

Keywords: nerve regeneration, spinal cord injury, methylprednisolone, rosiglitazone, inflammation, drug therapy, anti-inflammatory agents, functional recovery, neural regeneration
Abstract
Methylprednisolone exhibits anti-inflammatory antioxidant properties, and rosiglitazone acts as an anti-inflammatory and antioxidant by activating peroxisome proliferator-activated receptor-γ in the spinal cord. Methylprednisolone and rosiglitazone have been clinically used during the early stages of secondary spinal cord injury. Because of the complexity and diversity of the inflammatory process after spinal cord injury, a single drug cannot completely inhibit inflammation. Therefore, we assumed that a combination of methylprednisolone and rosiglitazone might promote recovery of neurological function after secondary spinal cord injury. In this study, rats were intraperitoneally injected with methylprednisolone (30 mg/kg) and rosiglitazone (2 mg/kg) at 1 hour after injury, and methylprednisolone (15 mg/kg) at 24 and 48 hours after injury. Rosiglitazone was then administered once every 12 hours for 7 consecutive days. Our results demonstrated that a combined treatment with methylprednisolone and rosiglitazone had a more pronounced effect on attenuation of inflammation and cell apoptosis, as well as increased functional recovery, compared with either single treatment alone, indicating that a combination better promoted recovery of neurological function after injury.
Introduction
Spinal cord injury (SCI) results in neurological deficits and motor dysfunction (Rintala et al., 1998). The pathophysiology of acute SCI is complex and two mechanisms can be distinguished as primary and secondary damage (Collins, 1983); primary traumatic mechanical injury to the spinal cord induces death in a number of neurons that cannot be recovered or regenerated (Profyris et al., 2004). These events are then exacerbated by secondary damage, including a variety of biochemical and pathological processes that begin within minutes after initial injury and continue for days and weeks (Diaz-Ruiz et al., 2002; Hausmann, 2003).
Several pharmacological agents have been developed to attenuate secondary damage and to maximize the extent of spared neurologic tissue following SCI. Methylprednisolone (MP) is a pharmacological agent used early in the treatment of experimental and human SCI because of the anti-inflammatory actions and potent anti-oxidation effects mediated by suppression of nuclear factor kappa B binding (Hsu and Dimitrijevic, 1990; Hall, 1992; Short et al., 2000; Nadeau and Rivest, 2003; Sayer et al., 2006). However, MP application is only effective within a relatively narrow time window after SCI. The onset of glucocorticoid receptor expression within the spinal cord is rapid, but is only transiently induced following traumatic injury (Yan et al., 1999), perhaps explaining the very limited therapeutic window of MP. Some studies have proposed the use of MP in combined treatments as an initial neuroprotective therapy coupled with pro-regenerative or other treatments (Baffour et al., 1995; Nash et al., 2002; Takami et al., 2002). Rosiglitazone (RSG), one type of thiazolidinedione, is a synthetic agonist of the ligand-activated transcription factor peroxisome proliferator-activated receptor-γ. Recent studies have shown that RSG exhibits anti-inflammatory and anti-oxidant properties in animal models of acute central nervous system insults involving focal cerebral ischemia, SCI, and traumatic brain injury (Sundararajan et al., 2005; Pereira et al., 2006; Park et al., 2007; Hyong et al., 2008).
However, based on the complexity and redundancy of the inflammatory response following SCI, it is unlikely that a single target can achieve complete inhibition of inflammation. The therapeutic effects of simultaneous administration of MP and RSG have not previously been evaluated. Therefore, the purpose of the present study was to investigate the neuroprotective effect of simultaneous administration of MP and RSG on experimental SCI, and to determine whether these mechanistically distinct pharmacological reagents can act synergistically.
Materials and Methods
Experimental animals
Ninety-six adult male Sprague-Dawley rats aged 10–12 weeks and weighing 200–250 g were provided by the Laboratory Animal Center of Zhejiang Province of China (SCXK (Zhe) 2014-0001). All animal experiments were approved by the Ethics Committee for Animal Experiments of Zhejiang University of China. Every effort was made to minimize the number and suffering of the animals used in the following experiments, in accordance with the United States National Institutes of Health Guide for the Care and Use of Laboratory Animal (NIH Publication No. 85-23, revised 1986).
Methods
Model establishment and administration
Rats were intraperitoneally anesthetized using chloral hydrate (400 mg/kg). A longitudinal incision was made on the midline of the back, exposing the paravertebral muscles. The spinal cord was exposed via a four-level T5–8 laminectomy, and compressive SCI was produced by transient extradural application of an aneurysm clip, which exerted a closing force of approximately 24 g on the spinal cord at T6–7 levels for 1 minute. This technique created a model of complete and irreversible SCI with complete paralysis of the lower extremities (Rivlin and Tator, 1978). After removal of the clip, the skin incision was closed and the rats were singly housed in a temperature-controlled room at 27°C for a survival period of 28 days. Manual massage of urinary bladder was performed twice daily until autonomous bladder voidance reflex developed.
