana crude extract, dichloromethane, ethyl acetate, aqueous fractions, and O. majorana essential oil have shown significant results in inhibiting the growth of bacteria and fungi and the synthesis of microbial metabolites [1, 4, 5]. Because of its antioxidant effects [6–8], O. majorana essential oil or extract can be used in the prevention of central nervous system disorders [2, 9]. O. majorana essential oil was also able to partially prevent the ethanol-induced decline in sperm quality, testosterone levels, and the weight of reproductive organs in male rats [2]. Previous studies have reported the potential use of O. majorana ethanolic extract as anticancer agent [10, 11], whereas the tea extract has been shown to have immunostimulant, antigenotoxic and antimutagenic properties [3, 12]. These activities are attributed to the chemical composition, which is characterized as rich in flavonoids and terpenoids [13].
In spite of the growing interest in using essential oils and other extracts in the treatment of diseases, it is necessary to perform toxicological studies to ensure that the chemical compounds found in the plant under study exert no adverse effects that would impair its use for therapeutic purposes. Genetic toxicology data are employed as a surrogate for long-term carcinogenicity data during early drug development and must be conducted when there is an indication of continuous use or prolonged therapeutic treatment. The genotoxicity battery includes gene and chromosome mutation detection tests for products for human use [14, 15].
Therefore, this study was performed to investigate the mutagenic activity of O. majorana essential oil using five strains of bacteria in the Salmonella/microsome test, with or without metabolic activation, and in the in vitro mammalian cell micronucleus test using Chinese hamster lung fibroblasts (V79 cells). Essential oil chemical characterization was also performed to identify the major components of the oil.
2. Material and Methods
2.1. Chemicals
Diethyl ether was acquired from Tedia Company, Inc. (Fairfield, OH, USA). Aflatoxin B1 (CAS 1162-65-8), 2-aminofluorene (CAS 153-78-6), 4-nitroquinoline-oxide (CAS 56-57-5), sodium azide (CAS 26628-22-8), colchicine (CAS 64-86-8), etoposide (CAS 33419-42-0), cytochalasin B (14930-96-2), dimethyl sulfoxide (DMSO) (CAS 67-68-5), glucose-6-phosphate (CAS 56-73-5), and NADP (CAS 698-999-85-8) were obtained from Sigma Chemical (St. Louis, USA). Dulbecco's Modified Eagle's Medium (DMEM), fetal bovine serum (FBS), trypsin-EDTA, L-glutamine, and antibiotics were purchased from Gibco BRL (Grand Island, NY, USA). The Aroclor 1254 induced male Sprague Dawley rat liver S9 fraction was obtained from Molecular Toxicology Inc. (Boone, NC, USA). All other chemicals were of analytical grade and were obtained from standard commercial suppliers.
2.2. Sample and Analysis by GC-MS
O. majorana dried leaves, acquired from Luar Sul Company (Santa Cruz do Sul, RS, Brazil) with an Egyptian Certificate of Origin and free of macroscopic contaminants, were used. The essential oil was obtained by steam distillation using a modified Clevenger device. The GC-MS was carried out on a Shimadzu mass spectrometer (GC/MS-QP5000) connected with cylindrical quadruple and operated at 70 eV ionization energy. An apolar Durabond-DB5 column (30 m × 0.25 mm × 0.25 μm) was used. To confirm the identity of the compounds, a polar column was also used (LM-120). The temperature was programmed from 60 to 300°C at 3°C/min and the injector and detector temperatures were set at 220°C and 250°C, respectively. GC-FID was used for quantification [16, 17]. The relative composition of the oils was obtained by electronic integration and the identification of the compounds was based on comparison to retention indices, determined relatively to the retention times of an homologous series of n-alkanes, and mass spectra of commercial database (NIST) and literature [18].
2.3. Bacterial and Mammalian Strains
Salmonella typhimurium strains TA98, TA97a, TA1535, TA100, and TA102 were obtained from Molecular Toxicology Inc. (Boone, NC, USA). TA98 detects frameshifts in the DNA target –C–G–C–G–C–G–C–G; TA97a detects frameshift mutations in –C–C–C–C–C–C–, +1 cytosine; TA1535 detects base pair substitutions and its corresponding isogenic strain TA100 detects base pair substitutions of a leucine [GAG] by proline [GGG]; and TA102 strain is sensitive to oxidative and alkylating mutagens and detects transversions or transitions in TAA DNA sequences [19].
