

Abstract
Background
Epidermal growth factor receptor (Egfr) has been the target of several drugs for cancers. The potential gender differences in genes in the Egfr axis have been suggested in humans and in animal models. Female and male mice from the same recombinant inbred (RI) strain have the same genomic components except the sex difference. A population of different RI mouse strains allows to conduct precise analysis of molecular pathways and regulation of Egfr between female and male mice.
Methods
The whole genome expression profiles of 70 genetically diverse RI strains of mice were used to compare three major molecular aspects of Egfr gene: the relative expression levels, gene network and expression quantitative trait loci (eQTL) that regulate the expression of Egfr between female and male mice.
Results
Our data showed that there is a significant sex difference in the expression levels in kidney. A considerable number of genes in the gene network of Egfr are sex differentially expressed. The expression levels of Egfr in mice are statistical significant different between C57BL/6 J (B6) and DBA/2 J (D2) genotypes in male while no difference in female mice. The eQTLs that regulate the expression levels of Egfr between female and male mice are also different. Furthermore, the differential expression levels of Egfr showed significantly different correlations with two known biological traits between male and female mice.
Conclusion
Overall there is a substantial sex difference in the Egfr pathways in mice. These data may have significant impact on drug target design, development, formulation, and dosage determinant for women and men in clinical trials.
Electronic supplementary material
The online version of this article (doi:10.1186/s12863-016-0449-3) contains supplementary material, which is available to authorized users.
Keywords: Drug, Egfr; Mice, Pathway, Sex
Background
The purpose of this work is to systematically investigate the sex differences of epidermal growth factor receptor (Egfr) in the kidney using high quality data from a population of mouse recombinant inbred (RI) strains. Sex differences have been brought to the attention of the public and health research community [1–3], and sex disparities in health throughout the lifespan of humans or mice have been documented [2, 3].
In humans, EGFR has been reported as an important player in pathways of kidney diseases. Liang et al. reported the inhibitory effect of silibinin on EGFR signal-induced renal cell carcinoma progression via suppression of the EGFR/MMP-9 signaling pathway [4]. The effect of EGR on renal cell carcinoma was also reported by others [5, 6]. EGFR activation is required to induce the renal fibrotic genes [7, 8]. EGFR has been received great attention in cancer research because of its connection to cancer development [9, 10]. Drugs targeting the EGFR pathway have been developed in recent years, and some clinic trials are underway. For example, Erlotinib, an EGFR and tyrosine kinase inhibitor, have been used for the treatment of kidney, lung, advanced adenocarcinoma of the oesophagus and gastro-oesophageal junction and hepatocellular carcinoma cancers [11, 12]. Lapatinib ditosylate (LAP), an anti-EGFR drug, has been used for treatment of gastro-oesophageal cancer and renal cancer [13, 14]. Ramucirumab has been used as monotherapy for previously treated advanced gastric or gastro-oesophageal junction adenocarcinoma [15–18]. While the drugs based on anti EGFR are in development and clinical trial stage, its gender specificity should be thoroughly investigated.
The potential gender differences in genes in the Egfr axis have been suggested in humans and in animal models [19–27]; however, the gender specificity on the molecular pathways has yet to be understood. At present, a systematic investigation of sex specificities in the Egfr axis, either in humans or animal models, has not been reported. Recently, substantial progress has been made in elucidating how diverse sex specific systems are integrated into developmental gene networks using animal models. In particular, the RI strains derived from C57BL/6 J (B6) X DBA/2 J (D2) have been used for studies of sex specific traits and genetic regulations [20–22]. Sex specific gene expressions profiling have been analyzed for the liver and other tissues [23–25]. Therefore, we believe that our optimized approach discloses information on sex differences in the Egfr axis that will eventually impact the management of all drug design and clinical trials in many diseases including cancer.
Several recent publications showed the sex specificity [20, 21, 24, 26] and tissue specificity [25, 27, 28] of gene expression levels or the association of gene expressions. Recent study indicated that the relative expression levels, gene network and eQTLs that regulate the expression of Egfr in liver are different between female and male mice [29]. We hypothesize that at least one step in the Egfr molecular pathway in kidney of the male is different from that of the female mice. We first investigated whether the expression levels of Egfr in the kidney is different between female and male mice. We then determined whether the gene network of Egfr between female and male mice are the same. We finally examined the genetic loci that regulate the expression of Egfr in females and males.
Methods
Expression levels of Egfr
For the expression data of Egfr and its associated genes, we collected the expression data of Egfr axis from whole genome expression data of the mouse kidney [29]. All data are from GeneNetwork (http://www.genenetwork.org/webqtl/main.py) and are available to the public. We used the Actin beta (B) as controls for the expression level of Egfr. The data set of gene expression profiles of mouse kidney at GeneNetwork were validated using sex specific probe sets such as X inactivation-specific transcript (Xist) and Dead/h box 3, y-linked (Dby) [29]. When multiple probes are presented for a gene, the probe with the highest expression level was chosen for the analysis while the others were used as references.
Data set for analysis of gene expression profiles
In this study, we used the Mouse Kidney M430v2 female and male (Aug06) RMA Database from the GeneNetwork (http://www.genenetwork.org/webqtl/main.py). RMA stands for statistical method of robust multi-array average for the raw data analysis [21] The data set includes mRNA expression in the adult kidney of both sexes of 70 genetically diverse strains of mice including 54 BXD RI strains, a set of 15 inbred strains, and 1 F1 hybrid: D2B6F1. Kidney samples were processed using a total of 153 Affymetrix Mouse Expression 430 2.0 arrays. Kidneys from two to six animals per strain were pooled. Forty-two (31 BXD, D2B6F1 and 10 inbred strains) are represented by male and female samples [21].
While samples of both sexes are collected at similar age, performed with same microarray platform, and grown in the same environment, most of them are from different RI strains. However, these RI strains are all derived from the same two progenitors. Also, the gene expression profiles of both the female and male progenitors are generated separately.
The association of the expression levels of Egfr between female and male mouse populations
The association analysis was conducted using the method as previously reported [23, 29, 30]. R values were compared between female and male mice, following standard criteria for the strong, correlation, and none correlation [29]. Unless noted in the figure legends, when the R value was equal or more than 0.7 or −0.7, the correlation was regarded as strong positive or negative. When the R value was between 0.50 and 0.69 or −0.50 and −0.69, the correlation existed but was not strong. When the R value was between 0.3 and 0.49 or −0.30 and −0.49, the correlation was weak. Any R value between 0 and 0.29 or 0 and −0.29 was regarded as none-correlation [29, 30].
Gene network construction
The gene networks were constructed using application tools in GeneNetwork. We constructed the gene network based on the Network Graph in combination with the Correlation Matrix [29]. For the Correlation Matrix, the Pearson product-moment correlations (the standard type of correlation) were used for the calculation of correlations [30, 31]. For each sex in each pair of samples, both the Network Graph and Correlation Matrix were obtained with the same set of parameters or criteria. For example, for the Line Threshold in the Network Graph, absolute values greater than 0.30 were used across all samples. The Spring Model layout (force reduction) was used for the graphic method for all graphic samples.
