

Abstract
Several species of the genus Dickeya provoke soft rot and blackleg diseases on a wide range of plants and crops. Dickeya solani has been identified as the causative agent of diseases outbreaks on potato culture in Europe for the last decade. Here, we report the complete genome of the D. solani IPO 2222T. Using PacBio and Illumina technologies, a unique circular chromosome of 4,919,833 bp was assembled. The G + C content reaches 56% and the genomic sequence contains 4,059 predicted proteins. The ANI values calculated for D. solani IPO 2222T vs. other available D. solani genomes was over 99.9% indicating a high genetic homogeneity within D. solani species.
Electronic supplementary material
The online version of this article (doi:10.1186/s40793-016-0208-0) contains supplementary material, which is available to authorized users.
Keywords: Short genome report, Dickeya solani, Blackleg, Soft rot, Genome, Potato
Introduction
Dickeya are pectinolytic enterobacteria that cause soft rot and blackleg diseases on a wide range of crops worldwide including potato plants (Solanum tuberosum) [1, 2]. They are equipped with an arsenal of plant-cell wall degrading enzymes that macerate tuber and stem tissues provoking disease symptoms [3]. In the beginning of the 2000′s, D. solani emerged as a novel species causing blackleg and soft rot diseases on potato in Europe and Mediterranean Basin [4]. Initially, several pectinolytic strains isolated from potatoes grown in Europe and Israel, were identified as members of the Dickeya genus, but shown to exhibit distinctive genetic and physiological traits (biovar 3). Thereafter, additional phylogenetic and biochemical analyses have brought these isolates into a distinct clade called D. solani [5–8]. The D. solani strain IPO 2222 T was isolated from infected potato plants in The Netherlands in 2007 [9].
To date, 12 draft genomes of D. solani are available in GenBank databases. Among them, the genome of the strain IPO 2222 T was sequenced using 454-pyrosequencing with a low average genome coverage (14×). The resulting draft genome is composed of 91 contigs that were assembled in a single scaffold [9]. In this report, we combined Illumina and Pacific Biosciences technologies to provide a complete genome sequence of the strain IPO 2222 T. We also highlighted some phylogenetic and phenotypic key-features of the D. solani species.
Organism information
Classification and features
D. solani IPO 2222 T belongs to the order of Enterobacteria and the class of Gammaproteobacteria. The gapA-based phylogenetic tree (Fig. 1) was congruent with the previously reported trees inferred from MLSA [8, 10], gathering all D. solani strains in a distinct clade within the Dickeya genus. The gapA housekeeping gene was chosen instead of 16S rRNA gene because the sequence analysis of gapA permit a highly resolved view of distinction between members of the Dickeya genus [8, 10].
Fig. 1.
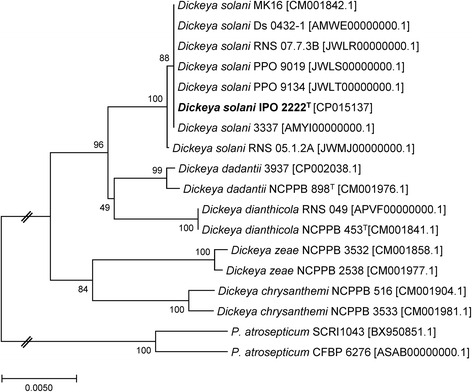
Phylogenetic tree highlighting the relative position of D. solani IPO 2222T within other Dickeya and Pectobacterium species. The unique gapA gene was retrieved from each of the complete and draft genomes that are available in NCBI database; alignment was generated using MUSCLE [23]; the evolutionary history was inferred using the Neighbor-Joining method [24] and the evolutionary distances were computed using the Maximum Composite Likelihood method [25]. Phylogenetic analyses were conducted using MEGA7 software [26]
D. solani IPO 2222 T is a Gram negative, non-spore-forming, motile and facultative anaerobic bacterium with rod shaped cells (0.9x2.0 μm) (Fig. 2) [8]. The strain IPO 2222 T grows in TY medium (tryptone 5 g/L, yeast extract 3 g/L and agar 1.5%) at 28 °C forming 1–2 mm colonies within 24 h. It produces phosphatase and indole and belongs to Dickeya biovar 3 as described previously [10]. Distinctive metabolic abilities of D. solani species were described using BIOLOG system [11]; among them, D. solani IPO 2222 T uses urea as sole nitrogen source (Additional file 1: Figure S1). D. solani IPO 2222 T was recovered form naturally infected potato plants showing blackleg and soft rot symptoms. Its aggressiveness was confirmed by infecting potato tubers and plants in greenhouse assays (Additional file 2: Figure S2). In addition, its ability to colonize the roots and stem tissues and to provoke disease symptoms has been reported using green fluorescent protein-tagged strain [12].
