

Abstract
Flavivirus nonstructural protein 1 (NS1) is a unique secreted non-structural glycoprotein. Although it is absent from the flavivirus virion, intracellular and extracellular forms of NS1 have essential roles in viral replication and the pathogenesis of infection. The fate of NS1 in insect cells has been more controversial, with some reports suggesting it is exclusively cell-associated. Here, we confirm NS1 secretion from cells of insect origin and characterize its physical, biochemical, and functional properties in the context of dengue virus (DENV) infection. Unlike mammalian cell-derived NS1, which displays both high mannose and complex type N-linked glycans, soluble NS1 secreted from DENV-infected insect cells contains only high mannose glycans. Insect cell-derived secreted NS1 also has different physical properties including smaller and more heterogeneous sizes, and the formation of less stable NS1 hexamers. Both mammalian and insect cell-derived NS1 bind to complement proteins C1s, C4 and C4 binding protein as well as to a novel partner, mannose-binding lectin (MBL). Binding of NS1 to MBL protects DENV virus against MBL-mediated neutralization by the lectin pathway of complement activation. As we detected secreted NS1 and DENV virus together in the saliva of infected Aedes aegypti mosquitoes, these findings suggest a mechanism of viral immune evasion at the very earliest phase of infection.
Keywords: dengue virus, flavivirus, nonstructural protein NS1, mannose-binding lectin, lectin pathway, neutralization, complement, pathogenesis, insect cell-derived NS1
INTRODUCTION
Dengue fever, dengue hemorrhagic fever, and dengue shock syndrome have increased markedly due to the global spread of dengue virus (DENV) and the expansion of its mosquito vectors, Aedes aegypti and Aedes albopictus. DENV is now the most important viral illness transmitted by insects (1) with an estimated 390 million infections per year (2). DENV belongs to the genus Flavivirus of the Flaviviridae family. It is a positive sense single stranded enveloped RNA virus with an approximately 11-kilobase genome encoding three structural proteins [capsid (C), pre-membrane/membrane (prM/M), and envelope (E)] and seven nonstructural (NS) proteins (NS1, NS2a, NS2b, NS3, NS4a, NS4b, and NS5) that are absent from the virion but function in viral replication and immune evasion within an infected cell. Among the NS proteins, only NS1 is displayed on cell surfaces and secreted from infected cells. DENV NS1 is a 46-kDa glycoprotein with two N-linked glycans and originally was described as a soluble complement-fixing antigen (3). DENV NS1 also functions intracellularly as a co-factor for viral replication by interacting with other structural and non-structural proteins, although the mechanistic basis for this activity remains poorly understood (4, 5).
NS1 is postulated to contribute to the pathogenicity of dengue diseases. High plasma levels of NS1 and terminal complement complexes C5b-9 observed in DENV-infected patients correlate with the development of severe dengue disease (6). Soluble NS1 enhances infection in hepatocyte cell lines (7), elicits autoantibodies that cross-react with platelets and extracellular matrix proteins, and promotes endothelial cell damage via antibody-dependent complement-mediated cytolysis (8-10). Moreover, soluble NS1 facilitates immune complex formation and complement activation, which can trigger microvesicle shedding from erythrocytes (6, 11). Soluble hexameric NS1 associates with lipids and forms lipoprotein particles that can impact vascular endothelial function and integrity (12, 13). Binding of soluble NS1 to endothelial cells triggers TLR-2, TLR-4, and TLR-6 activation resulting in proinflammatory cytokine generation and loss of endothelial cell junction integrity (13-15). Soluble NS1 also binds back to the plasma membrane of cells through an interaction with specific sulfated glycosaminoglycans (GAGs) (16), which could contribute to tissue-specific vascular leakage that occurs during a severe secondary DENV infection (6, 17). Moreover, NS1 has a separate immune evasion activity as it antagonizes complement activation, which limits inhibitory effects on flavivirus infection (18-20).
Previous studies have reported that flavivirus-infected mammalian but not insect cells secrete NS1 into the extracellular milieu (4, 18, 19). However, using a more sensitive assay for NS1 detection, we and others have detected NS1 in the culture supernatants of DENV-infected insect cells including Spodoptera frugiperda cells (20) and Aedes albopictus mosquito-derived C6/36 cells (21). The presence of NS1 in the culture medium of insect cells was not due to lysis but rather an active process requiring N-linked glycosylation and the protein secretory pathway. Here, we investigated the physical and functional properties of NS1 secreted from DENV-infected insect cells. We showed that soluble NS1 from DENV-infected insect cells, analogous to mammalian-cell derived NS1, formed hexamers and bound to human complement components C1s, C4, and C4b binding protein to restrict classical pathway-dependent complement activation. We also observed a novel complement evasion function of NS1 via an interaction with mannose binding lectin (MBL) to protect DENV from MBL-mediated neutralization. Finally, along with DENV, NS1 was detected in the saliva of infected mosquitoes suggesting a potential role for limiting complement recognition and activation at the site of the mosquito bite.
MATERIALS AND METHODS
Cells and viruses
All transformed cell lines were obtained from the ATCC. Three insect cell lines, the C6/36 clone of Aedes albopictus cells, the AP-61 cell line from Aedes pseudoscutellaris, and the TRA-284 cell line from Toxorhynchites amboinensis were grown in L-15 Medium (Gibco) supplemented with 10% tryptose phosphate broth (TPB, Sigma) and 10% fetal bovine serum (FBS, Hyclone) at 28°C. The swine fibroblast cell line (PscloneD) was grown in L-15 medium supplemented with 10% TPB and 10% FBS at 37°C. BHK and Vero cell lines were cultured in Dulbecco's Modified Eagle's medium (DMEM) supplemented with 10% FBS, 50 mM HEPES, 4 mM L-glutamine, 100 units/ml penicillin G, and 100 μg/ml streptomycin sulfate and complete MEM medium at 37°C. BHK cells that stably propagate a DENV-2 subgenomic replicon (BHK DENV-2 Rep cells (22)) were grown in DMEM containing 10% FBS and 3 μg/ml puromycin (Sigma).
Dengue virus (DENV) serotype 2 strain 16681 was propagated in C6/36 cells (17). DENV titer was determined by focus-forming assay (FFA) on Vero cells as previously described (8).
DENV NS1
Sources of insect cell-derived NS1 were from culture supernatants of 1) DENV-infected C6/36 cells; 2) DENV-infected AP-61 cells; 3) DENV-infected TRA-284 cells; and 4) DENV NS1 transfected Drosophila S2 cells. Mammalian cell derived DENV NS1 was prepared from culture supernatants of serum-free DENV-infected PscloneD or Vero cells and BHK DENV-2 Rep cells. For the PscloneD and Vero cells, cells were infected with DENV-2 at an MOI of 0.1 and cultured in serum-free L-15 medium. At 3 days post-infection, clarified supernatants were collected, aliquotted and stored at −70°C until use. In some experiments, supernatants were concentrated by ultrafiltration with a 30-kDa cut-off membrane (Millipore). Levels of NS1 in culture supernatants were determined by quantitative NS1-ELISA (6).
Drosophila S2 cells stably expressing recombinant NS1 of DENV-2 (S2-NS1 cells) were generated following co-transfection of plasmids pMT/DV2NS1 (gene encoding NS1 protein of dengue serotype 2 cloned into an expression vector, pMT/BiP/V5-His) and pCoHygro after hygromycin drug selection. The resulting S2/DV2NS1 stable cells were induced by CuSO4 for 5-7 days to produce and secrete S2-NS1 into serum-free culture supernatant. S2-NS1 was purified subsequently by immunoaffinity chromatography using anti-NS1 mAb 2G6 coupled beads and eluted with 20 mM diethylamine in phosphate buffered saline (PBS) (pH 11.3), as modified from a previously described protocol (6). Purified S2-NS1 possessed similar physical characteristics to that of naturally produced viral NS1 including a heat-sensitive dimeric form in SDS-PAGE and Western blot analysis and a hexameric form in solution, as judged by size exclusion chromatography (Puttikhunt et al., unpublished data).
Soluble NS1 from DENV-infected C6/36 or Vero supernatants was purified by affinity chromatography. Briefly, one liter of DENV-infected C6/36 or Vero supernatants was centrifuged at 13,000 × g for 10 min at 4°C before purification by monoclonal anti-NS1 antibodies (clone 2E11) conjugated to cyanogen bromide beads (6). Purified NS1 protein was dialyzed against PBS and stored at −70°C until use.
DENV infection
C6/36 or PscloneD cells were infected with DENV at MOIs of 0.01, 0.1 and 1.0. After viral adsorption for 2 h at 37°C, cell monolayers were washed once with culture medium and incubated for 2 days. Levels of NS1 and DENV in culture supernatants were determined by NS1-ELISA (6) and FFA as described above. To determine cell viability, cells were collected using a cell-scraper and washed twice with RPMI-1640 (Gibco) containing 1% FBS, then 106 cells were added to 300 μl of RPMI-1640 containing 1% FBS and 0.1 μg propidium iodide (Invitrogen) and analyzed by flow cytometry (BD Bioscience). To determine viral antigen expression, cells were fixed and permeabilized with 3.7% formaldehyde in PBS and 1% Triton X-100 in PBS prior to incubation with mAbs specific for NS1 (clone 2G6) or E (clone 3H5) proteins. After three washes, the cells were incubated with a rabbit anti-mouse immunoglobulin conjugated to fluorescein isothiocyanate (FITC) (1:50, Dako) before analysis by flow cytometry.
NS1 in the culture supernatants of DENV-infected C6/36, AP-61 and TRA-284 cells was concentrated by ultrafiltration as described above and analyzed by Western blotting with a mAb specific for NS1, clone 1F11, and horseradish peroxidase (HRP)-conjugated anti-goat IgG (GE Healthcare).
