

Abstract
Purpose of Review
This study aims to describe bone marrow fat changes in diabetes and to discuss the potential role of marrow fat in skeletal fragility.
Recent Findings
Advances in non-invasive imaging have facilitated marrow fat research in humans. In contrast to animal studies which clearly demonstrate higher levels of marrow fat in diabetes, human studies have shown smaller and less certain differences. Marrow fat has been reported to correlate with A1c, and there may be a distinct marrow lipid saturation profile in diabetes.
Summary
Greater marrow fat is associated with impaired skeletal health. Marrow fat may be a mediator of skeletal fragility in diabetes. Circulating lipids, growth hormone alterations, visceral adiposity, and hypoleptinemia have been associated with greater marrow fat and may represent potential mechanisms for the putative effects of diabetes on marrow fat, although other factors likely contribute. Additional research is needed to further define the role of marrow fat in diabetic skeletal fragility and to determine whether marrow fat is a therapeutic target.
Keywords: Diabetes, Bone marrow fat, Osteoporosis, Marrow adipose tissue
Introduction
Diabetes affects 382 million people worldwide, including 24 million people in the USA, or 10.9 % of the American population [1]. There is increasing recognition that compromised bone health is a complication of diabetes. The skeletal consequences are becoming more apparent as life expectancy for those with type 1 diabetes mellitus (T1DM) improves and prevalence of type 2 diabetes mellitus (T2DM) continues to rise. Epidemiologic studies demonstrate that those with diabetes are at higher fracture risk; hip fracture risk is elevated in T1DM (RR = 6.94, 95 % confidence interval (CI), 3.25–14.78) and T2DM (RR = 1.38, 95 % CI, 1.25–1.53) compared with those without diabetes [2]. Bone mineral density (BMD) is generally lower in T1DM; in contrast, those with T2DM have higher BMD than controls, yet they still have skeletal fragility. Other features of impaired bone health in diabetes, which have been recently reviewed [3], include reduced bone turnover [4] (particularly bone formation), impaired microarchitecture [5, 6], and decreased bone material strength in T2DM at a given BMD [7].
Although BMD is a valuable predictor of fracture in the non-diabetic population, BMD may not reflect true fracture risk in those with diabetes. Predicted hip fracture risk in T1DM based on BMD alone is significantly lower than the observed risk [2]. Similarly, T scores underestimate fracture risk in older adults with T2DM [8]. There are clearly factors not reflected by BMD that contribute to fracture risk, such as microvascular and macrovascular complications resulting in increased falls, although there is evidence that even after adjustment for falls fracture risk is still higher [9]. Other non-BMD-related risk factors include non-enzymatic glycation leading to impaired bone matrix properties [10] and increased cortical porosity [5, 6]. Identifying these non-BMD-related risk factors will be crucial for preventing clinically and economically costly fractures in this large and vulnerable population.
Recently bone marrow fat has been identified as a potential marker or mechanism for diabetes-related skeletal fragility. Traditionally marrow fat was thought to be an inert fat depot, but marrow fat is now known to be an endocrine organ with local and systemic effects. Marrow fat expands in response to metabolic conditions such as anorexia nervosa, aging, estrogen deficiency, glucocorticoid use, and growth hormone deficiency [11•], and there is evidence that marrow fat contributes to circulating adiponectin during caloric restriction [12]. Anatomically, marrow fat comprises approximately 70 % of the bone marrow, about 1 kg in weight, or 8 % of total fat mass, with marrow fat more predominant in the appendicular than axial skeleton [13]. The function of marrow fat is unclear, although it is postulated to play a role in lipid storage, skeletal remodeling, metabolic homeostasis, hematopoietic regulation, mechanical function, and thermogenesis [13].
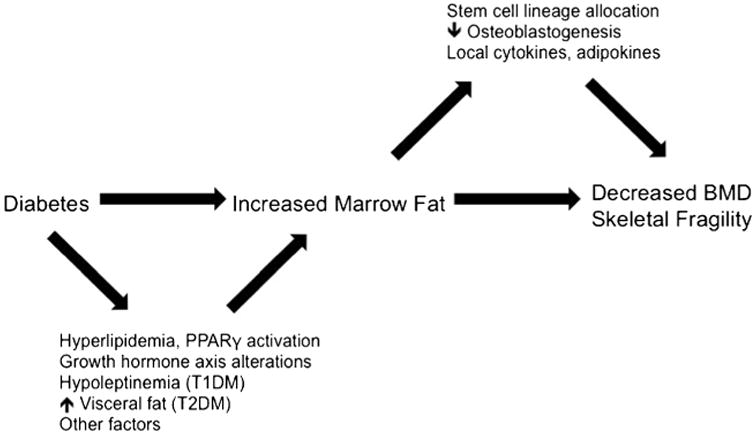
This review discusses marrow fat characteristics in diabetes, with a focus on potential mechanisms linking diabetes and marrow fat (Fig. 1). Data are summarized from animal and human studies of T1DM and T2DM that measured bone marrow adiposity and studies that examined potential mechanisms for differences in marrow fat by diabetes status. PubMed articles written in the English language were reviewed through July 2016, using the search terms “diabetes mellitus,” “bone marrow fat,” “bone marrow adiposity,” “bone marrow adipose tissue,” and “bone marrow adipocytes.” References from the retrieved articles were also used.
Fig. 1. Proposed model of increased marrow fat in diabetes.
Non-invasive Assessments of Marrow Fat
Insights into marrow fat have been advanced in part due to improvements in non-invasive imaging. Traditionally, measurements were performed histologically with bone biopsies, but now marrow fat is generally measured with magnetic resonance (MR) modalities. Two commonly used MR techniques are proton magnetic resonance spectroscopy (1H-MRS) and T1-weight MR imaging (MRI). In 1H-MRS, a single voxel is placed in the area of interest, and the water proton peak and saturated lipid proton peak are measured. Marrow fat is calculated as a percentage, rather than a total volume. Unfortunately, there is not a standardized equation used for calculation, with investigators variably reporting the lipid/water ratio or the fat fraction, calculated as lipid/(lipid + water). Thus, it can be challenging to compare marrow fat values between studies. Also, marrow fat values are location dependent, and while most human studies measure marrow fat at the lumbar vertebrae, other sites include the femur, tibia, and pelvis. There are also variations within sites, for example, marrow fat significantly increases from L1 to L4 [14]. T1-weight MRI is not limited to a small area defined by a voxel, although it is semi-quantitative and may also have errors due to partial volume effects [15]. At this time, MR assessment of marrow fat is not performed clinically.
