

Abstract
To determine the efficacy of a novel and safer (for gastrointestinal tract) aspirin (aspirin-PC) in preclinical models of ovarian cancer, in vitro dose-response studies were performed to compare the growth-inhibitory effect of aspirin-PC vs. aspirin on 3 human (A2780, SKOV3ip1, HeyA8), and a mouse (ID8) ovarian cancer cell line over an 8-day culture period. In the in vivo studies, the aspirin test drugs were studied alone and in the presence of a VEGF-A inhibitor (bevacizumab or B20), due to an emerging role for platelets in tumor growth following anti-angiogenic therapy, and we examined their underlying mechanisms. Aspirin-PC was more potent (vs. aspirin) in blocking the growth of both human and mouse ovarian cancer cells in monolayer culture. Using in vivo model systems of ovarian cancer, we found that aspirin-PC significantly reduced ovarian cancer growth by 50–90% (depending on the ovarian cell line/density). The efficacy was further enhanced in combination with Bevacizumab or B20. The growth-inhibitory effect on ovarian tumor mass and number of tumor nodules was evident, but less pronounced for aspirin and the VEGF inhibitors alone. There was no detectable gastrointestinal toxicity. Both aspirin and aspirin-PC also inhibited cell proliferation, angiogenesis and increased apoptosis of ovarian cancer cells. In conclusion, PC-associated aspirin markedly inhibits the growth of ovarian cancer cells, which exceeds that of the parent drug, in both cell culture and in mouse model systems. We also found that both aspirin-PC and aspirin have robust anti-neoplastic action in the presence of VEGF blocking drugs.
Keywords: ovarian cancer, aspirin, aspirin-PC, bevacizumab, B20, platelet
INTRODUCTION
Over the past fifty years there has been a growing awareness that aspirin consumption on a regular basis is associated with a reduced incidence and mortality of many cancers afflicting no less than 20 tissues (1–3). The strongest evidence is based on large meta-analyses (>100,000 subjects), following health professionals and nurses for over 10–20 years, where it has been reported that cancer incidence is reduced by 25–40% in subjects regularly taking low dose (75–325 mg) aspirin on a daily basis, with the most compelling evidence acquired for colorectal cancer. These findings have resulted in a subcommittee of the US Preventive Services Task Force (USPSTF), recently recommending that individuals aged 50–69 years old, daily take low-dose aspirin for 10 years or longer to prevent colorectal cancer, with the proviso they consult their physician beforehand, as aspirin can cause gastrointestinal (GI) ulceration and bleeding which can develop into life-threatening hemorrhage in susceptible individuals (http://www.uspreventiveservicestaskforce.org/Page/Document/draft-recommendation-statement/aspirin-to-prevent-cardiovascular-disease-and-cancer). Rothwell et al also reported convincing data for nine other cancers where chronic use of low dose aspirin was associated with a statistically significant reduction of cancer incidence and mortality (4, 5). It was also striking that the same group reported that aspirin use even when initiated post-cancer diagnosis could have a significant impact by reducing the metastatic spread of the primary cancer and prolong cancer patient survival as much as 2-fold (4, 5).
Ovarian cancer is the leading cause of gynecological cancer related mortality. Following initial response to therapy, there is frequently rapid emergence of drug resistance (6, 7). In a recent study, we also demonstrated that platelets play an important role in tumor regrowth following withdrawal of anti-VEGF therapy (8). Though prior studies have mostly focused on associating aspirin use to GI cancer, a number of clinical outcome studies have recently been reported that ovarian cancer is particularly sensitive to chronic consumption of aspirin, with ovarian cancer incidence being significantly reduced in regular users of low dose aspirin (9, 10), though other studies reported aspirin and NSAID use did not provide an effect that reached statistical significance (11). In addition, similar to both colorectal (5) and breast cancer (12) there is evidence that aspirin use even post-diagnosis can reduce the recurrence of ovarian cancer and possibly prolong patient survival (9).