Native PAGE
Fifty-fold concentrated supernatants from DENV-infected C6/36 or PscloneD cells were mixed with non-reducing sample buffer (40% glycerol, 0.02% bromophenol blue in 0.2 M Tris-HCl, pH 6.8) without boiling. After separation in a 4-12% gradient native-polyacrylamide gel (Native-PAGE) as previously described (23), Western blotting analysis was performed with anti-DENV NS1 mAb as described above.
Size exclusion chromatography
Two hundred microliters of fifty-fold concentrated DENV-2 infected C6/36 or PscloneD culture supernatants were loaded onto a Superdex 200 column (Pharmacia) at a rate of 0.4 ml/min. Thirty fractions (0.5 ml each) were collected and the NS1 concentration in each fraction was determined by NS1-ELISA (6). The reference protein size markers (0.5 mg/ml, 200 μl), including thyroglobulin (Sigma), ribonuclease A (Pharmacia), bovine serum albumin (Sigma), and human immunoglobulin (Alpha Therapeutic Corporation) were separated in parallel.
Sucrose density gradient centrifugation
Fifty-fold concentrated DENV-infected C6/36 or PscloneD supernatant was loaded on top of a 5% to 55% sucrose gradient (1 ml at each concentration with a 5% decrease in concentration at each step) and centrifuged at 35,000 × g for 20 h at 4°C using an SW-40ti rotor (Beckman, USA). One-milliliter fractions were collected from top to bottom. The presence of NS1 in each fraction was detected by Western blotting with anti NS1 mAb (clone 1F11).
NS1 deglycosylation
Fifty-fold concentrated culture supernatants from DENV-infected C6/36 or PscloneD cells were subjected to digestion with Endo H or PNGase F (New England Biolabs) according to the manufacturer's protocol. NS1 bands were determined by Western blotting assay as described above.
Complement-NS1 binding ELISA
The binding of NS1 to complement proteins including proC1s, C4, C4BP, factor H, and MBL was determined by ELISA as previously described (24). Briefly, microtiter plates were coated with bovine serum albumin (BSA) or purified human complement C1s proenzyme (proC1s), C4, C4BP, factor H and MBL (15 μg/ml in PBS). Nonspecific binding sites in each well were blocked with 2% BSA in PBS before incubation with equal concentrations (determined by NS1-ELISA) of NS1 in the culture supernatants from DENV-infected C6/36 or PscloneD cells, S2-NS1 cells or purified soluble NS1 from DENV-infected C6/36 or Vero cells. The plates were incubated with 10 μg/ml of DENV NS1-specific polyclonal Abs followed by biotinylated goat anti-mouse IgG (10 μg/ml, Sigma) and HRP-conjugated streptavidin (2 μg/ml). The signal was developed with ortho-phenylene diaminedihydrochloride (OPD, Sigma) and the reaction was terminated by the addition of 4M H2SO4 before evaluation on a 96 well-plate reader at an O.D. of 492 nm. Five washes with 0.05% Tween-20 in PBS were performed between each step. Mock-infected supernatants from each cell type were used as negative controls.
NS1-MBL co-immunoprecipitation
One ml of NS1-containing supernatants was incubated with purified human MBL (0.75 μg, Enzon, Inc.) at 4°C overnight and then immunoprecipitated with anti-NS1 mAb (2G6)-Sepharose at 4°C overnight. After five washes, the samples were added to SDS sample buffer and separated by 10% SDS-PAGE (reducing or non-reducing conditions). Western blotting was performed using 0.2 μg/ml of mouse anti-human MBL mAb (Quidel) followed by HRP-conjugated anti-mouse IgG (GE Healthcare).
Cell surface binding of secreted NS1
Binding of NS1 to cell surfaces was determined as previously described (16). Briefly, the human keratinocyte cell line HaCat and BHK cells were removed from tissue culture plates after incubation with an EDTA solution (4 mM EDTA plus 10% FBS in PBS). Cells (5 × 105) in suspension were incubated on ice for 1 h with 100 μl of 2-fold serially diluted NS1-containing culture supernatants from DENV-infected C6/36 and PscloneD or DENV NS1 transfected S2 cells. After washing once with 3 ml of medium, 50 μl of DENV-2 NS1 specific mAb 2G6 (10 μg/ml) was added to the cells and incubated on ice for 45 min. After subsequent washing, bound primary mAbs were detected after a 30 min incubation with a 1:500 dilution of Alexa Fluor 647-conjugated anti-mouse IgG (Invitrogen). In some experiments, 10 μg/ml of heparin was added to inhibit NS1 binding. Propidium iodide (0.2 μg/ml) was added immediately before analysis by flow cytometry to exclude dead cells.
Recruitment of C4BP to the cell surface by soluble NS1
The experiments were performed as previously described (25). Adherent BHK or HaCat cells were detached by an EDTA solution as described above. Concentrated serum-free supernatants from S2-NS1 cells (containing 32 μg/ml NS1 as judged by NS1-ELISA) or BSA (2.5 μg/ml) were mixed with C4BP (2.5 μg/ml) and the BSA-C4BP or NS1-C4BP mixture was added to cells (1 × 106). After a 1 h incubation on ice, the cells were washed three times with cold DMEM. Surface-bound C4BP and NS1 were detected after adding mouse anti-DENV NS1 2G6 mAb (26) or mouse anti-human C4BP MK 104 mAb (27) followed by a 1:500 dilution of Alexa Fluor 647-conjugated goat anti-mouse IgG (Invitrogen) and 0.2 μg/ml propidium iodide to exclude dead cells and analyzed by flow cytometry. BHK cells were incubated with BSA, supernatants containing NS1 or C4BP alone.
C4BP cofactor activity assay on the cell surface
Recruitment of C4BP by NS1 to the cell surface followed by measurement of the cofactor activity of C4BP was performed according to a previous study (25). Briefly, serum-free supernatants (200 μl) containing 12 μg/ml of NS1 from DENV-infected C6/36, DENV-infected PscloneD cells or S2-NS1 cells were incubated with C4BP (2.5 μg/ml, 50 μl) for 1 h on ice. Adherent BHK cells (1 × 106) were detached by PBS supplemented with 8 mM EDTA. Subsequently, 10% FBS was added to the C4BP solution (with or without NS1) and incubated for 1 h on ice. After three washes with 1 ml DMEM at 4°C, cells were washed with 1 ml dextrose veronal buffered saline (DVB, 2.5 mM veronal buffer pH 7.35, 25 mM NaCl, and 240 mM glucose) at 4°C. C4b (400 ng) and factor I (200 ng) diluted in DVB were added to cell pellets in a total volume of 50 μl. After a 15-min incubation at 37°C, cells were pelleted and supernatants were mixed with SDS sample buffer supplemented with 2-mercaptoethanol (5% v/v) and subjected to 12% SDS-PAGE. C4b fragmentation was analyzed by Western blotting using a 1:2,000 dilution of anti-C4d monoclonal antibody (Quidel) followed by a 1:5,000 dilution of HRP-conjugated anti-mouse IgG.
DENV-MBL binding ELISA
The binding of MBL to DENV was determined by a MBL-DENV capture ELISA as previously described (28). Briefly, microtiter plates were adsorbed with an anti-DENV prM protein-specific mAb 2H2 (10 μg/ml in PBS) (29) at 4°C overnight. DENV-2 stocks were diluted in DMEM to 5 × 105 PFU/ml in 50 μl and added to the wells. After five washes, MBL (3 μg/ml) diluted in binding buffer [20 mM Tris-HCl (pH 7.4), 0.05% Tween 20, 0.1% (w/v) BSA, 1 M NaCl, and 10 mM CaCl2] together with 2-fold increasing concentrations (0-50 ng/ml) of recombinant NS1 from S2 cell culture supernatants. After five washes, DENV-bound MBL was detected with biotin-conjugated mouse anti-human MBL mAb (1 μg/ml, Cedarlane) followed by HRP-conjugated streptavidin (2 μg/ml). After six final washes with PBS, the signal was detected as described above. For a negative control, purified human MBL was diluted in 20 mM Tris-HCl (pH 7.4) and 150 mM NaCl supplemented with 40 mM EDTA. In some experiments, NS1 was depleted by pre-incubation of culture supernatants with anti-NS1 mAb (2E11)-Sepharose (100 μl of supernatants per 30 μl of packed beads) at 4°C for 24 h. Anti-E mAb (4G2)-Sepharose was used as negative controls for NS1 depletion condition.
Inhibition of MBL mediated neutralization of DENV by DENV NS1
Experiments showing the effect of NS1 on MBL-mediated neutralization of DENV were modified from a previous paper (28). Briefly, 3 μg/ml of purified recombinant human MBL was pre-incubated with 10-fold increasing concentrations [0-150 ng/ml in gelatin veronal buffer supplemented with Mg2+ and Ca2+ (GVB++, Complement Technology)] of recombinant NS1 from S2 cells at 4°C for 2 h. DENV (102 FFU) was incubated with an MBL-NS1 mixture for 1 h at 37°C. Infectious DENV was quantified subsequently by FFA (8) or plaque forming assay (28). In some experiments, NS1 in culture supernatants was depleted as described above and MBL (3 μg/ml) premixed with purified hexameric NS1 (24) (4, 21 and 43 μg/ml), thyroglobulin (10, 50 and 100 μg/ml), or BSA (1, 5 and 10 μg/ml) with molar ratios of 1:1, 1:5, and 1:10 (MBL:protein). Soluble mannan (100 μg/ml) was used as an MBL inhibitor. To confirm this effect of NS1 derived from DENV-infected insect cells, purified soluble NS1 from DENV-infected C6/36 was used. Briefly, MBL (2 μg/ml) was premixed with purified soluble NS1 from DENV-infected C6/36 supernatant (0, 1, 2, 4, 8 and 16 μg/ml). An equal concentration of BSA was used as a negative control.