Fig. 2.

Photomicrographs of D. solani IPO 2222T using DAPI (4′,6-diamidino-2-phenylindole) staining (a), differential interference contrast (b) and blue methylene staining (c). These photomicrographs show the rod shaped forms of D. solani species
The strain IPO 2222 T has been registered at the Belgian Co-ordinated Collections of Micro-organisms (LMG 25993 T), the National Collection of Plant Pathogenic Bacteria in UK (NCPPB 4479 T), and the International Center for Microbial Resources - French collection of plant-associated bacteria (CFBP 8199 T). MIGS of D. solani strain IPO 2222 T is summarized in Table 1.
Table 1.
Classification and general features of Dickeya solani strain IPO 2222T [13]
| MIGS ID | Property | Term | Evidence codea |
|---|---|---|---|
| Classification | Domain Bacteria | TAS [15] | |
| Phylum Proteobacteria | TAS [27] | ||
| Class Gammaproteobacteria | TAS [28, 29] | ||
| Order “Enterobacteriales” | TAS [28, 29] | ||
| Family Enterobacteriaceae | TAS [30] | ||
| Genus Dickeya | TAS [1] | ||
| Species Dickeya solani | TAS [8] | ||
| Type strain: IPO 2222T (CP015137) | |||
| Gram stain | negative | TAS [8] | |
| Cell shape | Rod | TAS [8] | |
| Motility | Motile | IDA | |
| Sporulation | Non sporulating | NAS [8] | |
| Temperature range | Mesophilic | TAS [8] | |
| Optimum temperature | 39°C | TAS [8] | |
| pH range; Optimum | Not reported;7 | IDA | |
| Carbon source | D-Arabinose, Mannitol | TAS [8] | |
| MIGS-6 | Habitat | Rhizosphere | TAS [8] |
| MIGS-6.3 | Salinity | 0.5% NaCl (w/v) | TAS [31] |
| MIGS-22 | Oxygen requirement | Facultatively anaerobic | TAS [8] |
| MIGS-15 | Biotic relationship | free-living | TAS [8] |
| MIGS-14 | Pathogenicity | Pathogenic | NAS [8] |
| MIGS-4 | Geographic location | Netherlands | TAS [8, 9] |
| MIGS-5 | Sample collection | 2007 | TAS [8, 9] |
| MIGS-4.1 | Latitude | Not reported | NAS |
| MIGS-4.2 | Longitude | Not reported | NAS |
| MIGS-4.4 | Altitude | Not reported | NAS |
aEvidence codes - IDA: Inferred from Direct Assay; TAS: Traceable Author Statement (i.e., a direct report exists in the literature); NAS: Non-traceable Author Statement (i.e., not directly observed for the living, isolated sample, but based on a generally accepted property for the species, or anecdotal evidence). These evidence codes are from the Gene Ontology project [32]
Genome sequencing information
Genome project history
The genome sequence of D. solani strain IPO 2222 T was sequenced using two technologies, PacBio RSII and Illumina NextSeq 500. This organism was selected based on the agricultural relevance as an emerging pathogen with a significant impact on the potato production and trade in Europe and around the world. Project information is available from Genome Online database number Gp0138842 under the Gold study number Gs0118682 at Joint Genome Institute. The complete genome sequence is also deposited in GenBank under the accession number CP015137. In Table 2, we provide a summary of the project information and its association with MIGS [13].
Table 2.