As indicated by the USPSTF recommendations, aspirin use has been limited, due to the drug’s adverse event profile, which like other NSAIDs can cause potentially serious GI ulceration and bleeding, especially if taken at high anti-inflammatory doses. Evidence suggests that NSAIDs induce GI injury by both inhibiting epithelial COX activity and thereby depleting the tissue of “cytoprotective” prostaglandins and by a direct cytotoxic effect on the GI mucosae by compromising cellular (plasmalemmal and mitochondrial) extracellular/mucus function, thereby disrupting both barrier function and electron transport/ATP generation of the tissue (13, 14). The chemical association of NSAIDs with membrane and extracellular phospholipids has been of substantial interest to us (14–17), leading to the development of a family of phosphatidylcholine (PC)-associated NSAIDs (PC-NSAIDs) (16, 18–22). The formulation that has undergone most rigorous pre-clinical and clinical testing is aspirin-PC, in which aspirin is formulated with PC-enriched soy lecithin (called PL2200 Aspirin) and has recently been awarded a new drug approval (NDA, #203697) by the FDA.
Based upon the encouraging results in the literature on the chemopreventive activity of aspirin, we have previously performed an in vitro study on colon cancer cell lines, demonstrating that PC-associated aspirin had clear potential to inhibit cell growth, with a suggestion that the complex may be more potent than the parent drug (23). Here, we examined the biological effects of aspirin-PC formulation in ovarian cancer models alone and in combination with anti-VEGF monoclonal antibodies.
MATERIALS AND METHODS
Cell culture
We purchased the human ovarian cancer cell lines HeyA8, SKOV3ip1 and A2780 and the murine ovarian cancer cell line ID8 from the MD Anderson Characterized Cell Line Core Facility, which supplies authenticated cell lines. All experiments were performed with cell lines at 60% to 80% confluence. The HeyA8, SKOV3ip1 and A2780 cells were cultured in RPMI1640 medium supplemented with 10% (SKOV3ip1 and A2780) or 15% (HeyA8) fetal bovine serum and 0.5% gentamicin, maintained on plastic and incubated at 37°C in a mixture of 5% CO2 and 95% air. ID8 cells were cultured in DMEM medium containing 5% fetal bovine serum, 1x insulin-transferrin-selenium (ITS) and 0.5% gentamicin. The tumor cells were free of pathogenic murine viruses and Mycoplasma (assayed by M.A. Bioproducts, Walkersville, MD).
To study the effects of our aspirin test formulations on cell growth, at subculture each of the ovarian cancer cell lines were pre-incubated with the aspirin test drugs at a concentration range from 0–1.0 mM for 15 min, prior to pipetting the cells onto a 48-well plates at a density of 2×103 cells/well and cultured at 37°C in a mixture of 5% CO2 and 95% air. The cells were then cultured in the above growth medium in the presence and absence of the aspirin test formulations (at the same concentration, as used during the pre-incubation period) for 8 days (changing the medium with fresh medium on the 4th day). At the completion of the growth period, the number of viable cells/well were assessed using the established MTT method, which provides a linear relationship between the colored reaction product and cell number (23). Briefly this entails adding 3-(4,5-dimethylthiazolyl-2) 2, 5-diphenyltetra-zolium bromide (MTT, Sigma Aldrich Chemical Co., St Louis, MO), to the media of cells at a final concentration of 0.5 mg/ml for 4 hr. The solution containing the resultant purple-colored formazan product was then extracted into a solvent (90% isopropanol, 0.2% sodium dodecyl sulfate and 0.01N HCl) which was then collected from the wells and read at an absorbance of 570nm.
Tube forming assay
Matrigel (12.5 mg/mL) was thawed at 4°C, and 50 μL was added to each well (μ-Slide Angiogenesis, Ibidi, cat. no. 81506) and allowed to solidify for 20 min at 37°C. The wells with immortalized human vascular endothelial cells (RF24) cells (8,000 per well), which had previously been treated with or without Aspirin-PC or aspirin at 0.4 mM for 4 or 8 days were then incubated for 6 h at 37°C (which we have determined is sufficient time for tube formation to be complete). Experiments were performed in triplicate, using a previously described method (24). Employing an Olympus IX81 inverted microscope, images per well were taken at ×100 magnification. The amount of fully formed tubes per image was quantified.