The 96 rats were randomly allocated into six groups (n = 16). In the sham group, rats were subjected to laminectomy alone. In the SCI group, rats received laminectomy with SCI. In the SCI + vehicle group, rats were intraperitoneally injected with 0.9% saline after SCI. In the SCI + MP group, rats were intraperitoneally injected with MP (30 mg/kg at 1 hour, 15 mg/kg at 24 and 48 hours; Mustafa Nevzat Ilac Sanayi A.S., Turkey) after SCI. In the SCI + MP + RSG group, rats were intraperitoneally injected with RSG (2 mg/kg at 1 hour, and once every 12 hours for 7 days; Avandia GlaxoSmithKline, Philadelphia, PA, USA) after SCI. In the combined treatment group, rats were intraperitoneally injected with MP (30 mg/kg at 1 hour, 15 mg/kg at 24 and 48 hours) and RSG (2 mg/kg at 1 hour, and once every 12 hours for 7 days) after SCI. Among 16 rats, 10 were sacrificed 24 hours after SCI for myeloperoxidase (MPO), enzyme linked immunosorbent assay (ELISA), terminal dexynucleotidyl transferase-mediated dUTP nick end labeling (TUNEL), and western blot assays; the remaining six rats were used for functional assessment. The dose regimen used in the present study was chosen based on results from our preliminary dose-dependent study.
MPO activity assay
MPO activity, an indicator of neutrophil infiltration, was determined in spinal cord tissues at 24 hours post-injury as previously described (Mullane, 1989). MPO activity was measured in each sample according to manufacture instructions (Nanjing Jiancheng Biological Institute, Nanjing, China) and was recorded at U/g wet tissue.
Protein expression of tumor necrosis factor-alpha (TNF-α) and interleukin-1 beta (IL-1β)
Portions of spinal cord tissues collected at 24 hours after SCI were rapidly dissected and homogenized in 1 mL PBS containing protease inhibitors. TNF-α and IL-1β expression levels were assayed using the DuoSet ELISA Development System (R&D Systems Inc., Minneapolis, MN, USA). All assays were performed in duplicate using recommended buffers, diluents, and substrates. Standard samples and tissue samples were aliquoted into 96-well plates, and the optical density at 450 nm was measured for each well using a microplate reader. The optical densities for each sample were compared with a standard TNF-α and IL-1β concentration curve created in Excel to quantify serum TNF-α and IL-1β expression.
TUNEL assay
TUNEL assay was conducted using a TUNEL detection kit according to manufacture instructions (Roche, Basel, Switzerland) at 24 hours after SCI (Darzynkiewicz, 2008). Slides were observed by light microscopy and neurons with brown-stained nuclei or containing apoptotic bodies were considered apoptotic. All TUNEL-positive cells were counted and examined for typical pathological features of apoptosis. The mean number of TUNEL-positive cells in each group was calculated, and the apoptotic index was expressed as (TUNEL-positive cells/total cells) × 100%. Independent scoring was performed by a blinded investigator.
Western blot assay of Bax and Bcl-2 protein expression
Western blot assay was performed to determine expression of Bax and Bcl-2 protein within the injured spinal cord at 24 hours after SCI. Tissue samples from SCI-injured animals were collected and homogenized on ice in 10 mM Tris-HCl buffer (pH 7.4), 10 mM ethylenediamine tetraacetic acid, 30% TritonX-100, 10% sodium dodecyl sulfate, and NaCl using a homogenizer. Supernatant was collected and stored at –80°C. Samples (40 μg total protein/well) were subjected to 10–14% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and were electro-transferred to nitrocellulose membranes. The membranes were then blocked in 10% non-fat dry milk in saline buffer for 1 hour and incubated in primary antibodies specific to Bax, Bcl-2, and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (Santa Cruz Biotechnology, Santa Cruz, CA, USA) overnight at 4°C. Membranes were blocked in 10% non-fat milk for 1 hour at 37°C, then incubated in rabbit anti-rat Bax, rabbit anti-rat Bcl-2, or rabbit anti-rat GAPDH antibodies (all 1:400; Santa Cruz Biotechnology, Santa Cruz, CA, USA) overnight at 4°C. After washing three times with 0.1 M Tris buffered saline (pH 7.2) containing 0.1% Tween-20 (TBST) (10 minutes each), membranes were incubated with peroxidase-conjugated bovine anti-rabbit immunoglobulin G (1:2,000; Santa Cruz Biotechnology) for 2 hours at 37°C and washed three times with TBST (10 minutes each). Immunoreactive protein bands were visualized by enhanced chemiluminescence, according to manufacture instructions (Pierce, Rockford, IL, USA). Band optical density values were determined using Gel-Pro Analyzer Software, version 4.0 (Media Cybernetics, USA), and Bax and Bcl-2 optical density values were expressed relative to GAPDH.