Chinese hamster lung fibroblast V79 cells were obtained from Banco de Células do Rio de Janeiro (Rio de Janeiro, Brazil). V79 cells were chosen in this study because this cell line is largely adopted in cytogenetic assays mainly because of its stable karyotype and relatively short cellular cycle, varying between 12 and 16 h [20].
2.4. Salmonella/Microsome Mutagenicity Assay
Mutagenicity was assayed by the preincubation procedure where cells were incubated with and without metabolic activation in five strains of S. typhimurium. The S9 metabolic activation mixture (S9 mix) was prepared according to Maron and Ames [19]. Briefly, 100 μL of test bacterial cultures (1-2 × 109 cells/mL) was incubated at 37°C with different concentrations of O. majorana essential oil in the absence or in the presence of S9 mix (4% S9 fraction, 5 mM D-glucose-6-phosphate, 4 mM NADP, 33 mM MgCl2/8 mM KCl, and 100 mM phosphate buffer pH 7.4, as concentrations per tube) for 20–30 min, without shaking. Subsequently, 2 mL of soft agar (0.6% agar, 0.5% NaCl, 50 μM histidine, and 50 μM biotin, pH 7.4, 42°C) was added to the test tube and poured immediately onto a plate of minimal agar (1.5% agar, Vogel-Bonner E medium, containing 2% glucose).
Aflatoxin B1 (1 μg/plate) was used as positive control in the presence of metabolic activation (with S9 mix) for TA97a, TA98, and TA100 strains and 2-aminofluorene (2-AF, 10 μg/plate) was used for TA102 and TA1535 strains. In the absence of metabolic activation, 4-nitroquinoline-oxide (4-NQO, 0.5 μg/plate) was used for TA97a, TA98, and TA102 strains and sodium azide (1 μg/plate) was used for TA100 and TA1535 strains. Plates were incubated in the dark at 37°C for 48 h before counting the revertant colonies.
2.5. In Vitro Mammalian Cell Micronucleus Test (MNvit)
V79 cells were cultivated in standard conditions with high glucose DMEM medium supplemented with heat-inactivated 10% FBS and antibiotics. Cells were maintained in 25 cm2 culture bottles at 37°C and humid atmosphere with 5% CO2. For harvesting and culture establishment, PBS (phosphate buffer saline, pH 7.4) and 0.25% trypsin-EDTA were used. For the experiments, O. majorana essential oil was diluted in DMSO and further diluted in DMEM culture medium obtaining different concentrations in which 5 μL/mL was used as the highest test concentration. This concentration was chosen considering the extract solubility in the culture medium and the OECD suggestion for mutagenesis analysis [21]. The final concentration of DMSO in the cultures was 0.5%.
Thus, to verify the potential mutagenic effects of O. majorana essential oil, the MNvit assay was performed as described by Gonçalves et al. [22] and OECD 487 [21] with minor modifications. For this, 1 × 105 V79 cells were seeded per well in 6-well plates and exposed to 0.003125, 0.00625, 0.0125, and 0.025 μL/mL of O. majorana essential oil for 3 or 21 h. 0.5% DMSO was used as negative control and 0.75 μg/mL colchicine and 0.250 μg/mL etoposide were used as positive controls. In the 3 h treatment experiments, the medium was removed after treatment and replaced with fresh medium containing 3 μg/mL cytochalasin B to block the cytokinesis and then incubated for 21 h (period corresponding to 1.5–2 normal cell cycles). In the 21 h of treatment experiments, cultures were exposed to O. majorana essential oil in the presence of 3 μg/mL cytochalasin B. In both experiments, after the cytochalasin incubation, the cells were incubated in hypotonic solution (KCl 0.075 M) for 3 min at 4°C, prefixed and fixed with methanol and acetic acid (3 : 1) solution. This process was repeated and the cells were stored at 4°C overnight. Fixed cells were dropped on to microscope slides and stained with 2% Giemsa. The slide analysis was performed by means of a semiautomated scoring PathFinder Screen Tox system (IMSTAR, France) [23] or manually scored in optical microscope. It is important to note that the semiautomated scoring was previously validated in the laboratory demonstrating high correlation with the manual scoring results. Thus, for the analysis, the cell viability was first evaluated by the replication index [21]. For this, the number of mononucleated, binucleated, and multinucleated cells was counted in 500 cells/slide (manual analysis) or in all cells localized in a preselected area by slide (semiautomated analysis). For the analysis of micronucleus formation, at least 500 binucleated cells were considered for the presence of micronucleus, totalizing around 3000 cells per test group (two slides per treatment well, three wells per treatment group) when manual scoring was used. In the semiautomated scoring, the system identified in a preselected area the number of binucleated cells and the number of binucleated cells with micronucleus. Then, the technician confirmed the real presence of micronuclei in each binucleated cell previously identified by the software.