Virus attachment assay
The virus attachment assay was based on a published protocol (30) with some modifications. DENV (105 FFU/ml) was incubated with or without 80 μg/ml MBL (diluted with GVB++) together with purified NS1 (50 μg/ml) from NS1 transfected S2 cells or DENV-infected Vero cells at 37°C for 1 h. The mixture was then added to a BHK cell suspension (4 × 103, MOI = 25) followed by a 2-h incubation on ice. Cells were washed five times with GVB++ prior to RNA extraction using a conventional TRIzol-chloroform method. DENV genome copies per cell were determined by real time RT-PCR with capsid specific primers using SYBR green detection technique (6). The penultimate wash buffers were collected and subjected to RNA extraction and virus quantitation as negative controls.
Detection of DENV and soluble NS1 in the saliva of infected mosquitoes
Laboratory-reared female Aedes aegypti mosquitoes were used in this experiment. Mosquitoes were reared at 27°C ± 2°C at 80% relative humidity under a photo regime of 12:12 h (light:dark). Larvae were fed with fish food (C.P. Hi Pro®) on a regular basis. Adults were provided with 10% sucrose solutions as an energy source. Aedes aegypti females (4-6 day-old) were starved for 6 hours prior to the experiments.
Infectious blood meal comprising human blood mixed with DENV was used. Mosquitoes were infected using a membrane feeding technique for 30 min. Glass feeders containing infectious blood meal were maintained at 37°C during feeding using a water circulator system. After feeding, mosquitoes were anesthetized using a chill table. The fully engorged females were sorted and placed in new containers. Mosquitoes were incubated at 27°C ± 2 at 80% relative humidity for 12 days and provided with sucrose soaked cotton balls until use. After 12 days, mosquitoes were fed with fresh blood using a membrane feeding technique for 30 min. After feeding, the remaining blood was collected and NS1 and DENV viral RNA were determined by Western blotting, NS1-ELISA and semi-nested RT-PCR as previous described (6, 31). Mosquitoes were anesthetized using a chill table. Approximately 50 mosquito salivary glands were dissected, smeared, and fixed on glass slides to determine NS1 expression using an immunofluorescent staining assay as previously described (6). Another 50 dissected salivary glands were lysed with 0.5% SDS sample buffer for 15 min prior to 12% SDS-PAGE (heated and reduced conditions). Western blotting was performed using anti-DENV NS1 mAb as described above.
Statistical analysis
Data sets were statistically analyzed using Prism software (GraphPad). The Kolmogorov Smirnov test was used to test for normality. Comparisons between each group were done by t-test when the distribution of the variables was comparable to a normal parametric distribution; otherwise the Mann Whitney test was used. P ≤ 0.05 was considered to be statistically significant. All analyzed P values were 2-sided. All data were analyzed from 3 to 4 independent experiments.
RESULTS
Presence of extracellular NS1 in the culture supernatants of DENV-infected insect cells
Based on immunoprecipitation and Western blot analyses, it was believed previously that NS1 was secreted only from flavivirus-infected mammalian but not insect cells (18, 19). However, consistent with more recent studies (20, 21), our sensitive NS1 capture ELISA (6) detected soluble NS1 in the culture supernatants of DENV-infected C6/36 Aedes albopictus cells at 2 days post-infection albeit at lower levels than those from DENV-infected mammalian cells, including PscloneD (a pig fibroblast) and BHK (a baby hamster kidney fibroblast) cells (Fig. 1A). Increasing the inoculum of DENV virions per cell (multiplicity of infection, MOI) did not affect cell viability at the time evaluated (Fig. 1B), but did increase the percentage of cells infected as determined by intracellular staining with anti-NS1 (Fig. 1C) and anti-E (Fig. 1D) antibodies, virus production (Fig. 1E), and NS1 accumulation in the supernatants (Fig. 1A). Although marginal differences in the percentage of infected cells and virion production were observed between DENV-infected insect and mammalian cells (Fig. 1E), NS1 levels were higher (~2-fold, P = 0.012) in the supernatants of infected mammalian cells (Fig. 1A). These data imply that infected insect cells secrete NS1 less efficiently than infected mammalian cells. Notably, there was a direct correlation between the levels of NS1 in the supernatant and titers of infectious virus produced in all three cell types (Fig. 1F-H).
FIGURE 1. Detection of extracellular NS1 derived from DENV-infected insect cells.

C6/36 (black bars), PscloneD (white bars) and BHK (grey bars) cells were infected with DENV at an MOI of 0.01, 0.1 or 1.0 for 2 days. The levels of secreted NS1 (A), cell viability (B), percentage of NS1- (C) and E- (D) expressing cells and DENV production (E) were determined by NS1-ELISA, flow cytometry analysis using propidium iodide exclusion, anti-NS1 mAb (clone 1F11) and anti-E mAb (clone 4G2) and FFA, respectively. The data are shown as mean ± standard deviation (SD) of three to four independent experiments. Asterisks denote levels of NS1 detected in supernatants that are statistically different (* P < 0.05). The correlation coefficients between secreted NS1 and DENV production in DENV-infected C6/36 (F), DENV-infected PscloneD (G) and DENV-infected BHK (H) were analyzed by linear regression. NS1 in 50-fold concentrated supernatants of DENV-infected C6/36, AP-61 and TRA-284 cells was analyzed by Western blotting using anti-DENV NS1 mAb clone 1F11 (I).
To determine if NS1 secretion by mosquito cells was specific to Aedes albopictus cells, we extended our study to cells of other mosquito species, including AP-61 and TRA-284, which are derived from Aedes pseudoscutellaris and Toxorhynchites amboinensis, respectively. These two cell lines have been shown to efficiently generate DENV progeny and are used to isolate DENV from clinical specimens (32). Indeed, NS1 was detectable in the supernatants of all types of DENV-infected mosquito cells (Fig. 1I).
N-linked glycosylation is required for NS1 secretion from DENV-infected mosquito cells
N-linked glycosylation on NS1 is essential for its folding, surface expression, secretion, and hexamer stabilization in mammalian cells (33, 34). To confirm that NS1 in the supernatants of infected insect cells (Fig. 1A) was not caused by cell lysis, DENV-infected C6/36 cells (at 24 h post-infection) were washed extensively to remove NS1-containing culture medium and treated with fresh medium containing tunicamycin, an inhibitor of N-linked glycosylation (35, 36) (Fig. 2). NS1 accumulation in the supernatant of infected C6/36 cells was reduced 10-fold over a period of 6 h with tunicamycin treatment (10 μg/ml), compared to cells treated with the vehicle control, dimethyl sulfoxide (DMSO) (P = 0.02, Fig. 2A). Under these conditions, virtually no differences in the percentages of DENV-infected cells were observed (Fig. 2B-C) and, importantly, the intracellular expression levels of NS1 as indicated by mean fluorescence intensity (MFI) between tunicamycin-treated and untreated cells were equivalent (black bars, Fig. 2B). These data suggest that inhibition of N-linked glycosylation by tunicamycin interfered only with extracellular NS1 secretion but not intracellular NS1 or DENV infection. The decrease of NS1 secretion into the culture supernatants was not due to cell death or cell cycle arrest as there was no difference in cell viability and total cell number between tunicamycin-treated and DMSO-treated control cells (Fig. 2D-E). Similar to NS1 secretion, and as reported previously (37, 38), levels of DENV released into the supernatant also were reduced after tunicamycin treatment, suggesting that N-linked glycosylation is necessary for the egress of DENV from infected C6/36 cells (Fig. 2F). Overall, our data indicate that DENV-infected insect cells can secrete NS1 into the extracellular milieu and that this process requires N-linked glycosylation on the NS1 molecule.
FIGURE 2. Tunicamycin treatment inhibits NS1 and virion release from DENV-infected insect cells.

At 24 h after DENV infection, C6/36 cells were extensively washed to remove NS1-containing culture medium and treated with fresh medium containing either 0.1% DMSO or 10 μg/ml of tunicamycin in 0.1% DMSO for 6 h. Levels of NS1 in supernatants (A), percentage and mean fluorescent intensity (MFI) of NS1 positive cells (B) and percentages of E-positive cells (C), percentage of living cells (D), total cell number (E) and intracellular and extracellular DENV genome copies (F) were analyzed as described in Materials and Methods. The data are shown as mean ± SD from three independent experiments. Asterisks denote the means that are statistically different among indicated groups (* P < 0.05).
Physical properties of secreted NS1 from DENV-infected insect cells
Despite the heterogeneous sizing profile of secreted flavivirus NS1 obtained from culture supernatants of mammalian cells that stably propagate DENV or WNV subgenomic replicons (16, 32), the main species of soluble NS1 is hexameric, which is similar to the soluble NS1 generated by flavivirus-infected mammalian cells (18). The size of mosquito cell-derived soluble NS1 was investigated by native gel electrophoresis, size exclusion chromatography and sucrose density gradient centrifugation (Fig. 3). In native-PAGE analysis, secreted NS1 from DENV-infected C6/36 cells displayed three major bands, similar to NS1 secreted from DENV-infected Vero cells (molecular weight ranged between >200-660 kDa), but were slightly smaller in size and migrated faster in the gel [upper panel, lane 1 (C6/36-NS1) versus lane 3 (Vero-NS1); Fig. 3A]. The size of extracellular NS1 released from Vero cells was slightly larger, likely due to the addition of complex type N-linked glycans at position Asn130 of the NS1 molecule in the Golgi apparatus, which only occurs in mammalian cells (39) (lower panel, lane 3 versus lanes 1, 2 and 4; Fig. 3A). In contrast, intracellular NS1 derived from both DENV-infected mammalian and insect cells appeared more heterogeneous showing broadly smeared bands on the native PAGE (upper panel, lanes 2 and 4; Fig. 3A) whereas, in a regular SDS-PAGE under heat and reducing condition, both secreted and cell-associated NS1 from insect and mammalian cells migrated at 45-50 kDa (lower panel, Fig. 3A).
FIGURE 3. Oligomeric assembly, glycosylation pattern and stability of NS1 secreted from DENV-infected insect cells.