Marrow Fat Is Negatively Associated with Bone
Bone and marrow fat are intimately related within the bone microenvironment, and greater marrow fat is associated with lower BMD and increased skeletal fragility [11•, 15–17]. The negative association between marrow fat and bone has been seen in both sexes and in various ethnicities, as well as in conditions such as aging, osteoporosis, starvation, and mechanical unloading. Imaging-based findings have been confirmed by histology from bone biopsies [18, 19]. Although most studies are cross-sectional, a longitudinal study demonstrated that higher marrow fat predicted bone loss in postmenopausal women. Griffith et al. measured baseline MRS and perfusion imaging at the hip, with hip dual-energy X-ray absorptiometry (DXA) at baseline and after 2 and 4 years [20]. In women with lower than median femoral neck marrow fat content, BMD declined 1.6 %, compared with 4.7 % in those with higher than median marrow fat content. Other longitudinal studies show that marrow fat decreases with osteoporosis treatments, such as transdermal estrogen [21] and risedronate [22], providing additional support for an inverse relationship between marrow fat and bone. Marrow fat also correlates with prevalent fracture [19], independent of BMD [23••, 24], suggesting that marrow fat relates not only to bone mass but also to additional contributors to skeletal strength. Whether marrow fat changes precede or follow bone changes is not clear, as animal models have demonstrated both sequences [16]. Given observations that marrow fat and bone both peak in early adulthood, there are likely complex interactions between marrow fat and bone that remain unknown.
Mechanistically, marrow fat may negatively affect bone by creating an imbalance between adipogenesis and osteoblastogenesis. Bone and fat cells share a common mesenchymal stem cell precursor within the marrow, and differentiation favoring adipogenesis could occur at the expense of osteoblastogenesis. This mechanism for bone loss has been implicated in aging and thiazolidinedione use. Adipogenesis is regulated by several transcriptional factors, and peroxisome proliferator-activator receptor-γ (PPARγ) is thought to be of particular importance [25]. After differentiation, marrow adipocytes may directly influence bone metabolism. For example, upon culture with marrow adipocytes, osteoblasts express adipogenic messenger RNA (mRNA) [26], and fatty acids can inhibit osteoblast cell proliferation [27]. Additionally gene expression profiling of marrow adipocytes demonstrated increased expression of bone influencing factors compared with epididymal adipocytes, such as tumor necrosis factor alpha, transforming growth factor beta 1, and interleukin 6 [28].
Studies of Marrow Fat in Diabetes
Animal Studies: Higher Marrow Fat in Diabetes
T1DM
Animal models of T1DM have consistently demonstrated higher marrow adiposity and lower bone mass than controls. For example, in a study of two mouse models of T1DM, streptozotocin-induced diabetic mice had an 8-fold increase in tibial marrow adipocytes compared with controls, and non-obese diabetic (NOD) mice had a 3–4-fold increase in marrow adipocytes compared with controls [29]. Markers of adipocyte differentiation including PPARγ2 and adipocyte fatty acid-binding protein 2 were elevated. The diabetic models had significant trabecular bone loss, and cortical bone loss was present although not statistically significant. Interestingly, while earlier markers of osteoblast differentiation were similar to controls, there was a significant decrease in osteocalcin mRNA, a marker of mature osteoblasts, suggesting that later stages of osteoblast maturation may be influenced by T1DM. The effect of diabetes may be location dependent, as increased marrow adiposity in T1DM has been demonstrated in mouse femora and calvaria but not vertebrae [30]. The reciprocal relationship between bone and marrow fat has also been observed in long bones during bone formation; in a model of distraction osteogenesis, streptozotocin-induced T1DM mice displayed increased adipogenesis and decreased osteoblastogenesis compared with controls [31].
T2DM
Although there are fewer studies of marrow fat in animal models of T2DM than T1DM, marrow fat appears similarly elevated. In a model of early-onset T2DM, male TALLYHO/JngJ mice had high tibial marrow adiposity [32••]. Compared with control mice, Tallyho mice had a 50-and 7-fold increase in marrow fat at ages 8 and 17 weeks, respectively, with severe deficits in distal femur trabecular bone volume fraction, trabecular number, and connectivity density. It is unclear whether these differences were specifically due to T2DM, because these mice also had other metabolic abnormalities that might increase marrow fat, such as hyperlipidemia. In vitro, marrow stem cell differentiation into osteoblasts or adipocytes was similar in Tallyho and control mice, suggesting that the skeletal deficiencies were not due to stem cell lineage allocation and were more likely due to consequences of diabetes.
Human Studies: Smaller and Less Certain Increases in Marrow Fat
T1DM
In contrast to animal models, most human studies of T1DM do not show a significant elevation in marrow fat. However, some studies show a non-significant increase compared with controls. The largest study to examine marrow fat in T1DM was a cross-sectional study of 30 young women with T1DM, with median age of 22 years, body mass index (BMI) at 25 kg/m2, and hemoglobin A1c at 9.8 % [33•]. Median L3 marrow fat fraction by 1H-MRS was 31.1 % in those with T1DM, compared with 26.3 % in healthy controls (p = 0.20). MRI at the proximal tibia demonstrated lower apparent bone volume/total volume and apparent trabecular number, and greater apparent trabecular spacing.