Project information
| MIGS ID | Property | Term |
|---|---|---|
| MIGS 31 | Finishing quality | Complete genome |
| MIGS-28 | Libraries used | Paired-end |
| MIGS 29 | Sequencing platforms | Illumina NextSeq500, PacBio |
| MIGS 31.2 | Fold coverage | 450X |
| MIGS 30 | Assemblers | CLC Genomics |
| MIGS 32 | Gene calling method | NCBI Prokaryotic Genome Annotation Pipeline |
| Locus Tag | A4U42 | |
| Genbank ID | CP015137 | |
| GenBank Date of Release | 16 Mai 2016 | |
| GOLD ID | Gp0138842 | |
| BIOPROJECT | PRJNA317288 | |
| MIGS 13 | Source Material Identifier | IPO 2222T |
| Project relevance | Agricultural |
Growth conditions and genomic DNA preparation
D. solani IPO 2222 T was routinely cultured in TY medium at 28 °C. Genomic DNA extraction was performed from 5 mL overnight culture using a phenol-chloroform purification method followed by an ethanol precipitation as described by Wilson [14]. Quantification and quality control of the DNA was completed using a NanoDrop (ND 1000) device, Qubit® 2.0 fluorometer and agarose (1.0%) gel electrophoresis.
Genome sequencing and assembly
Second generation sequencing was performed using NextSeq 500 (Illumina, CA, USA) at the I2BC platform (Gif-sur-Yvette, France). A paired-end library was constructed with an insert size of 390 bp and sequencing was carried out using 2 × 151 bp paired-end read module. The de novo assembly (length fraction, 0.5; similarity, 0.8) was performed using CLC Genomics Workbench (v8.0) software (CLC Inc, Aarhus, Denmark). After quality (quality score threshold 0.05) and length (above 40 nucleotides) trimming of the sequences, 33 contigs (N50 = 266,602 bp) were generated (CLC parameters: automatic determination of the word and bubble sizes with no scaffolding) with a 450× average genome coverage. The largest contig length was 617,431 bp.
Third generation sequencing was performed using PacBio RSII (Pacific Biosciences, CA, USA) at the University of Malaya (Kuala Lumpur, Malaysia). The SMRTbell template library at the size of 20 kbp was constructed using the commercial Template Preparation Kit (Pacific Biosciences, CA, USA) followed by sequencing using P6/C4 sequencing chemistry with sequence collection time set at 240 min. Prior to assembly, short reads (less than 500 bp) were filtered off and the minimum polymerase read quality used for mapping of sub-reads from a single zero-mode waveguides was set at 0.75. In total 146,263 reads were obtained (N50 value was 9,161 bp) and total base pair number was at 1,070,191,526 resulting in a 217× average genome coverage. Reads were assembled using RS_HGAP_Assembly software (V2.0). The cut-off length of seeding reads was set at 13,304 bp in order to serve as a reference for the recruitment of shorter reads for preassembly. The resulted consensus accuracy based on multiple sequence alignment of the sub-reads was at 99.99%.
The de novo Illumina-contigs were used to verify the RS_HGAP assembly by blasting them against the PacBio sequence. In addition, the trimmed Illumina reads were mapped (length fraction, 0.5; similarity, 0.8) against the PacBio sequence and errors (SNPs and InDels), that might be generated by homopolymers during PacBio sequencing, were searched and corrected using basic variant calling tool from CLC genomic workbench. Using these two sets of sequences, the complete genome sequence was approved and circularized.
Genome annotation
The complete genome of D. solani IPO 2222 T was annotated using the NCBI prokaryotic genome annotation pipeline [15]. The protein coding gene prediction process begin by an alignment using ProSplign [16] where only complete alignments with 100% identity to a reference protein are kept for final annotation. Then the remaining frameshift or partial alignments were further analyzed by GeneMarkS+ [17]. To identify structural rRNA, the pipeline uses BLASTn search against the curated reference set. tRNAscan-SE was used to identify the tRNAs [18]. The CRISPRs are identified by using the CRISPR database [15].
Genome properties
The detailed information about Dickeya solani IPO 2222 T genome is provided in Table 3. The genome is constituted of one circular chromosome, 4,919,833 bp in size. The annotation predicted 4,208 genes including 4,059 CDSs (Table 4), 104 RNA genes (75 tRNA, 22 rRNA and 7 ncRNA genes) and 45 pseudo genes. The G + C reached 56%. The graphical genome map is provided in the Fig. 3.