Fifty-fold concentrated supernatants (lanes 1 and 3) and cell lysates (lanes 2 and 4) from DENV-infected C6/36 cells (lanes 1 and 2) and Vero cells (lanes 3 and 4) were separated by Native-PAGE (A, upper panel) and reducing 10% SDS-PAGE (A, lower panel) and Western blotted with 1F11 DENV NS1 specific mAb. B. Fifty-fold concentrated DENV-2 infected C6/36 (dashed line) and PscloneD (solid line) supernatants were passed over a Superdex 200 column and fractions were collected and quantitated for NS1 concentration by NS1-ELISA. Molecular weights of reference protein standards fractionated in parallel are depicted. C. Fifty-fold concentrated DENV-infected C6/36 or PscloneD supernatant was loaded on a 5% to 55% sucrose gradient. NS1 in each fraction was detected by Western blotting with 1F11 anti-NS1 mAb. Arrowheads indicate molecular weights of protein standards. D. Endoglycosidase (Endo H or PNGase F) treatment of extracellular NS1 derived from C6/36 (lanes 1, 2, 5 and 6) and PscloneD (lanes 3, 4, 7 and 8) cells prior to Western blot analysis with 1F11 anti-NS1 mAb. E-F. NS1 stability test.Supernatants from DENV-infected C6/36 and Vero cells were incubated in the presence of a protease inhibitor (10 mM PMSF) at 4°C or 37°C for 3 days. Amounts of NS1 at each time point of C6/36 cell-derived or Vero cell-derived at 4°C and 37°C were determined by ELISA and calculated by normalizing the zero-time point of each condition to 100 percent (E) and by Western blotting with 1F11 anti-NS1 mAb (F). Error bars indicate SD corresponding to three independent experiments. Values that are significantly different from the value for zero-time point are indicated by asterisks as follows: *, P < 0.05; **, P < 0.01; ***, P < 0.001.
To analyze the oligomeric states of NS1 secreted from infected insect cells in more detail, culture supernatants were subjected to size exclusion chromatography and sucrose density gradient centrifugation in parallel with those from infected mammalian cells (Fig. 3B-C). Consistent with secreted mammalian-cell derived NS1, the form of NS1 released from DENV-infected C6/36 cells ranged from monomer to oligomer. However, insect-cell derived NS1 exhibited a more heterogeneous profile with two separate peaks of monomer and higher-order oligomer whereas the sizing profile of NS1 from infected mammalian cells appeared as a broad single peak at around 44 to 660 kDa (Fig. 3B-C). Sucrose density gradient centrifugation analysis revealed a high-molecular weight, and possibly aggregated, form of insect-cell derived NS1, which was not observed in the supernatants of mammalian cells (fraction no. 1, Fig. 3C). As we previously reported that the complex type N-linked glycosylation at Asn130 is required for the stability of hexameric NS1 (34), and insect cells lack glycosyltransferase enzymes that synthesize hybrid and complex type N-linked glycans (40), we reasoned that the aggregated form of insect-cell derived NS1 was likely due to the lack of complex type glycans.
The glycosylation state of DENV NS1 secreted from infected mosquito cells was further analyzed by digestion with two glycosidase enzymes, endoglycosidase H (Endo H) which removes high mannose-content sugars (41) and peptide N-glycosidase F (PNGase F), which cleaves all types of N-linked glycans (42, 43). As expected, after digestion with either Endo H or PNGase F, NS1 secreted from DENV-infected C6/36 cells had similar sizes in SDS-PAGE (lanes 2 and 6, Fig. 3D), whereas Endo H digested DENV-infected mammalian cell-derived NS1 had a band that was higher in molecular weight compared to PNGase F digestion (lanes 4 versus 8, Fig. 3D). The presence of uncleaved NS1 after PNGase F treatment was likely from inefficient digestion (upper bands, lanes 6 and 8, Fig. 3D). These data confirm that secreted NS1 from DENV infected C6/36 cells had high-mannose type glycosylation whereas mammalian-cell derived NS1 displayed both Endo H-sensitive high mannose and Endo H-resistant complex type glycans.
Data from our previous study demonstrated that N-linked glycans regulate NS1 transport through the secretory pathway and affect protein stability (34). We therefore tested the stability of soluble NS1 protein derived from DENV-infected C6/36 and Vero cells by incubating the supernatants in the presence of a protease inhibitor (10 mM PMSF) at either 4°C or 37°C. The supernatants were collected daily for 3 days for NS1 quantification by ELISA (Fig. 3E) and Western blotting assay (Fig. 3F). As expected, NS1 derived from DENV-infected C6/36 cells which lacks complex-type N-linked glycans was degraded more rapidly especially at 37°C, compared with NS1 derived from DENV-infected Vero cells (Fig. 3E-F).
Functional characteristics of secreted NS1 from DENV-infected insect cells
Soluble NS1 from DENV-infected mammalian cells has immune evasion properties, as it directly antagonizes complement activation (24, 25); binding of DENV NS1 to C4 and C1s/C1s proenzyme resulted in C4 degradation and decreased complement activation (24). Additionally, DENV NS1 binds to C4b binding protein (C4BP), the major inhibitor of the classical/lectin pathway of complement activation in plasma, to control the activation of these pathways on cell surfaces (25). We hypothesize that insect-cell derived NS1 also has complement inhibitory functions. In the skin, at the site of the mosquito bite in which both DENV and, potentially soluble NS1 are simultaneously inoculated, complement antagonism might prevent the recognition and immediate complement-dependent neutralization of DENV and thus facilitate infection and spread.
To assess whether insect cell-derived NS1 could antagonize complement functions, we initially determined its capacity to bind human complement proteins (Fig 4A). Using comparable levels of NS1, as measured by ELISA (Fig. 4B), we observed that NS1 secreted from two different cells of insect origin (DENV-infected C6/36 cells and Drosophila S2 transfected cells) bound human complement proteins including C1s pro-enzyme (proC1s), C4, and C4BP to the same extent as mammalian cell-derived NS1 (Fig. 4A). As previously observed, and in contrast to West Nile virus (WNV) NS1 (18), DENV NS1 did not bind to factor H, the major complement regulatory protein of the alternative pathway (Fig. 4A).
FIGURE 4. Functional properties of extracellular NS1 derived from DENV-infected insect cells.

A. Insect-cell derived NS1 binds human complement. Microtiter plates were coated with BSA, factor H, C1s proenzyme (ProC1s), C4 and C4BP (5 μg/ml) and incubated with culture supernatants of DENV-infected C6/36, PscloneD and S2-NS1 cells. Bound NS1 was detected with specific mAbs. Values [optical density (O.D.) at 492 nm.] of NS1 binding to each complement component were subtracted from those of BSA control. Error bars indicate SD corresponding to three independent experiments. Asterisks denote NS1 binding that is statistically different from corresponding mock controls (** P < 0.01 and *** P < 0.001). Levels of NS1 in the supernatants were determined by NS1-ELISA (B). C-D. Insect-cell derived NS1 binds to cell surfaces via GAGs. Serum-free supernatants of DENV-infected PscloneD (C and D), C6/36 (C and D) and S2-NS1 (D) cells containing indicated concentrations of NS1 were incubated with BHK (C) and HaCat (D) cells and NS1 binding was detected by 2G6 anti-DENV NS1 mAb. Binding also was performed in the presence of 10 mM heparin (C). Error bars indicate SD of the mean corresponding to three independent experiments. Asterisks denote the means that are statistically different from 0 μg/ml of NS1 (* P < 0.05, ** P < 0.01 and *** P < 0.001). E. NS1 secreted from insect cells recruits C4BP to the cell surface. C4BP or BSA (2.5 μg/ml) was incubated with serum-free supernatants of S2-NS1 cells and 106 BHK cells. Binding of NS1 and C4BP to the cell surface was assessed by mAbs to DENV NS1 or C4BP and analyzed by flow cytometry. Examples of contour plots from three independent experiments are depicted. The y-axis indicates the levels of cell surface-associated C4BP or NS1, and the x-axis shows the forward scatter (FSC) of the cell population. Percentages of cells positive for surface-NS1 or C4BP staining are shown in the right upper quadrant. F. Extracellular NS1 derived from insect cells recruits C4BP to degrade C4b on the cell surface. Serum-free supernatants (containing 12 μg/ml NS1) from DENV-infected PscloneD (lane 2), DENV-infected C6/36 (lanes 3 and 6) or S2-NS1 (lane 4) cells were preincubated with 2.5 μg/ml C4BP and the NS1-C4BP solution was added to BHK cells. After three washes, the cells were incubated with C4b and factor I for 15 min. Cofactor activity was determined in the supernatants by the appearance of C4b cleavage products, a 70-kDa α3-C4d and a 45-kDa C4d after 12% SDS-PAGE under reducing condition followed by Western blot with anti-C4d mAb. G. Diagram of factor I (FI)-mediated cleavage of C4b in the presence of a cofactor C4BP. FI cleaves the α′chain of C4b at two sites (designated as A and B). Cleavage at either one of the two sites generates two partial cleavage fragments, C4d-α4 (59 kDa) and α3-C4d (70 kDa). Further cleavage by FI yields the final product C4d (45 kDa).