In a cross-sectional study of T1DM, Slade et al concluded that marrow fat levels were similar between diabetic participants and controls [34]. Eight men and eight women with T1DM, with mean A1c at 7.7 % and average age of 37 years (men) and 40 years (women), were compared with age- and BMI-matched controls. Marrow fat levels by MRI at the L4 vertebra, distal femur, and proximal tibia were similar between groups. For example, at the L4 vertebra, mean marrow fat level in diabetic men was 53.7 vs. 57.3 % for control men; in women, mean marrow fat level was 55.4 vs. 52.3 %. There was no correlation between marrow fat and A1c or disease duration in those with T1DM. The marrow fat sites had good correlation with one another (e.g., r = 0.411, p < 0.05 for vertebra and tibia). BMD by DXA at the femoral neck and hip correlated with femoral marrow fat, although vertebral BMD and marrow fat were not correlated.
T2DM
Some human studies demonstrate slightly higher levels of marrow fat in diabetic participants vs. controls, although these findings are often not statistically significant. This may reflect small sample sizes, because the largest study of marrow fat in T2DM did show a statistically significant difference. In the Osteoporotic Fracture in Men Study, 156 men aged 74–96 years, 24 % with T2DM, had lumbar spine marrow fat measured by 1H-MRS. In age-adjusted analysis, T2DM was associated with higher marrow fat (59 %) compared with controls (55 %, p = 0.03) [35]. Marrow fat positively correlated with bone formation and resorption markers, suggesting that in older men, higher marrow fat was associated with diabetes and higher bone turnover.
Two smaller studies have also suggested higher marrow fat levels in T2DM, but without statistical significance for a difference. Baum et al. studied 26 postmenopausal women without osteoporosis, 13 with T2DM and 13 age- and BMI-matched controls. Women with T2DM had mean age of 59 years, BMI at 27 kg/m2, and A1c at 7.6 %. Those with T2DM had higher L1–L3 marrow fat content by 1H-MRS (69.3 %) compared with the controls (67.5 %, p = 031) [36]. Importantly, within the diabetic women, those with higher A1c had higher marrow fat levels (r = 0.83, p < 0.01), supporting the hypothesis of an association between diabetes and marrow adiposity. Among all participants, those with higher marrow fat had lower spinal BMD (r = −0.45, p = 0.03).
Our group conducted a small pilot prospective study of marrow fat changes after Roux-en-Y gastric bypass (RYGB) surgery for morbid obesity, which suggested a possible difference in marrow fat by T2DM status. Six of eleven women had T2DM, with mean age of 49 years, BMI at 39 kg/m2, and A1c at 7.2 %; they were compared post hoc with the five non-diabetic women undergoing RYGB. The diabetic women were older and more likely to be postmenopausal, although differences were not statistically significant. Preoperatively those with T2DM had 54.2 % marrow fat content at the L3–L4 vertebrae by 1H-MRS, compared with 43.1 % in those without T2DM (p = 0.25) [37•]. There was no correlation between marrow fat and A1c in diabetic women, although diabetes medication use varied. In the entire group, marrow fat and vertebral volumetric BMD were inversely correlated, so those with higher marrow fat had lower BMD (r = −0.76, p < 0.01). Interestingly, 6 months after RYGB with a mean at 27 kg weight loss, there appeared to be a difference in marrow fat change between those with and without T2DM. Those with T2DM had a decline of −7.5 ± 7.3 % (p = 0.05), whereas mean change in those without T2DM was +0.9 ± 8.8 % (p = 0.84; p value between groups, 0.12). These findings suggest that marrow fat behavior during profound weight loss could differ by diabetes status. Bariatric surgery may be a useful model for studying marrow fat, as there is dramatic improvement or resolution of T2DM, accompanied by striking changes in potential mediators of marrow fat.
Patsch et al. measured L1–L3 marrow fat content by H-MRS in 69 diabetic and non-diabetic postmenopausal women with and without a history of fragility fracture [38••]. A1c was 7.6 % in diabetic women without fracture and 7.8 % in diabetic women with fracture. In a regression model adjusted for age, race, and spinal BMD by QCT, there was not an association between vertebral marrow fat content and diabetes (β = 0.02, p = 0.27). Interestingly, bone marrow fat composition was linked with fragility fracture and diabetes, as discussed later in this review.
There are limited data for those with pre-diabetes, but a study by de Paula et al. suggested a potential correlation between dysglycemia and marrow fat [39•]. L3 vertebral fat content was measured by 1H-MRS in 30 women, 11 of whom had pre-diabetes, with mean age of 47.8 years, BMI at 25.5 kg/m2, and A1c at 5.5 %. In the entire group, marrow fat content was 29.3 %, and marrow fat was positively associated with A1c and fasting blood glucose, although not with insulin level or the homeostatic model assessment of insulin resistance.
Human studies of marrow fat in T2DM have explored the role of marrow fat composition, which has not been examined to date in animal models or studies of T1DM. Imaging with 3 Tesla 1H-MRS can provide enhanced spectral resolution, so that unsaturated and residual lipids can be measured in addition to the standard saturated lipids. The fraction of lipids that are unsaturated vs. saturated can then be determined. Baum et al. found in their study of T2DM, the mean vertebral unsaturated lipid fraction was significantly lower in the diabetic group (6.7 ± 1.0 %) compared with the age- and BMI-matched control group (7.9 ± 1.6 %, p < 0.05) [36]. Similarly, Patsch et al. used the same calculations for lipid peaks and reported that in their participants, after adjustment for age, race, and spinal volumetric BMD, diabetes was associated with a 1.3 % lower unsaturation level (p = 0.018) and 3.3 % higher saturation level (p = 0.004) [38••]. Interestingly, there was also a relationship between marrow fat composition and fracture. With similar adjustments, the prevalence of fragility fracture was associated with a 1.7 % lower unsaturation level (p = 0.005) and 2.9 % higher saturation levels (p = 0.017). Diabetic women with fractures had the lowest marrow unsaturation and highest saturation levels. The relationship between lower unsaturated marrow fat levels and decreased BMD has also been reported in large study of non-diabetic women [40]. This may support the Women's Health Initiative study findings of an association between higher dietary unsaturated fatty acid intake and lower fracture risk, and an association between higher saturated fatty acid intake and higher fracture risk [41]. Additional research should further examine whether marrow fat composition plays a role in skeletal fragility in diabetes.