Basso, Beattie, and Bresnahan (BBB) assessment
Functional assessment was performed using the BBB (Basso et al., 1995) locomotor recovery scale on postoperative days 3, 7, 14, 21, and 28. The scale was based on the precise observation of hindlimb movements, stepping, and coordination in an open field. Rats were observed for 4 minutes by an observer blinded to the treatments and their behaviors were scored from 0 (no spontaneous locomotor activity) to 21 (normal movement-coordinated gait, with parallel paw placement). Scores from 0 to 7 indicate return of isolated movements in the hip, knee, and ankle joints. Scores from 8 to 13 indicate return of paw placement and coordinated movements with the forelimbs. Scores from 14 to 21 show return of toe clearance during stepping, predominant paw position, trunk stability, and tail position. Scores are represented as the average score of all animals (n = 6) in each group at each time point.
Statistical analysis
Data are expressed as the mean ± SD and were analyzed with SPSS 13.0 software (SPSS, Chicago, IL, USA). Multiple group comparisons of differences in quantitative measurement were made using one-way analysis of variance followed by Dunnett's t-test. A value of P < 0.05 was considered statistically significant.
Results
MPO activity after SCI
We investigated the effect of combined treatment with MP and RSG on MPO activity (Figure 1). MPO activity was significantly elevated in the spinal cord at 24 hours post-injury in rats with SCI compared with sham-operated rats (P < 0.01). MPO activity was significantly attenuated by early treatment with MP or RSG alone (P < 0.05). The protective effect of single treatment was further significantly enhanced by combined treatment with MP and RSG (P < 0.05).
Figure 1.
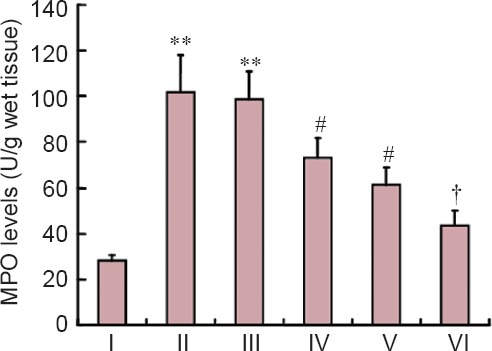
Effects of combined treatment with MP and RSG on MPO activity.
MPO activity in spinal cord after SCI was significantly increased at 24 hours after injury. **P < 0.01, vs. sham group; #P < 0.05, vs. SCI group and SCI + vehicle groups. †P < 0.05, vs. SCI + RSG group and SCI + MP group. Data are expressed as the mean ± SD (n = 10, one-way analysis of variance followed by Dunnett's t-test). SCI: Spinal cord injury; MP: methylprednisolone; RSG: rosiglitazone; MPO: myeloperoxidase. I: Sham; II: SCI; III: SCI + vehicle; IV: SCI + MP; V: SCI + RSG; VI: SCI + MP + RSG.
Effects of combined treatment with MP and RSG on TNF-α and IL-1β expression after SCI
To determine whether combined treatment with MP and RSG modulates the inflammatory process by regulating secretion of pro-inflammatory cytokines, we analyzed TNF-α and IL-1β expression levels (Figure 2). Rats with SCI showed dramatically up-regulated TNF-α and IL-1β expression in the injured regions of the spinal cord compared with rats with an intact spinal cord (P < 0.01). Although MP or RSG treatment alone significantly reduced TNF-α and IL-1β levels at 24 hours post-injury (P < 0.05), TNF-α and IL-1β levels were lower in the SCI + MP + RSG group than in the SCI + RSG group (P < 0.05).
Figure 2.

Effects of combined treatment with MP and RSG on TNF-α and IL-1β expression.
(A, B) TNF-α and IL-1β levels at the injury site were significantly enhanced at 24 hours after SCI. **P < 0.01, vs. sham group; #P < 0.05, vs. SCI group and SCI + vehicle group. †P < 0.05, vs. SCI + RSG group and SCI + MP group. Data are expressed as the mean ± SD (n = 10, one-way analysis of variance followed by Dunnett's t-test). SCI: Spinal cord injury; MP: methylprednisolone; RSG: rosiglitazone; TNF-α: tumor necrosis factor-α; IL-1β: interleukin-1β. I: Sham; II: SCI; III: SCI + vehicle; IV: SCI + MP; V: SCI + RSG; VI: SCI + MP + RSG.
