

Abstract
Background
Changes in immunoglobulin G (IgG) constant domain (Fc) glycosylation are associated with changes in rheumatoid arthritis (RA) disease activity in response to pregnancy. Here, we sought to determine whether the same holds true for variable domain (Fab) glycosylation.
Methods
IgGs were captured from RA and control sera obtained before (RA only), during and after pregnancy, followed by Fc and Fab separation, glycan release, and mass spectrometric detection. In parallel, glycans from intact IgG were analysed. The data was used to calculate glycosylation traits, and to estimate the level of Fab glycosylation.
Results
The overall level of Fab glycosylation was increased in RA patients compared to controls, while no differences in Fab glycosylation patterns were found. For the Fc and intact IgG (Total) previously observed differences in galactosylation and bisection were confirmed. Furthermore, increased galactosylation of Fc and Total were associated with lower disease activity and autoantibody positivity. In addition, the change in Fc galactosylation associated with the change in disease activity during pregnancy and after delivery, while this was not the case for Fab.
Conclusions
In contrast to changes in Fc glycosylation, changes in Fab glycosylation are not associated with improvement of RA during pregnancy and arthritis flare after delivery.
Electronic supplementary material
The online version of this article (doi:10.1186/s13075-016-1172-1) contains supplementary material, which is available to authorized users.
Keywords: Rheumatoid arthritis, Disease activity, Inflammation, Pregnancy
Background
Rheumatoid arthritis (RA) is an autoimmune disease, for which it is well known that patients may improve during pregnancy [1]. The immunoglobulin G (IgG) fragment crystallisable (Fc) N-glycan compositions, specifically the levels of galactosylation and sialylation (Fig. 1), have been recognised to be different in RA patients compared to healthy controls and to be associated with RA disease activity and its improvement during pregnancy [2–4].
Fig. 1.
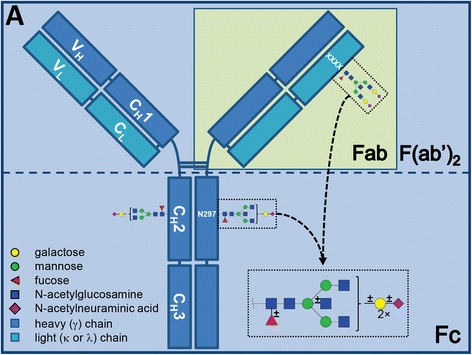
Schematic representation of immunoglobulin G. Indicated is what is generally considered as the Fc and Fab or F(ab’)2 portion. The glycans of both Fc and Fab consist of the same building blocks, although the most abundant glycans may differ vastly in their appearance. This figure was originally published in Bondt et al, Immunoglobulin G (IgG) Fab glycosylation analysis using a new mass spectrometric high-throughput profiling method reveals pregnancy-associated changes. Mol Cell Proteomics. 2014; 13:3029–3039. © the American Society for Biochemistry and Molecular Biology, and slightly adapted for the current manuscript
In addition to the Fc moiety, that always bears N-glycans, the hypervariable region of the antigen-binding fragment (Fab) may harbour N-glycans [5]. These glycans are generally present on approximately 15–25% of the Fab portions, and have been found to alter the binding properties of antibodies to their antigen, as well as effector functions, and are known to change during pregnancy [6–10]. Interestingly, for rheumatoid arthritis-specific autoantibodies (anti-citrullinated protein antibodies (ACPA)) the vast majority has been shown to carry these Fab glycans [11].
In view of the known differences between the Fc N-glycans of RA patients and healthy controls, and its association with disease activity and improvement during pregnancy, we aimed to determine whether similar changes and associations can be found regarding the Fab N-glycans. In addition, the level of Fab glycosylation was investigated.
Methods
Study population and data collection
For the current study we used sera from the Pregnancy-induced Amelioration of Rheumatoid Arthritis (PARA) study, a prospective cohort study on pregnancy and RA [12]. At all time points, disease activity (DAS28; based upon a count of 28 swollen and tender joints and C-reactive protein (CRP)) and medication was recorded. For the analysis we selected a subset of RA patients (n = 33) with the most pronounced (> = 0.6) or no change in disease activity, both during and after pregnancy, in search of maximum contrast. Only sera obtained before pregnancy, during the third trimester of pregnancy, and 26 weeks postpartum were analysed. In addition sera obtained from 32 healthy pregnant volunteers were analysed (third trimester and 6 months after delivery only). All patients fulfil the 1987 American College of Rheumatology criteria for RA. The study was in compliance with the Helsinki Declaration and was approved by the Ethics Review Board at the Erasmus University Medical Center, Rotterdam, The Netherlands.