Comparison of Human with Animal Model Findings
Why is marrow fat increased several fold in animal models of diabetes, whereas in human studies, elevations in marrow fat levels are smaller and less certain? Physiologic differences between mice and humans may explain some of the discrepancy. Mice and other small animals have proportionally less marrow fat compared with humans [13]. Further, animal models also have more severe forms of diabetes; for example, mice are confirmed to have T1DM when blood glucose is >300 mg/dl, and these animals are not treated with insulin. Anti-diabetic treatments may diminish findings in humans, and yet a human study of severe untreated diabetes would not be ethical. Additionally, most animal studies measure marrow fat in the long bones, whereas human studies tend to measure vertebral marrow fat. Finally, differences in the fraction of lipids that are unsaturated vs. saturated between those with and without diabetes could introduce variability that obscures differences in overall marrow fat content.
Potential Mechanism of Increased Marrow Fat in Diabetes
It is unclear why marrow fat may increase in diabetes. In T1DM, marrow fat may paradoxically increase in response to global substrate deficiency, as seen in caloric restriction and anorexia [16]. In T2DM, dysfunctional adipocytes and a globally adipogenic state may contribute to increased marrow adiposity. Although T1DM and T2DM have clear differences in pathophysiology and consequences, we will discuss specific potential mechanisms for increased marrow fat in the two diseases here, noting when a potential mechanism applies to just one disease or is shared. We focus on possible causal mechanisms and acknowledge that there may be other factors at play not discussed in this review, including oxidative stress [42], adiponectin, glucocorticoids, and sex hormones.
Hyperlipidemia and PPARγ
In diabetes, increased free fatty acids and triglycerides activate PPARγ, which is shown to promote adipocyte differentiation in the marrow. Slade et al. reported a positive correlation of serum lipid levels with vertebral and tibial marrow fat in humans with T1DM [34]. This relationship has also been seen in larger studies of non-diabetic participants and was found to be independent of age, BMI, insulin resistance, and exercise status [43].
PPARγ promotion of marrow adipogenesis and skeletal fragility in diabetes is an appealing proposed mechanism, given the association between fractures and thiazolidinediones [44], which are PPARγ agonists. A randomized controlled trial (RCT) of 20 participants with T2DM found that 6 months of pioglitazone treatment increased vertebral and proximal femoral marrow fat [45]. There was no association between changes in marrow fat and areal BMD, although 6 months may have been too short to see skeletal changes. Effects of thiazolidinediones on marrow fat may be drug and disease specific; in a different RCT of 53 non-diabetic postmenopausal women on rosiglitazone for 14 weeks, vertebral marrow fat actually decreased [46], although animal studies with rosiglitazone demonstrate an increase in marrow fat [47, 48].
If PPARγ agonism increases marrow fat and decreases bone mass, then PPARγ insufficiency or antagonism would be expected to decrease marrow fat and increase bone mass. Indeed, mice with PPARγ haploinsufficiency have fewer marrow adipocytes and enhanced bone mass compared with controls [49]. In T1DM mice treated with a PPARγ antagonist, bisphenol-A-diglycidyl ether (BADGE), BADGE treatment blocked diabetes-induced hyperlipidemia and marrow fat increase; however, there was still bone loss [50]. These findings might suggest that increased marrow fat is not linked to diabetic bone loss, although perhaps the length of PPARγ antagonism was not long enough or BADGE did not allow mesenchymal stem cells to switch to osteoblast differentiation [51].
Growth Hormone and Insulin-Like Growth Factor 1
Growth hormone (GH) stimulates bone formation and lipolysis and may play a role in marrow fat regulation. For example, in rats subjected to hypophysectomy, there was increased marrow fat and decreased bone growth, which reversed only with GH replacement and not with insulin-like growth factor 1 (IGF-1), estradiol, thyroxine, or cortisone [52]. The relationship between the GH axis and marrow fat has also been observed in non-diabetic humans. In a cross-sectional study of obese premenopausal women, vertebral marrow fat inversely correlated with IGF-1, independent of age and BMI [53]. Diabetes results in alterations to the GH axis. Adolescents with T1DM have been shown to have low IGF-1 levels despite GH elevations [54, 55]. Effects of T2DM on the GH axis are less clear, although obesity—often accompanying T2DM—is thought to blunt the axis [56].
Few studies address both the GH axis and marrow fat in the setting of diabetes. Abdalrahaman et al. did not find a significant correlation between IGF-1 and vertebral marrow fat in those with T1DM [33•]. While a plausible hypothesis, there is currently insufficient evidence that the GH axis meaningfully influences marrow adiposity in those with diabetes.
Increased Visceral Fat
Common mechanisms could regulate marrow fat and visceral fat depots. In those without diabetes, there appears to be a positive association between visceral fat and marrow fat. In premenopausal women, greater visceral fat correlated with higher vertebral marrow fat content, independent of BMD [53]. Similarly, in a study of women aged 18–88 years, visceral fat positively correlated with pelvic marrow fat [57]. In contrast, in a study of young healthy men and women, visceral fat did not correlate with marrow fat in the femoral shaft [58], suggesting that the relationship between visceral and marrow fat may depend on age as well as location of marrow fat. In T2DM, increased visceral fat is common, whereas visceral fat may not be increased in T1DM. In studies of marrow fat in diabetes, most studies that also examine body composition demonstrate a positive association between marrow and visceral fat [36] or a trend towards a positive association [33•, 37•]; these interestingly include a study of T1DM.