Effects of combined treatment with MP and RSG on cell apoptosis after SCI
Almost no apoptotic cells were detected in the spinal cords of sham-operated rats (Figure 3A). However, many TUNEL-positive cells were observed within spinal cord tissues from non-treated and vehicle-treated rats (P < 0.01; Figures 3B, C).
Figure 3.

Effects of combined treatment with MP and RSG on cell apoptosis after SCI.
(A) Apoptotic cells were quantified at 24 hours after SCI. Almost no TUNEL-positive cells were observed in the sham group. (B, C) Numerous TUNEL-positive cells are detected in the SCI and SCI + vehicle groups. (D, E) The number of TUNEL-positive cells after SCI was significantly decreased in the SCI + RSG and SCI + MP groups. (F) There are significantly less TUNEL-positive cells in the SCI + MP + RSG group compared with the SCI + RSG group and the SCI + MP group. Scare bar: 100 μm. (G) Cell apoptosis index [(TUNEL-positive cells/total cells) × 100%] after SCI. **P < 0.01, vs. sham group; #P < 0.05, vs. SCI group and SCI + vehicle group; †P < 0.05, vs. + RSG group and SCI + MP group. Data are expressed as the mean ± SD (n = 10; one-way analysis of variance followed by Dunnett's; t-test). SCI: Spinal cord injury; MP: methylprednisolone; RSG: rosiglitazone; TUNEL: terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling. I: Sham; II: SCI; III: SCI + vehicle; IV: SCI + MP; V: SCI + RSG; VI: SCI + MP + RSG.
MP, RSG, and the combination of both significantly reduced the number of TUNEL-positive cells compared with vehicle-treatment (P < 0.05; Figure 3D–F). However, there was a significant difference between the SCI + MP + RSG group and the SCI + MP and SCI + RSG groups (P < 0.05).
Effects of combined treatment with MP and RSG on Bax and Bcl-2 expression after SCI
At 24 hours post-injury, Bax levels were appreciably increased in the spinal cord of rats subjected to SCI. Bcl-2 expression was significantly decreased in whole extracts obtained from spinal cord tissues at 24 hours after SCI. MP and RSG administered as a single treatment significantly reduced SCI-induced Bax expression and blunted SCI-induced inhibition of Bcl-2 expression, respectively. However, combined treatment with MP and RSG had a more pronounced effect on SCI-induced Bax and Bcl-2 expression (Figure 4).
Figure 4.
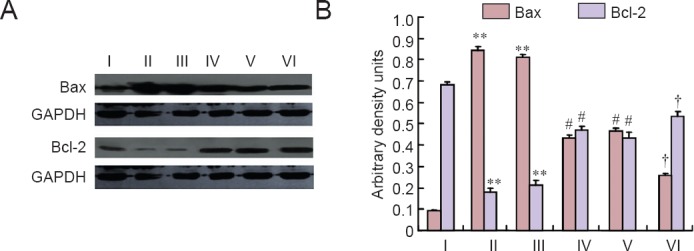
Effects of combined MP and RSG treatment on Bax and Bcl-2 expression at 24 hours after SCI.
Western blot assay was performed in spinal cord samples collected at 24 hours after injury. Representative immunoblots of Bax and Bcl-2 in the sham group, SCI group, SCI + vehicle group, SCI + MP group, SCI + RSG group, and SCI + MP + RSG group. GAPDH was used as a loading control. At 24 hours after SCI, Bax levels were significantly increased and Bcl-2 expression was significantly decreased in the SCI and SCI + vehicle groups. Combined treatment with MP and RSG significantly inhibited Bax expression and blunts SCI-induced inhibition of Bcl-2 expression compared with other treatment groups. **P < 0.01, vs. sham group; #P < 0.05, vs. SCI group and SCI + vehicle group; †P < 0.05, vs. SCI + RSG groups and SCI + MP group. Data are expressed as the mean ± SD (n = 10; one-way analysis of variance followed by Dunnett's t-test). SCI: Spinal cord injury; MP: methylprednisolone; RSG: rosiglitazone; GAPDH: glyceraldehyde-3-phosphate dehydrogenase. I: Sham; II: SCI; III: SCI + vehicle; IV: SCI + MP; V: SCI + RSG; VI: SCI + MP + RSG.
Effects of combined treatment with MP and RSG on functional recovery after SCI
BBB locomotor recovery scale after SCI in all six groups is shown in Figure 5. Spinal cord contusion resulted in a score of 0 with bilateral hind limb paralysis in all rats subjected to SCI. Although spontaneous recovery of function was observed over the next 3 weeks after SCI, neurological function in rats of the SCI + MP group, SCI + RSG group, and SCI + MP + RSG group improved more rapidly and showed statistically significantly better scores compared with the SCI group and SCI + vehicle group (P < 0.05). However, there was a significant difference in BBB locomotor recovery scales between combined treatment group and single treatment group from 3 to 21 days after injury (P < 0.05).