2.6. Statistical Analysis
The results of the Salmonella/microsome mutagenicity assay were analyzed by using the Salmonella Assay Software (Environmental Monitoring System Laboratory, EPA, software version 2.3). This program allows assessment of the dose-response effect by calculating the analysis of variance (ANOVA) between the measurements of the average number of revertants tested at different concentrations and the average number of revertants per plate from the negative control (mutagenic index, MI). A test substance was considered mutagenic when significant dose response and ANOVA variance were observed, and the increase in the mean number of revertants on test plates was at least twofold higher than that observed in the negative control plates (or MI ≥ 3 for TA1535 strain).
In the MNvit test, all experiments were independently repeated at least three times, with duplicate samples for each treatment. The results are expressed as mean ± standard deviation (SD) and were analyzed by one-way ANOVA followed by Dunnett's Multiple Comparison Test or Student's t-test when needed by using GraphPad Prism 5.0 software (GraphPad Inc., San Diego, CA). p < 0.05 was considered statistically significant.
3. Results and Discussion
3.1. Essential Oil Characterization
The analysis of the chemical composition of samples obtained from different geographical locations indicates that the biological activity is directly related to the concentration of the essential oil components, which may vary according to the region [4, 7, 24, 25]. Moreover, season, climate, stage of plant development at harvest, and the technique of extraction of the product may influence the quantity of the plant compounds [24, 26].
Fifteen compounds were identified in the O. majorana essential oil (Table 1). The most abundant compounds were γ-terpinene (25.73%), α-terpinene (17.35%), terpinen-4-ol (17.24%), and sabinene (10.8%). This chemical profile is in accordance with what is reported in the literature, with some quantitative variations. Rodrigues et al. [27] and Vági et al. [13] also reported the presence of terpenes as the major components of the O. majorana essential oil. Usually, terpinen-4-ol and γ-terpinene are described as the most abundant compounds in O. majorana essential oil and sabinene and α-terpinene are also observed [4, 7, 13, 24, 25].
Table 1.
Chemical composition of the Origanum majorana essential oil.
| Compound | RIa | % |
|---|---|---|
| α-Thujene | 921 | 3.96 |
| α-Pinene | 927 | 1.24 |
| Sabinene | 966 | 10.80 |
| Myrcene | 986 | 2.08 |
| α-Phellandrene | 1000 | 1.70 |
| α-Terpinene | 1012 | 17.35 |
| o-Cymene | 1019 | 2.24 |
| β-Phellandrene | 1023 | 7.05 |
| γ-Terpinene | 1053 | 25.73 |
| Terpinolene | 1082 | 3.76 |
| N.I.b | 1097 | 1.06 |
| Terpinen-4-ol | 1174 | 17.24 |
| Trans-sabinene hydrate acetate | 1248 | 0.13 |
| Linalool acetate | 1251 | 1.38 |
| Terpinenen-4-ol acetate | 1293 | 0.88 |
| (Z)-caryophyllene | 1404 | 2.72 |
| N.I.b | 1480 | 0.67 |
| Total identified | 98.26 |
aRelative retention index experimentally determined against n-alkanes on Durabond-DB5 column. bCompound not identified.
3.2. Evaluation of Mutagenesis in Bacteria
The dose range of the O. majorana essential oil was determined in range finding experiments using the TA100 strain, with or without S9 mix, using five serially diluted concentrations (0.008–5 μL/plate), where the highest dose tested was determined by the solubility of the substance and as suggested by the Ames Test Guideline (Table 2) [28]. Cytotoxicity (mutagenic index ≤0.6) can be observed in Table 3 at the highest concentrations used for TA98, TA100, TA102, and TA1535 in absence of metabolic activation and for TA100 in presence of metabolic activation. In regard to the range finding study, it was performed only in TA100 strain in accordance with OECD suggestion.
Table 2.