IgG capturing and N-glycan release
IgGs were affinity-captured in duplicate from human serum in a 96-well format, as described before, with minor modifications [9]. One set was used for IdeS digestion to separate the Fab and Fc portions, whereas the other set was kept intact to analyse the total IgG glycans. Dried samples were reconstituted in 5 μL phosphate-buffered saline and 10 μL 2% (w/v) sodium dodecyl sulfate solution, instead of 10 and 20, respectively. Similarly, PNGase F was added in 10 μL.
Sialic acid derivatization and HILIC enrichment
To prevent the loss of sialic acids in the mass spectrometric detection of released glycans a derivatization protocol was applied as described before, with minor modifications [9, 13]. Briefly, 2 μL released glycans were added to 20 μL ethyl esterification reagent in a V-bottom 96-well plate, and incubated at 37 °C for 1 h. After the incubation 20 μL acetonitrile was added. Ethyl esterified N-glycans were purified from the reaction mixture using hydrophilic interaction liquid chromatography (HILIC) solid phase extraction (SPE) with a piece of cotton thread as the solid phase in a micro tip as described before [9].
MALDI-TOF-MS and data processing
Five microliter of eluate was mixed on the spot with sodium hydroxide-spiked Super-DHB matrix (Sigma-Aldrich, St. Louis, MO, USA) on an AnchorChip plate. Automated matrix-assisted laser desorption/ionization (MALDI)-time of flight (TOF) mass spectrometry (MS) measurements were performed summing 10,000 shots per spot with 250 shot steps and a full spot random walk. Details on data processing, extraction, and the calculation of glycosylation traits are described in Additional file 1.
Statistical analysis
Statistical analysis was performed in Stata 13SE (StataCorp, College Station, TX, USA). Bonferroni correction for multiple testing was performed throughout, with final significance thresholds depicted in the tables with results. First, a Wilcoxon rank-sum test was performed to explore the differences in glycosylation between RA patients and healthy controls outside pregnancy, at ≥26 weeks after delivery. Next we investigated which clinical covariates (use of medication, autoantibody (AAb) positivity, age at delivery and disease activity) were associated with the glycosylation in RA patients using linear regression analysis. To study changes in glycosylation over time the Wilcoxon matched-pairs signed-rank test was used, comparing pre-conception with the third trimester for the patients, and third trimester with 26 weeks postpartum for both patients and controls. Association of the changes in glycosylation over time and the accompanying changes in disease activity were explored using Spearman correlation.
Results
Response during pregnancy and flare after delivery
Clinical characteristics of the patients and controls are given in Additional file 1: Table S1. The patients who improved (n = 14) during pregnancy showed a mean decrease in DAS28 (delta DAS28; ΔDAS) of -1.7 (SD 0.5), whereas the nine patients that worsened showed a ΔDAS of +1.6 (SD 0.5). Ten patients did not change during pregnancy (ΔDAS = 0.0; SD 0.4). After delivery the 12 patients that flared had a mean ΔDAS of +1.5 (SD 0.8), while the 12 improving patients showed a ΔDAS of −1.5 (SD 0.6). Nine patients without a change in disease activity between the third trimester and 6 months postpartum (ΔDAS = −0.1; SD 0.4) were selected.
Fab glycosylation is higher in RA patients outside pregnancy when compared to controls, yet with similar glycans
The calculated level of Fab glycosylation in the patients (21.4%; [IQR 19.5–25.4%]) was significantly higher than in controls (16.5%; [IQR 13.3–18.2%]) at the non-pregnant time point 26 weeks postpartum (Tables 1, 2 and 3). However, no differences between the levels of galactosylation, sialylation, fucosylation or presence of bisecting N-acetylglucosamine were observed for the Fab glycosylation (Tables 1, 2 and 3) compared between healthy controls and RA patients. However, for both Fc and total IgG (Total) there was a lower level of galactosylation and higher level of bisection in RA patients compared to the healthy controls.
Table 1.