Figure 5.
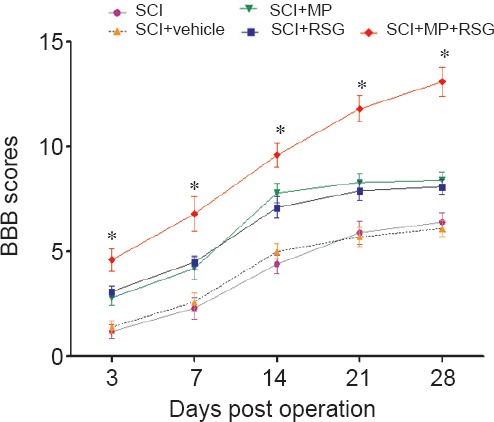
Effects of combined treatment with MP and RSG on locomotor activity during 4 weeks following SCI.
Combined treatment with MP and RSG significantly ameliorated hindlimb motor disturbances compared with other groups. *P < 0.05, vs. other groups. Data are expressed as the mean ± SD (n = 6; one-way analysis of variance followed by Dunnett's t-test). Higher BBB scores indicate better neurological function. SCI: Spinal cord injury; MP: methylprednisolone; RSG: rosiglitazone; BBB: Basso, Beattie and Bresnahan.
Discussion
The current study demonstrated that combined treatment with MP and RSG after spinal cord contusion significantly suppressed infiltration of neutrophils, proinflammatory cytokine expression, and occurrence of neuronal apoptosis compared with MP or RSG alone. The combined treatment group also exhibited greater locomotor recovery during the 3-week follow-up period. The difference in neurological and histological outcomes between the single and combined treatment groups could be explained by the synergistic or additive effect of MP and RSG during the acute phase of SCI.
The neuroprotective effects of high-dose MP on SCI have been previously reported. Anti-inflammatory actions and reduction of lipid peroxidation have been postulated to be the two major factors in improved outcomes associated with MP (Hsu et al., 1990; Hall, 1992; Nadeau and Rivest, 2003). RSG is a novel experimental therapy for SCI that mitigates secondary damage by activating peroxisome proliferator-activated receptor-γ in the central nervous system. Peroxisome proliferator-activated receptor-γ is a ligand-activated transcription factor from the nuclear hormone receptor superfamily, and is thought to modulate multiple cellular functions, including glucose absorption, lipid metabolism, cell growth and differentiation, apoptosis, and inflammation (Tolman and Chandramouli, 2003). Recent studies showed that RSG is extremely neuroprotective in animal models of acute central nervous system insults, including neurodegonerative diseases, focal ischemia, SCI, and surgical trauma (Sundararajan et al., 2005; Pereira et al., 2006; Park et al., 2007; Hyong et al., 2008; Madeira et al., 2015). However, it is unrealistic to achieve disease remission by blocking a single early mediator in the inflammatory cascade, because a large number of inflammatory mediators are involved in the secondary injury processes after SCI. Therefore, combination therapies with drugs that can target more than one mechanism are beneficial. Additionally, compounds that simultaneously target multiple pathways can be more effective.
Secondary degeneration after traumatic SCI is affected by a variety of cellular and molecular mechanisms. Inflammation is one important mechanism that may exacerbate the effects of primary mechanical trauma. During the acute phase of SCI, an inflammatory reaction develops within hours after initial injury and is characterized by the neutrophil infiltration and microglial activation (McTigue et al., 1989). These reactive cells produce several proinflammatory cytokines, including IL-1, TNF-α, IL-6, and IL-3, which mediate the inflammatory response and regulate cellular events after SCI.
The proinflammatory cytokines TNF-α and IL-1β play an important function in the induction of inducible nitric oxide synthase, which is known to have a key role in the development of SCI (Genovese et al., 2006). Several experimental studies have confirmed that TNF-α and IL-1β levels in the injured spinal cord are significantly upregulated within the first few hours after SCI, which significantly influences the degree of secondary injury after SCI (Blight, 1992). Even within 1 hour after injury, increased TNF-α and IL-1β mRNA expression is detected within the spinal cord (Bartholdi and Schwab, 1997; Wang et al., 1997). In the current study, the number of MPO-positive neutrophils and TNF-α and IL-1β expression were significantly increased in the injured rats compared with sham-operated rats at 24 hours post-injury. Although, to a certain extent, administration of MP or RSG alone reduced the inflammatory response following SCI, the combined treatment attenuated infiltration of neutrophils and upregulated TNF-α and IL-1β expression to a greater degree within spinal cord after injury.