Range finding study of the O. majorana essential oil using the TA100 strain.
| Substance | Concentration | Without metabolic activation (−S9) | With metabolic activation (+S9) | ||
|---|---|---|---|---|---|
| Rev/platea | MIb | Rev/platea | MIb | ||
| NCa | — | 141.33 ± 17.56 | — | 148.00 ± 4.58 | — |
| Essential oil | 0.008 µL/mL | 105.67 ± 22.01 | 0.75 | 145.33 ± 18.15 | 0.98 |
| 0.04 µL/mL | 53.00 ± 9.64 | 0.38b | 127.00 ± 19.47 | 0.86 | |
| 0.2 µL/mL | 33.00 ± 23.07 | 0.23b | 72.67 ± 13.05 | 0.49b | |
| 1 µL/mL | 0.33 ± 0.58 | 0.00 | 0.33 ± 0.58 | 0.00 | |
| 5 µL/mL | 0.00 ± 0.00 | 0.00 | 0.00 ± 0.00 | 0.00 | |
aNegative control: DMSO (10 µL). bCytotoxicity.
Table 3.
Induction of his+ revertants in S. typhimurium strains by Origanum majorana essential oil with and without metabolic activation (S9 mix).
| Substance | Concentration | S. typhimurium strains | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| TA98 | TA97a | TA100 | TA1535 | TA102 | |||||||
| Rev/platea | MIb | Rev/platea | MIb | Rev/platea | MIb | Rev/platea | MIb | Rev/platea | MIb | ||
| Without metabolic activation (−S9) | |||||||||||
| NCc | — | 19.7 ± 8.2 | — | 69.7 ± 3.1 | — | 103.7 ± 6.1 | — | 4.3 ± 2.5 | — | 291.7 ± 10.5 | — |
| Essential oil (µL/plate) |
0.0025 | 24.7 ± 6.4 | 1.03 | 91.7 ± 7.5 | 1.32 | 102.3 ± 9.3 | 0.99 | 4.7 ± 2.1 | 1.09 | 268 ± 12.5 | 0.92 |
| 0.005 | 14.5 ± 3.4 | 0.74 | 84.7 ± 18.9 | 1.22 | 110.7 ± 10.5 | 1.07 | 6.3 ± 4 | 1.45 | 255.3 ± 26.5 | 0.88 | |
| 0.01 | 15.2 ± 6.2 | 0.77 | 89.3 ± 23 | 1.28 | 95.7 ± 15.6 | 0.92 | 2 ± 0 | 0.46 | 253 ± 46.5 | 0.87 | |
| 0.02 | 10.5 ± 2.3 | 0.53 | 71.7 ± 15.5 | 1.03 | 75.5 ± 30.4 | 0.73 | 4 ± 1.7 | 0.92 | 174.7 ± 15.6 | 0.60 | |
| 0.04 | 9.2 ± 2.5 | 0.47 | 72 ± 13.1 | 1.03 | 41.7 ± 4 | 0.40 | 1.3 ± 1.2 | 0.31 | 111.3 ± 9.1 | 0.38 | |
| PCd (µg/plate) | 0.5 (4NQO) | 332 ± 22.6∗∗∗ | 20.68 | 464.3 ± 91.8∗∗∗ | 6.66 | — | — | — | — | 1031 ± 93.5∗∗∗ | 3.54 |
| 1 (NaN3) | — | — | — | — | 427.3 ± 31.2∗∗∗ | 4.12 | 312.3 ± 9∗∗∗ | 72.13 | — | — | |
|
| |||||||||||
| With metabolic activation (+S9) | |||||||||||
| NCc | — | 15.3 ± 1.5 | — | 100.7 ± 9.9 | — | 179.7 ± 11.1 | — | 9.3 ± 2.9 | — | 325.7 ± 15.5 | — |
| Essential oil (µL/plate) |
0.005 | 19 ± 3.6 | 1.24 | 109.3 ± 8.5 | 1.09 | 155.7 ± 10 | 0.87 | 10.3 ± 4.5 | 1.1 | 310 ± 3.6 | 0.95 |
| 0.01 | 23.3 ± 1.5 | 1.52 | 112 ± 20.1 | 1.11 | 141.3 ± 12.6 | 0.79 | 8 ± 3 | 0.86 | 338 ± 12.7 | 1.04 | |
| 0.02 | 15.7 ± 1.5 | 1.01 | 115 ± 10.5 | 1.14 | 135.3 ± 6.7 | 0.75 | 7.7 ± 1.5 | 0.83 | 332.7 ± 29 | 1.02 | |
| 0.04 | 21 ± 9.5 | 1.37 | 113.7 ± 8.7 | 1.13 | 112 ± 14.9 | 0.62 | 8.3 ± 3.5 | 0.89 | 301.7 ± 26 | 0.93 | |
| 0.08 | 18.7 ± 1.2 | 1.22 | 131.7 ± 10.6 | 1.31 | 99.3 ± 14 | 0.55 | 7.7 ± 2.3 | 0.83 | 295.7 ± 31.1 | 0.91 | |
| PCe (µg/plate) | 1 (AFB1) | 1587 ± 91.1∗∗∗ | 103.5 | 587 ± 88.5∗∗ | 5.83 | 660.7 ± 59.9∗∗ | 3.68 | — | — | — | — |
| 10 (2-AF) | — | — | — | — | — | — | 104.3 ± 12.1∗∗ | 11.18 | 767.3 ± 53.6∗∗∗ | 2.36 | |
aNumber of revertants/plate: mean of three independent experiments ± SD; bMI: mutagenic index (number of his+ induced in the sample/number of spontaneous his+ in the negative control). cNegative control: DMSO (10 µL) used as solvent for the extract. dPositive control (−S9): NaN3 (sodium azide) to TA100 and TA1535 and 4-NQO to TA97a, TA98, and TA102. ePositive control (+S9): AFB1 (aflatoxin B1) for TA97a, TA98, and TA100 and 2-aminofluorene for TA102 and TA1535; ∗∗ p < 0.01; ∗∗∗ p < 0.001, in relation to the negative control.
