

Abstract
Background
Mouse infection studies have shown that interferon-γ (IFN-γ), a T helper 1 (Th1) cytokine, is required for the development of severe pathology induced by chronic Helicobacter infection. This finding is largely based on studies performed using mice that have polarised Th1 responses i.e. C57BL/6 animals. The current work aims to investigate the role of IFN-γ in Helicobacter-induced inflammation in BALB/c mice which have Th2-polarised immune responses.
Results
At 7 months post-infection with Helicobacter felis, IFN-γ deficiency in BALB/c mice had no significant effect on H. felis colonisation levels in the gastric mucosa, nor on humoral responses, or gastritis severity. Ifng −/− animals with chronic H. felis infection did, however, develop significantly fewer lymphoid follicle lesions, as well as increased IL-4 splenocyte responses, when compared with infected Ifng +/+ mice (P = 0.015 and P = 0.0004, respectively).
Conclusions
The work shows that in mice on a BALB/c background, IFN-γ is not required for bacterial clearance, antibody responses, nor gastric inflammation. Conversely, IFN-γ appears to play a role in the development of gastric lymphoid follicles, which are precursor lesions to mucosa-associated lymphoid tissue (MALT) lymphoma. This study highlights the importance of mouse host background on the susceptibility to Helicobacter-induced pathologies.
Keywords: Helicobacter, Interferon-gamma, Lymphoid follicle, Mucosa-associated lymphoid tissue, MALT lymphoma, BALB/c, T helper response, Gastric inflammation
Background
The prolonged immune responses generated during Helicobacter pylori infection in humans drive the development of diseases that vary in severity, ranging from peptic ulcers to gastric adenocarcinomas and lymphomas [1]. One of the key soluble mediators induced during Helicobacter infection is interferon-γ (IFN-γ), a pro-inflammatory cytokine that contributes to gastric inflammation and is a hallmark of T helper (Th) type 1 responses [2–4].
Helicobacter infection has been reported to induce both Th1 and Th2 responses [5–7]. Th1 responses are characterised by immune cell production of IFN-γ and interleukin-12 (IL-12) [8, 9], whereas Th2 responses are associated with strong humoral responses and the production of cytokines, such as IL-4 and IL-13 [10–12]. Intraepithelial lymphocytes have been shown to be a major source of IFN-γ and IL-4 production within the gastric mucosa of Helicobacter-infected murine and human hosts [2, 3, 5, 13]. High levels of IFN-γ production have been observed in both gastric CD4+ T cells and splenocyte cultures from infected hosts, thereby implicating severe inflammation with skewed Th1 responses [5, 9]. IFN-γ was also reported to play a role in bacterial clearance [4, 13].
Mice that have been experimentally infected with the feline/canine Helicobacter sp., Helicobacter felis, develop many aspects of the pathological changes observed in H. pylori-infected humans, including the progression from chronic gastritis to cancer, commonly known as the “Correa model” [1, 14–18]. Although the type(s) and severity of pathology observed in Helicobacter infection models are known to be influenced by the genetic background of the mice [3, 19, 20], little is still known regarding the role of host factors in Helicobacter-induced immunopathology. Previously, we proposed that the default Th phenotypes of mice may be one factor contributing to the different types of pathology seen in animals [21].
Wild-type C57BL/6 mice are relatively susceptible to Helicobacter infection and develop severe atrophic gastritis and pre-neoplastic lesions [1, 19, 22]. In contrast, mice on a BALB/c background appear to be more resistant to Helicobacter infection [20, 22, 23] and usually develop little to no gastritis in the first few months post-infection [16, 22, 24]. However, after long-term infection (i.e. 18–24 months) with either H. pylori or H. felis, BALB/c mice develop structured gastric lymphoid follicles resembling mucosa-associated lymphoid tissue (MALT) lymphoma in humans [16, 20, 22, 25].
Many of the Helicobacter infection studies in the literature have used mice on a C57BL/6 background. These animals have Th1-polarised immune responses [7, 9, 26]. Given that IFN-γ is a key mediator of these responses [7, 9, 26], we sought to determine whether IFN-γ is required for Helicobacter-induced inflammation in mice with Th2-biased responses. To address this question, we used H. felis to experimentally infect Ifng +/+ and Ifng −/− mice on a BALB/c background which have Th2-polarised immune responses. From these experiments, we were able to show that IFN-γ plays no role in cellular or humoral immune responses to chronic H. felis infection in BALB/c mice. Importantly, however, the absence of IFN-γ in BALB/c animals resulted in a significant reduction in lymphoid aggregate formation, when compared with infected Ifng +/+ mice. These results demonstrate the influence of the mouse genetic background on the pathology observed in Helicobacter infection models.
Methods
Animals
Ifng +/+ and Ifng −/− BALB/c mice (The Jackson Laboratory, Bar Harbor, ME, USA) were housed in polycarbonate cages in isolators and fed a commercial pellet diet with water. Groups of mice (n = 8–10) were inoculated with >106 H. felis CS1 bacteria or given broth alone [21]. At 7 months post-infection, mice were sacrificed. All animal experimentation was performed in accordance with institutional guidelines, prescribed by the committee of Hygiène Sécurité et Protection de L’Environnement (Institut Pasteur), according to French Law 87-848.
Samples
Gastric tissues from mice were divided into two sections, each containing the antrum and body mucosa and used to determine H. felis colonisation, gastric antibody levels and histology studies [21]. Sera were collected in Sarstedt microtubes (Sarstedt) and stored at −20 °C for further testing. Gastric secretions were collected as described previously [27]. Briefly, mouse stomachs were opened along the greater curvature and the contents released into phosphate-buffered saline (pH 7.4) in six-well tissue culture plates (Falcon, Becton–Dickinson Labware). Protease inhibitors were added to samples and stored at −20 °C. Spleens were collected in RPMI 1640 medium (Gibco) containing 5% (vol/vol) foetal calf serum, 200 mM l-glutamine, 10,000 IU/mL penicillin and 10,000 mg/mL streptomycin (RPMI complete medium; all solutions from Life Technologies™) [27].