The current study also demonstrated that the number of TUNEL-positive cells in the combined treatment group was significantly less than in the single treatment groups at the injury site. Recent studies have revealed that most neuronal and glial cell loss after injury results from apoptosis (Charriaut-Marlangue et al., 1996; Crowe et al., 1997; Kato et al., 1997; Liu et al., 1997). Additionally, apoptotic death of oligodendrocytes is also observed along the longitudinal axis of the spinal cord, which is involved in the delayed and prolonged process of demyelination and deterioration of sensorimotor function of the cord (Abe et al., 1999). Therefore, interventions targeting apoptosis may be beneficial for preventing secondary degeneration following traumatic SCI. Apoptosis can be triggered by intracellular and extracellular signals and involves a complex cascade of molecular and biochemical reactions in the cell. It has been well demonstrated that Bax, a pro-apoptotic gene, plays an important role in developmental cell death and in central nervous system injury (Wu et al., 2006). Likewise, administration of Bcl-2 protein into the injured spinal cord increases neuronal survival, suggesting that the SCI-induced decrease in Bcl-2 expression contributes considerably to neuronal death (Lu et al., 2006). In this study, Bax levels appreciably increased in spinal cords from rats subjected to SCI at 24 hours post-injury, while Bcl-2 expression significantly decreased in SCI rats at 24 hours post-injury. Combined treatment with MP and RSG significantly decreased Bax expression and increased Bcl-2 expression following SCI. These results suggest that combined treatment reduced apoptotic cell death by preventing the attenuation of the anti-apoptotic pathway and blocking activation of the pro-apoptotic pathway.
During the 3-week follow-up period, gradual functional recovery was assessed using the BBB locomotor recovery scale in all rats with SCI. However, BBB scores were significantly better in the combined treatment group than in other experimental groups. Better BBB scores indicated improved coordination between different muscles, better timing sequence and scaling of muscle activity, and progress in inter-joint and inter-limb coordination. These findings demonstrated that combined treatment with MP and RSG significantly increased recovery of motor function in SCI rats. Our results suggested that RSG administration after spinal cord contusion attenuated inflammatory reactions and diminished apoptosis, resulting in improved locomotor activity.
In conclusion, the strategies targeting multiple proinflammatory pathways may be more effective than targeting a single effector molecule. Combined treatment with MP and RSG exerts therapeutic effects on SCI.
Footnotes
Conflicts of interest: None declared.
Plagiarism check: This paper was screened twice using CrossCheck to verify originality before publication.
Peer review: This paper was double-blinded and stringently reviewed by international expert reviewers.
Copyedited by Cooper C, Hindle A, Wang J, Qiu Y, Li CH, Song LP, Zhao M
References
- Abe Y, Yamamoto T, Sugiyama Y, Watanabe T, Saito N, Kayama H, Kumagai T. Apoptotic cells associated with Wallerian degeneration after experimental spinal cord injury: a possible mechanism of oligodendroglial death. J Neurotrauma. 1999;16:946–952. doi: 10.1089/neu.1999.16.945. [DOI] [PubMed] [Google Scholar]
- Baffour R, Achanta K, Kaufman J, Berman J, Garb JL, Rhee S, Friedmann P. Synergistic effect of basic fibroblast growth factor and methylprednisolone on neurological function after experimental spinal cord injury. J Neurosurg. 1995;83:105–110. doi: 10.3171/jns.1995.83.1.0105. [DOI] [PubMed] [Google Scholar]
- Bartholdi D, Schwab ME. Expression of pro-inflammatory cytokine and chemokine mRNA upon experimental spinal cord injury in mouse: an in situ hybridization study. Eur J Neurosci. 1997;9:1422–1438. doi: 10.1111/j.1460-9568.1997.tb01497.x. [DOI] [PubMed] [Google Scholar]