Table 3.
Genome statistics
| Attribute | Value | % of total |
|---|---|---|
| Genome size (bp) | 4,919,833 | 100.00 |
| DNA coding (bp) | 4,243,944 | 86.33 |
| DNA G + C (bp) | 2,767,155 | 56.24 |
| DNA scaffolds | 1 | 100.00 |
| Total genes | 4,208 | 100.00 |
| Protein coding genes | 4,104 | 97.5 |
| RNA genes | 104 | 2.5 |
| Pseudo genes | 45 | 1.06 |
| Genes in internal clusters | 1,093 | 25.97 |
| Genes with function prediction | 3,670 | 87.21 |
| Genes assigned to COGs | 3,365 | 79.97 |
| Genes with Pfam domains | 3,788 | 99.02 |
| Genes with signal peptides | 386 | 9.17 |
| Genes with transmembrane helices | 953 | 22.65 |
| CRISPR repeats | 1 | - |
Table 4.
Number of genes associated with general COG functional categories
| Code | Value | % agea | Description |
|---|---|---|---|
| J | 234 | 6.09 | Translation, ribosomal structure and biogenesis |
| A | 1 | 0.03 | RNA processing and modification |
| K | 305 | 7.94 | Transcription |
| L | 112 | 2.92 | Replication, recombination and repair |
| B | 0 | 0.00 | Chromatin structure and dynamics |
| D | 42 | 1.09 | Cell cycle control, Cell division, chromosome partitioning |
| V | 90 | 2.34 | Defense mechanisms |
| T | 216 | 5.62 | Signal transduction mechanisms |
| M | 245 | 6.38 | Cell wall/membrane biogenesis |
| N | 106 | 2.76 | Cell motility |
| U | 82 | 2.14 | Intracellular trafficking and secretion |
| O | 138 | 3.59 | Posttranslational modification, protein turnover, chaperones |
| C | 222 | 5.78 | Energy production and conversion |
| G | 324 | 8.44 | Carbohydrate transport and metabolism |
| E | 438 | 11.41 | Amino acid transport and metabolism |
| F | 96 | 2.5 | Nucleotide transport and metabolism |
| H | 192 | 5.0 | Coenzyme transport and metabolism |
| I | 130 | 3.39 | Lipid transport and metabolism |
| P | 281 | 7.32 | Inorganic ion transport and metabolism |
| Q | 93 | 2.42 | Secondary metabolites biosynthesis, transport and catabolism |
| R | 282 | 7.34 | General function prediction only |
| S | 175 | 4.56 | Function unknown |
| - | 843 | 20.03 | Not in COGs |
| bTotal | 4,683 | 120 |
aThe percentage is based on the total number of protein coding genes in the annotated genome
bThe total does not correspond to 4,208 CDS because some genes are associated with more than one COG functional categories
Fig. 3.
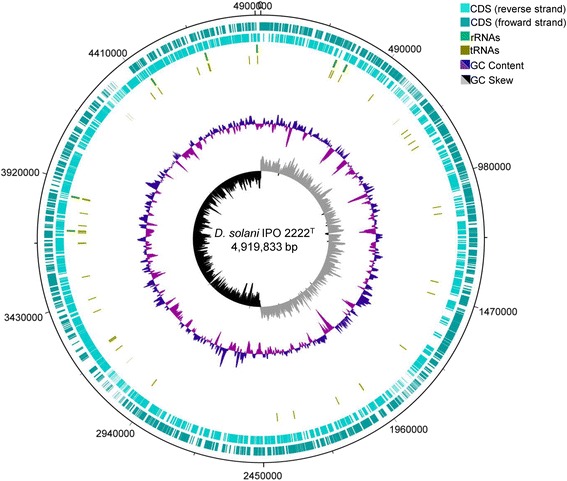
Graphical circular map of D. solani IPO 2222T chromosome
Insights from the genome sequence
D. solani species is genetically highly homogenous with 99.9% in genomic similarity (ANI value) [19, 20]. Between two given D. solani genomes, the number of variations (SNPs/InDels) is below one hundred. For example, when D. solani strain 3337 and D. solani strain IPO 2222 T were compared, 49 variations were observed: 15 were located out of CDS and 34 within CDS [19]. Only a few of D. solani genomes (strains RNS 07.7.3B, PPO 9019 and PPO 9134) exhibited a higher number of variations (>1000) because they acquired D. dianthicola genes by horizontal gene transfer [19]. None horizontal gene transfer from D. dianthicola was observed in strain IPO 2222 T.