Histology
Formalin-fixed gastric tissue segments were stained with Harris’ Hematoxylin and Eosin or Giemsa and then assessed blind for histopathological changes [28] and Helicobacter felis colonisation levels [29], respectively. Briefly, inflammatory scores for polymorphoneutrophils (PMNs) and lymphocytes were graded according to a previously described six-point scheme ([21, 30]; Table 1). Lymphoid aggregates were graded according to the actual numbers of glands affected. Since H. felis does not normally form isolated colonies on culture plates, we assessed the proportion of Giemsa-stained fields according to the following scale: 0, none; 1, 1–10; 2, 10–100; and 3, >100 bacteria [29]. A total of 34–82 fields were graded accordingly per mouse. Cumulative scores for each mouse were calculated according to the formula: cumulative score = n0/t + n1/t + n2/t + n3/t, where n0, n1, n2 and n3 = the numbers of fields with respective scores of 0, 1, 2 or 3. t = total numbers of fields assessed.
Table 1.
Histological scoring scheme to grade PMN and lymphocyte inflammatory scores
| Score | Criteria | Definition of criteria |
|---|---|---|
| 0 | No inflammation | Absence of immune cells in the mucosa |
| 1 | Milda multifocal | Scattered clumps of two or three immune cells |
| 2 | Mild widespread OR moderateb multifocal | Mild widespread = widespread scattering of two or three immune cells across most of the antrum or fundus; moderate multifocal = larger clumps of immune cells seen in a few fields per region |
| 3 | Mild widespread AND moderate OR severec multifocal | As per 2. Severe multifocal = large infiltrations of immune cells across the whole width of the mucosa |
| 4 | Moderate widespread | Large clumps of immune cells seen throughout the whole width of mucosa |
| 5 | Moderate widespread and severe multifocal | As per 4 but also with areas of dense concentrations of immune cells |
| 6 | Severe widespread | Dense sheets of immune cells throughout the mucosa |
aMild inflammation was defined as an influx of inflammatory cells in the basal region of the mucosa
bModerate inflammation describes inflammatory cells extending up to the mid-region of the mucosa
cSevere inflammation describes immune cell infiltration throughout the full thickness of the mucosa
Enzyme-linked immunoassay (ELISA)
Total and H. felis-specific gastric (IgA, IgG) and serum (IgG1, IgG2a) antibodies were detected by ELISA [27]. Ninety-six well Nunc Maxisorp plates (Nunc) were coated with either goat anti-mouse IgA (SouthernBiotech) or anti-mouse IgG antibodies (GE Healthcare Life Sciences; 100 ng/well), or sonicated H. felis extracts (25 µg protein/well). Serum and gastric lavage samples were diluted 1:1000 and 1:100, respectively. Biotinylated goat anti-mouse antibodies and streptavidin-peroxidase conjugate (GE Healthcare Life Sciences) were used to detect total or antigen-specific immunoglobulins. Immune complexes were detected as described previously [21]. Total antibody levels were determined using standard curves derived from purified polyclonal IgG and monoclonal IgA mouse antibodies (Sigma). Helicobacter felis-specific IgG1 and IgG2c were determined from absorbance readings at 405 nm (test) and 492 nm (reference) wavelengths.
Splenocyte responses
To isolate single cell suspensions, tissues were homogenised using Teflon grinders and recovered by centrifugation in Ficoll-Paque solution (GE Healthcare Life Sciences), as described previously [27]. Splenocytes were recovered in RPMI complete medium and seeded at 2 × 106 cells/mL in 24-well tissue cultures. Cells were left untreated or stimulated with an anti-CD3 antibody (5 µg/mL; BD Pharmingen), then incubated for 3 days at 37 °C in 5% CO2. IL-4 and IFN-γ levels in culture supernatants were tested by sandwich ELISA (BD Pharmingen™).
Statistical analyses
All analyses were performed using Graphpad Prism version 6.0c. Data were analysed by either the Mann–Whitney test, one- or two-way analysis of variance (ANOVA), as appropriate. Presented are the median values or means and standard error of the mean (SEM). Differences were considered significant when P < 0.05.
Results
IFN-γ is not required to control H. felis colonisation in BALB/c mice
To determine whether the absence of IFN-γ in BALB/c mice may have an effect on Helicobacter bacterial burden, we infected Ifng +/+ and Ifng −/− mice on a BALB/c background with H. felis. Control Ifng +/+ mice received broth medium alone. As H. felis does not normally form isolated colonies on culture plates, colonisation levels were assessed by examination of Giemsa-stained fields [29] (Fig. 1a). All H. felis-challenged mice (n = 19) became infected (data not shown). However, no significant differences in H. felis colonisation levels were observed between Ifng +/+ and Ifng −/− mice (Fig. 1b), thus showing that IFN-γ is not required to control H. felis colonisation in the gastric mucosa of BALB/c animals.
Fig. 1.
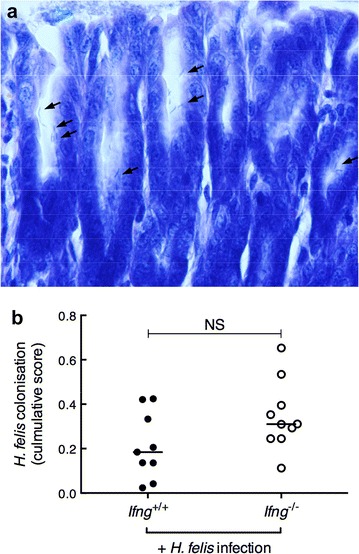
H. felis colonisation levels were similar in Ifng +/+ and Ifng −/− BALB/c mice. a Giemsa-stained gastric section showing glands lined with H. felis bacteria, indicated by arrows (magnification ×40). b H. felis colonisation levels, as determined from the assessment of Giemsa-stained sections, are presented as cumulative scores (see “Methods” section). Values for individual mice are presented. Horizontal bars correspond to the median value. Mann–Whitney test: NS not significant
Ifng−/− mice display normal gastric and serum antibody responses to H. felis infection
IFN-γ is known to play a role in mucosal antibody responses and to promote B cell isotype switching [31–34]. IFN-γ deficiency in BALB/c mice, however, had no effect on total IgA and IgG levels in gastric juices (Fig. 2). Furthermore, no differences were observed in either Helicobacter-specific IgA and IgG levels in the gastric compartment, nor in serum IgG1 and IgG2a levels, which are indicative of Th2- and Th1-responses, respectively (Fig. 3). These results indicate that IFN-γ does not regulate mucosal antibody responses in BALB/c animals during chronic H. felis infection.