Drugs and antibodies
Aspirin (uncoated) for the animal studies was purchased from Walgreens. Aspirin-PC was prepared as described below for the culture and animal studies. Bevacizumab (Genentech/Roche) is a humanized mAb (monoclonal antibody) that inhibits VEGF-A. Anti-VEGF antibody B20 (Genentech) is a cross-species reactive, function-blocking mAb targeting both human and murine VEGF-A. The antibodies used in immunofluorescence and immunohistochemistry were CD31 (BD Pharmingen, San Diego, CA), Ki67 (Neomarkers Inc., Fremont, CA), cleaved caspase-3 (Biocare Medical LLC, Concord, CA), CA9 (Novus Biologicals, Littleton, CO), horseradish peroxidase-conjugated goat anti-rabbit immunoglobulin G (Serotec Harlan Bioproducts for Science, Inc., Madison, WI), biotinylated goat anti-rabbit secondary antibody (Biocare Medical, Concord, CA) and Alexa Fluor 594-conjugated anti-rat antibody (Jackson ImmunoResearch Laboratories, West Grove, PA).
Aspirin test formulations
We used established procedures to prepare our PC-associated aspirin test formulations for cell culture and intragastric dosing (18, 21). For cell culture, to prepare the drug stock solution, aspirin was dissolved in the serum free culture medium at 10mM and then combined with an equimolar amount of purified soy phosphatidylcholine/PC (S-100, provided by Lipoid Inc, GE), which was previously dissolved in chloroform and then blown dry under nitrogen. The tubes were then sonicated at room temperature in a bath-type sonicator for 20 min until a homogenous suspension was obtained. (See Supplementary Material, Fig S-1 for the chemical structures of aspirin and soy PC). For animal dosing we followed the procedure established for preparing aspirin-PC for oral administration. This involved adding equal concentrations of protonated aspirin (Rhodia) to PC-enriched soy lecithin (P35SB, provided by Lipoid Inc.) which was pre-warmed to 37°C under constant stirring until a homogenous clear suspension was formed. This suspension, was made weekly and stored at room temperature in the dark until immediately before animal dosing at which time the drug solution at 5 mg aspirin/ml was made with water to the desired concentration, and sonicated in a bath-type sonicator as outlined above prior to intragastric administration to the mice with a feeding needle. The aspirin comparator formulations were prepared at the equivalent concentration, in either the culture medium or vehicle (water) for the in vitro and in vivo studies, respectively.
ELISA
Blood samples were allowed to clot for 2 hours at room temperature before centrifuging for 20 minutes at 2000 × g. Serum samples were diluted 1:50 for TGF-β1 and 1:10 for TXB2 measurement. Total TGF-β1 in serum was measured after converting latent TGF-β1 to active TGF-β1 by acidification (10-minute incubation at RT with 1 N HCl for serum, followed by neutralization by adding the same volume of 1.2 N NaOH in 0.5M HEPES) with a sandwich ELISA specific for the activated form of TGF-β1 (R&D Systems). TXB2 was measured by a competitive ELISA kit (R&D Systems).
Fecal hemoglobin (Hb) analysis
Fecal Hb was monitored by collecting the fecal droppings at regular intervals from the bedding and storing them at −20°C until the day of analysis. At this time the feces were allowed to thaw to room temperature, weighed and then distilled water was added at a 1g:10ml feces: water ratio. After standing for 1 hour, the feces were disrupted into a homogenous suspension by vortexing for 2 minutes and then the Hb analyzed by a previously described method (19).