We next tested whether NS1 secreted from DENV-infected insect cells bound to the surface of cells. Again, analogous to mammalian-cell derived NS1 (16), NS1 secreted from DENV-infected and transfected insect cells bound to the surface of BHK cells in a dose-dependent manner (Fig. 4C). This binding required glycosaminoglycans (GAGs) on the BHK cell surface, as the soluble GAGs (e.g., heparin) competitively inhibited NS1 attachment to the cells (Fig. 4C). A dose-dependent binding of soluble NS1 from three different sources (DENV-infected mammalian PscloneD cells, DENV-infected insect C6/36 cells and S2-NS1 cells) also bound equivalently to the human skin keratinocyte cell line, HaCat (Fig. 4D). Keratinocytes are one of the major target cells for flavivirus infection in the skin and the cells also have immunomodulatory functions during virus inoculation by infected mosquitoes (44-47). Importantly, the interaction of insect cell-derived NS1 with the complement regulatory protein C4BP in solution led to the attachment of C4BP to the plasma membrane via the cell binding property of NS1 (Fig. 4E). C4BP on cell surfaces that was recruited by insect and mammalian cell-derived NS1 acted as a cofactor for factor I to cleave the α′ chain of C4b to inactive forms, the 70-kDa-partial-cleavage fragment α3-C4d and the final end product C4d (Fig. 4F, lanes 3 and 4; Fig. 4G). Of note, the 70-kDa partial cleavage product α3-C4d, also was detected without NS1 (Fig. 4F, lane 5) suggesting a small background interaction of C4BP with cells Fig. 4E, upper panel). However, C4d was not observed when NS1 or factor I was absent (Fig. 4F, lanes 1, 5 and 6). Collectively, these experiments establish that NS1 secreted from insect cells behaves similarly to mammalian-cell derived NS1 as it binds to cell surface glycosaminoglycans, specific human complement proteins, and recruits C4BP through its cofactor activity to degrade C4b on the surface of cells.
A novel antagonism function of soluble NS1: Inhibition of MBL mediated neutralization of DENV infection
Using a complement protein-binding ELISA, we observed that soluble NS1 derived from both mammalian and insect cells bound in a dose-dependent manner to mannose-binding lectin (MBL), a pattern recognition molecule that initiates the lectin pathway of complement activation (Fig. 5A-C). The interaction of secreted DENV NS1 and MBL was confirmed by co-immunoprecipitation experiments (Fig. 5D). Because we previously demonstrated that direct binding of MBL to DENV neutralized infection through both complement activation-dependent and -independent mechanisms (28, 30), we reasoned that direct binding of soluble NS1 to MBL might interfere with its recognition and neutralization of DENV, independent of complement activation. To test this, purified human MBL was pre-incubated with culture supernatants containing increasing concentrations of insect cell-derived NS1 prior to addition to microtiter wells containing immobilized virions (Fig. 5E). As expected, NS1 competitively bound MBL in solution, which resulted in decreased MBL binding to captured DENV (Fig. 5E). Removal of NS1 from the culture supernatants using NS1-specific mAb 2E11 (26) coupled Sepharose beads but not anti-E mAb (4G2)-Sepharose prior to incubating with purified MBL restored the MBL recognition of captured DENV (Fig. 5F). Of note, abrogation of Ca2+-dependent MBL binding to DENV in the presence of EDTA served as negative controls for these experiments (Fig. 5E-F).
FIGURE 5. NS1 protects DENV from MBL-mediated neutralization.

A-C. NS1 binds directly to MBL. Purified human MBL was coated on a microtiter plate and incubated with NS1 containing supernatants from DENV-infected C6/36, DENV-infected PscloneD and S2-NS1 cells (A) or with increasing concentrations of NS1 in culture supernatants of DENV-infected PscloneD and S2-NS1 cells (B) or with increasing concentrations of purified soluble NS1 from culture supernatants of DENV-infected C6/36 or Vero cells (C). Bound NS1 was detected with an anti-NS1 specific mAb. Error bars indicate SD for three independent experiments. D. Co-immunoprecipitation experiments. Serum-free supernatants from BHK-DENV2-Rep or control BHK cells were incubated with purified human MBL (0.75 μg) and Western blots were performed after immunoprecipitation with anti-DENV NS1 2G6 mAb-Sepharose. Immunoprecipitates were probed with a mouse anti-human MBL mAb after non-reduced (upper panel) or reduced (lower panel) SDS-PAGE. E-F. NS1 reduces MBL binding to DENV. DENV virions were captured on a microtiter plate using an anti-DENV prM protein-specific mAb and incubated with purified human MBL (3 μg/ml) with or without increasing concentrations of NS1 containing supernatants from S2-NS1 cells (E). MBL binding to captured DENV was detected with an anti-MBL specific mAb and inhibited by the presence of 20 mM EDTA. F. Culture supernatants from S2-NS1 cells (containing 12.5 μg/ml NS1) were incubated with anti-NS1 mAb (2E11)-Sepharose, anti-E mAb (4G2)-Sepharose or without Sepharose (no beads) prior to addition of purified human MBL and subjected to ELISA as described in E. MBL binding to DENV captured on a microtiter plate in GVB++ buffer (without NS1) or in buffer containing 20 mM EDTA were used as positive and negative controls, respectively. G-J. NS1 inhibits MBL-mediated neutralization of DENV. DENV virions were incubated with purified human MBL without or with indicated concentrations of NS1 in supernatants from S2-NS1 cells or with NS1 supernatants alone. The mixtures were then added to a BHK monolayer, and after carboxymethylcellulose overlay, cells were cultured for 3 days. After fixation and staining with anti-DENV E mAb, infected foci were counted (G). H. DENV virions were incubated with purified MBL in GVB++ buffer without or with NS1 containing supernatants (12 μg/ml) from S2-NS1 cells that were preincubated with anti-NS1 mAb (2E11)- or anti-E mAb (4G2)-Sepharose. The samples were added to a BHK monolayer and DENV-infected foci were determined as described in G. MBL-mediated neutralization of DENV was abolished by 20 mM EDTA. I. DENV virions were incubated with purified human MBL with or without indicated concentrations of purified soluble NS1 from DENV-infected C6/36 cells or with BSA control protein prior to the addition to a BHK monolayer to assess viral infectivity as described in G. J. DENV virions were incubated with MBL premixed with purified hexameric NS1 derived from BHK-DENV2-Rep, thyroglobulin or bovine serum albumin (BSA) at molar ratios of 1:1, 1:5, and 1:10 (MBL:protein). The samples were added to BHK monolayers, and after agarose overlay, cultured for 5 days. After fixation and staining of wells, plaques were counted. Soluble mannan (100 μg/ml) was used as an MBL inhibitor. K. Soluble NS1 facilitates DENV attachment to cells in the presence of human MBL. DENV was incubated with 80 μg/ml MBL (diluted with GVB++ buffer) alone or with MBL plus BSA or purified NS1 (50 μg/ml) derived from S2-NS1 cells or DENV-infected Vero cells. The mixture was then added to BHK cell suspensions for 2 h on ice. Cell-bound DENV was determined by real time RT-PCR. Error bars indicate SD corresponding to three independent experiments. Asterisks denote the means that are statistically different among indicated groups (* P < 0.05, ** P < 0.01 and *** P < 0.001).
Preincubation of MBL with increasing concentrations of soluble NS1 derived from insect cells (NS1 in supernatants from S2-NS1 cells) also resulted in an increased number of DENV-infected foci (Fig. 5G). Conversely, neutralization of DENV by MBL was restored when MBL was pre-incubated with NS1-containing supernatants that had been pre-cleared with anti-NS1 (2E11) but not anti-E (4G2) Sepharose beads (Fig. 5H). The dose-dependent activity of insect cell-derived NS1 to inhibit DENV neutralization by human MBL was further demonstrated using NS1 purified from culture supernatants of DENV-infected C6/36 cells (Fig. 5I). Consistent with these results, complement activation-independent neutralization of DENV by MBL was also inhibited by mammalian cell-derived NS1 (Fig. 5J). Pre-incubation of MBL with increasing concentrations of hexameric NS1 purified from the supernatants of BHK cells that propagate DENV-2 subgenomic replicons (22, 34), but not with the control proteins, abrogated MBL-mediated neutralization of DENV (Fig. 5J).
We next investigated the mechanism by which NS1 inhibited MBL-mediated neutralization of DENV. We speculated that MBL binding to DENV prevented the attachment of viruses to cells and in the presence of NS1, the protein competitively bound MBL and thus allowed DENV to bind and enter target cells. To test this hypothesis, we incubated BHK cells with DENV in the presence or absence of purified human MBL. After a 2 h incubation at 4°C, cell-bound DENV was measured by real-time RT-PCR for viral genome copies. As expected, MBL efficiently inhibited DENV attachment to BHK cells. However, pre-incubation of MBL with either purified mammalian or insect cell-derived NS1 but not with the control protein BSA increased the binding of DENV to BHK cells (Fig. 5K). Overall, these experiments establish that MBL inhibits DENV infection at the step of virus attachment and that NS1 competitively binds MBL, which facilitates DENV binding and entry to initiate infection.
Detection of soluble NS1 in the saliva of DENV-infected Aedes aegypti mosquitoes
Our data showed that mosquito-cell derived NS1 retained its biological functions, despite being secreted at a lower level than mammalian-cell derived NS1. We reasoned that during blood feeding when DENV is inoculated into the skin of a susceptible host, the presence of NS1 in the saliva of infected mosquitoes could protect DENV from immune recognition, and thus facilitate its spread from the site of inoculation. To demonstrate the relevance of our in vitro findings, we evaluated whether NS1 is present in the saliva of DENV-infected mosquitoes. A total of 1,500 female Aedes aegypti mosquitoes were challenged orally with DENV via an infectious blood meal as previously described (48). Successfully fed mosquitoes (~1,200) with a blood-filled midgut were sorted and subsequently incubated for 14 days until a second non-infectious blood meal was offered. The blood that remained in a container after mosquito feeding was subjected to both viral RNA and NS1 analysis. In addition, salivary glands from 50 mosquitoes were harvested and processed for immunofluorescence staining and Western blotting for DENV NS1. As expected, NS1 was detected in the salivary glands of about 30 to 50% of blood-fed mosquitoes (Fig. 6A-B and data not shown). Along with the DENV genome (Fig. 6C), NS1 was detected in the residual blood after mosquito feeding (Fig. 6D-E) indicating that both DENV and soluble NS1 were present in the saliva of infected mosquitoes and were released during blood feeding.
FIGURE 6. Both DENV and soluble NS1 are released during blood feeding from infected mosquitoes.