Fig. 2.
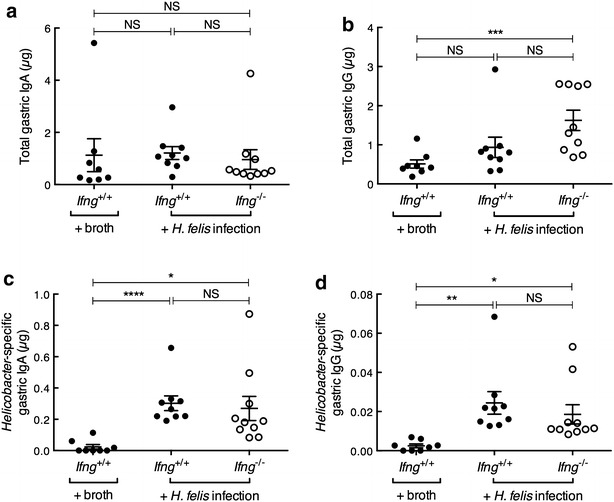
Gastric antibody levels in Ifng +/+ and Ifng −/− BALB/c mice. Total (a) IgA and (b) IgG and Helicobacter-specific (c) IgA and (d) IgG antibodies. Values for individual mice are presented and were determined from triplicate measurements. Horizontal bars correspond to the mean ± SEM. One-way ANOVA: NS not significant; *P = 0.05. **P = 0.01; ***P = 0.008; ****P = 0.005
Fig. 3.
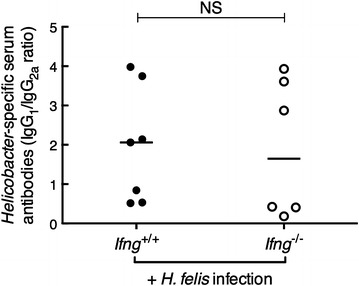
Helicobacter-specific serum IgG1 and IgG2a responses were similar in Ifng +/+ and Ifng −/− BALB/c mice. Values for individual mice are presented as ratios calculated from paired serum samples (n = 6–7/group). Horizontal bars correspond to the median value. Mann–Whitney test: NS not significant
Ifng−/− BALB/c mice exhibit normal levels of gastritis but reduced lymphoid follicle formation in response to chronic H. felis infection
We next sought to investigate whether IFN-γ may be important for histopathological changes in the gastric tissues of H. felis-infected mice on a Th2-biased BALB/c background. As controls, uninfected Ifng −/− BALB/c mice were also assessed histologically but showed a normal gland architecture (data not shown), with only some immune cells occasionally present in the submucosal region of the stomach (Fig. 4). In contrast, infected Ifng +/+ BALB/c mice displayed general thickening of the glandular tissues, immune cell infiltration of the mucosa and invasion of mucosal areas by large lymphoid cell aggregates (Fig. 4). The levels of PMN and mononuclear cell infiltration did not significantly differ between Ifng +/+ and Ifng −/− mice (Fig. 5). Gland atrophy was not detected in tissues of infected or uninfected mice (data not shown). Interestingly, Ifng −/− mice developed significantly fewer lymphoid aggregates within the gastric mucosa when compared with infected Ifng +/+ animals (Fig. 5; P = 0.015). These findings indicate that IFN-γ does not play a major role in the gastritis induced by chronic H. felis infection in BALB/c mice. In contrast, this cytokine is likely to be important for gastric lymphoid tissue formation, a precursor to MALT lymphoma.
Fig. 4.
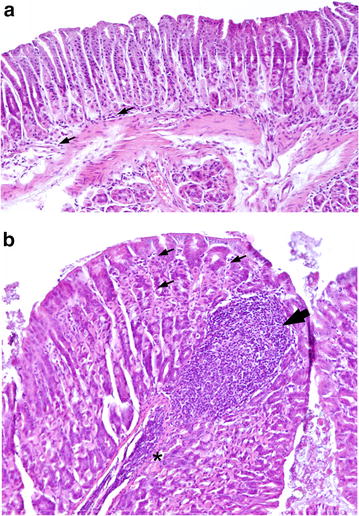
Chronic H. felis infection promoted mucosal inflammation and lymphoid cell formation in Ifng +/+ BALB/c mice when compared with uninfected animals. Gastric tissue sections from (a) uninfected and (b) H. felis-infected Ifng +/+ BALB/c mice. Small arrows show immune cells (mainly lymphocytes) present either in the (a) submucosa or (b) between the glands in the mucosal region. Massive immune cell infiltration of the submucosa (asterisk) and mucosal lymphoid cell formation (arrowhead) in an H. felis-infected mouse. Haematoxylin and Eosin stain; magnification ×10
Fig. 5.