Median percentages and corresponding interquartile ranges for all time points and glycosylation traits
| Healthy | RA | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 3rd trima | 26 wkpp | pre-conception | 3rd trim | 26 wkpp | ||||||
| Median (%) | IQR | Median (%) | IQR | Median (%) | IQR | Median (%) | IQR | Median (%) | IQR | |
| Fc | ||||||||||
| Galactosylation | 81.5 | [78.1–83.2] | 71.6 | [67.9–74.7] | 60.8 | [52.2–64.3] | 73.6 | [66.5–79.5] | 61.0 | [56.8–66.7] |
| Sialylation | 18.4 | [17.1–20.5] | 13.1 | [11.5–15.8] | 11.0 | [9.0–13.1] | 16.0 | [14.0–19.7] | 11.5 | [9.4–14.1] |
| Fucosylation | 96.8 | [95.8–97.4] | 97.3 | [96.6–98.0] | 97.9 | [97.5–98.3] | 97.5 | [96.5–98.0] | 97.7 | [97.2–98.2] |
| Bisection | 8.7 | [7.3–9.7] | 9.2 | [8.3–10.5] | 10.3 | [9.0–11.5] | 8.6 | [7.4–10.2] | 11.0 | [9.5–13.4] |
| Total | ||||||||||
| Galactosylation | 82.9 | [80.6–84.6] | 74.8 | [70.3–77.0] | 66.0 | [59.9–69.0] | 77.5 | [73.5–82.2] | 68.6 | [59.1–73.5] |
| Sialylation | 26.2 | [23.9–28.7] | 21.3 | [19.4–22.6] | 20.8 | [19.1–23.8] | 27.3 | [24.4–28.4] | 22.6 | [20.0–24.5] |
| Fucosylation | 95.4 | [94.7–96.2] | 96.5 | [95.5–97.2] | 96.9 | [96.2–97.3] | 96.2 | [95.4–96.7] | 96.7 | [96.0–97.1] |
| Bisection | 12.6 | [10.6–14.1] | 14.8 | [13.3–17.3] | 18.1 | [15.8–19.8] | 15.2 | [13.6–17.7] | 18.1 | [16.8–19.6] |
| Fab | ||||||||||
| Galactosylation | 96.8 | [96.3–97.0] | 97.2 | [96.5–97.8] | 96.6 | [95.7–97.7] | 97.3 | [96.9–97.8] | 96.8 | [96.0–97.3] |
| Sialylation | 81.6 | [80.4–84.3] | 79.0 | [77.8–82.6] | 79.7 | [77.0–81.1] | 82.7 | [80.6–84.6] | 80.2 | [76.9–81.6] |
| Fucosylation | 70.6 | [68.0–75.3] | 76.6 | [74.5–80.6] | 75.6 | [71.9–80.5] | 73.2 | [61.6–75.8] | 77.7 | [72.8–79.7] |
| Bisection | 37.0 | [36.1–41.4] | 46.6 | [41.7–52.2] | 51.5 | [46.5–55.2] | 42.1 | [33.8–44.4] | 52.2 | [46.7–54.6] |
| % glycosylation | 17.6 | [13.7–21.0] | 16.5 | [13.3–18.2] | 22.0 | [18.7–27.4] | 28.0 | [23.3–32.7] | 21.4 | [19.5–25.4] |
a Abbreviations: trim trimester, wkpp weeks postpartum, IQR interquartile range
Table 2.
Number of observations obtained for each category and time point
| Healthy | RA | ||||
|---|---|---|---|---|---|
| 3rd trima | 26 wkpp | pre-conc. | 3rd trim | 26 wkpp | |
| Fc | |||||
| Galactosylation | 29 | 25 | 33 | 31 | 33 |
| Sialylation | 29 | 25 | 33 | 31 | 33 |
| Fucosylation | 29 | 25 | 33 | 31 | 33 |
| Bisection | 29 | 25 | 33 | 31 | 33 |
| Total | |||||
| Galactosylation | 25 | 25 | 33 | 30 | 31 |
| Sialylation | 25 | 25 | 33 | 30 | 31 |
| Fucosylation | 25 | 25 | 33 | 30 | 31 |
| Bisection | 25 | 25 | 33 | 30 | 31 |
| Fab | |||||
| Galactosylation | 17 | 15 | 29 | 25 | 23 |
| Sialylation | 17 | 15 | 29 | 25 | 23 |
| Fucosylation | 17 | 15 | 29 | 25 | 23 |
| Bisection | 17 | 15 | 29 | 25 | 23 |
| % glycosylation | 15 | 15 | 29 | 24 | 21 |
a Abbreviations: trim trimester, wkpp weeks postpartum, pre-conc. pre-conception
Table 3.