Hypoleptinemia
Leptin is a hormone secreted by adipocytes that plays an important role in food intake, energy expenditure, fat metabolism, as well as bone metabolism. Hypoleptinemia could represent a mechanism for effects of T1DM on marrow adiposity, as hypoleptinemia is prevalent in T1DM [59] and in other states of increased marrow fat [52]. Also, the T1DM bone and marrow fat phenotype is similar to that of leptin-deficient ob/ob mice [60]. In ob/ob mice, leptin administered centrally [61] or peripherally [62] decreased marrow adipogenesis and increased osteogenesis. However, these studies used supraphysiological doses of leptin [13], and other studies have demonstrated a dissociation between leptin levels and marrow fat changes [63]. For example, caloric restriction in rabbits resulted in hypoleptinemia, but marrow fat expansion did not occur [64]. Few data exists about leptin and marrow fat in diabetic animals. In a study of streptozotocin-induced T1DM mice receiving chronic subcutaneous leptin infusions, leptin prevented the diabetes-associated marrow fat increase [65]. However, diabetes-associated bone loss was not prevented, suggesting that leptin deficiency may only be responsible for increased marrow fat, which is not required for bone loss in T1DM.
Conclusions
Skeletal fragility in diabetes is a growing concern. Because BMD may not adequately predict fracture risk in those with diabetes, there is interest in other markers and mediators of bone health, including bone marrow fat. Greater marrow fat is associated with lower BMD and compromised bone strength. Marrow fat is clearly elevated in animal models of diabetes. In humans, the relationship with marrow fat and diabetes is less clear, although marrow fat has been shown to correlate with A1c and may have a different saturated and unsaturated marrow fat composition. Circulating lipids, PPARγ, GH axis alterations, visceral adiposity, and hypoleptinemia are candidate mechanisms for the putative effects of diabetes on marrow fat; other factors likely play roles as well. Additional research is needed to define the role of marrow fat in diabetic skeletal fragility and ultimately to determine whether it is a therapeutic target.
Acknowledgments
T. Kim has been supported by a National Institute of Diabetes, Digestive and Kidney Diseases (NIDDK), National Institutes of Health (NIH) training grant (5T32 DK007418-35) and the Wilsey Family Foundation. A. Schafer has been supported by the Department of Veterans Affairs, Veterans Health Administration, Clinical Science Research and Development Service, Career Development Award-2 (5 IK2 CX000549); the NIDDK, NIH (R01 DK107629); and the American Society for Bone and Mineral Research (Junior Faculty Osteoporosis Clinical Research Award). The authors thank Robert Nissenson for his expert review of the manuscript.
Footnotes
Conflict of Interest: Anne Schafer and Tiffany Kim declare no conflicts of interest.
Compliance with Ethical Standards: Human and Animal Rights and Informed Consent All reported studies by the authors involving human subjects were performed after approval by the appropriate institutional review boards. When required, written informed consent was obtained from all participants.
References
Papers of particular interest, published recently, have been highlighted as:
• Of importance
•• Of major importance
- 1.Guariguata L, Whiting DR, Hambleton I, Beagley J, Linnenkamp U, Shaw JE. Global estimates of diabetes prevalence for 2013 and projections for 2035. Diabetes Res ClinPract. 2014;103(2):137–49. doi: 10.1016/j.diabres.2013.11.002. [DOI] [PubMed] [Google Scholar]
- 2.Vestergaard P. Discrepancies in bone mineral density and fracture risk in patients with type 1 and type 2 diabetes—a meta-analysis. Osteoporos Int. 2007;18(4):427–44. doi: 10.1007/s00198-006-0253-4. [DOI] [PubMed] [Google Scholar]
- 3.Sellmeyer DE, Civitelli R, Hofbauer LC, Khosla S, Lecka-Czernik B, Schwartz AV, et al. Skel metabolism, fracture risk, and fracture outcomes in type 1 and type 2 diabetes. Diabetes. 2016;65(7):1757–66. doi: 10.2337/db16-0063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Starup-Linde J, Vestergaard P. Biochemical bone turnover markers in diabetes mellitus—a systematic review. Bone. 2016;82:69–78. doi: 10.1016/j.bone.2015.02.019. [DOI] [PubMed] [Google Scholar]
- 5.Shanbhogue VV, Hansen S, Frost M, Jorgensen NR, Hermann AP, Henriksen JE, et al. Bone geometry, volumetric density, microarchitecture, and estimated bone strength assessed by HR-pQCT in adult patients with type 1 diabetes mellitus. J Bone Miner Res. 2015;30(12):2188–99. doi: 10.1002/jbmr.2573. [DOI] [PubMed] [Google Scholar]
- 6.Burghardt AJ, Issever AS, Schwartz AV, Davis KA, Masharani U, Majumdar S, et al. High-resolution peripheral quantitative computed tomographic imaging of cortical and trabecular bone microarchitecture in patients with type 2 diabetes mellitus. J Clin Endocrinol Metab. 2010;95(11):5045–55. doi: 10.1210/jc.2010-0226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Farr JN, Drake MT, Amin S, Melton LJ, 3rd, McCready LK, Khosla S. In vivo assessment of bone quality in postmenopausal women with type 2 diabetes. J Bone Miner Res. 2014;29(4):787–95. doi: 10.1002/jbmr.2106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Schwartz AV, Vittinghoff E, Bauer DC, Hillier TA, Strotmeyer ES, Ensrud KE, et al. Association of BMD and FRAX score with risk of fracture in older adults with type 2 diabetes. JAMA. 2011;305(21):2184–92. doi: 10.1001/jama.2011.715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Bonds DE, Larson JC, Schwartz AV, Strotmeyer ES, Robbins J, Rodriguez BL, et al. Risk of fracture in women with type 2 diabetes: the Women's Health Initiative Observational Study. J Clin Endocrinol Metab. 2006;91(9):3404–10. doi: 10.1210/jc.2006-0614. [DOI] [PubMed] [Google Scholar]