Thus, in the mutagenicity test the dose ranged between 0.0025 and 0.04 μL/plate in the absence of S9 mix and between 0.005 and 0.08 μL/plate in the presence of S9 mix. It can be seen in Table 3 that no mutagenic effect was detected on strains TA98 and TA97a in the presence or absence of metabolic activation. In addition, no mutagenicity was seen in the strain TA1535 and its corresponding isogenic strain TA100. Negative results were also observed in the TA102 strain, which is sensitive to oxidative and alkylating mutagens [19].
3.3. Evaluation of Mutagenesis in Mammalian Cells
The MNvit assay is a genotoxicity test for the detection of micronuclei in the cytoplasm of interphase cells [29] and was carried out to evaluate the potential of the O. majorana essential oil to induce chromosome mutation. The O. majorana essential oil concentrations were chosen based on replication index previously analyzed at 0.0005, 0.005, 0.05, 0.5, and 5 μL/mL (Table 4) with 3 h of exposure, where a concentration-dependent decrease in cellular viability was observed mainly in concentrations ≥0.05 μL/mL. Moreover, 100% cytotoxicity was observed at the concentrations of 0.5 and 5 μL/mL. The analysis of micronucleus formation was performed in concentrations lower than 0.05 μL/mL to provide the appropriate range of cytotoxicity (up to 55 ± 5%).
Table 4.
Replication index of cultures exposed for 3 h to Origanum majorana essential oil.
| Substance | Concentration | Replication factor (mean ± S.D.) | Replication index (%) |
|---|---|---|---|
| NCa | — | 0.809 ± 0.012 | 100 |
| Essential oil | 0.0005 μL/mL | 0.823 ± 0.027 | 102 |
| 0.005 μL/mL | 0.800 ± 0.035 | 99 | |
| 0.05 μL/mL | 0.126 ± 0.028 | 15 | |
| 0.5 μL/mL | 0 ± 0 | 0 | |
| 5 μL/mL | 0 ± 0 | 0 |
aNegative control: DMSO (0.5%).
Micronuclei may originate from acentric chromosome fragments (i.e., lacking a centromere) or whole chromosomes that are unable to migrate to the poles during the anaphase stage of cell division. The assay detects the activity of clastogenic and aneugenic chemicals in cells that have undergone cell division during or after exposure to the test substance ([29]; reviewed in [21]). As illustrated in Table 5, the O. majorana essential oil did not decrease the cell viability after 3 h or 21 h of exposure. In relation to micronucleus formation in binucleated cells, O. majorana essential oil was not able to induce increase in the micronucleus frequency when compared to the negative control, in both periods evaluated (Figure 1). It is important to note that these results were observed in the absence of S9-mix and additional studies in the presence of metabolic activation are needed.
Table 5.