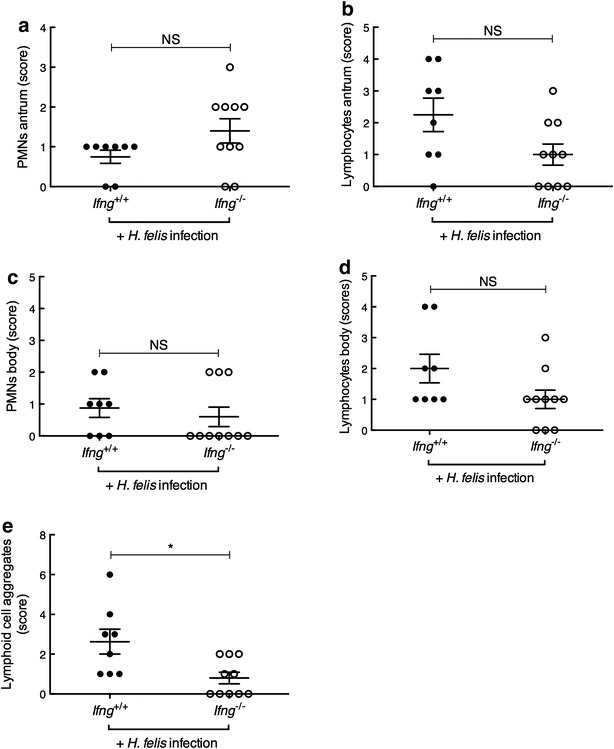
Chronic H. felis infection induced similar levels of gastritis but less lymphoid aggregate formation in Ifng −/− BALB/c mice than in infected Ifng +/+ animals. The levels of PMN and lymphocyte infiltration in the a, b antrum and c, d body of the gastric mucosa were scored blind according to the grading scheme in the “Methods” section. e Lymphoid aggregate numbers were assessed over the entirety of gastric tissue sections. Histopathological scores for individual mice are presented. One-way ANOVA: NS not significant; *P = 0.015
IFN-γ deficiency in BALB/c mice is associated with increased IL-4 splenocyte responses following chronic H. felis infection
As BALB/c mice have a polarised Th2 profile [21, 35], we speculated that the absence of Ifng −/− would result in enhanced Th2-type cytokine responses to chronic H. felis infection in these mice. Indeed, anti-CD3 treatment of splenocyte cultures from H. felis-infected Ifng −/− mice induced significantly increased levels of IL-4 production when compared with untreated splenocytes from these mice (Fig. 6; P < 0.0001). Most importantly, stimulated splenocytes from H. felis-infected Ifng −/− mice produced significantly more IL-4 when compared with splenocytes from either H. felis-infected Ifng +/+ animals or uninfected Ifng −/− mice (Fig. 6; P = 0.0004). Anti-CD3-treatment did not induce detectable levels of IFN-γ production in Ifng −/− splenocytes (Fig. 6). Taken together, the data are consistent with the Th2-polarised immune response of BALB/c mice [21, 35] and, moreover, show that chronic H. felis infection promotes an enhanced production by splenic immune cells of IL-4, a Th2 cytokine. The skewed Th2 response may influence the degree or development of gastric diseases induced by chronic H. felis infection.
Fig. 6.
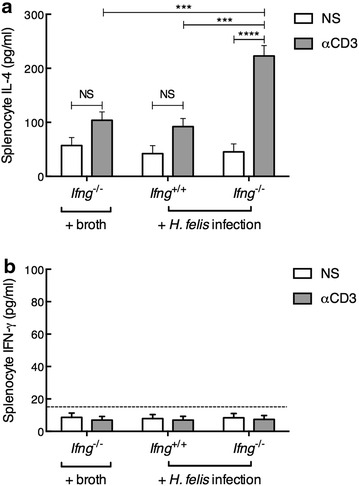
Increased IL-4 production in response to stimulation by splenocytes from H. felis-infected Ifng −/− mice when compared with infected Ifng +/+ BALB/c animals. The levels of a IL-4 and b IFN-γ production were determined in splenocyte cultures that were either not stimulated (NS) or stimulated with anti-CD3 antibody (αCD3, 5 µg/mL). Data are presented as the mean value (±SEM) determined from triplicate measurements for 8–10 mice/group. Two-way ANOVA: NS not significant; ***P = 0.004; ****P < 0.0001
Discussion
IFN-γ has been reported to be an important mediator of the immunopathology induced by chronic Helicobacter infection [4, 7, 9, 13, 36, 37] and may also be important for Helicobacter clearance, but the findings are not conclusive [4, 10, 13, 23]. Importantly, these observations were largely based on studies performed in C57BL/6 mice which have default Th1 type immune responses. We now show in a Th2-polarised mouse model that IFN-γ was not required for host defence against experimental Helicobacter infection, nor for the induction of humoral immune responses or gastritis. Interestingly, however, IFN-γ was found to be important for lymphoid aggregate formation. This work highlights the impact of the mouse genetic background on host immune responses and the accompanying immunopathology induced by chronic Helicobacter infection.
The influence of mouse genetic background on Helicobacter-induced immunopathology was first described two decades ago [3, 24], yet our understanding of the reasons for varying disease severity between mouse strains remains poor. In humans, it was reported that host factors, such as cytokine gene polymorphisms, can influence disease outcome to H. pylori infection [38]. Of particular relevance here, Th1/Th2 profiles in human hosts were shown to have an impact on the immunopathogenesis of H. pylori infection, with Th1 cytokines associated with stronger cellular responses and early stages of gastric cancer, whereas Th2 polarised responses mediated humoral immunity and advanced stages of carcinogenesis [39]. There is clear evidence that the default T helper cell responses of mouse strains contribute to inflammation severity [13, 21, 26, 36, 40, 41]. However, other immunological characteristics intrinsic to specific mouse strains, such as their major histocompatibility complex haplotype, are also likely to be important [3, 20, 21, 23, 30, 35, 42, 43].
Helicobacter-infected C57BL/6 mice develop more intense gastric inflammation than BALB/c animals [3, 9, 20, 22, 24], with disease severity correlating with higher levels of IFN-γ production [3, 4, 9]. Interestingly, H. pylori infection in Il4 −/− mice on a C57BL/6 background resulted in more severe gastritis and higher levels of IFN-γ production by stimulated splenocytes [9]. It has been suggested that the ability of IFN-γ to promote macrophage secretion of pro-inflammatory cytokines, and/or to downregulate the production of anti-inflammatory factors (e.g. IL-10, transforming growth factor β), may account for the severity of gastritis in C57BL/6 mice [14, 44, 45]. Consistent with this suggestion, Ifng −/− mice on a C57BL/6 background did not develop gastritis to H. pylori infection, while the splenocytes from these animals produced significantly more IL-4 in response to stimulation than those from wild type animals [9]. In the current study, we observed that Ifng −/− BALB/c mice exhibited enhanced Th2 responses to H. felis infection, as reflected by increased splenocyte IL-4 production (Fig. 4). Conversely, Th2-type humoral responses (Figs. 2, 3) were unchanged in the Ifng −/− BALB/c mice. Although the reason for this is unclear, a similar lack of concordance between humoral and cytokine responses was reported previously in human subjects given a measles vaccine [46].