- 10.Vashishth D. The role of the collagen matrix in skeletal fragility. Curr Osteoporos Rep. 2007;5(2):62–6. doi: 10.1007/s11914-007-0004-2. [DOI] [PubMed] [Google Scholar]
- 11•.Fazeli PK, Horowitz MC, MacDougald OA, Scheller EL, Rodeheffer MS, Rosen CJ, et al. Marrow fat and bone—new perspectives. J Clin Endocrinol Metab. 2013;98(3):935–45. doi: 10.1210/jc.2012-3634. This recent review of animal and human data examines the behavior of marrow fat and its potential significance in metabolic diseases associated with skeletal fragility, including diabetes. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Cawthorn WP, Scheller EL, Learman BS, Parlee SD, Simon BR, Mori H, et al. Bone marrow adipose tissue is an endocrine organ that contributes to increased circulating adiponectin during caloric restriction. Cell Metab. 2014;20(2):368–75. doi: 10.1016/j.cmet.2014.06.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Scheller EL, Cawthorn WP, Burr AA, Horowitz MC, MacDougald OA. Marrow adipose tissue: trimming the fat. Trends Endocrinol Metab. 2016;27(6):392–403. doi: 10.1016/j.tem.2016.03.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Li X, Kuo D, Schafer AL, Porzig A, Link TM, Black D, et al. Quantification of vertebral bone marrow fat content using 3 Tesla MR spectroscopy: reproducibility, vertebral variation, and applications in osteoporosis. J Magn Reson Imaging. 2011;33(4):974–9. doi: 10.1002/jmri.22489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Schwartz AV. Marrow fat and bone: review of clinical findings. Front Endocrinol (Lausanne) 2015;6:40. doi: 10.3389/fendo.2015.00040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Devlin MJ, Rosen CJ. The bone-fat interface: basic and clinical implications of marrow adiposity. Lancet Diabetes Endocrinol. 2015;3(2):141–7. doi: 10.1016/S2213-8587(14)70007-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Paccou J, Hardouin P, Cotten A, Penel G, Cortet B. The role of bone marrow fat in skeletal health: usefulness and perspectives for clinicians. J Clin Endocrinol Metab. 2015;100(10):3613–21. doi: 10.1210/jc.2015-2338. [DOI] [PubMed] [Google Scholar]
- 18.Verma S, Rajaratnam JH, Denton J, Hoyland JA, Byers RJ. Adipocytic proportion of bone marrow is inversely related to bone formation in osteoporosis. J Clin Pathol. 2002;55(9):693–8. doi: 10.1136/jcp.55.9.693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Justesen J, Stenderup K, Ebbesen EN, Mosekilde L, Steiniche T, Kassem M. Adipocyte tissue volume in bone marrow is increased with aging and in patients with osteoporosis. Biogerontology. 2001;2(3):165–71. doi: 10.1023/a:1011513223894. [DOI] [PubMed] [Google Scholar]
- 20.Griffith JF, Yeung DK, Leung JC, Kwok TC, Leung PC. Prediction of bone loss in elderly female subjects by MR perfusion imaging and spectroscopy. Eur Radiol. 2011;21(6):1160–9. doi: 10.1007/s00330-010-2054-6. [DOI] [PubMed] [Google Scholar]
- 21.Syed FA, Oursler MJ, Hefferanm TE, Peterson JM, Riggs BL, Khosla S. Effects of estrogen therapy on bone marrow adipocytes in postmenopausal osteoporotic women. Osteoporos Int. 2008;19(9):1323–30. doi: 10.1007/s00198-008-0574-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Duque G, Li W, Adams M, Xu S, Phipps R. Effects of risedronate on bone marrow adipocytes in postmenopausal women. Osteoporos Int. 2011;22(5):1547–53. doi: 10.1007/s00198-010-1353-8. [DOI] [PubMed] [Google Scholar]
- 23••.Schwartz AV, Sigurdsson S, Hue TF, Lang TF, Harris TB, Rosen CJ, et al. Vertebral bone marrow fat associated with lower trabecular BMD and prevalent vertebral fracture in older adults. J Clin Endocrinol Metab. 2013;98(6):2294–300. doi: 10.1210/jc.2012-3949. Higher marrow fat was associated with prevalent vertebral fracture in men, independent of BMD. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Wehrli FW, Hopkins JA, Hwang SN, Song HK, Snyder PJ, Haddad JG. Cross-sectional study of osteopenia with quantitative MR imaging and bone densitometry. Radiology. 2000;217(2):527–38. doi: 10.1148/radiology.217.2.r00nv20527. [DOI] [PubMed] [Google Scholar]
- 25.Sadie-Van Gijsen H, Hough FS, Ferris WF. Determinants of bone marrow adiposity: the modulation of peroxisome proliferator-activated receptor-gamma2 activity as a central mechanism. Bone. 2013;56(2):255–65. doi: 10.1016/j.bone.2013.06.016. [DOI] [PubMed] [Google Scholar]
- 26.Clabaut A, Delplace S, Chauveau C, Hardouin P, Broux O. Human osteoblasts derived from mesenchymal stem cells express adipogenic markers upon coculture with bone marrow adipocytes. Differentiation. 2010;80(1):40–5. doi: 10.1016/j.diff.2010.04.004. [DOI] [PubMed] [Google Scholar]
- 27.Maurin AC, Chavassieux PM, Meunier PJ. Expression of PPARgamma and beta/delta in human primary osteoblastic cells: influence of polyunsaturated fatty acids. Calcif Tissue Int. 2005;76(5):385–92. doi: 10.1007/s00223-004-0108-y. [DOI] [PubMed] [Google Scholar]