Plant-cell wall degrading enzymes comprising pectinases, proteinases and cellulases, play a major role in the plant tissue maceration process [21]. Indeed, 10 pectates lyase enzymes (genes pelABCDEILXWZ) were predicted in strain IPO 2222 T genome; they showed a 93.3% average nucleotide identity when compared to the orthologous genes of D. dadantii 3937.
Recent comparative analyses underlined the major genetic and metabolic divergences between Dickeya solani species and the nearest clades that are D. dandatii (ANI 94%) and D. dianthicola (ANI 92%) [11, 19]. D. solani is characterized by a low content of phages elements and CRISPR system: in strain IPO 2222 T genome, only one CRISPR cluster (208 bp) was identified. Using PHAST tool [22], the strain IPO 2222 T harbors one questionable prophage (11 CDSs) in a 10,687 bp region. In addition, some genomic regions were shown to be specific for D. solani species and contain some metabolic and NRPS/PKS encoding genes [11].
Conclusions
The complete sequence of D. solani IPO 2222 T is the first complete genome of a member of this species, the type strain. This work provides a substantial resource in terms of knowledge of the bacterial genetic material. It may help to understand the successful fitness of D. solani in invading potato fields, opening the way to new control strategies against this phytopathogen.
Acknowledgements
We thank Peter Mergaert (I2BC, CNRS) for help in microscopy imaging. This work has benefited from the platforms and expertise of the High-throughput Sequencing Platform of I2BC (Gif-sur-Yvette, France). This work is supported by CNRS (I2BC-SE2015), Agence Nationale de la Recherche (ANR-15-CE21-0003), the High Impact Research Grant (UM.C/625/1/HIR/MOHE/CHAN/14/01, Grant number A-000001-50001) and the French-Malaysian exchange program awarded by French Embassy in Malaysia, the continuous support of which is acknowledged here.
Authors’ contributions
SK, PB, TMC performed experiments, SK assembled and analysed genome, SK, PB, TMC, KGC, DF wrote the paper. All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Abbreviations
- CDS
Coding DNA sequence
- CRISPR
Clustered regularly interspaced short palindromic repeats
- MIGS
Minimum information on the genome sequence
- MLSA
Multi-locus sequence analysis
Additional files
Growth curves of D. solani IPO 2222T, D. dadantii 3937 and D. dianthicola RNS 049 in the presence of urea as a sole nitrogen source. Data were collected from duplicates. (TIFF 296 kb)
Symptoms of D. solani IPO 2222T on potato plants (a) and tubers (b). (TIFF 8904 kb)
References
- 1.Samson R, Legendre JB, Christen R, Fischer-Le Saux M, Achouak W, Gardan L. Transfer of Pectobacterium chrysanthemi (Burkholder et al. 1953) Brenner et al. 1973 and Brenneria paradisiaca to the genus Dickeya gen. nov. as Dickeya chrysanthemi comb. nov. and Dickeya paradisiaca comb. nov. and delineation of four novel species, Dickeya dadantii sp. nov., Dickeya dianthicola sp. nov., Dickeya dieffenbachiae sp. nov. and Dickeya zeae sp. nov. Int J Syst Evol Microbiol. 2005;55:1415–1427. doi: 10.1099/ijs.0.02791-0. [DOI] [PubMed] [Google Scholar]
- 2.Gardan L, Gouy C, Christen R, Samson R. Elevation of three subspecies of Pectobacterium carotovorum to species level: Pectobacterium atrosepticum sp. nov., Pectobacterium betavasculorum sp. nov. and Pectobacterium wasabiae sp. nov. Int J Syst Evol Microbiol. 2003;53:381–391. doi: 10.1099/ijs.0.02423-0. [DOI] [PubMed] [Google Scholar]