T cell responses have been associated with the growth and development of lymphoid follicles which arise at various tissue sites, including the gastric mucosa [6, 40, 47, 48]. These follicles predominantly consist of B cell clones which form premalignant lesions, leading to gastric MALT lymphoma [48–50]. Studies using C57BL/6 mice identified IFN-γ to be an important mediator in the B and T cell interactions required for gastric MALT lymphoma formation [37, 40, 51]. Furthermore, IFN-γ was shown to be highly expressed in the gastric mucosa and to be crucial for B cell clonal expansion and lymphoid follicle formation in C57BL/6 mice infected with Helicobacter suis [37]. Interestingly, IL-4 was dispensable in this infection model [37]. The authors also showed that B lymphocytes, and not CD4+ T cells, were the main source of gastric IFN-γ and that this cytokine was secreted independently of T cell help [37]. Our study similarly demonstrates a role for IFN-γ in lymphoid aggregate formation in the gastric mucosa of H. felis-infected BALB/c mice, which are prone to develop gastric MALT lymphoma [16, 50, 52]. Asides from the potential direct actions of IFN-γ on the immune system, it is possible that its absence in Ifng −/− BALB/c mice resulted in an increased abundance of IL-4-producing Th2 cells, thereby priming naïve B cells and promoting the differentiation of naïve T cells into Th2 cells [35, 52]. Consistent with this suggestion, studies in humans and mice found that B cell proliferation in gastric MALT lymphoma was driven by Th2-polarised T cell responses [52–54]. More work is required to fully elucidate the respective roles of different Th responses in Helicobacter-induced gastric MALT lymphoma.
Conclusions
The fact that IFN-γ was not required for gastritis and bacterial control in BALB/c mice raises an interesting question regarding the role of this cytokine in Helicobacter infection. Consistent with previous studies, BALB/c mice appeared to develop less severe inflammation than that typically observed in C57BL/6 animals [18, 24]. Although the differences in pathology observed in BALB/c and C57BL/6 mice may be attributed to their respective Th profiles, it is likely that differences in other immunological and genetic factors between the two murine backgrounds may also be important [3, 20, 23, 30, 35, 43, 55]. This work highlights the importance of considering the genetic background of the host when performing immunological or vaccine studies in Helicobacter infection models. Furthermore, the findings reinforce the importance of host genetics on disease outcome, as has been observed in humans with H. pylori infection.
Authors’ contributions
MC analysed and interpreted the data, and wrote the manuscript. PA performed the experiments. MH performed histological grading and contributed to data analysis and interpretation. RLF performed the experiments, analysed and interpreted the data and revised the manuscript. All authors read and approved the final manuscript.
Acknowledgements
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Availability of data and material
The datasets during and/or analyzed during the current study are available from the corresponding author on reasonable request.
Ethics approval and consent to participate
All animal experimentation was performed in accordance with institutional guidelines and French legislation (Law 87-848).
Funding
This work was supported by a Projet Transversal de Recherche (PTR38) from the Institut Pasteur and a project grant to R.L.F. from the National Health and Medical Research Council (NHMRC, APP1107930). Research at the Hudson Institute of Medical Research is supported by the Victorian Government’s Operational Infrastructure Support Program. R.L.F is an NHMRC Senior Research Fellow (APP1079904).
Contributor Information
Michelle Chonwerawong, Email: Michelle.Chonwerawong@Hudson.org.au.
Patrick Avé, Email: patrick.ave@pasteur.fr.
Michel Huerre, Email: michel.huerre@curie.fr.
Richard L. Ferrero, Phone: +61 03 8572 2728, Email: Richard.Ferrero@Hudson.org.au
References
- 1.Correa P, Houghton J. Carcinogenesis of Helicobacter pylori. Gastroenterology. 2007;133(2):659–672. doi: 10.1053/j.gastro.2007.06.026. [DOI] [PubMed] [Google Scholar]
- 2.Karttunen R, Karttunen T, Ekre HP, MacDonald TT. Interferon gamma and interleukin 4 secreting cells in the gastric antrum in Helicobacter pylori positive and negative gastritis. Gut. 1995;36(3):341–345. doi: 10.1136/gut.36.3.341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Mohammadi M, Czinn S, Redline R, Nedrud J. Helicobacter-specific cell-mediated immune responses display a predominant Th1 phenotype and promote a delayed-type hypersensitivity response in the stomachs of mice. J Immunol. 1996;156(12):4729–4738. [PubMed] [Google Scholar]