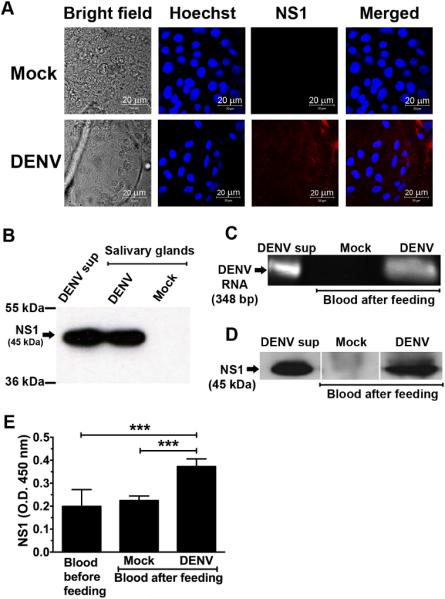
A. Representative bright field and immunofluorescence images of the salivary glands of mock- and DENV-infected mosquitoes. Aedes aegypti female mosquitoes were challenged orally with DENV by feeding them with an infectious blood meal and incubated for 14 days. Salivary glands of mosquitoes were harvested, smeared, and fixed on glass slides. NS1 was detected using a mouse anti-DENV NS1 mAb 1F11 (1 μg/ml) followed by Cy3-conjugated goat anti-mouse IgG. Nuclei were stained with Hoechst, a DNA-specific dye. Salivary glands harvested from uninfected mosquitoes served as a negative control. Analysis was performed by confocal microscopy. B. Expression of NS1 in the salivary glands of DENV-infected mosquitoes was analyzed by Western blotting. Fifty-fold concentrated supernatants from DENV-infected C6/36 cells (DENV sup) and the salivary glands harvested from uninfected mosquitoes (Mock) served as positive and negative controls, respectively. C-D. At 14 days after initial challenge with DENV, a total of 600 to 1200 mosquitoes were fed with a second non-infectious blood meal, the presence of DENV and NS1 in the remaining blood were determined by RT-PCR (C), Western blotting with DENV NS1 specific mAb 1F11 (D) and NS1-ELISA (E), respectively. Error bars indicate SD corresponding to three independent experiments. Asterisks denote the means that are statistically different among indicated groups (*** P < 0.001).
DISCUSSION
It has been reported previously that NS1 is secreted from DENV infected mammalian cells but not from cells of insect origin (4, 18). The claim of detectable soluble NS1 in the culture medium of DENV-infected mosquito cells has been controversial as it could reflect the liberation of intracellular NS1 from dying cells as opposed to an active cellular secretory process (21). Our data provide the following supportive evidence that NS1 is secreted from infected insect cells: 1) Tunicamycin treatment affected extracellular NS1 and virion release but not levels of NS1 protein expression within infected insect cells; 2) Extracellular NS1 derived from infected insect cells appeared to form higher order oligomers, likely hexameric in nature unlike cell-associated NS1, which exhibits different characteristics under native conditions (12); 3) Soluble insect cell-derived NS1 possessed functional characteristics similar to mammalian cell-derived NS1, as it bound to glycosaminoglycans on cell surfaces and inhibited complement activation. We also observed a novel complement antagonism function of NS1 secreted from both mammalian and insect cells as it targets MBL, the major recognition receptor of the lectin pathway of complement activation. Importantly, we showed that soluble NS1 was released together with DENV virions in the context of mosquito blood feeding.
Although NS1 was released from DENV-infected insect cells, concentrations of soluble NS1 in culture media were lower (up to a 10-fold difference) than those observed in infected mammalian cell cultures. The disparity between the two cell types could be attributed to different glycosylation machinery, which affects NS1 processing and secretion efficiency (34). Insect cells lack glycosyltransferases that are needed for synthesis of hybrid and complex-type glycans (40); DENV NS1 produced in insect cells had polymannose-type glycans at both N130 and N207 positions whereas N-linked glycosylation at N130 is further processed to complex-type glycans in mammalian cells (39, 49). In support of the lower levels of NS1 in the supernatants from insect cells, loss of complex-type glycans at N130 results in a decrease in secretion of soluble NS1 (18, 50). Additionally, low levels of NS1 detected in the culture medium of infected insect cells could result from instability of NS1 hexamers resulting in protein aggregation, which we observed when NS1 was subjected to density gradient fractionation and a protein stability test. These results are consistent with a study in which mutation of Asn130 resulted in a loss of hexameric NS1 and an increase in the formation of higher-order NS1 oligomers (34). Indeed, our data agree with recent studies showing that NS1 secreted from DENV-infected C6/36 cells is hexameric in nature (51).
Compared to mammalian-cell derived NS1, soluble NS1 derived from insect cells bound comparably to the plasma membrane via interactions with sulfated glycosaminoglycans like heparan sulfate despite lacking a complex-type glycan at position Asn130 (16). This is consistent with our previous observation that the N-linked glycan at position Asn207 but not at Asn130 was required for cell surface attachment of NS1 (34). NS1 binding back to the cell surface could have an important role in immune evasion through interaction with the plasma complement regulator C4BP (25). Analogous to mammalian-cell derived NS1, recruitment of C4BP to the cell surface by insect cell-derived soluble NS1 attenuated classical and lectin pathway activation through C4BP's cofactor activity, which resulted in C4b degradation by factor I. Insect-cell derived NS1 also interacted with other complement components including C4 and C1s/ProC1s. This could facilitate C4 degradation and protect DENV from C4-mediated neutralization in solution similar to what has been previously described for soluble NS1 from mammalian cells (24).
It has been suggested that the complement system has dual roles in pathogenesis and protection against DENV infection depending on the type of immune response (primary versus secondary infection), disease period (early versus critical phase), and host genetic factors (6, 17, 25). The lectin pathway of complement activation as triggered by the direct interaction of DENV and MBL has been shown to neutralize all four DENV serotypes and thus may control the spread of DENV within a host, especially during a primary infection (28). We described a novel interaction between soluble DENV NS1 and human MBL. NS1 competitively bound to MBL, which decreased DENV recognition by MBL and protected the virus from MBL-mediated neutralization. Mechanistically, without a requirement for further complement activation, MBL directly bound DENV and prevented DENV attachment to target cells. Soluble NS1 competitively inhibited MBL binding, and thus allowed the virus to bind, enter and replicate within susceptible host cells. Consistent with this, binding of MBL to surface glycoproteins of influenza A virus (52), hepatitis C virus (53), and human immunodeficiency virus (54) blocked virus attachment to target cells in a complement activation-independent manner. In WNV infection, however, MBL-mediated neutralization occurs by blocking viral fusion, not cellular attachment (30). Nevertheless, independent interference of MBL with DENV fusion could not be ruled out.
The finding that DENV-infected insect cells could secrete soluble hexameric NS1 that retains complement inhibitory activity prompted us to explore its biological relevance. Detection of soluble NS1 and DENV in a blood meal of DENV-infected Aedes aegypti mosquitoes implies that the mosquito saliva likely contains both soluble NS1 and infectious DENV virions, which could be released simultaneously into the skin and possibly blood during feeding. Complement antagonism functions of soluble NS1 (both insect-cell and mammalian cell-derived) could protect DENV from complement-mediated neutralization, thereby facilitating viral spread either at the earliest stages of infection (during mosquito feeding) or during subsequent replication cycles of DENV in a human host (Fig. 7). First, soluble NS1 forms a tripartite complex with C4 and C1s/C1s proenzyme which results in C4 degradation and decreases complement activation (24) [Pathway 1), Fig. 7]. Second, DENV NS1 binds to C4BP, the major inhibitor of the classical/lectin pathway of complement activation in plasma, to control the activation of the classical and lectin pathways on cell surfaces (25) [Pathway 2), Fig. 7]. Third, a novel interaction between DENV NS1 and human MBL described in this study results in the protection of DENV from MBL-mediated neutralization [Pathway 3), Fig. 7]. However, further studies are required to define sites on the NS1 molecule that interact with complement components and to determine which interactions play more significant roles for DENV evasion in vivo.
FIGURE 7. Model of complement antagonism by DENV NS1.

Together with DENV virions, soluble NS1 is released from DENV-infected insect or mammalian cells. Attenuation of the classical and lectin pathways of complement activation in tissue is promoted by three mechanisms: 1) NS1 binds proC1s and C4 forming a tripartite complex promoting the cleavage of C4 to C4b which is rapidly inactivated by hydrolysis in the fluid phase (C4 degradation), thereby limiting the supply of native C4 (24). 2) NS1 binds and recruits C4BP to the cell surface to restrict the classical and lectin pathway C3 and C5 convertases via the cofactor activity of C4BP on cells (25). 3) NS1 competitively binds MBL and prevents the recognition of DENV by MBL, thereby protecting the virus from MBL-mediated neutralization. Overall, NS1 possesses complement evasion functions and protects DENV and DENV-infected cells from complement activation and inhibition.
NS1 immunization has been demonstrated to have a protective effect against a DENV challenge in animals [reviewed in (5)]. Mechanisms of protection by non-neutralizing anti-NS1 antibodies have been suggested to involve complement-mediated lysis of infected cells (55, 56). It is also possible that NS1 specific antibodies blocking the interaction of NS1 with complement components like C1s, C4BP, and MBL could afford protection. Epitopes on NS1 that confer protection could serve as a basis for future NS1-based vaccine development that circumvent the undesired risk of enhancing antibodies against the prM and E structural proteins.
ACKNOWLEDGEMENTS
We thank all staff members at the Department of Entomology, Armed Forces Research Institute of Medical Sciences (AFRIMS), Bangkok, Thailand for advice on the mosquito experiments. We thank Mr. Kasima Wasuworawong for his help with SDS-PAGE analyses. We also thank Dr. Wichit Suthammarak at the Department of Biochemistry, Faculty of Medicine Siriraj Hospital, Mahidol University for providing materials and advice on native-PAGE analysis and Lee Greenberger (Enzon, Inc.) for providing the purified human MBL protein.