- Basso DM, Beattie MS, Bresnahan JC. A sensitive and reliable locomotor rating scale for open field testing in rats. J Neurotrauma. 1995;12:1–21. doi: 10.1089/neu.1995.12.1. [DOI] [PubMed] [Google Scholar]
- Blight AR. Macrophages and inflammatory damage in spinal cord injury. J Neurotrauma. 1992;9:83–91. [PubMed] [Google Scholar]
- Charriaut-Marlangue C, Margaill I, Represa A, Popovici T, Plotkine M, Ben-Ari Y. Apoptosis and necrosis after reversible focal ischemia: an in situ DNA fragmentation analysis. J Cereb Blood Flow Metab. 1996;16:186–194. doi: 10.1097/00004647-199603000-00002. [DOI] [PubMed] [Google Scholar]
- Collins WF. A review and update of experimental and clinical studies of spinal cord injury. Paraplegia. 1983;21:204–219. doi: 10.1038/sc.1983.34. [DOI] [PubMed] [Google Scholar]
- Crowe MJ, Bresnahan JC, Shuman SL, Masters JN, Beattie MS. Apoptosis and delayed degeneration after spinal cord injury in rats and monkeys. Nat Med. 1997;3:73–76. doi: 10.1038/nm0197-73. [DOI] [PubMed] [Google Scholar]
- Díaz-Ruiz A, Ibarra A, Pérez-Severiano F, Guízar-Sahagún G, Grijalva I, Ríos C. Constitutive and inducible nitric oxide synthase activities after spinal cord contusion in rats. Neurosci Lett. 2002;319:129–132. doi: 10.1016/s0304-3940(01)02540-x. [DOI] [PubMed] [Google Scholar]
- Darzynkiewicz Z, Galkowski D, Zhao H. Analysis of apoptosis by cytometry using TUNEL assay. Methods. 2008;44:250–254. doi: 10.1016/j.ymeth.2007.11.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Genovese T, Mazzon E, Crisafulli C, Di Paola R, Muià C, Bramanti P, Cuzzocrea S. Immunomodulatory effects of etanercept in an experimental model of spinal cord injury. J Pharmacol ExpTher. 2006;316:1006–1016. doi: 10.1124/jpet.105.097188. [DOI] [PubMed] [Google Scholar]
- Hall ED. The neuroprotective pharmacology of methylprednisolone. J Neurosurg. 1992;76:13–22. doi: 10.3171/jns.1992.76.1.0013. [DOI] [PubMed] [Google Scholar]
- Hausmann ON. Post-traumatic inflammation following spinal cord injury. Spinal Cord. 2003;41:369–378. doi: 10.1038/sj.sc.3101483. [DOI] [PubMed] [Google Scholar]
- Hsu CY, Dimitrijevic MR. Methylprednisolone in spinal cord injury: the possible mechanism of action. J Neurotrauma. 1990;7:115–119. doi: 10.1089/neu.1990.7.115. [DOI] [PubMed] [Google Scholar]
- Hyong A, Jadhav V, Lee S, Tong W, Rowe J, Zhang JH, Tang J. Rosiglitazone, a PPAR gamma agonist, attenuates inflammation after surgical brain injury in rodents. Brain Res. 2008;1215:218–224. doi: 10.1016/j.brainres.2008.04.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kato H, Kanellopoulos GK, Matsuo S, Wu YJ, Jacquin MF, Hsu CY, Kouchoukos NT, Choi DW. Neuronal apoptosis and necrosis following spinal cord ischemia in the rat. Exp Neurol. 1997;148:464–474. doi: 10.1006/exnr.1997.6707. [DOI] [PubMed] [Google Scholar]
- Liu XZ, Xu XM, Hu R, Du C, Zhang SX, McDonald JW, Dong HX, Wu YJ, Fan GS, Jacquin MF, Hsu CY, Choi DW. Neuronal and glial apoptosis after traumatic spinal cord injury. J Neurosci. 1997;17:5395–5406. doi: 10.1523/JNEUROSCI.17-14-05395.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu B, Wang L, Stehlik C, Medan D, Huang C, Hu S, Chen F, Shi X, Rojanasakul Y. Phosphatidylinositol 3-kinase/Akt positively regulates Fas (CD95)-mediated apoptosis in epidermal Cl41 cells. J Immunol. 2006;176:6785–6793. doi: 10.4049/jimmunol.176.11.6785. [DOI] [PubMed] [Google Scholar]
- Madeira JM, Schindler SM, Klegeris A. A new look at auranofin, dextromethorphan and rosiglitazone for reduction of glia-mediated inflammation in neurodegenerative diseases. Neural Regen Res. 2015;10:391–393. doi: 10.4103/1673-5374.153686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McTigue DM, Tani M, Krivacic K, Chernosky A, Kelner GS, Maciejewski D, Maki R, Ransohoff RM, Stokes BT. Selective chemokine mRNA accumulation in the rat spinal cord after contusion injury. J Neurosci Res. 1989;53:368–376. doi: 10.1002/(SICI)1097-4547(19980801)53:3<368::AID-JNR11>3.0.CO;2-1. [DOI] [PubMed] [Google Scholar]
- Mullane K. Neutrophil-platelet interactions and post-ischemic myocardial injury. Prog Clin Biol Res. 1989;301:39–51. [PubMed] [Google Scholar]