- 4.Sawai N, Kita M, Kodama T, Tanahashi T, Yamaoka Y, Tagawa Y, Iwakura Y, Imanishi J. Role of gamma interferon in Helicobacter pylori-induced gastric inflammatory responses in a mouse model. Infect Immun. 1999;67(1):279–285. doi: 10.1128/iai.67.1.279-285.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bamford KB, Fan X, Crowe SE, Leary JF, Gourley WK, Luthra GK, Brooks EG, Graham DY, Reyes VE, Ernst PB. Lymphocytes in the human gastric mucosa during Helicobacter pylori have a T helper cell 1 phenotype. Gastroenterology. 1998;114(3):482–492. doi: 10.1016/S0016-5085(98)70531-1. [DOI] [PubMed] [Google Scholar]
- 6.D’Elios MM, Amedei A, Manghetti M, Costa F, Baldari CT, Quazi AS, Telford JL, Romagnani S, Del Prete G. Impaired T-cell regulation of B-cell growth in Helicobacter pylori–related gastric low-grade MALT lymphoma. Gastroenterology. 1999;117(5):1105–1112. doi: 10.1016/S0016-5085(99)70395-1. [DOI] [PubMed] [Google Scholar]
- 7.Eaton KA, Mefford M, Thevenot T. The role of T cell subsets and cytokines in the pathogenesis of Helicobacter pylori gastritis in mice. J Immunol. 2001;166(12):7456–7461. doi: 10.4049/jimmunol.166.12.7456. [DOI] [PubMed] [Google Scholar]
- 8.Pellicanò A, Sebkova L, Monteleone G, Guarnieri G, Imeneo M, Pallone F, Luzza F. Interleukin-12 drives the Th1 signaling pathway in Helicobacter pylori-infected human gastric mucosa. Infect Immun. 2007;75(4):1738–1744. doi: 10.1128/IAI.01446-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Smythies LE, Waites KB, Lindsey JR, Harris PR, Ghiara P, Smith PD. Helicobacter pylori-induced mucosal inflammation is Th1 mediated and exacerbated in IL-4, but not IFN-gamma, gene-deficient mice. J Immunol. 2000;165(2):1022–1029. doi: 10.4049/jimmunol.165.2.1022. [DOI] [PubMed] [Google Scholar]
- 10.Garhart CA, Nedrud JG, Heinzel FP, Sigmund NE, Czinn SJ. Vaccine-induced protection against Helicobacter pylori in mice lacking both antibodies and interleukin-4. Infect Immun. 2003;71(6):3628–3633. doi: 10.1128/IAI.71.6.3628-3633.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Lucas B, Bumann D, Walduck A, Koesling J, Develioglu L, Meyer TF, Aebischer T. Adoptive transfer of CD4 + T Cells specific for subunit A of Helicobacter pylori urease reduces H. pylori stomach colonization in mice in the absence of interleukin-4 (IL-4)/IL-13 receptor signaling. Infect Immun. 2001;69(3):1714–1721. doi: 10.1128/IAI.69.3.1714-1721.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Saldinger PF, Porta N, Launois P, Louis JA, Waanders GA, Bouzourène H, Michetti P, Blum AL, Corthésy-Theulaz IE. Immunization of BALB/c mice with Helicobacter urease B induces a T helper 2 response absent in Helicobacter infection. Gastroenterology. 1998;115(4):891–897. doi: 10.1016/S0016-5085(98)70261-6. [DOI] [PubMed] [Google Scholar]
- 13.Sayi A, Kohler E, Hitzler I, Arnold I, Schwendener R, Rehrauer H, Müller A. The CD4 + T cell-mediated IFN-γ response to Helicobacter infection is essential for clearance and determines gastric cancer risk. J Immunol. 2009;182(11):7085–7101. doi: 10.4049/jimmunol.0803293. [DOI] [PubMed] [Google Scholar]
- 14.Berg DJ, Lynch NA, Lynch RG, Lauricella DM. Rapid development of severe hyperplastic gastritis with gastric epithelial dedifferentiation in Helicobacter felis-infected IL-10−/− mice. Am J Pathol. 1998;152(5):1377–1386. [PMC free article] [PubMed] [Google Scholar]
- 15.Cai X, Carlson J, Stoicov C, Li H, Wang TC, Houghton J. Helicobacter felis eradication restores normal architecture and inhibits gastric cancer progression in C57BL/6 mice. Gastroenterol. 2005;128(7):1937–1952. doi: 10.1053/j.gastro.2005.02.066. [DOI] [PubMed] [Google Scholar]
- 16.Enno A, O’Rourke JL, Howlett CR, Jack A, Dixon MF, Lee A. MALToma-like lesions in the murine gastric mucosa after long-term infection with Helicobacter felis. A mouse model of Helicobacter pylori-induced gastric lymphoma. Am J Pathol. 1995;147(1):217–222. [PMC free article] [PubMed] [Google Scholar]
- 17.Lee JY, Kim N, Choi YJ, Nam RH, Choi YJ, Kwon YH, Yoon K, Suh JH, Lee SM, Lee HS, et al. Histologic findings and inflammatory reactions after long-term colonization of Helicobacter felis in C57BL/6 mice. J Cancer Prev. 2014;19(3):224–230. doi: 10.15430/JCP.2014.19.3.224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Roth KA, Kapadia SB, Martin SM, Lorenz RG. Cellular immune responses are essential for the development of Helicobacter felis-associated gastric pathology. J Immunol. 1999;163(3):1490–1497. [PubMed] [Google Scholar]
- 19.Lee A, O’Rourke J, De Ungria MC, Robertson B, Daskalopoulos G, Dixon MF. A standardized mouse model of Helicobacter pylori infection: introducing the Sydney strain. Gastroenterology. 1997;112(4):1386–1397. doi: 10.1016/S0016-5085(97)70155-0. [DOI] [PubMed] [Google Scholar]
- 20.Thompson LJ, Danon SJ, Wilson JE, O’Rourke JL, Salama NR, Falkow S, Mitchell H, Lee A. Chronic Helicobacter pylori infection with Sydney strain 1 and a newly identified mouse-adapted strain (Sydney strain 2000) in C57BL/6 and BALB/c mice. Infect Immun. 2004;72(8):4668–4679. doi: 10.1128/IAI.72.8.4668-4679.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ferrero RL, Ave P, Radcliff FJ, Labigne A, Huerre MR. Outbred mice with long-term Helicobacter felis infection develop both gastric lymphoid tissue and glandular hyperplastic lesions. J Pathol. 2000;191(3):333–340. doi: 10.1002/1096-9896(2000)9999:9999<::AID-PATH619>3.0.CO;2-H. [DOI] [PubMed] [Google Scholar]