This research project was supported by the Faculty of Medicine Siriraj Hospital, Mahidol University (R015633003), the Thailand Research Fund (P.A.) and NIH grant R01 AI077955 (M.S.D.). P.A. has been supported by a Siriraj Chalermprakiat Grant and a Research Lecturer Grant, Faculty of Medicine Siriraj Hospital, Mahidol University. S.T. is a Ph.D. scholar in the Royal Golden Jubilee Ph.D. Program (PHD/0101/2554).
ABBREVIATIONS
- BHK
baby hamster kidney fibroblast
- C4BP
C4b binding protein
- DENV
dengue virus
- DENV-2 Rep
DENV-2 subgenomic replicon
- DHF/DSS
dengue hemorrhagic fever/dengue shock syndrome
- DVB
dextrose veronal buffered saline
- E
Envelope
- Endo H
endoglycosidase H
- FFA
Focus forming assay
- GVB++
gelatin veronal buffer supplemented with Mg2+ and Ca2+
- GAGs
glycosaminoglycans
- MBL
Mannose blinding lectin
- MFI
mean fluorescence intensity
- MOI
Multiplicity of infection
- NEAA
Non-essential amino acids
- NS1
nonstructural protein 1
- OPD
ortho-phenylene diaminedihydrochloride
- PNGase F
peptide N-glycosidase F
- proC1s
C1s pro-enzyme
- rNS1
recombinant NS1
- S2-NS1 cells
Drosophila S2 cells stably expressing recombinant NS1 of DENV-2
REFERENCES
- 1.WHO . Comprehensive Guidelines for Prevention and Control of Dengue and Dengue Haemorrhagic Fever, Revised and expanded edition ed. WHO Library Cataloguing-in-Publication data, World Health Organization, Regional Office for South-East Asia; 2011. [Google Scholar]
- 2.Bhatt S, Gething PW, Brady OJ, Messina JP, Farlow AW, Moyes CL, Drake JM, Brownstein JS, Hoen AG, Sankoh O, Myers MF, George DB, Jaenisch T, Wint GR, Simmons CP, Scott TW, Farrar JJ, Hay SI. The global distribution and burden of dengue. Nature. 2013;496:504–507. doi: 10.1038/nature12060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Brandt WE, Chiewslip D, Harris DL, Russell PK. Partial purification and characterization of a dengue virus soluble complement-fixing antigen. J Immunol. 1970;105:1565–1568. [PubMed] [Google Scholar]
- 4.Muller DA, Young PR. The flavivirus NS1 protein: molecular and structural biology, immunology, role in pathogenesis and application as a diagnostic biomarker. Antiviral Res. 2013;98:192–208. doi: 10.1016/j.antiviral.2013.03.008. [DOI] [PubMed] [Google Scholar]
- 5.Scaturro P, Cortese M, Chatel-Chaix L, Fischl W, Bartenschlager R. Dengue Virus Non-structural Protein 1 Modulates Infectious Particle Production via Interaction with the Structural Proteins. PLoS Pathog. 2015;11:e1005277. doi: 10.1371/journal.ppat.1005277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Avirutnan P, Punyadee N, Noisakran S, Komoltri C, Thiemmeca S, Auethavornanan K, Jairungsri A, Kanlaya R, Tangthawornchaikul N, Puttikhunt C, Pattanakitsakul SN, Yenchitsomanus PT, Mongkolsapaya J, Kasinrerk W, Sittisombut N, Husmann M, Blettner M, Vasanawathana S, Bhakdi S, Malasit P. Vascular leakage in severe dengue virus infections: a potential role for the nonstructural viral protein NS1 and complement. J Infect Dis. 2006;193:1078–1088. doi: 10.1086/500949. [DOI] [PubMed] [Google Scholar]
- 7.Alcon-LePoder S, Drouet MT, Roux P, Frenkiel MP, Arborio M, Durand-Schneider AM, Maurice M, Le Blanc I, Gruenberg J, Flamand M. The secreted form of dengue virus nonstructural protein NS1 is endocytosed by hepatocytes and accumulates in late endosomes: implications for viral infectivity. J Virol. 2005;79:11403–11411. doi: 10.1128/JVI.79.17.11403-11411.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Chang HH, Shyu HF, Wang YM, Sun DS, Shyu RH, Tang SS, Huang YS. Facilitation of cell adhesion by immobilized dengue viral nonstructural protein 1 (NS1): arginine-glycine-aspartic acid structural mimicry within the dengue viral NS1 antigen. J Infect Dis. 2002;186:743–751. doi: 10.1086/342600. [DOI] [PubMed] [Google Scholar]
- 9.Lin CF, Lei HY, Shiau AL, Liu CC, Liu HS, Yeh TM, Chen SH, Lin YS. Antibodies from dengue patient sera cross-react with endothelial cells and induce damage. J Med Virol. 2003;69:82–90. doi: 10.1002/jmv.10261. [DOI] [PubMed] [Google Scholar]
- 10.Sun DS, King CC, Huang HS, Shih YL, Lee CC, Tsai WJ, Yu CC, Chang HH. Antiplatelet autoantibodies elicited by dengue virus non-structural protein 1 cause thrombocytopenia and mortality in mice. J Thromb Haemost. 2007;5:2291–2299. doi: 10.1111/j.1538-7836.2007.02754.x. [DOI] [PubMed] [Google Scholar]
- 11.Punyadee N, Mairiang D, Thiemmeca S, Komoltri C, Pan-Ngum W, Chomanee N, Charngkaew K, Tangthawornchaikul N, Limpitikul W, Vasanawathana S, Malasit P, Avirutnan P. Microparticles provide a novel biomarker to predict severe clinical outcomes of dengue virus infection. J Virol. 2015;89:1587–1607. doi: 10.1128/JVI.02207-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Gutsche I, Coulibaly F, Voss JE, Salmon J, d'Alayer J, Ermonval M, Larquet E, Charneau P, Krey T, Megret F, Guittet E, Rey FA, Flamand M. Secreted dengue virus nonstructural protein NS1 is an atypical barrel-shaped high-density lipoprotein. Proc Natl Acad Sci U S A. 2011;108:8003–8008. doi: 10.1073/pnas.1017338108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Beatty PR, Puerta-Guardo H, Killingbeck SS, Glasner DR, Hopkins K, Harris E. Dengue virus NS1 triggers endothelial permeability and vascular leak that is prevented by NS1 vaccination. Sci Transl Med. 2015;7:304ra141. doi: 10.1126/scitranslmed.aaa3787. [DOI] [PubMed] [Google Scholar]
- 14.Chen J, Ng MM, Chu JJ. Activation of TLR2 and TLR6 by Dengue NS1 Protein and Its Implications in the Immunopathogenesis of Dengue Virus Infection. PLoS Pathog. 2015;11:e1005053. doi: 10.1371/journal.ppat.1005053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Modhiran N, Watterson D, Muller DA, Panetta AK, Sester DP, Liu L, Hume DA, Stacey KJ, Young PR. Dengue virus NS1 protein activates cells via Toll-like receptor 4 and disrupts endothelial cell monolayer integrity. Sci Transl Med. 2015;7:304ra142. doi: 10.1126/scitranslmed.aaa3863. [DOI] [PubMed] [Google Scholar]
- 16.Avirutnan P, Zhang L, Punyadee N, Manuyakorn A, Puttikhunt C, Kasinrerk W, Malasit P, Atkinson JP, Diamond MS. Secreted NS1 of dengue virus attaches to the surface of cells via interactions with heparan sulfate and chondroitin sulfate E. PLoS Pathog. 2007;3:e183. doi: 10.1371/journal.ppat.0030183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Avirutnan P, Malasit P, Seliger B, Bhakdi S, Husmann M. Dengue virus infection of human endothelial cells leads to chemokine production, complement activation, and apoptosis. J Immunol. 1998;161:6338–6346. [PubMed] [Google Scholar]
- 18.Flamand M, Megret F, Mathieu M, Lepault J, Rey FA, Deubel V. Dengue virus type 1 nonstructural glycoprotein NS1 is secreted from mammalian cells as a soluble hexamer in a glycosylation-dependent fashion. J Virol. 1999;73:6104–6110. doi: 10.1128/jvi.73.7.6104-6110.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Mason PW. Maturation of Japanese encephalitis virus glycoproteins produced by infected mammalian and mosquito cells. Virology. 1989;169:354–364. doi: 10.1016/0042-6822(89)90161-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Leblois H, Young PR. Maturation of the dengue-2 virus NS1 protein in insect cells: effects of downstream NS2A sequences on baculovirus-expressed gene constructs. J Gen Virol. 1995;76(Pt 4):979–984. doi: 10.1099/0022-1317-76-4-979. [DOI] [PubMed] [Google Scholar]
- 21.Ludert JE, Mosso C, Ceballos-Olvera I, del Angel RM. Use of a commercial enzyme immunoassay to monitor dengue virus replication in cultured cells. Virol J. 2008;5:51. doi: 10.1186/1743-422X-5-51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Whitby K, Pierson TC, Geiss B, Lane K, Engle M, Zhou Y, Doms RW, Diamond MS. Castanospermine, a potent inhibitor of dengue virus infection in vitro and in vivo. J Virol. 2005;79:8698–8706. doi: 10.1128/JVI.79.14.8698-8706.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Haider SR, Reid HJ, Sharp BL. Modification of tricine-SDS-PAGE for online and offline analysis of phosphoproteins by ICP-MS. Anal Bioanal Chem. 2010;397:655–664. doi: 10.1007/s00216-010-3588-9. [DOI] [PubMed] [Google Scholar]
- 24.Avirutnan P, Fuchs A, Hauhart RE, Somnuke P, Youn S, Diamond MS, Atkinson JP. Antagonism of the complement component C4 by flavivirus nonstructural protein NS1. J Exp Med. 2010;207:793–806. doi: 10.1084/jem.20092545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Avirutnan P, Hauhart RE, Somnuke P, Blom AM, Diamond MS, Atkinson JP. Binding of flavivirus nonstructural protein NS1 to C4b binding protein modulates complement activation. J Immunol. 2011;187:424–433. doi: 10.4049/jimmunol.1100750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Puttikhunt C, Kasinrerk W, Srisa-ad S, Duangchinda T, Silakate W, Moonsom S, Sittisombut N, Malasit P. Production of anti-dengue NS1 monoclonal antibodies by DNA immunization. J Virol Methods. 2003;109:55–61. doi: 10.1016/s0166-0934(03)00045-4. [DOI] [PubMed] [Google Scholar]