Animal experiments
Female athymic nude mice (NCr-nu) and C57BL/6 mice were purchased from Taconic Farms. HeyA8 and SKOV3ip1 cells were injected into the intraperitoneal cavity of NCr-nu mice (at a concentration of 2.5×105 cells/0.2 ml for HeyA8 and 1×106 cells/0.2 ml for SKOV3ip1 cells) per mouse as described in previous publications using the orthotopic mouse model of ovarian cancer (25–27). ID8 cells at a concentration of 2×106 cells/0.2 ml per mouse were injected into the intraperitoneal cavity of C57BL/6 mice. All mouse studies were approved by the Institutional Animal Care and Use Committee and were cared for in accordance with guidelines set forth by the American Association for Accreditation of Laboratory Animal Care and the US Public Health Service Policy on Human Care and Use of Laboratory Animals. In most of the in vivo studies the mice were randomly divided into 6 treatment groups: untreated control, aspirin (20 mg/kg intragastrically, dissolving the drug in water, daily), Aspirin-PC (20 mg aspirin/kg intragastrically, dissolving the drug in water, daily), Bev (Bevacizumab, 6.25 mg/kg intraperitoneally, dissolving the drug in PBS, twice weekly), B20 (5 mg/kg intraperitoneally, dissolving the drug in PBS, twice weekly), or aspirin or aspirin-PC plus Bev or B20. Treatment was initiated 7–10 days after cancer cell injection into the intraperitoneal cavity. After 3–4 weeks of treatment, the mice were sacrificed, and total body weight, total blood, tumor location and weight, and number of tumor nodules were recorded. Tumor specimens were preserved in either optimum cutting temperature medium (Miles Inc., Elkhart, IN) (for frozen slides), or fixed in formalin (for paraffin slides) for further analysis.
Immunohistochemical and immunofluorescence staining
Paraffin-embedded tumor tissue samples were incubated with Ki67 (1:400) or cleaved caspase-3 (1:100). Staining with rat monoclonal anti-mouse CD31 (1:800; BD Pharmingen) was performed on the frozen sections using immunofluorescence (IF) staining using a previously described technique (28). After incubation overnight at 4°C, Alexa Fluor 594 conjugated anti-rat antibody (1:1000) was incubated for 1 hour at room temperature. After the samples were washed with PBS, they were incubated with DAPI (Invitrogen) for 10 minutes at room temperature. Mounted samples were visualized using a fluorescent microscope (Leica Microsystems CMS GmbH, Wetzlar, Germany) with the appropriate filter (Original magnification ×200). Two independent investigators evaluated immunohistochemical staining. Expression was scored as intensity (1 = no/focal, 2 = weak, 3 = moderate, 4 = strong), ordinal ranking of percent positive cells (0 = none, 1 = 1–25%, 2 = 26–50%, 3 = 51–75%, 4 = 76–100%), and an overall score calculated as the intensity and ordinal ranking of percent positive cells.
To detect hypoxia in the collected ovarian tumor tissues, we examined carbonic anhydrase-9 (CA9) expression using a previously described immunohistochemical staining and scoring method (29). Briefly this entailed incubating paraffin tumor tissue sections with anti-rabbit CA9 antibody (1:500, cat. no. NB100-417, Novus Biologicals) 4°C overnight. After washing the slides with PBS, tumor sections were incubated with a biotinylated goat anti-rabbit secondary antibody (cat.no. GR602H, Biocare Medical) followed by incubation with Streptavidin-HRP (cat.no. HP604H, Biocare Medical). Signal development was done using DAB. CA9 tumor expression was scored by multiplying grade of intensity (0 = negative, 1 = weak, 2 = moderate, 3 = strong) with percentage of positive cells (0 = no positive cells, 1 = <10% positive cells, 2 = 10–50% positive cells, 3 = 51–80% positive cells, 4 = >80% positive cells; range 0–12).
HPLC analysis
Aspirin and salicylic acid from culture media were analyzed by High Performance Liquid Chromatography (HPLC) using a previously described method (30). Briefly this entailed extracting the samples in acetonitrile (1 part media: 3 parts acetonitrile), followed by centrifugation with 10 μl samples supernatant applied with a Model 2707 autosampler onto an Agilent Zorbax 300 SBC18 column (at 45°C) using a Model 1525 binary HPLC pump with mixtures of 50 mM phosphoric acid/acetonitrile (80%/20% initially), followed by a 20%/80% mixture. Detection was through a Model 2489 UV detector at 230 nm. The salicylic acid and aspirin concentrations were determined by comparing the elution peaks to that of purified standards of acetylsalicylic and salicylic acid.