Transcriptomic loci (eQTL) that regulates the expression level of Egfr in female and male mice
Transcriptome mapping with GeneNetwork was used to identify the chromosomal regions that affect the expression of Egfr in female and male mice, which includes three major steps. First, Egfr probes of gene expression were identified from female and male strains of tissues. Second, interval mapping was done to establish Egfr transcriptome maps for the entire genome. Permutations of 5000 tests were used to assess the strength and consistency of the linkages. Third, genomic regions and locations on chromosomes were compared [30–32].
Results
Expression levels of Egfr between female and male in mouse kidney
Basical statistical analysis was conducted with data of gene expression of Actin B and Egfr in both sexes from a total of 43 strains [29]. Five probes for Actin B on the Affymetrix Mouse Genome 430 2.0 array chip was identified. The Probe of 1436722_a_at was chosen for the analysis because its high expression level and it contains exon 3, 4, 5 and proximal 3′ UTR. Figure 1a and b showed the expression levels of Actin A in female and male mice of different strains. The P value from the T test between female and male is 0.55 and the R value from correlation analysis is 0.25.
Fig. 1.
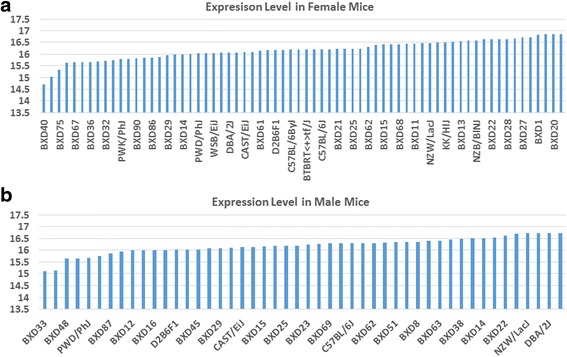
Expression levels of Actin B in kidney between female and male mice in BXD strains. Numbers on left vertical bar are for the relative levels of expression of Actin B. The information of strains and names of standard inbred strains are listed under the horizontal bar. a. The expression levels of Actin B in kidney of female mice. b. The expression levels of Actin B in kidney of male mice
Seven probes for Egfr were found from data from the Affymetrix Mouse Genome 430 2.0 array chip. The Probe 1460420_a_at was chosen for the analysis because its high expression level. Figure 2a and b shows the expression levels of Egfr in female and male mice of different strains. For Egfr, the P value from T test between female and male mice is 0.00 and the R value is −0.05.
Fig. 2.
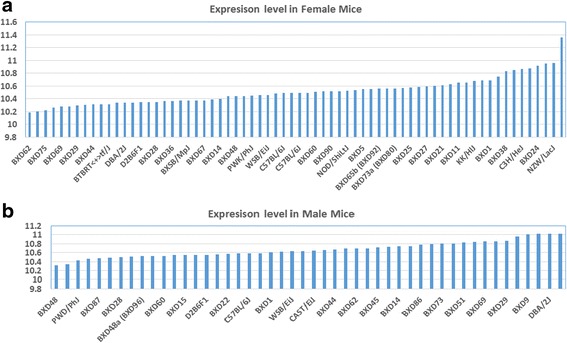
Expression levels of Egfr in kidney between female and male mice in BXD strains. Numbers on left vertical bar are for the relative levels of expression of Egfr. The information of strains and names of standard inbred strains are listed under the horizontal bar. a. The expression levels of Egfr in kidney of female mice. b. The expression levels of Egfr in kidney of male mice
Thus, there was no sex difference in the expression level of Actin B in kidney while there was a significant sex difference in the Egfr expression levels in these mouse strains.
Gene network of Egfr between female and male in mouse kidney
With Probe 1460420, the top 50 probes of genes with expression levels most correlated to that of Egfr were identified from the mouse data of Mouse kidney M430v2 Male (Aug06) RMA Database. These probes represent genes with diversity of biological functions (Additional file 1: Table S1).
As shown in Fig. 3a, Egfr is either positively or negatively correlated to the top 50 probes of genes. The expression of mouse Egfr is strongly positively correlated to ribosomal protein S4, X-linked (Rps4x) and zinc inger protein 261 (Zfp261); it is strongly negatively correlated to dedicator of cytokinesis 9 (Dock9). In order to compare the correlation of these genes to Egfr in female mice, we then constructed a gene network using the same probes from female gene expression profiles of the kidney (Fig. 3b).
Fig. 3.
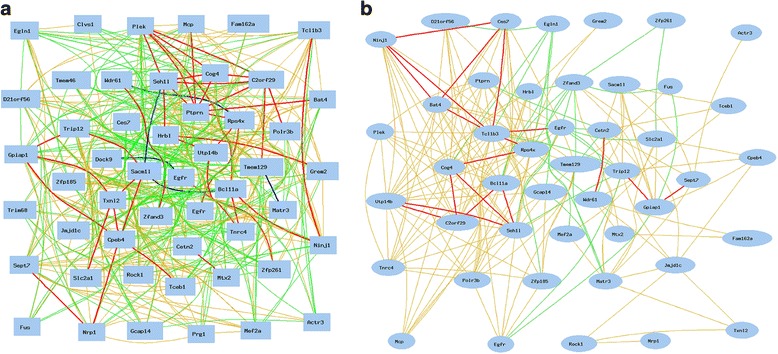
Gene network of Egfr in mouse Kidney. The 50 nodes in the graph below show the selected traits. Only nodes with edges are displayed. The 208 edges between the nodes, filtered from the 1225 total edges and drawn as curves, show Pearson correlation coefficients greater than 0.5 or less than −0.5. The graph’s canvas is 40.0 by 40.0 cm, and the node labels are drawn with a 16.0 point font, and the edge labels are drawn with a 16.0 point font. a. Gene network of top 50 genes that closely associated to Egfr in male mouse kidney. b. Gene network of male top 50 genes that closely associated to Egfr in female mouse kidney
Several differences in the gene network of the kidney were found between female and male mice. In males, Egfr showed a strong negative correlation with Dock9 but in females, such a negative correlation did not appear. In males, Egfr showed a negative correlation with clavesin 1 (retinaldehyde binding protein 1-like 1) (Clvs1); HIF1 alpha hypoxia growth and transformation-dependent protein, proapoptotic (Fam162a); neuropilin 1 (Nrp1); proteoglycan 1, secretory granule (Prg1); septic 7 (cell division cycle 10 homolog) (Sept7); transcription elongation factor B (SIII), polypeptide 1 (Tceb1); and transmembrane protein 46 (Tmem46). However, in female mice, these negative correlations did not show up. In addition, in male mice, Egfr showed a positive correlation with membrane cofactor protein (Mcp); pleckstrin associated EST AK008484 (Plek); and trinucleotide repeat containing 4 (Tnrc4), although this positive correlation did not show up in female mice.
Confirmation of sex difference in kidney with multiple probes
The above results are mainly based on Egfr probe 1460420. There were seven probes of Egfr on the Affymetrix Mouse Genome 430 2.0 array chip. In order to find out whether these sex differences are caused by the bias in one probe, we examined correlations between all the Egfr probes and these groups of sex different genes.