- 3.Collmer A, Keen NT. The role of pectic enzymes in plant pathogenesis. Annu Rev Phytopathol. 1986;24:383–409. doi: 10.1146/annurev.py.24.090186.002123. [DOI] [Google Scholar]
- 4.Toth IK, van der Wolf JM, Saddler G, Lojkowska E, Hélias V, Pirhonen M, et al. Dickeya species: an emerging problem for potato production in Europe: Dickeya spp. on potato in Europe. Plant Pathol. 2011;60:385–399. doi: 10.1111/j.1365-3059.2011.02427.x. [DOI] [Google Scholar]
- 5.Tsror L, Erlich O, Lebiush S, Hazanovsky M, Zig U, Slawiak M, et al. Assessment of recent outbreaks of Dickeya sp. (syn. Erwinia chrysanthemi) slow wilt in potato crops in Israel. Eur J Plant Pathol. 2009;123:311–320. doi: 10.1007/s10658-008-9368-0. [DOI] [Google Scholar]
- 6.Parkinson N, Stead D, Bew J, Heeney J, Tsror Lahkim L, Elphinstone J. Dickeya species relatedness and clade structure determined by comparison of recA sequences. Int J Syst Evol Microbiol. 2009;59:2388–2393. doi: 10.1099/ijs.0.009258-0. [DOI] [PubMed] [Google Scholar]
- 7.Laurila J, Ahola V, Lehtinen A, Joutsjoki T, Hannukkala A, Rahkonen A, et al. Characterization of Dickeya strains isolated from potato and river water samples in Finland. Eur J Plant Pathol. 2008;122:213–225. doi: 10.1007/s10658-008-9274-5. [DOI] [Google Scholar]
- 8.van der Wolf JM, Nijhuis EH, Kowalewska MJ, Saddler GS, Parkinson N, Elphinstone JG, et al. Dickeya solani sp. nov., a pectinolytic plant-pathogenic bacterium isolated from potato (Solanum tuberosum) Int J Syst Evol Microbiol. 2014;64:768–774. doi: 10.1099/ijs.0.052944-0. [DOI] [PubMed] [Google Scholar]
- 9.Pritchard L, Humphris S, Baeyen S, Maes M, Vaerenbergh JV, Elphinstone J, et al. Draft Genome Sequences of Four Dickeya dianthicola and Four Dickeya solani Strains. Genome Announc. 2013;1:e00087–12. doi: 10.1128/genomeA.00087-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Sławiak M, van Beckhoven JRCM, Speksnijder AGCL, Czajkowski R, Grabe G, van der Wolf JM. Biochemical and genetical analysis reveal a new clade of biovar 3 Dickeya spp. strains isolated from potato in Europe. Eur J Plant Pathol. 2009;125:245–261. doi: 10.1007/s10658-009-9479-2. [DOI] [Google Scholar]
- 11.Pédron J, Mondy S, Raoul des Essarts Y, Van Gijsegem F, Faure D. Genomic and metabolic comparison with Dickeya dadantii 3937 reveals the emerging Dickeya solani potato pathogen to display distinctive metabolic activities and T5SS/T6SS-related toxin repertoire. BMC Genomics. 2014;15:283. doi: 10.1186/1471-2164-15-283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Czajkowski R, de Boer WJ, Velvis H, van der Wolf JM. Systemic colonization of potato plants by a soilborne, green fluorescent protein-tagged strain of Dickeya sp. biovar 3. Phytopathology. 2010;100:134–142. doi: 10.1094/PHYTO-100-2-0134. [DOI] [PubMed] [Google Scholar]
- 13.Field D, Garrity G, Gray T, Morrison N, Selengut J, Sterk P, et al. The minimum information about a genome sequence (MIGS) specification. Nat Biotechnol. 2008;26:541–547. doi: 10.1038/nbt1360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Wilson K. Preparation of Genomic DNA from Bacteria. Curr Protoc Mol Biol John Wiley & Sons, Inc; 2001. Available from: http://dx.doi.org/10.1002/0471142727.mb0204s56 [DOI] [PubMed]