Replication index of cultures exposed to Origanum majorana essential oil for 3 h or 21 h.
| Substance | Concentration | 3 h | 21 h | ||
|---|---|---|---|---|---|
| Replication factor (mean ± S.D.) | Replication index (%) | Replication factor (mean ± S.D.) | Replication index (%) | ||
| NCa | — | 0.726 ± 0.021 | 100 | 0.820 ± 0.050 | 100 |
| Essential oil | 0.003125 μL/mL | 0.735 ± 0.019 | 101 | 0.822 ± 0.036 | 100 |
| 0.00625 μL/mL | 0.698 ± 0.021 | 96 | 0.809 ± 0.014 | 99 | |
| 0.0125 μL/mL | 0.731 ± 0.012 | 101 | 0.837 ± 0.027 | 102 | |
| 0.025 μL/mL | 0.722 ± 0.031 | 99 | 0.769 ± 0.024 | 94 | |
| Colchicineb | 0.75 μg/mL | 0.501 ± 0.013 | 69 | — | — |
| Etoposideb | 0.25 μg/mL | — | — | 0.767 ± 0.028 | 94 |
aNegative control: DMSO (0.5%). bPositive control.
Figure 1.
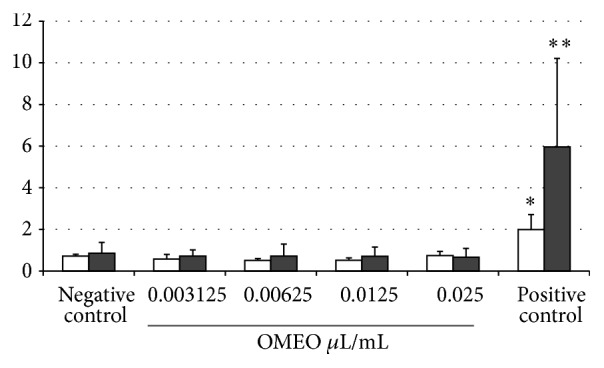
Micronucleus frequency of V79 cells exposed to different O. majorana essential oil (OMEO) concentrations for 3 h (white columns) and 21 h (black columns). Negative control: DMSO (0.5%); positive control: 0.75 μg/mL colchicine (3 h) and 0.25 μg/mL etoposide (21 h). ∗ p < 0.05; ∗∗ p < 0.01, in relation to the negative control (Student's t-test).
Several studies have reported that sweet marjoram leaves, especially in the form of an herbal tea, may be useful as immunostimulant and in reducing genotoxicity in patients under chemotherapeutic interventions, besides inhibiting the production of free radicals [3, 6]. Indeed, Khan et al. [12] proposed that reducing the availability of the genotoxic metabolites of chemical carcinogens from the body, by forming more water soluble conjugates, will consequently decrease the chance of an interaction with DNA through adduct formation that may lead to cancer.
Terpinen-4-ol, one of the main compounds found in O. majorana essential oil, is also the main active compound of the Melaleuca alternifolia essential oil. Both the M. alternifolia essential oil and the terpinen-4-ol compound were able to induce caspase-dependent apoptosis of melanoma cells [30]. Gomes-Carneiro et al. [31] reported that α-terpinene was not able to induce mutagenicity in Salmonella strains when tested alone.
In conclusion, the findings from the present study indicated that, at the concentrations and conditions used, O. majorana essential oil is not able to induce in vitro gene (S. typhimurium strains) and chromosome (V79 Chinese hamster lung fibroblast cells) mutations, respectively, contributing to the knowledge in the safe use of this essential oil. Further studies are now in progress to explore possible reproductive toxicity, teratology, and development toxicology induced by O. majorana essential oil in vivo.
Competing Interests
The authors declare that there are no competing interests regarding the publication of this paper.