- 27.Hardig Y, Hillarp A, Dahlback B. The amino-terminal module of the C4b-binding protein alpha-chain is crucial for C4b binding and factor I-cofactor function. Biochem J. 1997;323(Pt 2):469–475. doi: 10.1042/bj3230469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Avirutnan P, Hauhart RE, Marovich MA, Garred P, Atkinson JP, Diamond MS. Complement-mediated neutralization of dengue virus requires mannose-binding lectin. MBio. 2011;2 doi: 10.1128/mBio.00276-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Gentry MK, Henchal EA, McCown JM, Brandt WE, Dalrymple JM. Identification of distinct antigenic determinants on dengue-2 virus using monoclonal antibodies. Am J Trop Med Hyg. 1982;31:548–555. doi: 10.4269/ajtmh.1982.31.548. [DOI] [PubMed] [Google Scholar]
- 30.Fuchs A, Lin TY, Beasley DW, Stover CM, Schwaeble WJ, Pierson TC, Diamond MS. Direct complement restriction of flavivirus infection requires glycan recognition by mannose-binding lectin. Cell Host Microbe. 2010;8:186–195. doi: 10.1016/j.chom.2010.07.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Yenchitsomanus PT, Sricharoen P, Jaruthasana I, Pattanakitsakul SN, Nitayaphan S, Mongkolsapaya J, Malasit P. Rapid detection and identification of dengue viruses by polymerase chain reaction (PCR). Southeast Asian J Trop Med Public Health. 1996;27:228–236. [PubMed] [Google Scholar]
- 32.Kuno G, Gubler DJ, Velez M, Oliver A. Comparative sensitivity of three mosquito cell lines for isolation of dengue viruses. Bull World Health Organ. 1985;63:279–286. [PMC free article] [PubMed] [Google Scholar]
- 33.Whiteman MC, Wicker JA, Kinney RM, Huang CY, Solomon T, Barrett AD. Multiple amino acid changes at the first glycosylation motif in NS1 protein of West Nile virus are necessary for complete attenuation for mouse neuroinvasiveness. Vaccine. 2011;29:9702–9710. doi: 10.1016/j.vaccine.2011.09.036. [DOI] [PubMed] [Google Scholar]
- 34.Somnuke P, Hauhart RE, Atkinson JP, Diamond MS, Avirutnan P. N-linked glycosylation of dengue virus NS1 protein modulates secretion, cell-surface expression, hexamer stability, and interactions with human complement. Virology. 2011;413:253–264. doi: 10.1016/j.virol.2011.02.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Butters TD, Hughes RC. Effects of tunicamycin on insect cells. Biochem Soc Trans. 1980;8:170–171. doi: 10.1042/bst0080170. [DOI] [PubMed] [Google Scholar]
- 36.Metz SW, Geertsema C, Martina BE, Andrade P, Heldens JG, van Oers MM, Goldbach RW, Vlak JM, Pijlman GP. Functional processing and secretion of Chikungunya virus E1 and E2 glycoproteins in insect cells. Virol J. 2011;8:353. doi: 10.1186/1743-422X-8-353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Gruters RA, Neefjes JJ, Tersmette M, de Goede RE, Tulp A, Huisman HG, Miedema F, Ploegh HL. Interference with HIV-induced syncytium formation and viral infectivity by inhibitors of trimming glucosidase. Nature. 1987;330:74–77. doi: 10.1038/330074a0. [DOI] [PubMed] [Google Scholar]
- 38.Mehta A, Block TM, Dwek RA. The role of N-linked glycosylation in the secretion of hepatitis B virus. Adv Exp Med Biol. 1998;435:195–205. doi: 10.1007/978-1-4615-5383-0_19. [DOI] [PubMed] [Google Scholar]
- 39.Pryor MJ, Wright PJ. Glycosylation mutants of dengue virus NS1 protein. J Gen Virol. 1994;75(Pt 5):1183–1187. doi: 10.1099/0022-1317-75-5-1183. [DOI] [PubMed] [Google Scholar]
- 40.Altmann F, Staudacher E, Wilson IB, Marz L. Insect cells as hosts for the expression of recombinant glycoproteins. Glycoconj J. 1999;16:109–123. doi: 10.1023/a:1026488408951. [DOI] [PubMed] [Google Scholar]
- 41.Tarentino AL, Maley F. Purification and properties of an endo-beta-N-acetylglucosaminidase from Streptomyces griseus. J Biol Chem. 1974;249:811–817. [PubMed] [Google Scholar]
- 42.Tarentino AL, Gomez CM, Plummer TH., Jr. Deglycosylation of asparagine-linked glycans by peptide:N-glycosidase F. Biochemistry. 1985;24:4665–4671. doi: 10.1021/bi00338a028. [DOI] [PubMed] [Google Scholar]
- 43.Maley F, Trimble RB, Tarentino AL, Plummer TH., Jr. Characterization of glycoproteins and their associated oligosaccharides through the use of endoglycosidases. Anal Biochem. 1989;180:195–204. doi: 10.1016/0003-2697(89)90115-2. [DOI] [PubMed] [Google Scholar]
- 44.Limon-Flores AY, Perez-Tapia M, Estrada-Garcia I, Vaughan G, Escobar-Gutierrez A, Calderon-Amador J, Herrera-Rodriguez SE, Brizuela-Garcia A, Heras-Chavarria M, Flores-Langarica A, Cedillo-Barron L, Flores-Romo L. Dengue virus inoculation to human skin explants: an effective approach to assess in situ the early infection and the effects on cutaneous dendritic cells. Int J Exp Pathol. 2005;86:323–334. doi: 10.1111/j.0959-9673.2005.00445.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Brown AN, Kent KA, Bennett CJ, Bernard KA. Tissue tropism and neuroinvasion of West Nile virus do not differ for two mouse strains with different survival rates. Virology. 2007;368:422–430. doi: 10.1016/j.virol.2007.06.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Lim PY, Behr MJ, Chadwick CM, Shi PY, Bernard KA. Keratinocytes are cell targets of West Nile virus in vivo. J Virol. 2011;85:5197–5201. doi: 10.1128/JVI.02692-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Surasombatpattana P, Hamel R, Patramool S, Luplertlop N, Thomas F, Despres P, Briant L, Yssel H, Misse D. Dengue virus replication in infected human keratinocytes leads to activation of antiviral innate immune responses. Infect Genet Evol. 2011;11:1664–1673. doi: 10.1016/j.meegid.2011.06.009. [DOI] [PubMed] [Google Scholar]
- 48.Lambrechts L, Paaijmans KP, Fansiri T, Carrington LB, Kramer LD, Thomas MB, Scott TW. Impact of daily temperature fluctuations on dengue virus transmission by Aedes aegypti. Proc Natl Acad Sci U S A. 2011;108:7460–7465. doi: 10.1073/pnas.1101377108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Falgout B, Markoff L. Evidence that flavivirus NS1-NS2A cleavage is mediated by a membrane-bound host protease in the endoplasmic reticulum. J Virol. 1995;69:7232–7243. doi: 10.1128/jvi.69.11.7232-7243.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Muylaert IR, Chambers TJ, Galler R, Rice CM. Mutagenesis of the N-linked glycosylation sites of the yellow fever virus NS1 protein: effects on virus replication and mouse neurovirulence. Virology. 1996;222:159–168. doi: 10.1006/viro.1996.0406. [DOI] [PubMed] [Google Scholar]
- 51.Alcala AC, Medina F, Gonzalez-Robles A, Salazar-Villatoro L, Fragoso-Soriano RJ, Vasquez C, Cervantes-Salazar M, Del Angel RM, Ludert JE. The dengue virus non-structural protein 1 (NS1) is secreted efficiently from infected mosquito cells. Virology. 2015;488:278–287. doi: 10.1016/j.virol.2015.11.020. [DOI] [PubMed] [Google Scholar]
- 52.Kase T, Suzuki Y, Kawai T, Sakamoto T, Ohtani K, Eda S, Maeda A, Okuno Y, Kurimura T, Wakamiya N. Human mannan-binding lectin inhibits the infection of influenza A virus without complement. Immunology. 1999;97:385–392. doi: 10.1046/j.1365-2567.1999.00781.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Brown KS, Keogh MJ, Owsianka AM, Adair R, Patel AH, Arnold JN, Ball JK, Sim RB, Tarr AW, Hickling TP. Specific interaction of hepatitis C virus glycoproteins with mannan binding lectin inhibits virus entry. Protein Cell. 2010;1:664–674. doi: 10.1007/s13238-010-0088-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Ezekowitz RA, Kuhlman M, Groopman JE, Byrn RA. A human serum mannose-binding protein inhibits in vitro infection by the human immunodeficiency virus. J Exp Med. 1989;169:185–196. doi: 10.1084/jem.169.1.185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Schlesinger JJ, Brandriss MW, Walsh EE. Protection against 17D yellow fever encephalitis in mice by passive transfer of monoclonal antibodies to the nonstructural glycoprotein gp48 and by active immunization with gp48. J Immunol. 1985;135:2805–2809. [PubMed] [Google Scholar]
- 56.Henchal EA, Henchal LS, Schlesinger JJ. Synergistic interactions of anti-NS1 monoclonal antibodies protect passively immunized mice from lethal challenge with dengue 2 virus. J Gen Virol. 1988;69(Pt 8):2101–2107. doi: 10.1099/0022-1317-69-8-2101. [DOI] [PubMed] [Google Scholar]