We first obtained the data of all seven Egfr probes and probes of all 11 genes that showed positive or negative correlations with Egfr from kidney of male strains, and then constructed the gene network for all of them. As shown in Fig. 4a, three positive regulated genes (Mcp, Plek, Tnrc4) were all being connected positively to at least one of the Egfr probes, while eight negatively regulated genes (Dock9, Clvs1, Fam162a, Nrp1, Prg1, Sept7, Tceb1, Tmem46) were negatively connected to at least one of the Egfr probes.
Fig. 4.
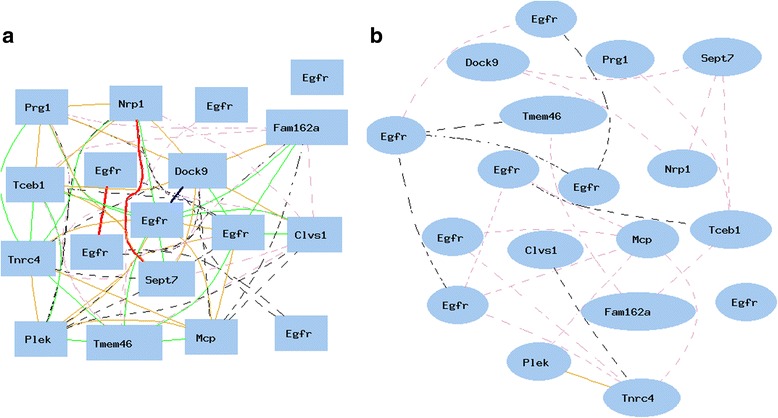
Confirmation of negative and positive correlations between Egfr and key genes showing sex difference. The 18 nodes in the graph below show the selected traits. The graph’s canvas is 40.0 by 40.0 cm, and the node labels are drawn with a 16.0 point font, and the edge labels are drawn with a 16.0 point font. Curves show Pearson correlation coefficients greater than 0.3 or less than −0.3. a. Correlations between Egfr and key genes in kidney of male mice. All nodes are displayed. The 73 edges between the nodes, filtered from the 153 total edges and drawn as curves. b. Correlations between Egfr and key genes in kidney of female mice. Lack of positive and negative correlations between Egfr probes and 11 key genes. All nodes are displayed. The 23 edges between the nodes, filtered from the 153 total edges and drawn as curves
We next obtained the data of all seven Egfr probes and probes of all above 11 genes from kidneys of female mouse strains. Figure 4b showed that there was no significant connection between these 11 genes to any probe of Egfr. The R values were all under 0.3, at a non-significant level. These data confirm the significant difference in the molecular network of Egfr between female and male mice.
EQTL regulation of Egfr between female and male mice
The eQTLs of all seven probes of Egfr in female and male mice were identififed. Figure 5 shows the eQTL mapped from each probe in female and in male mice. The eQTL of these seven probes of Egfr in kidneys of female mice were mapped on to chromosomes 1, 3, 9, 11, 12. 14, 15, 16 and 19. The probe, 1460420_a_at with the highest expression level, was mapped on chromosome 14. The other probe, 1451530_ at, mapped an eQTL on chromosome 15, with the highest LRS score of 15.6. We then found eQTLs for the regulation of Egfr from the male kidney from these seven probes. The eQTL from these probes were on chromosome 4, 9, 12, 14, 15, 17 and 19. The QTL with highest LRS value is located on chromosome 4, which is 14.2 (Fig. 5).
Fig. 5.
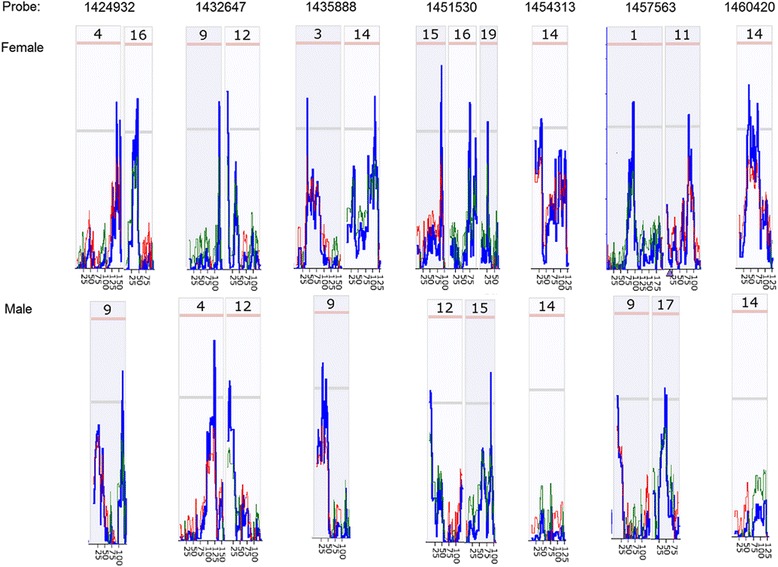
Detection of eQTL for regulation of Egfr expression levels in kidney of mouse RI strains using probes of Egfr. Top line is the numbers of Egfr probes, second line is the eQTL detected from female mice. Bottom line shows the eQTL detected from male mice. The numbers on top of each individual figure in second and bottom lines indicate the number of chromosome. Pink color lines on top of the individual figure indicate the threshold for significant level. Light grey lines indicate the threshold for suggestive level
Although there were overlaps on some chromosomes of eQTL between female and male mice, the locations of most of these eQTL are different between sexes. The eQTLs from three probes were all different from each other between female and male mice. Mapping with probe 1424932_at, eQTLs were detected from chromosome 4 and 16 from female, while the eQTL was located on chromosome 9 from the male. Mapping with probe 1435888_at, eQTLs were detected from chromosome 3 and 14 from female, while the eQTL was located on chromosome 9 from the male. Mapping with probe 1457563_at, eQTLs were detected from chromosome 1 and 11 from female, while the eQTLs were mapped on chromosome 9 and 17 from the male. The other two probes each detected one overlapped eQTL and other non-overlapped eQTLs between female and male. Mapping with probe 1432647_at, eQTLs were detected from chromosomes 9 and 12 from female, while the eQTLs were mapped on chromosomes 4 and 12 from the male. Mapping with probe 1451530_at, eQTLs were detected from chromosome 12, 15, and 19 from females, while the eQTL were mapped on chromosomes 12 and 15 from the males. The final two probes had eQTL from female but not from the male mice. Mapping with probes 1454313_at and 1460420_a_at an eQTL was detected on chromosome 14, for female for both probes, while no eQTL was mapped from the male mice. Overall, the difference is much larger than the similarity.