p values obtained for comparing glycosylation traits of controls vs. cases and over time
| RA vs. healthy at 26 wkppa | Healthy | RA | ||
|---|---|---|---|---|
| 3rd trim vs. 26 wkpp | pre-conc. vs. 3rd trim | 3rd trim vs. 26 wkpp | ||
| Fc | ||||
| Galactosylation | 0.000 | 0.000 | 0.000 | 0.000 |
| Sialylation | 0.077 | 0.000 | 0.000 | 0.000 |
| Fucosylation | 0.072 | 0.000 | 0.003 | 0.048 |
| Bisection | 0.004 | 0.003 | 0.000 | 0.000 |
| Total | ||||
| Galactosylation | 0.001 | 0.000 | 0.000 | 0.000 |
| Sialylation | 0.179 | 0.000 | 0.000 | 0.000 |
| Fucosylation | 0.889 | 0.000 | 0.000 | 0.012 |
| Bisection | 0.000 | 0.000 | 0.000 | 0.000 |
| Fab | ||||
| Galactosylation | 0.347 | 0.333 | 0.004 | 0.148 |
| Sialylation | 0.754 | 0.028 | 0.000 | 0.007 |
| Fucosylation | 0.709 | 0.005 | 0.000 | 0.003 |
| Bisection | 0.104 | 0.005 | 0.000 | 0.001 |
| % glycosylation | 0.002 | 0.401 | 0.009 | 0.249 |
For the comparison of RA vs. healthy a significance threshold of p <0.0038 was used (Bonferroni corrected for 13 tests), and for the time comparisons p <0.0013 (corrected for 42 tests). Significant observations are highlighted in bold font
a Abbreviations: wkpp weeks postpartum, trim trimester
Galactosylation of Fc and intact IgG, but not Fab, associate with disease activity and autoantibody positivity
In addition, it was explored which clinical covariates were associated with the levels of the calculated glycosylation traits at 26 weeks postpartum. Covariates were first tested univariate, and those with p <0.2 were included in the multivariate analysis. No Fab glycosylation models remained significant after Bonferroni correction. However, for galactosylation of both Fc and Total multivariate models could be composed with R2 values of 0.4 and 0.5, respectively. For these models the main contributing factors were disease activity (beta = −0.5) and autoantibody positivity (beta = +0.4). The results for all multivariate models are shown in Additional file 1: Table S2.
Fab glycosylation changes during pregnancy and after delivery are not associated with changes in disease activity
Galactosylation and sialylation of Fab, Fc and Total increased between pre-conception and the third trimester of pregnancy in RA patients (Tables 1, 2 and 3; Fig. 2). The opposite was observed after delivery in both patients and healthy controls, except for the Fab portion where only a decrease in sialylation was observed. For fucosylation and bisection a decrease during pregnancy (RA only) and increase after delivery was observed for Fab, Fc and Total, in both patients and controls (Tables 1, 2 and 3). In addition, during pregnancy an increase in the level of Fab glycosylation was found in RA patients. Of note, changes in glycosylation traits that were high, e.g. Fab galactosylation and Fc fucosylation, were often not significant after Bonferroni correction (Tables 1, 2 and 3; Fig. 2).
Fig. 2.
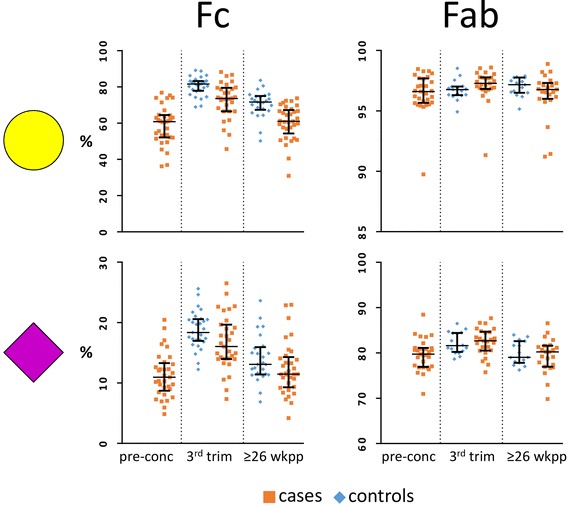
Fc and Fab galactosylation and sialylation in RA and healthy controls. The percentages of galactosylation (yellow circle) and sialylation (purple diamond) on the Fc show a clear increase during pregnancy (from pre-conception to the 3rd trimester; RA only) and decrease after delivery (from 3rd trim to 26 weeks postpartum), as well as differences between cases and controls. No differences between cases and controls, and only minor pregnancy-associated changes were observed for the Fab glycosylation. Abbreviations: pre-conc pre-conception, trim trimester, wkpp weeks postpartum
The Fc galactosylation change from pre-conception to the third trimester of pregnancy was negatively associated with the change in disease activity in that time span (rho = −0.39, p = 0.029; Additional file 1: Table S3, Additional file 1: Figure S1). The opposite was observed for the change Fc and Total galactosylation from third trimester to 26 weeks postpartum (rho = 0.48 and 0.62, p = 0.006 and 0.001, respectively). For neither the Fab glycosylation traits nor the Fab glycosylation level these associations were observed.