- Nadeau S, Rivest S. Glucocorticoids play a fundamental role in protecting the brain during innate immune responses. J Neurosci. 2003;23:5336–5544. doi: 10.1523/JNEUROSCI.23-13-05536.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nash HH, Borke RC, Anders JJ. Ensheathing cells and methylprednisolone promote axonal regeneration and functional recovery in the lesioned adult rat spinal cord. J Neurosci. 2002;22:7111–7120. doi: 10.1523/JNEUROSCI.22-16-07111.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park SW, Yi JH, Miranpuri G, Satriotomo I, Bowen K, Resnick DK, Vemuganti R. Thiazolidinedione class of peroxisome proliferator-activated receptor gamma agonists prevents neuronal damage, motor dysfunction, myelin loss, neuropathic pain, and inflammation after spinal cord injury in adult rats. J Pharmacol Exp Ther. 2007;320:1002–1012. doi: 10.1124/jpet.106.113472. [DOI] [PubMed] [Google Scholar]
- Pereira MP, Hurtado O, Cárdenas A, Boscá L, Castillo J, Dávalos A, Vivancos J, Serena J, Lorenzo P, Lizasoain I, Moro MA. Rosiglitazone and 15-deoxy-delta12, 14-prostaglandin J2 cause potent neuroprotection after experimental stroke through noncompletely overlapping mechanisms. J Cereb Blood Flow Metab. 2006;26:218–229. doi: 10.1038/sj.jcbfm.9600182. [DOI] [PubMed] [Google Scholar]
- Profyris C, Cheema SS, Zang D, Azari MF, Boyle K, Petratos S. Degenerative and regenerative mechanisms governing spinal cord injury. Neurobiol Dis. 2004;15:415–436. doi: 10.1016/j.nbd.2003.11.015. [DOI] [PubMed] [Google Scholar]
- Rintala DH, Loubser PG, Castro J, Hart KA, Fuhrer MJ. Chronic pain in a community-based sample of men with spinal cord injury: prevalence, severity, and relationship with impairment, disability, handicap, and subjective well-being. Arch Phys Med Rehabil. 1998;79:604–614. doi: 10.1016/s0003-9993(98)90032-6. [DOI] [PubMed] [Google Scholar]
- Rivlin AS, Tator CH. Effect of duration of acute spinal cord compression in a new acute cord injury model in the rat. Surg Neurol. 1978;10:38–43. [PubMed] [Google Scholar]
- Sayer FT, Kronvall E, Nilsson OG. Methylprednisolone treatment in acute spinal cord injury: the myth challenged through a structured analysis of published literature. Spine J. 2006;6:335–343. doi: 10.1016/j.spinee.2005.11.001. [DOI] [PubMed] [Google Scholar]
- Short DJ, El Masry WS, Jones PW. High dose methylprednisolone in the management of acute spinal cord injury-a systematic review from a clinical perspective. Spinal Cord. 2000;38:273–286. doi: 10.1038/sj.sc.3100986. [DOI] [PubMed] [Google Scholar]
- Sundararajan S, Gamboa JL, Victor NA, Wanderi EW, Lust WD, Landreth GE. Peroxisome proliferator-activated receptor-gamma ligands reduce inflammation and infarction size in transient focal ischemia. Neuroscience. 2005;130:685–696. doi: 10.1016/j.neuroscience.2004.10.021. [DOI] [PubMed] [Google Scholar]
- Takami T, Oudega M, Bethea JR, Wood PM, Kleitman N, Bunge MB. Methylprednisolone and interleukin-10 reduce gray matter damage in the contused Fischer rat thoracic spinal cord but do not improve functional outcome. J Neurotrauma. 2002;19:653–666. doi: 10.1089/089771502753754118. [DOI] [PubMed] [Google Scholar]
- Tolman KG, Chandramouli J. Hepatotoxicity of the thiazolidinediones. Clin Liver Dis. 2003;7:369–379. doi: 10.1016/s1089-3261(03)00020-5. [DOI] [PubMed] [Google Scholar]
- Wang CX, Olschowka JA, Wrathall JR. Increase of interleukin-1beta mRNA and protein in the spinal cord following experimental traumatic injury in the rat. Brain Res. 1997;759:190–196. doi: 10.1016/s0006-8993(97)00254-0. [DOI] [PubMed] [Google Scholar]
- Wu CH, Ding XY, Wang HY, Ye XB, Huang SY, Huang AM, Li HZ, Wu SY, Yu J, Yan XH. Neural apoptosis and apoptosis-related genes in intracerebral hemorrhage patients. Zhonghua Yi Xue Za Zhi. 2006;86:3073–3076. [PubMed] [Google Scholar]
- Yan P, Xu J, Li Q, Chen S, Kim GM, Hsu CY, Xu XM. Glucocorticoid receptor expression in the spinal cord after traumatic injury in adult rats. J Neurosci. 1999;19:9355–9363. doi: 10.1523/JNEUROSCI.19-21-09355.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]