References
- 1.Abdel-Massih R. M., Abraham A. Extracts of Rosmarinus officinalis, Rheum rhaponticum, and Origanum majorana exhibit significant anti-staphylococcal activity. International Journal of Pharmaceutical Sciences and Research. 2014;5:819–828. [Google Scholar]
- 2.El-Ashmawy I. M., Saleh A., Salama O. M. Effects of marjoram volatile oil and grape seed extract on ethanol toxicity in male rats. Basic and Clinical Pharmacology and Toxicology. 2007;101(5):320–327. doi: 10.1111/j.1742-7835.2007.00125.x. [DOI] [PubMed] [Google Scholar]
- 3.Ramadan G., El-Beih N. M., Zahra M. M. Egyptian sweet marjoram leaves protect against genotoxicity, immunosuppression and other complications induced by cyclophosphamide in albino rats. British Journal of Nutrition. 2012;108(6):1059–1068. doi: 10.1017/s0007114511006210. [DOI] [PubMed] [Google Scholar]
- 4.Busatta C., Vidal R. S., Popiolski A. S., et al. Application of Origanum majorana L. essential oil as an antimicrobial agent in sausage. Food Microbiology. 2008;25(1):207–211. doi: 10.1016/j.fm.2007.07.003. [DOI] [PubMed] [Google Scholar]
- 5.Prakash B., Singh P., Kedia A., Dubey N. K. Assessment of some essential oils as food preservatives based on antifungal, antiaflatoxin, antioxidant activities and in vivo efficacy in food system. Food Research International. 2012;49(1):201–208. doi: 10.1016/j.foodres.2012.08.020. [DOI] [Google Scholar]
- 6.Jin Jun W., Kyung Han B., Won Yu K., et al. Antioxidant effects of Origanum majorana L. on superoxide anion radicals. Food Chemistry. 2001;75(4):439–444. doi: 10.1016/s0308-8146(01)00233-3. [DOI] [Google Scholar]
- 7.Alizadeh A., Khosh-Khui M., Javidnia K., Firuzi O., Jokar S. M. Chemical composition of the essential oil, total phenolic content and antioxidant activity in Origanum majorana L. (Lamiaceae) cultivated in Iran. Advances in Environmental Biology. 2011;5(8):2326–2331. [Google Scholar]
- 8.Roby M. H. H., Sarhan M. A., Selim K. A.-H., Khalel K. I. Evaluation of antioxidant activity, total phenols and phenolic compounds in thyme (Thymus vulgaris L.), sage (Salvia officinalis L.), and marjoram (Origanum majorana L.) extracts. Industrial Crops and Products. 2013;43(1):827–831. doi: 10.1016/j.indcrop.2012.08.029. [DOI] [Google Scholar]
- 9.Rezaie A., Jafari B., Mousavi G., et al. Comparative study of sedative, pre-anesthetic and anti-anxiety effect of Origanum majorana extract with diazepam on rats. Research Journal of Biological Sciences. 2011;6:611–614. doi: 10.3923/rjbsci.2011.611.614. [DOI] [Google Scholar]
- 10.Al Dhaheri Y., Attoub S., Arafat K., et al. Anti-metastatic and anti-tumor growth effects of Origanum majorana on highly metastatic human breast cancer cells: inhibition of NFκB signaling and reduction of nitric oxide production. PLoS ONE. 2013;8(7) doi: 10.1371/journal.pone.0068808.e68808 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Rao S., Timsina B., Nadumane V. K. Evaluation of the anticancer potentials of Origanum marjorana on fibrosarcoma (HT-1080) cell line. Asian Pacific Journal of Tropical Disease. 2014;4(1):S389–S394. doi: 10.1016/s2222-1808(14)60476-x. [DOI] [Google Scholar]
- 12.Khan J. A., Jalal J. A., Ioanndes C., Moselhy S. S. Impact of aqueous doash extract on urinary mutagenicity in rats exposed to heterocyclic amines. Toxicology and Industrial Health. 2013;29(2):142–148. doi: 10.1177/0748233711427053. [DOI] [PubMed] [Google Scholar]
- 13.Vági E., Simándi B., Suhajda Á., Héthelyi É. Essential oil composition and antimicrobial activity of Origanum majorana L. extracts obtained with ethyl alcohol and supercritical carbon dioxide. Food Research International. 2005;38(1):51–57. doi: 10.1016/j.foodres.2004.07.006. [DOI] [Google Scholar]
- 14.Kirkland D., Reeve L., Gatehouse D., Vanparys P. A core in vitro genotoxicity battery comprising the Ames test plus the in vitro micronucleus test is sufficient to detect rodent carcinogens and in vivo genotoxins. Mutation Research. 2011;721(1):27–73. doi: 10.1016/j.mrgentox.2010.12.015. [DOI] [PubMed] [Google Scholar]
- 15.ICH S2 (R1) Guidance on Genotoxicity Testing and Data Interpretation for Pharmaceuticals Intended for Human Use. 2011. [PubMed] [Google Scholar]