Potential candidate for the Egfr expression level in male mice on chromosome 4
According to the map based on probe #1432647, the peak region of the eQTL on chromosome 4 is between 120.6 Mb and 124 Mb (Fig. 6a). Within the region, 35 genetic elements, including 25 known genes, exist. We further examined the known SNP in this region among BXD strains. Within the region, we find 13 polymorphic markers between B6 and DBA/1. They are divided into 4 haploid groups. The first one is the SNP rs4224744, which is located on 120.476225. The second group include rs3675629 and gnf04.116.914, which located between 120.731447 Mb and 120.875373 Mb. The third group includes 7 markers, gnf04.117.102, CEL-4_120039566, UT_4_121.927901, gnf04.119.329, rs13477959, and rs3714811, which is located between 121.069747 Mb and 124.048406 Mb. The fourth group includes three markers, rs3677161, rs13474356 and rs3704486, which is located between 124.517595 Mb and 125.265631 Mb. We then compared the expression level of Egfr with B (B6) and D (DBA/1) genotypes in male mice of each group of haploid types. The P values of T-test for each of these four groups are 0.062186655, 0.035691806, 0.007229499, and 0.009439615, respectively (Fig. 6b). Thus, the candidate genes are most likely located within the region covered by the polymorphic markers in group 3. Within this region, there are 29 genetic elements, Including 18 known genes. These genes are Ppt1, Cap1, Mfsd2, Mycl1, Trit1, Bmp8b, Oxct2b, Ppie, Hpcal4, Nt5c1a, Heyl, Pabpc4, Bmp8a, Oxct2a, Macf1, Ndufs5, Rhbdl2, Mycbp and Rragc. We next examined the relationship of expression level between these candidate genes and that of Egfr. No expression level of these genes showed a strong correlation to that of Egfr. In order to examine whether this eQTL is male specific, we examined the Egfr expression of B and D genotypes in female. Our data showed that the P values between the B and D genotypes in different haploid groups varied from 0.869525556 to 0.978478759 (Fig. 6c). Thus, there is no difference in the Egfr expression level between the B and D genotypes in female mice. These data confirm the difference in the regulation of expression levels of Egfr between female and male mice.
Fig. 6.
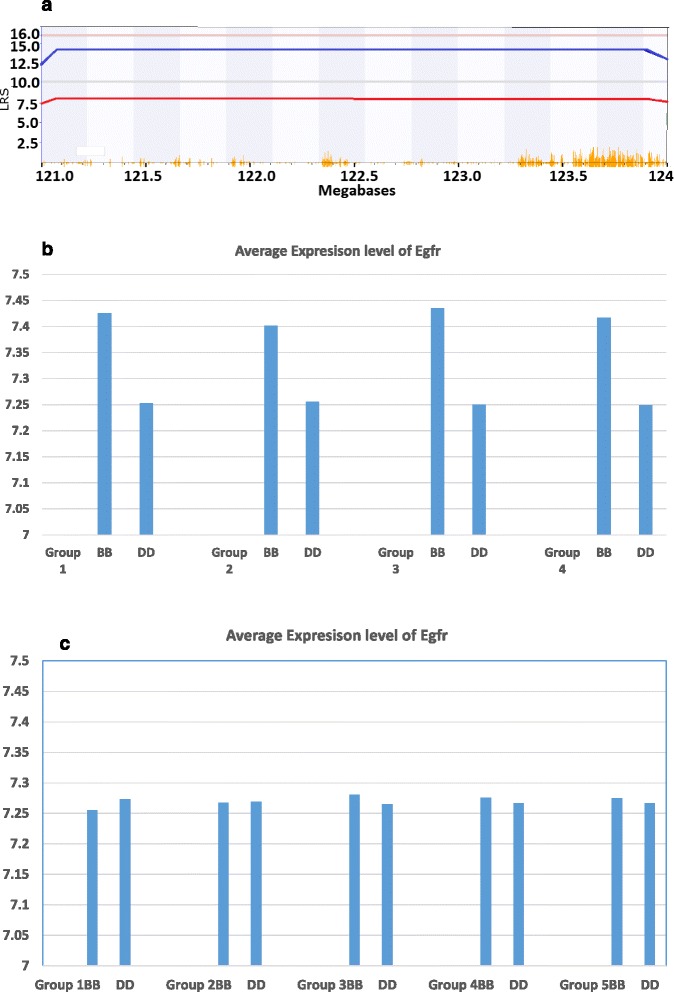
Comparison of level of Egfr expression of different genotypes in peak region of eQTL on chromosome 4. a. Location of peak region of eQTL of Egfr in male mice. Pink color lines on top of the individual figure indicate the threshold for significant level. Light grey lines indicate the threshold for suggestive level. Numbers on the Y bar are the LRS scores. Numbers on the X bar are the position on the chromosome measured by Mb. b. The average expression levels of Egfr in male mice with BB and DD genotypes. Numbers on the Y bar are the relative expression levels of Egfr. X bar indicates the groups of SNP and their genotypes. * P value is less than 0.05 but larger than 0.01. ** P values are less than 0.01. c. The average expression levels of Egfr in female mice with BB and DD genotypes. Numbers on the Y bar are the relative expression levels of Egfr. X bar indicates the groups of SNP and their genotypes
Potential impact of sex difference in Egfr expression on immunological traits
Because of these sex differences, we expect some biological traits may be affected. In order to estimate the potential impact of such a sex difference in the Egfr expression on the immune system, we examined the correlation between the Egfr expression level and some available traits in GeneNetwork. While a majority of the straits did not show a difference in terms of correlation with the expression level of Egfr in both sexes, some did show significant differences. Figure 6a and b shows the significantly different effect of expression level of Egfr between sexes on the level of interleukin-4 (Il4) from draining lymph node, 3 weeks post infection (GeneNework ID: 12711). In the male, the expression of Il4 is strongly positively correlated to that of Egfr, (Fig. 7a) in female, with a Rho value of 0.929 for Spearman rank correlation. However, in the female, they are negatively correlated (Fig. 7b) with a Rho value of −0.445. Figure 7c and d shows the significantly different effect of expression level of Egfr between sexes on the level of alpha-aminoadipate levels measured by LC-MS/MS in pooled plasma after overnight fasting in males at 29 weeks of age [33]. While in the male the expression of alpha-aminoadipate is strongly negatively correlated to that of Egfr with a Rho value of −0.721 (Fig. 7c), in female they are weakly positively correlated with a Rho value of 0.265 (Fig. 7d).
Fig. 7.