Discussion
There is a strong association of IgG Fc galactosylation with RA disease activity and the pregnancy-associated improvement thereof [2, 3]. For the Fab portion of IgG this has not yet been investigated. Therefore, we applied our recently developed method for the analysis of Fab glycosylation on samples from a set of RA and healthy individuals in the context of pregnancy. Interestingly, we found no qualitative differences in Fab glycosylation between cases and controls nor an association with changes in disease activity when we compared several glycosylation traits. This may in part be caused by the fact that Fab galactosylation and sialylation levels are already high in the non-pregnant state, leaving less room for a potential pregnancy-associated increase.
Since the Fab portion of IgG – in contrast to the Fc portion – is not always glycosylated, the association of changes in Fab glycosylation with disease activity could be quantitative rather than qualitative. In line with this, it has recently been published that the RA-specific ACPA autoantibodies express high levels of Fab glycosylation [11]. Indeed, in this study we did observe increased levels of Fab glycosylation. However, even though the levels are higher in RA patients compared to controls, they were not associated with disease activity.
In the current study we chose to include only a limited number of patients and time points. Therefore minor associations between changes in RA disease activity and changes in Fab glycosylation could have been missed.
Conclusions
In conclusion, this study clearly demonstrates that, in contrast to changes in the glycosylation of the IgG Fc domain, changes in the glycosylation of the IgG Fab domain do not seem to play a major role in the pregnancy-associated improvement of RA.
Acknowledgements
The authors thank Mahmoud Habeeb for his efforts in testing purification and measurement conditions.
Funding
AB was funded by the Dutch Arthritis Foundation (NR-10-1-411), AB and MW were supported by funding from the European Union’s Seventh Framework Programme (FP7-Health-F5-2011) under grant agreement no 278535 (HighGlycan). The funding bodies had no role in the design of the study and collection, analysis, and interpretation of data and in writing the manuscript.
Availability of data and materials
Extracted MS data is supplied in Additional file 2. Clinical parameters will not be shared for confidentiality reasons.
Authors’ contributions
AB designed and performed the experiments, acquired and analyzed the data, drafted the manuscript, gave final approval of the version to be published, and agreed to be accountable for all aspects of the work. MW designed the experiments, supported the interpretation of the data, was involved in drafting and revising the manuscript, gave final approval of the version to be published, and agreed to be accountable for all aspects of the work. MK supported the interpretation of the data, revised the manuscript, gave final approval of the version to be published, and agreed to be accountable for all aspects of the work. JH was involved in drafting and revising the manuscript, gave final approval of the version to be published, and agreed to be accountable for all aspects of the work. RD designed the experiments, supported the interpretation of the data, was involved in drafting and revising the manuscript, gave final approval of the version to be published, and agreed to be accountable for all aspects of the work. All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Ethics approval and consent to participate
The study was in compliance with the Helsinki Declaration and was approved by the Ethics Review Board at the Erasmus University Medical Center, Rotterdam, The Netherlands. All patients gave informed consent.
Abbreviations
- AAb
autoantibody
- ACPA
anti-citrullinated protein antibodies
- DAS28
based upon a count of 28 swollen and tender joints
- Fab
antigen-binding fragment
- Fc
fragment crystallisable
- HILIC
hydrophilic interaction liquid chromatography
- IgG
immunoglobulin G
- MALDI
matrix-assisted laser desorption/ionization
- MS
mass spectrometry
- PARA
Pregnancy-induced Amelioration of Rheumatoid Arthritis
- RA
rheumatoid arthritis
- SPE
solid phase extraction
- TOF
time of flight
Additional files
Additional Methods, Figure and Tables. The additional file contains an addition to the Methods, a figure, and three additional tables. (DOCX 262 kb)
Data used for analysis. Contains the relative abundances of the observed glycans and the calculated glycosylation traits for the RA patients and healthy controls used in the current study. (XLSX 120 kb)
Contributor Information
Albert Bondt, Email: a.bondt@lumc.nl.