- 28.Liu LF, Shen WJ, Ueno M, Patel S, Kraemer FB. Characterization of age-related gene expression profiling in bone marrow and epididymal adipocytes. BMC Genomics. 2011;12:212. doi: 10.1186/1471-2164-12-212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Botolin S, McCabe LR. Bone loss and increased bone adiposity in spontaneous and pharmacologically induced diabetic mice. Endocrinology. 2007;148(1):198–205. doi: 10.1210/en.2006-1006. [DOI] [PubMed] [Google Scholar]
- 30.Martin LM, McCabe LR. Type I diabetic bone phenotype is location but not gender dependent. Histochem Cell Biol. 2007;128(2):125–33. doi: 10.1007/s00418-007-0308-4. [DOI] [PubMed] [Google Scholar]
- 31.Fowlkes JL, Bunn RC, Liu L, Wahl EC, Coleman HN, Cockrell GE, et al. Runt-related transcription factor 2 (RUNX2) and RUNX2-related osteogenic genes are down-regulated throughout osteogenesis in type 1 diabetes mellitus. Endocrinology. 2008;149(4):1697–704. doi: 10.1210/en.2007-1408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32••.Devlin MJ, Van Vliet M, Motyl K, Karim L, Brooks DJ, Louis L, et al. Early-onset type 2 diabetes impairs skeletal acquisition in the male TALLYHO/JngJ mouse. Endocrinology. 2014;155(10):3806–16. doi: 10.1210/en.2014-1041. Marrow fat levels were significantly elevated in a mouse model of T2DM. These mice also had reduced BMD and severe deficits in trabecular microarchitecture. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33•.Abdalrahaman N, McComb C, Foster JE, McLean J, Lindsay RS, McClure J, et al. Deficits in trabecular bone microarchitecture in young women with type 1 diabetes mellitus. J Bone Miner Res. 2015;30(8):1386–93. doi: 10.1002/jbmr.2465. Young women with T1DM had slightly higher levels of vertebral marrow fat than nondiabetic controls, and higher marrow fat levels correlated with greater visceral fat. [DOI] [PubMed] [Google Scholar]
- 34.Slade JM, Coe LM, Meyer RA, McCabe LR. Human bone marrow adiposity is linked with serum lipid levels not T1-diabetes. J Diabetes Complications. 2012;26(1):1–9. doi: 10.1016/j.jdiacomp.2011.11.001. [DOI] [PubMed] [Google Scholar]
- 35.Sheu Y, Schwartz AV, Amati F, Goodpaster BH, Li X, Bauer DC, et al. Bone marrow adiposity is elevated in older men with type 2 diabetes. Diabetes. 2012;61:A367. [Google Scholar]
- 36.Baum T, Yap SP, Karampinos DC, Nardo L, Kuo D, Burghardt AJ, et al. Does vertebral bone marrow fat content correlate with abdominal adipose tissue, lumbar spine bone mineral density, and blood biomarkers in women with type 2 diabetes mellitus? J Magn Reson Imaging. 2012;35(1):117–24. doi: 10.1002/jmri.22757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37•.Schafer AL, Li X, Schwartz AV, Tufts LS, Wheeler AL, Grunfeld C, et al. Changes in vertebral bone marrow fat and bone mass after gastric bypass surgery: a pilot study. Bone. 2015;74:140–5. doi: 10.1016/j.bone.2015.01.010. Marrow fat content decreased after gastric bypass surgery in obese women with diabetes, while it was maintained in women without diabetes. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38••.Patsch JM, Li X, Baum T, Yap SP, Karampinos DC, Schwartz AV, et al. Bone marrow fat composition as a novel imaging biomarker in postmenopausal women with prevalent fragility fractures. J Bone Miner Res. 2013;28(8):1721–8. doi: 10.1002/jbmr.1950. Marrow fat composition was associated with diabetes and prevalent fracture, such that postmenopausal women with diabetes with prevalent fracture had the lowest marrow unsaturation and highest marrow saturation levels. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39•.de Paula FJ, de Araujo IM, Carvalho AL, Elias Jr J, Salmon CE, Nogueira-Barbosa MH. The relationship of fat distribution and insulin resistance with lumbar spine bone mass in women. PLoS ONE. 2015;10(6):e0129764. doi: 10.1371/journal.pone.0129764. Hemoglobin A1c level and marrow fat were positively correlated in a group of women, some of whom had pre-diabetes. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Yeung DK, Griffith JF, Antonio GE, Lee FK, Woo J, Leung PC. Osteoporosis is associated with increased marrow fat content and decreased marrow fat unsaturation: a proton MR spectroscopy study. J Magn Reson Imaging. 2005;22(2):279–85. doi: 10.1002/jmri.20367. [DOI] [PubMed] [Google Scholar]
- 41.Orchard TS, Cauley JA, Frank GC, Neuhouser ML, Robinson JG, Snetselaar L, et al. Fatty acid consumption and risk of fracture in the Women's Health Initiative. Am J Clin Nutr. 2010;92(6):1452–60. doi: 10.3945/ajcn.2010.29955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Coe LM, Lippner D, Perez GI, McCabe LR. Caspase-2 deficiency protects mice from diabetes-induced marrow adiposity. J Cell Biochem. 2011;112(9):2403–11. doi: 10.1002/jcb.23163. [DOI] [PubMed] [Google Scholar]
- 43.Bredella MA, Gill CM, Gerweck AV, Landa MG, Kumar V, Daley SM, et al. Ectopic and serum lipid levels are positively associated with bone marrow fat in obesity. Radiology. 2013;269(2):534–41. doi: 10.1148/radiol.13130375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Zhu ZN, Jiang YF, Ding T. Risk of fracture with thiazolidinediones: an updated meta-analysis of randomized clinical trials. Bone. 2014;68:115–23. doi: 10.1016/j.bone.2014.08.010. [DOI] [PubMed] [Google Scholar]
- 45.Grey A, Beckley V, Doyle A, Fenwick S, Horne A, Gamble G, et al. Pioglitazone increases bone marrow fat in type 2 diabetes: results from a randomized controlled trial. Eur J Endocrinol. 2012;166(6):1087–91. doi: 10.1530/EJE-11-1075. [DOI] [PubMed] [Google Scholar]