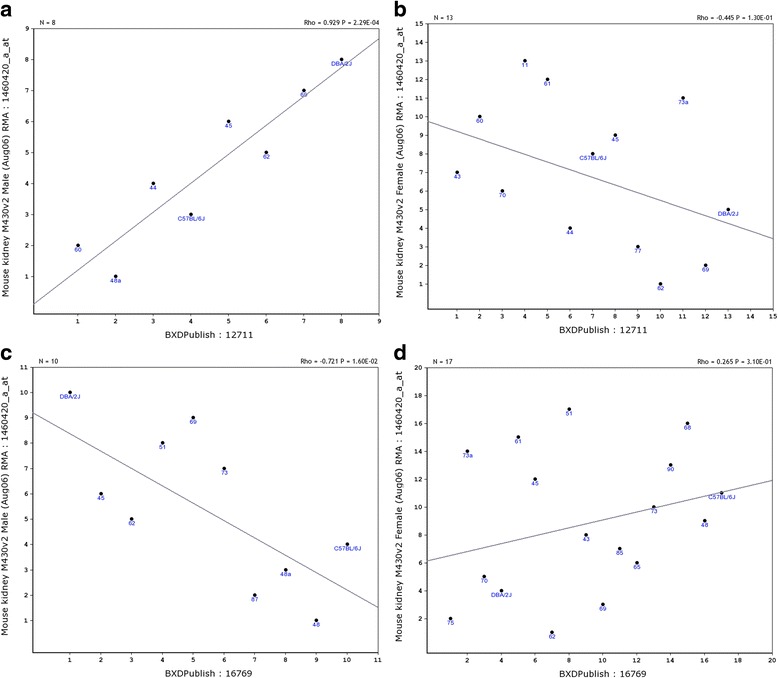
Correlation between the expression level of Egfr and known traits between male and female mice. a. Expression of Il-4 from draining lymph node in male mice is strongly positively correlated to that of Egfr. Numbers on the Y bar indicate the expression levels of Egfr. Numbers on the X bar indicate expression levels of Il-4. b. Expression of Il-4 in female and Egfr are weakly negatively correlated. Numbers on the Y bar indicate the expression levels of Egfr. Numbers on the X bar indicate expression levels of Il-4. c. In the male the expression of alpha-aminoadipate is negatively correlated to that of Egfr. Numbers on the Y bar indicate the expression levels of Egfr. Numbers on the X bar indicate expression levels of alpha-aminoadipate in male mice. d. In the female the expression of alpha-aminoadipate and that of Egfr are weakly positively correlated. Numbers on the Y bar indicate the expression levels of Egfr. Numbers on the X bar indicate expression levels of alpha-aminoadipate in female mice
Potential sex difference in Egfr ligands
As function of Egfr is regulated by the availability of its ligand, we further reexamined the expression levels of two Egfr ligands, the heparin-binding epidermal growth factor (hbEgf) and Epithelial Mitogen (Epgn). The eQTLs from two ligands were all different from each other between female and male mice. Figure 8 shows the eQTL detected from these two Egfr ligands. Mapping with the probe from Epgn, 3 eQTLs at suggestive levels were detected from chromosome 8, 13 and X from female, while no eQTL was detected from the male. Mapping with the probe from hbEgf, one eQTL at significant level was detected from chromosome 18 from female, while an eQTL at suggestive level on chromosome 2 was detected from the male. These data strengthen the importance of sex difference in drug design and treatment when targeting at Egfr.
Fig. 8.
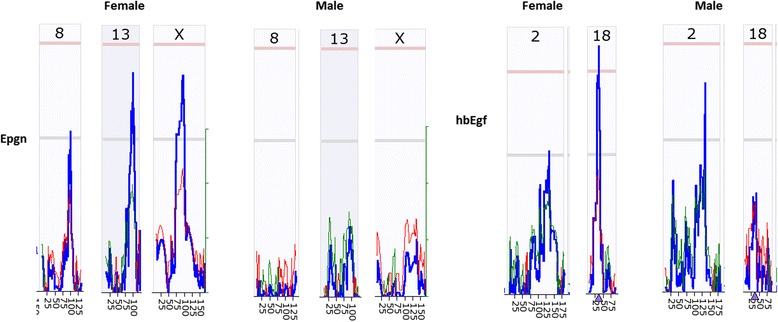
Detection of eQTL for regulation of Egfr ligands Epgn and hbEgf expression levels in kidney of mouse RI strains using probes of Egfr. The number on top of each individual figure indicates the number of chromosome. Pink color lines on top of the individual figure indicate the threshold for significant level. Light grey lines indicate the threshold for suggestive level. Numbers on the bottom indicate the Mb on the chromosomes
Discussion
Our data clearly demonstrated that there are sex differences in the molecular pathways of Egfr in the kidney. Gene expression profiles from each RI strain are generated from multiple homozygous mice [20–22, 29]. Female and male mice in the same strains are kept in the same animal facility with the same environment. The reliability of the data was demonstrated by similar expression levels of Actin B between female and male mice in these strains. The significant sex difference in the expression levels of Egfr in the kidney of these stains provided the solid foundation for this important study. Accordingly, the information on sex differences from these RI strains are important resources for further study.
Majority sex differentially regulated genes from this study have not been reported. However, a few of them have demonstrated their important roles in the pathways of Egfr. For example, NRP1 is widely expressed in cancer cells and in advanced human tumors. Most importantly, NRP1 is the co-receptor of EGFR [34]. It has been reported that NRP1 can control EGFR signaling and tumor growth [35]. The expression of Nrp1 is strongly negatively associated with that of Egfr in female mice but did not show a strong connection with Egfr in male mice. In such a case, it is important to know whether such a difference exists during tumor development in human kidney. Accordingly, these differences should be brought to attention for the development of drugs for cancers targeting Egfr in kidney diseases. It is also known that in humans VEGF signaling controls the Gonadotropin-releasing hormone (GnRH) neurons survival via NRP1 [36]. Hormones play an important role in the expression of VEGF as well as NRP1 [33–35]. The significant difference in correlation with two important traits to Egfr confirms that the sex difference may have a significant impact on a significant number of traits including the immune system. For the drug targeted Egfr pathway, attention should be paid to the gender differences of the drug response in the clinical trials. Dosages and methods of application of these drugs may need to be modified based on the gender.
It is known that in humans EGFR is a hormone related gene [27, 28]. It is reasonable to assume that its gender differential regulation to a certain degree hormone relevant. In addition to the Nrp1, we found that several other differentially expressed genes between female and male mice are also hormone related genes. For example, Prg1 is a progestin-responsive gene [37]. In humans, TCEB1 promotes invasion of prostate cancer cells and is involved in the development of hormone-refractory prostate cancer [38]. TSLP is involved in the regulation of estrogen on the secretion of MCP-1 and IL-8, and the growth of ESCs through JNK and NF-κB signal pathways [39, 40].
Interestingly, the eQTL that regulate the Egfr expression levels between female and male mice showed significant differences. Although several eQTL between mice are located on the same chromosomes in both sexes, these same eQTL are usually not being mapped with the same probes between sexes. For example, eQTL on chromosome 4 was mapped in females by probe 1424932_at while it was mapped in males by probe 1432647_at. EQTL on chromosome 9 was mapped in females by probe 1432647_at while it was mapped in males by probe 1424932_at, 1435888_at, and 1457563_at. Furthermore, eQTL was mapped on chromosome 14 by three probes 1435888_at, 1454313_at and 1460420_a_at in females but not by any probe in male mice. This complicated regulation of Egfr between females and males suggest that it is very likely that various mechanisms such as different splicing sites, multiple promote sites as well as differences in binding sites may involve in the regulation of Egfr. Further studies are necessary to clarify their mechanisms.
Similarly, our data also showed that potentially the expression levels of many Egfr ligands and/or relevant genes may also have sex-related differences. We demonstrated that different eQTL regulate the sex difference of hbEgf and Epgn. We believe that the expression of many Egfr ligands may be regulated differently in females and males. The sex differential expression may directly or indirectly affect the disease incidents and response to drug treatment. For example, hbEGF involves in kidney diseases such as glomerulonephritis and diabetic kidney disease [41, 42]. The interaction between Egfr and hbEgf in females and males may play an essential role on the effect of hbEgf on the diseases. Overexpression of Epgn during embryonic development induces reversible, epidermal growth factor receptor-dependent sebaceous gland hyperplasia [43]. We also notice that although Egfr is ubiquitously expressed [44, 45], its function is regulated by its ligands which are highly spatially restricted. Therefore further analysis of the sex differences of expression of Egfr ligands is necessary.