Statistical analyses
Statistical analyses were performed using the Statistical Package for Social Science software version 18.0 (SPSS, Inc., Chicago, IL). Continuous variables were compared using a Student’s t test (for 2 groups) or analysis of variance (for all groups) if the data were normally distributed. For nonparametric distributions, the Mann-Whitney U test (for 2 groups) was used. A P value < 0.05 with 2-tailed testing was deemed statistically significant.
RESULTS
In vitro effects of aspirin and aspirin-PC on the growth of ovarian cells in culture
We first examined the effects of our aspirin test drugs on the growth of four ovarian cancer cell lines (Figure 1A–D). Aspirin had only a modest to negligible inhibitory effect on the growth of these cell lines within the dose-range studied, with A2780 and SKOVip1 only showing a response at the highest doses tested and HeyA8 human and ID8 mouse cells showing a clear response at doses of ≥0.16mM. Of particular note, all four cell lines showed a more prominent growth-inhibitory response to aspirin-PC at most of the doses tested over the 8-day culture period. It was determined that this growth inhibitory effect was not attributable to our aspirin test formulations interfering with the initial attachment of the cells to the surface of the culture plate, as the cell density was not affected by either aspirin of aspirin-PC after incubation periods of ≤ 4 days. We also determined by HPLC that aspirin (when added to the medium as either unmodified aspirin or Aspirin-PC was stable (<10% breakdown to salicylate) in over a 4-day culture period, after which it was exchanged for fresh aspirin-containing medium (data not shown).
Figure 1. In vitro evidence that aspirin-PC and aspirin inhibit the growth of four ovarian cancer cell lines derived from human; (A) HeyA8, (B) SKOV3ip1, (C) A2780; and mouse (D) ID8.

Dose response experiments using MTT to assess cell growth over an 8 day culture period were studies demonstrating the increased effectiveness of PC-associated aspirin vs unmodified aspirin to inhibit ovarian cell growth in cell culture. *= p<0.05 (using Fisher PLSD test. △= p<0.05 (using t-test).
In vivo effects of aspirin-PC, aspirin and VEGF inhibitors in a mouse model of ovarian cancer
On the basis of our in vitro studies, we next examined the in vivo efficacy of our aspirin test drugs alone and in combination with VEGF-A antagonists on the growth of ovarian tumors using established orthotopic and syngeneric mouse model systems. Aspirin-PC (at a dose of 20mg aspirin/kg) alone and in combination with a VEGF-A inhibitor (either Bev or B20) resulted in a significant inhibition of the growth of HeyA8 and SKOV3ip1 ovarian tumors; similar results were noted with the syngeneic ID8 model (Fig. 2A–C) and the number of tumor nodules (Fig. 2D–F). Of particular note, was the observation that the highly significant growth inhibitory action of our aspirin test drugs (aspirin and aspirin-PC) was equivalent to or exceeded that of the VEGF inhibitor, whereas the combination of the two classes of drugs induced a further reduction in both tumor mass and number of tumor nodules that was appreciably lower than either test agent alone.
Figure 2. In vivo effects of aspirin, aspirin-PC, and Bev or B20 in ovarian cancer growth in mice: Tumor mass (A–C); and Tumor nodules (E–F).

In vivo effect of aspirin, aspirin-PC, and Bev or B20 on average tumor weight of mice (n=10/group) in three independent experiments are shown using: (A) SKOV3ip1; (B) HeyA8 cells and (C) ID8 cells. The in vivo effects of aspirin, Aspirin-PC, and bevacizumab (BEV) or B20 on average tumor nodule number of mice (n=10/group) in three independent experiments are shown using: (D) SKOV3ip1; (E) HeyA8 cells and (F) ID8 cells. Error bars indicate the standard error of the mean. * p<0.05, ** p<0.01.