- 46.Harslof T, Wamberg L, Moller L, Stodkilde-Jorgensen H, Ringgaard S, Pedersen SB, et al. Rosiglitazone decreases bone mass and bone marrow fat. J Clin Endocrinol Metab. 2011;96(5):1541–8. doi: 10.1210/jc.2010-2077. [DOI] [PubMed] [Google Scholar]
- 47.Rzonca SO, Suva LJ, Gaddy D, Montague DC, Lecka-Czernik B. Bone is a target for the antidiabetic compound rosiglitazone. Endocrinology. 2004;145(1):401–6. doi: 10.1210/en.2003-0746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Ali AA, Weinstein RS, Stewart SA, Parfitt AM, Manolagas SC, Jilka RL. Rosiglitazone causes bone loss in mice by suppressing osteoblast differentiation and bone formation. Endocrinology. 2005;146(3):1226–35. doi: 10.1210/en.2004-0735. [DOI] [PubMed] [Google Scholar]
- 49.Akune T, Ohba S, Kamekura S, Yamaguchi M, Chung UI, Kubota N, et al. PPARgamma insufficiency enhances osteogenesis through osteoblast formation from bone marrow progenitors. J Clin Invest. 2004;113(6):846–55. doi: 10.1172/JCI19900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Botolin S, McCabe LR. Inhibition of PPARgamma prevents type I diabetic bone marrow adiposity but not bone loss. J Cell Physiol. 2006;209(3):967–76. doi: 10.1002/jcp.20804. [DOI] [PubMed] [Google Scholar]
- 51.McCabe LR. Understanding the pathology and mechanisms of type I diabetic bone loss. J Cell Biochem. 2007;102(6):1343–57. doi: 10.1002/jcb.21573. [DOI] [PubMed] [Google Scholar]
- 52.Menagh PJ, Turner RT, Jump DB, Wong CP, Lowry MB, Yakar S, et al. Growth hormone regulates the balance between bone formation and bone marrow adiposity. J Bone Miner Res. 2010;25(4):757–68. doi: 10.1359/jbmr.091015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Bredella MA, Torriani M, Ghomi RH, Thomas BJ, Brick DJ, Gerweck AV, et al. Vertebral bone marrow fat is positively associated with visceral fat and inversely associated with IGF-1 in obese women. Obesity (Silver Spring, Md) 2011;19(1):49–53. doi: 10.1038/oby.2010.106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Amiel SA, Sherwin RS, Hintz RL, Gertner JM, Press CM, Tamborlane WV. Effect of diabetes and its control on insulin-like growth factors in the young subject with type I diabetes. Diabetes. 1984;33(12):1175–9. doi: 10.2337/diab.33.12.1175. [DOI] [PubMed] [Google Scholar]
- 55.Moyer-Mileur LJ, Slater H, Jordan KC, Murray MA. IGF-1 and IGF-binding proteins and bone mass, geometry, and strength: relation to metabolic control in adolescent girls with type 1 diabetes. J Bone Miner Res. 2008;23(12):1884–91. doi: 10.1359/jbmr.080713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Pijl H, Langendonk JG, Burggraaf J, Frolich M, Cohen AF, Veldhuis JD, et al. Altered neuroregulation of GH secretion in viscerally obese premenopausal women. J Clin Endocrinol Metab. 2001;86(11):5509–15. doi: 10.1210/jcem.86.11.8061. [DOI] [PubMed] [Google Scholar]
- 57.Shen W, Chen J, Punyanitya M, Shapses S, Heshka S, Heymsfield SB. MRI-measured bone marrow adipose tissue is inversely related to DXA-measured bone mineral in Caucasian women. Osteoporos Int. 2007;18(5):641–7. doi: 10.1007/s00198-006-0285-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Di Iorgi N, Mittelman SD, Gilsanz V. Differential effect of marrow adiposity and visceral and subcutaneous fat on cardiovascular risk in young, healthy adults. Int J Obes (Lond) 2008;32(12):1854–60. doi: 10.1038/ijo.2008.170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Kiess W, Anil M, Blum WF, Englaro P, Juul A, Attanasio A, et al. Serum leptin levels in children and adolescents with insulin-dependent diabetes mellitus in relation to metabolic control and body mass index. Eur J Endocrinol. 1998;138(5):501–9. doi: 10.1530/eje.0.1380501. [DOI] [PubMed] [Google Scholar]
- 60.Hamrick MW, Pennington C, Newton D, Xie D, Isales C. Leptin deficiency produces contrasting phenotypes in bones of the limb and spine. Bone. 2004;34(3):376–83. doi: 10.1016/j.bone.2003.11.020. [DOI] [PubMed] [Google Scholar]
- 61.Bartell SM, Rayalam S, Ambati S, Gaddam DR, Hartzell DL, Hamrick M, et al. Central (ICV) leptin injection increases bone formation, bone mineral density, muscle mass, serum IGF-1, and the expression of osteogenic genes in leptin-deficient ob/ob mice. J Bone Miner Res. 2011;26(8):1710–20. doi: 10.1002/jbmr.406. [DOI] [PubMed] [Google Scholar]
- 62.Hamrick MW, Della-Fera MA, Choi YH, Pennington C, Hartzell D, Baile CA. Leptin treatment induces loss of bone marrow adipocytes and increases bone formation in leptin-deficient ob/ob mice. J Bone Miner Res. 2005;20(6):994–1001. doi: 10.1359/JBMR.050103. [DOI] [PubMed] [Google Scholar]
- 63.Devlin MJ, Brooks DJ, Conlon C, Vliet M, Louis L, Rosen CJ, et al. Daily leptin blunts marrow fat but does not impact bone mass in calorie-restricted mice. J Endocrinol. 2016;229(3):295–306. doi: 10.1530/JOE-15-0473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Cawthorn WP, Scheller EL, Parlee SD, Pham HA, Learman BS, Redshaw CM, et al. Expansion of bone marrow adipose tissue during caloric restriction is associated with increased circulating glucocorticoids and not with hypoleptinemia. Endocrinology. 2016;157(2):508–21. doi: 10.1210/en.2015-1477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Motyl KJ, McCabe LR. Leptin treatment prevents type I diabetic marrow adiposity but not bone loss in mice. J Cell Physiol. 2009;218(2):376–84. doi: 10.1002/jcp.21608. [DOI] [PMC free article] [PubMed] [Google Scholar]