Similar to our previous study, this study is that our analysis is at one time point [29]. The differentially expressed genes between female and male mice may vary at different life stages. Future studies at different ages may lead to a comprehensive understanding of the sex difference in the molecular pathways of the Egfr axis in kidney. Similar to many studies, our data set is from the RI strains that were derived from two mouse strains, C57BL/6 J and DBA/2 J. Mouse strain specific pathways have been known [46, 47]. The results may represent sex difference under certain genomic backgrounds. Studies using other strains may reveal different mechanisms..
Like all other studies using animal models, the sex differences from this study may or may not represent that in humans [48]. Thus, not all the sex differentially expressed genes in mouse kidney are differentially expressed between males and females in human kidney. However, this study and our other study [29] strongly suggest that a gender difference in the EGFR pathway in humans exists.
We understand that, although the RNA expression levels are matched to the protein levels in a majority of genes, in some cases, the gene expression level does not necessarily represent the protein level. As to Egfr, a considerable number of publications have indicated the consistency between the RNA expression level and protein level [49–51]. However, a relatively large number of genes including its ligands are in the pathways in regulation of Egfr. It is important to examine whether their RNA expression levels agree with their protein levels in the future. On the other hand, the disagreement between protein and RNA levels does not necessarily invalidate the importance of the gene expression. We now know that RNA itself plays an important role in many biological functions including regulation of the expression of genes.
Conclusion
Our study reveals that there are differences in the molecular pathways of Egfr in the kidney between female and male mice. Many of sex differential expressed genes are related to hormone metabolism. These data reinforce the importance of being aware of the gender difference in drug development targeting Egfr and its ligands and their pathways.
Acknowledgements
Authors thank Drs. Robert Williams, Lu Lu, and Lei Yan for suppling data at GeneNetowrk and assistance on the data analysis.
Funding
This work was partially supported by the Center of Genomics and Bioinformatics and Center of Connective Tissue Research at the University of Tennessee Health Science Center; The department of technology in Hebei province, PR China (to QL); the Department of Veterans Affairs (BX000671 to WG), USA.
Availability of data and materials
All raw collected data which used in this manuscript are freely available from http://www.genenetwork.org/webqtl/main.py.
Authors’ contributions
WG, QL, and Yan J. designed experiments. FL, FG and WG analyzed data, constructed the gene networks, and mapped the eQTLs. QL and WG conceived and managed the project. FL, Yan J, Yun J. FG and WG contributed to manuscript preparation. All authors contribute to manuscript finalization. All authors have read and approved the final version of the manuscript.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Ethics approval
The data of moue tissues for this study from GeneNetwork (http://www.genenetwork.org/webqtl/main.py). Experimental animal procedures and mouse husbandry were performed in accordance with the National Institutes of Health’s Guide for the Care and Use of Laboratory Animals and approved by the UTHSC Institutional Animal Care and Use Committee.
Abbreviations
- EGFR
Epidermal growth factor receptor
- eQTL
Expression quantitative trait loci
- LAP
Lapatinib ditosylate
- RI
Recombinant inbred
Additional file
Correlation matrix of top 50 genes that are close correlated to Egfr in male mice. (DOCX 82 kb)
Contributor Information
Fengxia Liu, Email: lfx135246@sina.com.
Yan Jiao, Email: Yjiao2@uthsc.edu.
Yun Jiao, Email: Yun.Jiao@stjude.org.
Franklin Garcia-Godoy, Email: godoy@uthsc.edu.
Weikuan Gu, Email: wgu@uthsc.edu.
Qingyi Liu, Phone: 1-901-448-2259, Email: zhongmeijian-lqy@163.com.
References
- 1.Clayton JA, Collins FS. Policy: NIH to balance sex in cell and animal studies. Nature. 2014;509(7500):282–3. doi: 10.1038/509282a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Institute of Medicine (US) Committee on Understanding the Biology of Sex and Gender Differences. Wizemann TM, Pardue ML. Exploring the biological contributions to human health: does Sex matter? Washington (DC): National Academies Press (US); 2001. [PubMed] [Google Scholar]
- 3.Williams TM, Carroll SB. Genetic and molecular insights into the development and evolution of sexual dimorphism. Nat Rev Genet. 2009;10(11):797–804. doi: 10.1038/nrg2687. [DOI] [PubMed] [Google Scholar]
- 4.Liang L, Li L, Zeng J, Gao Y, Chen YL, Wang ZQ, Wang XY, Chang LS, He D. Inhibitory effect of silibinin on EGFR signal-induced renal cell carcinoma progression via suppression of the EGFR/MMP-9 signaling pathway. Oncol Rep. 2012;28(3):999–1005. doi: 10.3892/or.2012.1874. [DOI] [PubMed] [Google Scholar]
- 5.Thomasson M, Hedman H, Ljungberg B, Henriksson R. Gene expression pattern of the epidermal growth factor receptor family and LRIG1 in renal cell carcinoma. BMC Res Notes. 2012;5:216. doi: 10.1186/1756-0500-5-216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Dorđević G, Matušan Ilijaš K, Hadžisejdić I, Maričić A, Grahovac B, Jonjić N. EGFR protein overexpression correlates with chromosome 7 polysomy and poor prognostic parameters in clear cell renal cell carcinoma. J Biomed Sci. 2012;19:40. doi: 10.1186/1423-0127-19-40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Samarakoon R, Dobberfuhl AD, Cooley C, Overstreet JM, Patel S, Goldschmeding R, Meldrum KK, Higgins PJ. Induction of renal fibrotic genes by TGF-β1 requires EGFR activation, p53 and reactive oxygen species. Cell Signal. 2013;25(11):2198–209. doi: 10.1016/j.cellsig.2013.07.007. [DOI] [PubMed] [Google Scholar]
- 8.Liu N, He S, Ma L, Ponnusamy M, Tang J, Tolbert E, Bayliss G, Zhao TC, Yan H, Zhuang S. Blocking the class I histone deacetylase ameliorates ameliorateand inhibits renal fibroblast activation via modulating TGF-beta and EGFR signaling. PLoS One. 2013;8(1):e54001. doi: 10.1371/journal.pone.0054001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Jacot W, Mollevi C, Fina F, Lopez-Crapez E, Martin PM, Colombo PE, Bibeau F, Romieu G, Lamy PJ. High EGFR protein expression and exon 9 PIK3CA mutations are independent prognostic factors in triple negative breast cancers. BMC Cancer. 2015;15:986. [DOI] [PMC free article] [PubMed]