In two of the animal experiments, we compared the inhibitory activity of aspirin-PC vs. aspirin on the growth of human (HeyA8) and mouse (ID8) ovarian cancer cells in the presence and absence of Bev or B20, respectively. As demonstrated in Fig. 2 B & C and E & F), aspirin-PC and aspirin both significantly inhibited the growth of both human and mouse ovarian tumors (p<0.01), with the PC-associated aspirin tending to have a greater growth inhibitory effect than the unmodified NSAID.
In the HeyA8 model, treatment with unmodified aspirin significantly reduced ovarian tumor growth (76.4% reduction in tumor weight vs. control [p<0.01] and 69.1% decrease in the number of tumor nodules vs. control [p<0.05]) (Fig. 2B & E). Aspirin-PC also resulted in profound anti-tumor activity (86.4% reduction in tumor weight vs. control [p<0.01] and 79.4% decrease in the number of tumor nodules vs. control [p<0.01]) (Fig. 2B & E). In this study, Bev also exhibited significant anti-tumor activity (63.6% reduction in tumor weight vs. control [p<0.01] and 58.8% decrease in the number of tumor nodules vs. control [p<0.05]) (Fig. 2B & E). Most interestingly, the combination of aspirin and Bev resulted in a large reduction in tumor weight (91.4% vs. control [p<0.01]) and tumor nodules (85.3% vs. control [p<0.01]) (Fig. 2B & E). In our statistical analysis it was determined that the combination of aspirin plus Bev resulted in a reduction in tumor weight and tumor nodules, compared to either aspirin or Bev alone (p<0.05 tumor weight and number of nodules of combo vs Bev; with the tumor weight and number of nodules of the combo vs aspirin not reaching statistical significance having p values of 0.06 and 0.14 respectively). This potentially synergistic interaction with the VEGF inhibitor, was also observed for aspirin-PC, as the combination of aspirin-PC plus Bev resulted in a large reduction in tumor weight (96.4% vs. control [p<0.01]) and tumor nodules (85.3% vs. control [p<0.01]) (Fig. 2B & E). In our statistical analysis it was determined that the combination of aspirin-PC plus Bev resulted in a significant reduction in tumor weight and nodule number, compared to aspirin-PC or Bev alone (p<0.05). In the HeyA8 study, there are no significant differences between aspirin and aspirin-PC alone and/or in combination with the VEGF inhibitor.
In the ID8 model, aspirin-treatment significantly reduced ovarian tumor growth (49.2% reduction in tumor weight vs. control [p<0.01] and 42.9% decrease in the number of tumor nodules vs. control [p<0.05]) (Fig. 2C & F). Aspirin-PC also possessed highly significant anti-tumor activity (72.3% reduction in tumor weight vs. control [p<0.01] and 73.3% decrease in the number of tumor nodules vs. control [p<0.01]) (Fig. 2C & F). The B20 treatment on its own, exhibited profound anti-tumor activity (73.8% reduction in tumor weight vs. control [p<0.01] and 73.9% decrease in the number of tumor nodules vs. control [p<0.01]) (Fig. 2C & F). The combination of aspirin plus B20 resulted in an even greater reduction in tumor weight (86.2% vs. control [p<0.01]) and tumor nodules (85.1% vs. control [p<0.01]) (Fig. 2C & F). Similar to the effects of aspirin ± VEGF inhibitor, the combination of aspirin-PC plus B20 resulted in a large reduction in tumor weight (96.9% vs. control [p<0.01]) and tumor nodules (91.9% vs. control [p<0.01]) (Fig. 2C & E). In our statistical analysis, it was determined that the combination of aspirin plus B20 resulted in a significant reduction in tumor weight, and nodule number compared to aspirin or B20 alone (p<0.05). Further, the combination of aspirin-PC plus B20 resulted in a significant reduction in tumor weight and number of nodules, compared to aspirin-PC or B20 alone (p<0.05). Of particular note, aspirin-PC demonstrated significantly greater anti-tumor activity than plain aspirin (45.5% reduction in tumor weight [p<0.05] and 53.3% decrease in the number of tumor nodules [p<0.05]). Lastly, aspirin-PC plus B20 demonstrated significantly greater anti-tumor activity than unmodified aspirin plus B20 (77.8% reduction in tumor weight [p<0.01]).