Manfred Wuhrer, Email: m.wuhrer@lumc.nl.
T. Martijn Kuijper, Email: t.kuijper@erasmusmc.nl.
Johanna M. W. Hazes, Email: j.hazes@erasmusmc.nl
Radboud J. E. M. Dolhain, Phone: +31 (0)10 70 33805, Email: r.dolhain@erasmusmc.nl
References
- 1.Ince-Askan H, Dolhain RJ. Pregnancy and rheumatoid arthritis. Best Pract Res Clin Rheumatol. 2015;29(4-5):580–96. doi: 10.1016/j.berh.2015.07.001. [DOI] [PubMed] [Google Scholar]
- 2.Parekh RB, Dwek RA, Sutton BJ, et al. Association of rheumatoid arthritis and primary osteoarthritis with changes in the glycosylation pattern of total serum IgG. Nature. 1985;316(6027):452–7. doi: 10.1038/316452a0. [DOI] [PubMed] [Google Scholar]
- 3.Bondt A, Selman MHJ, Deelder AM, et al. Association between galactosylation of immunoglobulin G and improvement of rheumatoid arthritis during pregnancy is independent of sialylation. J Proteome Res. 2013;12(10):4522–31. doi: 10.1021/pr400589m. [DOI] [PubMed] [Google Scholar]
- 4.van de Geijn FE, Wuhrer M, Selman MH, et al. Immunoglobulin G galactosylation and sialylation are associated with pregnancy-induced improvement of rheumatoid arthritis and the postpartum flare: results from a large prospective cohort study. Arthritis Res Ther. 2009;11(6):R193. doi: 10.1186/ar2892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Abel CA, Spiegelberg HL, Grey HM. Carbohydrate content of fragments and polypeptide chains of human.gamma.G-myeloma proteins of different heavy-chain subclasses. Biochemistry. 1968;7(4):1271–78. doi: 10.1021/bi00844a004. [DOI] [PubMed] [Google Scholar]
- 6.Wright A, Tao MH, Kabat EA, et al. Antibody variable region glycosylation: position effects on antigen binding and carbohydrate structure. EMBO J. 1991;10(10):2717–23. doi: 10.1002/j.1460-2075.1991.tb07819.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Coloma MJ, Trinh RK, Martinez AR, et al. Position effects of variable region carbohydrate on the affinity and in vivo behavior of an anti-(1 → 6) dextran antibody. J Immunol. 1999;162(4):2162–70. [PubMed] [Google Scholar]
- 8.Jacquemin M, Radcliffe CM, Lavend’homme R, et al. Variable region heavy chain glycosylation determines the anticoagulant activity of a factor VIII antibody. J Thromb Haemost. 2006;4(5):1047–55. doi: 10.1111/j.1538-7836.2006.01900.x. [DOI] [PubMed] [Google Scholar]
- 9.Bondt A, Rombouts Y, Selman MHJ, et al. Immunoglobulin G (IgG) Fab glycosylation analysis using a new mass spectrometric high-throughput profiling method reveals pregnancy-associated changes. Mol Cell Proteomics. 2014;13(11):3029–39. doi: 10.1074/mcp.M114.039537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.van de Bovenkamp FS, Hafkenscheid L, Rispens T, et al. The emerging importance of IgG Fab glycosylation in immunity. J Immunol. 2016;196(4):1435–41. doi: 10.4049/jimmunol.1502136. [DOI] [PubMed] [Google Scholar]
- 11.Rombouts Y, Willemze A, van Beers JJ, et al. Extensive glycosylation of ACPA-IgG variable domains modulates binding to citrullinated antigens in rheumatoid arthritis. Ann Rheum Dis. 2016;75(3):578––85. doi: 10.1136/annrheumdis-2014-206598. [DOI] [PubMed] [Google Scholar]
- 12.de Man YA, Dolhain RJEM, van de Geijn FE, et al. Disease activity of rheumatoid arthritis during pregnancy: results from a nationwide prospective study. Arthritis Rheum. 2008;59(9):1241–48. doi: 10.1002/art.24003. [DOI] [PubMed] [Google Scholar]
- 13.Reiding KR, Blank D, Kuijper DM, et al. High-throughput profiling of protein N-glycosylation by MALDI-TOF-MS employing linkage-specific sialic acid esterification. Anal Chem. 2014;86(12):5784–93. doi: 10.1021/ac500335t. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Extracted MS data is supplied in Additional file 2. Clinical parameters will not be shared for confidentiality reasons.