- 10.Wang L, Xiao J, Gu W, Chen H. Sex Difference of Egfr Expression and Molecular Pathway in the Liver: Impact on Drug Design and Cancer Treatments? J Cancer. 2016;7(6):671–80. [DOI] [PMC free article] [PubMed]
- 11.Togashi Y, Masago K, Fukudo M, Terada T, Ikemi Y, Kim YH, Fujita S, Irisa K, Sakamori Y, Mio T, Inui K, Mishima M. Pharmacokinetics of erlotinib and its active metabolite OSI-420 in patients with non-small cell lung cancer and chronic renal failure who are undergoing hemodialysis. J Thorac Oncol. 2010;5(5):601–5. [DOI] [PubMed]
- 12.Chen J, Chen JK, Harris RC. Deletion of the epidermal growth factor receptor in renal proximal tubule epithelial cells delays recovery from acute kidney injury. Kidney Int. 2012;82(1):45–52. doi: 10.1038/ki.2012.43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Cheng W, Hu Y, Sheng R. Development of EGFR family small molecule inhibitors for anticancer intervention: an overview of approved drugs and clinical candidates. Curr Med Chem. 2014;21(38):4374–404. doi: 10.2174/0929867321666140915142809. [DOI] [PubMed] [Google Scholar]
- 14.Passmore JC, Fleming JT, Tyagi SC, Falcone JC. Tyrosine kinase receptor alteration of renal vasoconstriction in rats is sex- and age-related. Can J Physiol Pharmacol. 2012;90(10):1372–9. doi: 10.1139/y2012-093. [DOI] [PubMed] [Google Scholar]
- 15.Gordon MS, Hussey M, Nagle RB, Lara PN, Jr, Mack PC, Dutcher J, Samlowski W, Clark JI, Quinn DI, Pan CX, Crawford D. Phase II study of erlotinib in patients with locally advanced or metastatic papillary histology renal cell cancer: SWOG S0317. J Clin Oncol. 2009;27(34):5788–93. doi: 10.1200/JCO.2008.18.8821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Wainberg ZA, Lin LS, DiCarlo B, Dao KM, Patel R, Park DJ, Wang HJ, Elashoff R, Ryba N, Hecht JR. Phase II trial of modified FOLFOX6 and erlotinib in patients with metastatic or advanced adenocarcinoma of the oesophagus and gastro-oesophageal junction. Br J Cancer. 2011;105(6):760–5. doi: 10.1038/bjc.2011.280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Zhou C, Wu YL, Chen G, Feng J, Liu XQ, Wang C, Zhang S, Wang J, Zhou S, Ren S, Lu S, Zhang L, Hu C, Hu C, Luo Y, Chen L, Ye M, Huang J, Zhi X, Zhang Y, Xiu Q, Ma J, Zhang L, You C. Final overall survival results from a randomised, Phase III study of erlotinib versus chemotherapy as first-line treatment of EGFR mutation-positive advanced non-small-cell lung cancer (OPTIMAL, CTONG-0802). Ann Oncol. 2015;26(9):1877–83. [DOI] [PubMed]
- 18.Chiorean EG, Ramasubbaiah R, Yu M, Picus J, Bufill JA, Tong Y, Coleman N, Johnston EL, Currie C, Loehrer PJ. Phase II trial of erlotinib and docetaxel in advanced and refractory hepatocellular and biliary cancers: Hoosier Oncology Group GI06-101. Oncologist. 2012;17(1):13. doi: 10.1634/theoncologist.2011-0253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Lorenzen S, Riera Knorrenschild J, Haag GM, Pohl M, Thuss-Patience P, Bassermann F, Helbig U, Weißinger F, Schnoy E, Becker K, Stocker G, Rüschoff J, Eisenmenger A, Karapanagiotou-Schenkel I, Lordick F. Lapatinib versus lapatinib plus capecitabine as second-line treatment in human epidermal growth factor receptor 2-amplified metastatic gastro-oesophageal cancer: a randomised phase II trial of the Arbeitsgemeinschaft Internistische Onkologie. Eur J Cancer. 2015;51(5):569–76. doi: 10.1016/j.ejca.2015.01.059. [DOI] [PubMed] [Google Scholar]
- 20.van Meerten E, Franckena M, Wiemer E, van Doorn L, Kraan J, Westermann A, Sleijfer S. Phase I study of cisplatin, hyperthermia, and lapatinib in patients with recurrent carcinoma of the uterine cervix in a previously irradiated area. Oncologist. 2015;20(3):241–2. doi: 10.1634/theoncologist.2014-0365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Pal SK, Figlin RA. Ramucirumab in metastatic renal cell carcinoma: The beginning or the end? Cancer. 2014 doi: 10.1002/cncr.28636. [DOI] [PubMed] [Google Scholar]
- 22.Garcia JA, Hudes GR, Choueiri TK, Stadler WM, Wood LS, Gurtler J, Bhatia S, Joshi A, Hozak RR, Xu Y, Schwartz JD, Thompson JA. A phase 2, single-arm study of ramucirumab in patients with metastatic renal cell carcinoma with disease progression on or intolerance to tyrosine kinase inhibitor therapy. Cancer. 2014 doi: 10.1002/cncr.28634. [DOI] [PubMed] [Google Scholar]
- 23.Fuchs CS, Tomasek J, Yong CJ, et al. Ramucirumab monotherapy for previously treated advanced gastric or gastro-oesophageal junction adenocarcinoma (REGARD): an international, randomised, multicentre, placebo-controlled, phase 3 trial. Lancet. 2014;383:31–9. doi: 10.1016/S0140-6736(13)61719-5. [DOI] [PubMed] [Google Scholar]
- 24.Malamitsi-Puchner A, Tziotis J, Tsonou A, Protonotariou E, Sarandakou A, Creatsas G. Changes in serum levels of vascular endothelial growth factor in males and females throughout life. J Soc Gynecol Investig. 2000;7:309–12. doi: 10.1016/S1071-5576(00)00073-3. [DOI] [PubMed] [Google Scholar]
- 25.Baserga M, Bares AL, Hale MA, Callaway CW, McKnight RA, Lane PH, Lane RH. Uteroplacental insufficiency affects kidney VEGF expression in a model of IUGR with compensatory glomerular hypertrophy and hypertension. Early Hum Dev. 2009;85:361–7. doi: 10.1016/j.earlhumdev.2008.12.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Gatti DM, Zhao N, Chesler EJ, Bradford BU, Shabalin AA, Yordanova R, Lu L, Rusyn I. Sex-specific gene expression in the BXD mouse liver. Physiol Genomics. 2010;42(3):456–68. doi: 10.1152/physiolgenomics.00110.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hayes KS, Hager R, Grencis RK. Sex-dependent genetic effects on immune responses to a parasitic nematode. BMC Genomics. 2014;15:193. doi: 10.1186/1471-2164-15-193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Mozhui K, Lu L, Armstrong WE, Williams RW. Sex-specific modulation of gene expression networks in murine hypothalamus. Front Neurosci. 2012;6:63. doi: 10.3389/fnins.2012.00063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Wang L, Xiao J, Gu W, Chen H. Sex difference of egfr expression and molecular pathway in the liver: impact on drug design and cancer treatments? J Cancer. 2016;7(6):671–80. doi: 10.7150/jca.13684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Jiao Y, Chen H, Yan J, Wang L, Huang Y, Liu X, WilIiams RW, Lu L, Wang Y, Gu W. Genome-wide gene expression profiles in antioxidant pathways and their potential sex differences and connections to vitamin C in mice. Int J Mol Sci. 2013;14(5):10042–62. doi: 10.3390/ijms140510042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Gatti D, Maki A, Chesler EJ, Kirova R, Kosyk O, Lu L, Manly KF, Williams RW, Perkins A, Langston MA, Threadgill DW, Rusyn I. Genome-level analysis of genetic regulation of liver gene expression networks. Hepatology. 