- 16.Limberger R. P., Sobral M., Henriques A. T., Menut C., Bessière J.-M. Óleos voláteis de espécies de Myrcia nativas do Rio Grande do Sul. Química Nova. 2004;27:916–919. [Google Scholar]
- 17.Simões-Pires C. A., Debenedetti S., Spegazzini E., et al. Investigation of the essential oil from eight species of Baccharis belonging to sect. Caulopterae (Asteraceae, Astereae): a taxonomic approach. Plant Systematics and Evolution. 2005;253(1–4):23–32. doi: 10.1007/s00606-005-0296-6. [DOI] [Google Scholar]
- 18.Adams R. P. Identification of Essential Oil Components by Gas Chromatograph/Quadrupole Mass Spectrometry. Carol Stream, Ill, USA: Allured Publishing; 2007. [Google Scholar]
- 19.Maron D. M., Ames B. N. Revised methods for the Salmonella mutagenicity test. Mutation Research/Environmental Mutagenesis and Related Subjects. 1983;113(3-4):173–215. doi: 10.1016/0165-1161(83)90010-9. [DOI] [PubMed] [Google Scholar]
- 20.Bradley M. O., Bhuyan B., Francis M. C., Langenbach R., Peterson A., Huberman E. Mutagenesis by chemical agents in V79 Chinese hamster cells: a review and analysis of the literature. A report of the gene-tox program. Mutation Research/Reviews in Genetic Toxicology. 1981;87(2):81–142. doi: 10.1016/0165-1110(81)90029-4. [DOI] [PubMed] [Google Scholar]
- 21.OECD. Test no. 487: in vitro mammalian cell micronucleus test, OECD guidelines for the testing of chemicals, section 4. Health Effects. 2014:26. doi: 10.1787/9789264224438-en. [DOI] [Google Scholar]
- 22.Gonçalves T. S., Menezes L. M. D., Trindade C., et al. Cytotoxicity and genotoxicity of orthodontic bands with or without silver soldered joints. Mutation Research—Genetic Toxicology and Environmental Mutagenesis. 2014;762:1–8. doi: 10.1016/j.mrgentox.2014.01.011. [DOI] [PubMed] [Google Scholar]
- 23.Decordier I., Papine A., Vande Loock K., Plas G., Soussaline F., Kirsch-Volders M. Automated image analysis of micronuclei by IMSTAR for biomonitoring. Mutagenesis. 2011;26(1):163–168. doi: 10.1093/mutage/geq063. [DOI] [PubMed] [Google Scholar]
- 24.Jelali N., Dhifi W., Chahed T., Marzouk B. Salinity effects on growth, essential oil yield and composition and phenolic compounds content of marjoram (origanum majorana l.) leaves. Journal of Food Biochemistry. 2011;35(5):1443–1450. doi: 10.1111/j.1745-4514.2010.00465.x. [DOI] [Google Scholar]
- 25.Ramos S., Rojas L. B., Lucena M. E., Meccia G., Usubillaga A. Chemical composition and antibacterial activity of Origanum majorana L. essential oil from the Venezuelan Andes. Journal of Essential Oil Research. 2011;23(5):45–49. doi: 10.1080/10412905.2011.9700481. [DOI] [Google Scholar]
- 26.Ncube B., Finnie J. F., Van Staden J. Quality from the field: the impact of environmental factors as quality determinants in medicinal plants. South African Journal of Botany. 2012;82:11–20. doi: 10.1016/j.sajb.2012.05.009. [DOI] [Google Scholar]
- 27.Rodrigues M. R. A., Caramão E. B., Arce L., Ríos A., Valcárcel M. Determination of monoterpene hydrocarbons and alcohols in Majorana hortensis Moench by micellar electrokinetic capillary chromatographic. Journal of Agricultural and Food Chemistry. 2002;50(15):4215–4220. doi: 10.1021/jf011667n. [DOI] [PubMed] [Google Scholar]
- 28.OECD. OECD. 471. OECD Publishing; 1997. Test No. 471: bacterial reverse mutation test, OECD guidelines for the testing of chemicals, section 4: health effects. [DOI] [Google Scholar]
- 29.Fenech M., Kirsch-Volders M., Natarajan A. T., et al. Molecular mechanisms of micronucleus, nucleoplasmic bridge and nuclear bud formation in mammalian and human cells. Mutagenesis. 2011;26(1):125–132. doi: 10.1093/mutage/geq052. [DOI] [PubMed] [Google Scholar]
- 30.Calcabrini A., Stringaro A., Toccacieli L., et al. Terpinen-4-ol, the main component of Melaleuca alternifolia (Tea Tree) oil inhibits the in vitro growth of human melanoma cells. The Journal of Investigative Dermatology. 2004;122(2):349–360. doi: 10.1046/j.0022-202x.2004.22236.x. [DOI] [PubMed] [Google Scholar]
- 31.Gomes-Carneiro M. R., Viana M. E. S., Felzenszwalb I., Paumgartten F. J. R. Evaluation of β-myrcene, α-terpinene and (+)- and (−)-α-pinene in the Salmonella/microsome assay. Food and Chemical Toxicology. 2005;43(2):247–252. doi: 10.1016/j.fct.2004.09.011. [DOI] [PubMed] [Google Scholar]