- 22.Wang X, Willen R, Svensson M, Ljungh A, Wadstrom T. Two-year follow-up of Helicobacter pylori infection in C57BL/6 and Balb/cA mice. APMIS. Acta Pathol Microbiol et Immunol Scand. 2003;111(4):514–522. doi: 10.1034/j.1600-0463.2003.1110410.x. [DOI] [PubMed] [Google Scholar]
- 23.Kamradt AE, Greiner M, Ghiara P, Kaufmann SH. Helicobacter pylori infection in wild-type and cytokine-deficient C57BL/6 and BALB/c mouse mutants. Microbes Infect. 2000;2(6):593–597. doi: 10.1016/S1286-4579(00)00367-1. [DOI] [PubMed] [Google Scholar]
- 24.Sakagami T, Dixon M, O’Rourke J, Howlett R, Alderuccio F, Vella J, Shimoyama T, Lee A. Atrophic gastric changes in both Helicobacter felis and Helicobacter pylori infected mice are host dependent and separate from antral gastritis. Gut. 1996;39(5):639–648. doi: 10.1136/gut.39.5.639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Enno A, O’Rourke J, Braye S, Howlett R, Lee A. Antigen-dependent progression of mucosa-associated lymphoid tissue (MALT)-type lymphoma in the stomach. Effects of antimicrobial therapy on gastric MALT lymphoma in mice. Am J Pathol. 1998;152(6):1625–1632. [PMC free article] [PubMed] [Google Scholar]
- 26.Mohammadi M, Nedrud J, Redline R, Lycke N, Czinn SJ. Murine CD4 T-cell response to Helicobacter infection: tH1 cells enhance gastritis and TH2 cells reduce bacterial load. Gastroenterology. 1997;113(6):1848–1857. doi: 10.1016/S0016-5085(97)70004-0. [DOI] [PubMed] [Google Scholar]
- 27.Ferrero RL, Thiberge J-M, Labigne A. Local immunoglobulin G antibodies in the stomach may contribute to immunity against Helicobacter infection in mice. Gastroenterology. 1997;113:185–194. doi: 10.1016/S0016-5085(97)70094-5. [DOI] [PubMed] [Google Scholar]
- 28.Ferrero RL, Ave P, Ndiaye D, Bambou JC, Huerre MR, Philpott DJ, Memet S. NF-kappaB activation during acute Helicobacter pylori infection in mice. Infect Immun. 2008;76(2):551–561. doi: 10.1128/IAI.01107-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Viala J, Chaput C, Boneca IG, Cardona A, Girardin SE, Moran AP, Athman R, Mémet S, Huerre MR, Coyle AJ, et al. Nod1 responds to peptidoglycan delivered by the Helicobacter pylori cag pathogenicity island. Nat Immunol. 2004;51:166–174. doi: 10.1038/ni1131. [DOI] [PubMed] [Google Scholar]
- 30.Sutton P, Wilson J, Genta R, Torrey D, Savinainen A, Pappo J, Lee A. A genetic basis for atrophy: dominant non-responsiveness and Helicobacter induced gastritis in F(1) hybrid mice. Gut. 1999;45(3):335–340. doi: 10.1136/gut.45.3.335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Huang S, Hendriks W, Althage A, Hemmi S, Bluethmann H, Kamijo R, Vilcek J, Zinkernagel RM, Aguet M. Immune response in mice that lack the interferon-gamma receptor. Science. 1993;259(5102):1742–1745. doi: 10.1126/science.8456301. [DOI] [PubMed] [Google Scholar]
- 32.Collins JT, Dunnick WA. Germline transcripts of the murine immunoglobulin gamma 2a gene: structure and induction by IFN-gamma. Int Immunol. 1993;5(8):885–891. doi: 10.1093/intimm/5.8.885. [DOI] [PubMed] [Google Scholar]
- 33.Snapper CM, Paul WE. Interferon-gamma and B cell stimulatory factor-1 reciprocally regulate Ig isotype production. Science. 1987;236(4804):944–947. doi: 10.1126/science.3107127. [DOI] [PubMed] [Google Scholar]
- 34.Finkelman FD, Katona IM, Mosmann TR, Coffman RL. IFN-gamma regulates the isotypes of Ig secreted during in vivo humoral immune responses. J Immunol. 1988;140(4):1022–1027. [PubMed] [Google Scholar]
- 35.Uchida K, Okazaki K, Debrecceni A, Nishi T, Iwano H, Inai M, Uose S, Nakase H, Ohana M, Oshima C, et al. Analysis of cytokines in the early development of gastric secondary lymphoid follicles in Helicobacter pylori-infected BALB/c mice with neonatal thymectomy. Infect Immun. 2001;69(11):6749–6754. doi: 10.1128/IAI.69.11.6749-6754.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Gray BM, Fontaine CA, Poe SA, Eaton KA. Complex T cell interactions contribute to Helicobacter pylori gastritis in mice. Infect Immun. 2013;81(3):740–752. doi: 10.1128/IAI.01269-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Yang L, Yamamoto K, Nishiumi S, Nakamura M, Matsui H, Takahashi S, Dohi T, Okada T, Kakimoto K, Hoshi N, et al. Interferon-gamma-producing B cells induce the formation of gastric lymphoid follicles after Helicobacter suis infection. Muc Immunol. 2015;8(2):279–295. doi: 10.1038/mi.2014.66. [DOI] [PubMed] [Google Scholar]
- 38.Rad R, Dossumbekova A, Neu B, Lang R, Bauer S, Saur D, Gerhard M, Prinz C. Cytokine gene polymorphisms influence mucosal cytokine expression, gastric inflammation, and host specific colonisation during Helicobacter pylori infection. Gut. 2004;53(8):1082–1089. doi: 10.1136/gut.2003.029736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Wang SK, Zhu HF, He BS, Zhang ZY, Chen ZT, Wang ZZ, Wu GL. CagA + H. pylori infection is associated with polarization of T helper cell immune responses in gastric carcinogenesis. World J Gastroenterol. 2007;13(21):2923–2931. doi: 10.3748/wjg.v13.i21.2923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Riedel S, Kraft M, Kucharzik T, Pauels HG, Tiemann M, Steinbuchel A, Domschke W, Lugering N. CD4 + Th1-cells predominate in low-grade B-cell lymphoma of gastric mucosa-associated lymphoid tissue (MALT type) Scand J Gastroenterol. 2001;36(11):1198–1203. doi: 10.1080/00365520152584842. [DOI] [PubMed] [Google Scholar]