- 15.Woese CR, Kandler O, Wheelis ML. Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya. Proc Natl Acad Sci. 1990;87:4576–4579. doi: 10.1073/pnas.87.12.4576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Wheeler DL. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2006;34:D173–D180. doi: 10.1093/nar/gkj158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.GeneMarkS BJ. a self-training method for prediction of gene starts in microbial genomes. Implications for finding sequence motifs in regulatory regions. Nucleic Acids Res. 2001;29:2607–2618. doi: 10.1093/nar/29.12.2607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lowe TM, Eddy SR. tRNAscan-SE: A program for improved detection of transfer RNA Genes in genomic sequence. Nucleic Acids Res. 1997;25:955–964. doi: 10.1093/nar/25.5.0955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Khayi S, Blin P, Pédron J, Chong T-M, Chan K-G, Moumni M, et al. Population genomics reveals additive and replacing horizontal gene transfers in the emerging pathogen Dickeya solani. BMC Genomics. 2015;16:788. doi: 10.1186/s12864-015-1997-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Khayi S, Mondy S, Beury-Cirou A, Moumni M, Helias V, Faure D. Genome Sequence of the Emerging Plant Pathogen Dickeya solani Strain RNS 08.23.3.1A. Genome Announc. 2014;2:e01270–13. doi: 10.1128/genomeA.01270-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Hugouvieux-Cotte-Pattat N, Condemine G, Nasser W, Reverchon S. Regulation of pectinolysis in Erwinia chrysanthemi. Annu Rev Microbiol. 1996;50:213–257. doi: 10.1146/annurev.micro.50.1.213. [DOI] [PubMed] [Google Scholar]
- 22.Zhou Y, Liang Y, Lynch KH, Dennis JJ, Wishart DS. PHAST: a fast phage search tool. Nucleic Acids Res. 2011;39:W347–W352. doi: 10.1093/nar/gkr485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Edgar RC. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004;32:1792–1797. doi: 10.1093/nar/gkh340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Saitou N, Nei M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol. 1987;4(4):406–25. [DOI] [PubMed]
- 25.Tamura K, Nei M, Kumar S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc Natl Acad Sci U S A. 2004;101:11030–11035. doi: 10.1073/pnas.0404206101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kumar S, Stecher G, Tamura K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol Biol Evol. 2016;33:1870–1874. doi: 10.1093/molbev/msw054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Garrity GM, Bell JA, Lilburn T. Phylum XIV. Proteobacteria Phyl. nov. In: Garrity GM, Brenner DJ, Krieg NR, Staley JT, editors. Bergey’s Manual of Systematic Bacteriology. Volume 2 (Part B) 2. New York: Springer; 2005. p. 1. [Google Scholar]
- 28.Garrity GM, Bell JA, Lilburn T. Class I. Alphaproteobacteria class. nov. Bergey’s Manual Syst. Bacteriol. Springer; 2005. p. 1–574
- 29.Williams KP, Kelly DP. Proposal for a new class within the phylum Proteobacteria, Acidithiobacillia classis nov., with the type order Acidithiobacillales, and emended description of the class Gammaproteobacteria. Int J Syst Evol Microbiol. 2013;63:2901–2906. doi: 10.1099/ijs.0.049270-0. [DOI] [PubMed] [Google Scholar]
- 30.Ewing WH, Farmer J, III, Brenner DJ. Proposal of Enterobacteriaceae fam. nov., nom. rev. to replace Enterobacteriaceae Rahn 1937, nom. fam. cons. (Opin. 15, Jud. Comm. 1958), which lost standing in nomenclature on 1 January 1980. Int J Syst Evol Microbiol. 1980;30:674–675. [Google Scholar]
- 31.Perombelon MCM, Kelman A. Ecology of the soft rot erwinias. Annu Rev Phytopathol. 1980;18:361–387. doi: 10.1146/annurev.py.18.090180.002045. [DOI] [Google Scholar]
- 32.Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, et al. Gene ontology: tool for the unification of biology. The Gene Ontology Consortium. Nat Genet. 2000;25:25–29. doi: 10.1038/75556. [DOI] [PMC free article] [PubMed] [Google Scholar]