2007;46(2):548–57. doi: 10.1002/hep.21682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Davies MN, Lawn S, Whatley S, Fernandes C, Williams RW, Schalkwyk LC. To what extent is blood a reasonable surrogate for brain in gene expression studies: estimation from mouse hippocampus and spleen. Front Neurosci. 2009;3:54. doi: 10.3389/neuro.15.002.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Wu Y, Williams EG, Dubuis S, Mottis A, Jovaisaite V, Houten SM, Argmann CA, Faridi P, Wolski W, Kutalik Z, Zamboni N, Auwerx J, Aebersold R. Multilayered genetic and omics dissection of mitochondrial activity in a mouse reference population. Cell. 2014;158(6):1415–30. doi: 10.1016/j.cell.2014.07.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Xu R, Li Q, Zhou J, Zhou X, Perelman JM, Kolosov VP. Secretoneurin induces airway mucus hypersecretion by enhancing the binding of EGF to NRP1. Cell Physiol Biochem. 2014;33(2):446–56. doi: 10.1159/000358625. [DOI] [PubMed] [Google Scholar]
- 35.Rizzolio S, Rabinowicz N, Rainero E, Lanzetti L, Serini G, Norman J, Neufeld G, Tamagnone L. Neuropilin-1-dependent regulation of EGF-receptor signaling. Cancer Res. 2012;72(22):5801–11. doi: 10.1158/0008-5472.CAN-12-0995. [DOI] [PubMed] [Google Scholar]
- 36.Cariboni A, Davidson K, Dozio E, Memi F, Schwarz Q, Stossi F, Parnavelas JG, Ruhrberg C. VEGF signalling controls GnRH neuron survival via NRP1 independently of KDR and blood vessels. Development. 2011;138(17):3723–33. doi: 10.1242/dev.063362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Hamilton JA, Callaghan MJ, Sutherland RL, Watts CK. Identification of PRG1, a novel progestin-responsive gene with sequence homology to 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase. Mol Endocrinol. 1997;11(4):490–502. doi: 10.1210/mend.11.4.9909. [DOI] [PubMed] [Google Scholar]
- 38.Jalava SE, Porkka KP, Rauhala HE, Isotalo J, Tammela TL, Visakorpi T. TCEB1 promotes invasion of prostate cancer cells. Int J Cancer. 2009;124(1):95–102. doi: 10.1002/ijc.23916. [DOI] [PubMed] [Google Scholar]
- 39.Chang KK, Liu LB, Li H, Mei J, Shao J, Xie F, Li MQ, Li DJ. TSLP induced by estrogen stimulates secretion of MCP-1 and IL-8 and growth of human endometrial stromal cells through JNK and NF-κB signal pathways. Int J Clin Exp Pathol. 2014;7(5):1889–99. [PMC free article] [PubMed] [Google Scholar]
- 40.Hillenbrand A, Fassler J, Huber N, Xu P, Henne-Bruns D, Templin M, Schrezenmeier H, Wolf AM, Knippschild U. Changed adipocytokine concentrations in colorectal tumor patients and morbidly obese patients compared to healthy controls. BMC Cancer. 2012;12:545. doi: 10.1186/1471-2407-12-545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Bollée G, Flamant M, Schordan S, Fligny C, Rumpel E, Milon M, Schordan E, Sabaa N, Vandermeersch S, Galaup A, Rodenas A, Casal I, Sunnarborg SW, Salant DJ, Kopp JB, Threadgill DW, Quaggin SE, Dussaule JC, Germain S, Mesnard L, Endlich K, Boucheix C, Belenfant X, Callard P, Endlich N, Tharaux PL. Epidermal growth factor receptor promotes glomerular injury and renal failure in rapidly progressive crescentic glomerulonephritis. Nat Med. 2011;17(10):1242–50. doi: 10.1038/nm.2491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Papagregoriou G, Erguler K, Dweep H, Voskarides K, Koupepidou P, Athanasiou Y, Pierides A, Gretz N, Felekkis KN, Deltas C. A miR-1207-5p binding site polymorphism abolishes regulation of HBEGF and is associated with disease severity in CFHR5 nephropathy. PLoS One. 2012;7(2):e31021. doi: 10.1371/journal.pone.0031021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Dahlhoff M, Frances D, Kloepper JE, Paus R, Schäfer M, Niemann C, Schneider MR. Overexpression of epigen during embryonic development induces reversible, epidermal growth factor receptor-dependent sebaceous gland hyperplasia. Mol Cell Biol. 2014;34(16):3086–95. doi: 10.1128/MCB.00302-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.McIntyre E, Blackburn E, Brown PJ, Johnson CG, Gullick WJ. The complete family of epidermal growth factor receptors and their ligands are co-ordinately expressed in breast cancer. Breast Cancer Res Treat. 2010;122(1):105–10. doi: 10.1007/s10549-009-0536-5. [DOI] [PubMed] [Google Scholar]
- 45.Abbott BD, Buckalew AR, DeVito MJ, Ross D, Bryant PL, Schmid JE. EGF and TGF-alpha expression influence the developmental toxicity of TCDD: dose response and AhR phenotype in EGF, TGF-alpha, and EGF + TGF-alpha knockout mice. Toxicol Sci. 2003;71(1):84–95. doi: 10.1093/toxsci/71.1.84. [DOI] [PubMed] [Google Scholar]
- 46.Mo Y, Chen J, Schlueter CF, Hoyle GW. Differential susceptibility of inbred mouse strains to chlorine-induced airway fibrosis. Am J Physiol Lung Cell Mol Physiol. 2013;304:L92–102. doi: 10.1152/ajplung.00272.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Haston CK, Cory S, Lafontaine L, Dorion G, Hallett MT. Strain-dependent pulmonary gene expression profiles of a cystic fibrosis mouse model. Physiol Genomics. 2006;25:336–45. doi: 10.1152/physiolgenomics.00208.2005. [DOI] [PubMed] [Google Scholar]
- 48.Wang L, Liu H, Jiao Y, Wang E, Clark SH, Postlethwaite AE, Gu W, Chen H. Differences between mice and humans in regulation and the molecular network of collagen, type III, alpha-1 at the gene expression level: obstacles that translational research must overcome. Int J Mol Sci. 2015;16(7):15031–56. doi: 10.3390/ijms160715031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Darr J, Klochendler A, Isaac S, Geiger T, Eden A. Phosphoproteomic analysis reveals Smarcb1 dependent EGFR signaling in malignant rhabdoid tumor cells. Mol Cancer. 2015;14:167. doi: 10.1186/s12943-015-0439-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Mirza Z, Schulten HJ, Farsi HM, Al-Maghrabi JA, Gari MA, Chaudhary AG, Abuzenadah AM, Al-Qahtani MH, Karim S. Molecular interaction of a kinase inhibitor midostaurin with anticancer drug targets, S100A8 and EGFR: transcriptional profiling and molecular docking study for kidney cancer therapeutics. PLoS One. 2015;10(3):e0119765. doi: 10.1371/journal.pone.0119765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Matušan-Ilijaš K, Damante G, Fabbro D, Dorđević G, Hadžisejdić I, Grahovac M, Avirović M, Grahovac B, Jonjić N, Lučin K. EGFR expression is linked to osteopontin and Nf-κB signaling in clear cell renal cell carcinoma. Clin Transl Oncol. 2013;15(1):65–71. doi: 10.1007/s12094-012-0889-9. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All raw collected data which used in this manuscript are freely available from http://www.genenetwork.org/webqtl/main.py.