- 41.Shi Y, Liu XF, Zhuang Y, Zhang JY, Liu T, Yin Z, Wu C, Mao XH, Jia KR, Wang FJ, et al. Helicobacter pylori-induced Th17 responses modulate Th1 cell responses, benefit bacterial growth, and contribute to pathology in mice. J Immunol. 2010;184(9):5121–5129. doi: 10.4049/jimmunol.0901115. [DOI] [PubMed] [Google Scholar]
- 42.Arnold IC, Hitzler I, Engler D, Oertli M, Agger EM, Muller A. The C-terminally encoded, MHC class II-restricted T cell antigenicity of the Helicobacter pylori virulence factor CagA promotes gastric preneoplasia. J Immunol. 2011;186(11):6165–6172. doi: 10.4049/jimmunol.1003472. [DOI] [PubMed] [Google Scholar]
- 43.van Doorn NE, Namavar F, Sparrius M, Stoof J, van Rees EP, van Doorn LJ, Vandenbroucke-Grauls CM. Helicobacter pylori-associated gastritis in mice is host and strain specific. Infect Immun. 1999;67(6):3040–3046. doi: 10.1128/iai.67.6.3040-3046.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Liu S, Luo J, Liu Y, Tang S, Chen C, Cai H, Yu M, Zhang Y. CD4(+) Foxp3(+) regulatory T-cell number increases in the gastric tissue of C57BL/6 mice infected with Helicobacter pylori. APMIS Acta Pathol Microbiol et Immunol Scand. 2015;123(7):571–579. doi: 10.1111/apm.12388. [DOI] [PubMed] [Google Scholar]
- 45.Strober W, Kelsall B, Fuss I, Marth T, Ludviksson B, Ehrhardt R, Neurath M. Reciprocal IFN-gamma and TGF-beta responses regulate the occurrence of mucosal inflammation. Immunol Today. 1997;18(2):61–64. doi: 10.1016/S0167-5699(97)01000-1. [DOI] [PubMed] [Google Scholar]
- 46.Dhiman N, Ovsyannikova IG, Ryan JE, Jacobson RM, Vierkant RA, Pankratz VS, Jacobsen SJ, Poland GA. Correlations among measles virus-specific antibody, lymphoproliferation and Th1/Th2 cytokine responses following measles-mumps-rubella-II (MMR-II) vaccination. Clin Exp Immunol. 2005;142(3):498–504. doi: 10.1111/j.1365-2249.2005.02931.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Bergman MP, D’Elios MM. Cytotoxic T cells in H. pylori-related gastric autoimmunity and gastric lymphoma. J Biomed Biotechnol. 2010;2010:104–918. doi: 10.1155/2010/104918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Craig VJ, Cogliatti SB, Arnold I, Gerke C, Balandat JE, Wundisch T, Muller A. B-cell receptor signaling and CD40 ligand-independent T cell help cooperate in Helicobacter-induced MALT lymphomagenesis. Leukemia. 2010;24(6):1186–1196. doi: 10.1038/leu.2010.76. [DOI] [PubMed] [Google Scholar]
- 49.Hussell T, Isaacson PG, Crabtree JE, Spencer J. Helicobacter pylori-specific tumour-infiltrating T cells provide contact dependent help for the growth of malignant B cells in low-grade gastric lymphoma of mucosa-associated lymphoid tissue. J Pathol. 1996;178(2):122–127. doi: 10.1002/(SICI)1096-9896(199602)178:2<122::AID-PATH486>3.0.CO;2-D. [DOI] [PubMed] [Google Scholar]
- 50.O’Rourke JL, Dixon MF, Jack A, Enno A, Lee A. Gastric B-cell mucosa-associated lymphoid tissue (MALT) lymphoma in an animal model of ‘Helicobacter heilmannii’ infection. J Pathol. 2004;203(4):896–903. doi: 10.1002/path.1593. [DOI] [PubMed] [Google Scholar]
- 51.Hauer AC, Finn TM, MacDonald TT, Spencer J, Isaacson PG. Analysis of TH1 and TH2 cytokine production in low grade B cell gastric MALT-type lymphomas stimulated in vitro with Helicobacter pylori. J Clin Pathol. 1997;50(11):957–959. doi: 10.1136/jcp.50.11.957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Mueller A, O’Rourke J, Chu P, Chu A, Dixon MF, Bouley DM, Lee A, Falkow S. The role of antigenic drive and tumor-Infiltrating accessory cells in the pathogenesis of Helicobacter-induced mucosa-associated lymphoid tissue lymphoma. Am J Pathol. 2005;167(3):797–812. doi: 10.1016/S0002-9440(10)62052-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Greiner A, Knorr C, Qin Y, Sebald W, Schimpl A, Banchereau J, Muller-Hermelink HK. Low-grade B cell lymphomas of mucosa-associated lymphoid tissue (MALT-type) require CD40-mediated signaling and Th2-type cytokines for in vitro growth and differentiation. Am J Pathol. 1997;150(5):1583–1593. [PMC free article] [PubMed] [Google Scholar]
- 54.Knorr C, Amrehn C, Seeberger H, Rosenwald A, Stilgenbauer S, Ott G, Muller Hermelink HK, Greiner A. Expression of costimulatory molecules in low-grade mucosa-associated lymphoid tissue-type lymphomas in vivo. Am J Pathol. 1999;155(6):2019–2027. doi: 10.1016/S0002-9440(10)65521-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Sellers RS, Clifford CB, Treuting PM, Brayton C. Immunological variation between inbred laboratory mouse strains: points to consider in phenotyping genetically immunomodified mice. Vet Pathol. 2012;49(1):32–43. doi: 10.1177/0300985811429314. [DOI] [PubMed] [Google Scholar]