Effect of aspirin-based treatment on tumor vessels, cell proliferation and apoptosis
To examine the biological effects of our test agents alone and in combination, we examined tumor specimens from the HeyA8 mouse model for markers of proliferation (Ki67), apoptosis (cleaved caspase-3), and microvessel density (MVD) (CD31). Representative images of immunohistochemical staining are presented in Fig. 3A. We observed that rates of proliferation were significantly decreased in the treatment groups over the control group (p<0.01). In addition, the combination of aspirin plus Bev resulted in a large reduction in Ki67 expression (37.3% vs. aspirin, p<0.01; 57.4% vs. Bev, p<0.01) (Fig. 3B). Further, the combination of aspirin-PC plus Bev resulted in a large reduction in Ki67 expression (29.6% vs. aspirin-PC, p<0.01; 61.2% vs. Bev, p<0.01) (Fig. 3B). It was also determined that there was a significant difference between aspirin and aspirin-PC in Ki67 expression (p<0.05).
Figure 3. Immunohistochemical and immunofluorescence staining in the HeyA8 model.

(A) Immunohistochemical staining showing the effects of aspirin, Aspirin-PC, Bev and the combination on cell proliferation (Ki67), apoptosis (cleaved caspase 3), and MVD (CD31) in the HeyA8 model. (B) Statistical analysis for Ki67. (C) Statistical analysis for cleaved caspase 3. (D) Statistical analysis for CD31.* p<0.05, ** p<0.01. Original magnification ×200.
The rates of apoptosis were modestly increased in the treatment groups over cleaved Caspase-3 levels of the control group (p<0.01). Interestingly, the combination of aspirin plus Bev resulted in a large increment in cleaved caspase-3 expression (65.3% vs. aspirin, p<0.01; 71.3% vs. Bev, p<0.01) (Fig. 3C). The combination of aspirin-PC plus Bev also resulted in a large increase in the expression of cleaved caspase-3 expression which significantly exceeded the effects of either agent alone (67.9% vs. aspirin-PC, p<0.01; 72.3% vs. Bev, p<0.01) (Fig. 3C).
We observed that the number of tumor micro-vessels scored in fixed/IFC (CD31) stained ovarian tumor tissue was significantly decreased in the treatment groups over in the vehicle-treated control group (p<0.01). In addition, the combination of aspirin or aspirin-PC plus Bev resulted in a large reduction in MVD, thus providing evidence that co-joint therapy of aspirin drugs with a VEGF-A inhibitor is more effective in blocking angiogenesis in ovarian tumor tissue than either agent alone (p<0.01) (Fig. 3D).
We also evaluated the direct inhibitory effect of aspirin and aspirin-PC in vitro on tube formation (over a 6 hr period) using an immortalized cell line (RF24) of human umbilical vein endothelial cells (HUVECs). The results using an aspirin dose of 0.4 mM (which was ~ ID50 dose for ovarian cancer cell growth) showed no effect at 4-days and a modest, but non-significant inhibitory effect on tube formation after 8-days of culture in medium containing our test agents (see Supplementary Material, Fig S-2).
Serum levels of total TGF-β1 and TXB2
We examined serum level of total TGF-β1 and TXB2 in the HeyA8 mouse model and the ID8 mouse models. In the first analysis, we observed that serum levels of total TGF-β1 in the HeyA8 mouse model were significantly decreased in the treatment groups over those measured in the vehicle-treated control group (p<0.01). In addition, the combination of aspirin plus Bev resulted in a large reduction in serum levels of total TGF-β1 (25.5±2.2 vs. 77.2±5.3, p<0.01; 25.5±2.2 vs. 57.7±2.0, p<0.01) (Fig. 4A). A similar synergistic interaction was recorded for the combination of aspirin-PC plus Bev, which resulted in a large reduction in serum TGF-β1 levels (25.3±3.0 vs. 60.1±5.4, p<0.01; 25.3±3.0 vs. 57.7±2.0, p<0.01) (Fig. 4A).
Figure 4. Serum levels of total TGF-β1 and TXB2 in vivo.
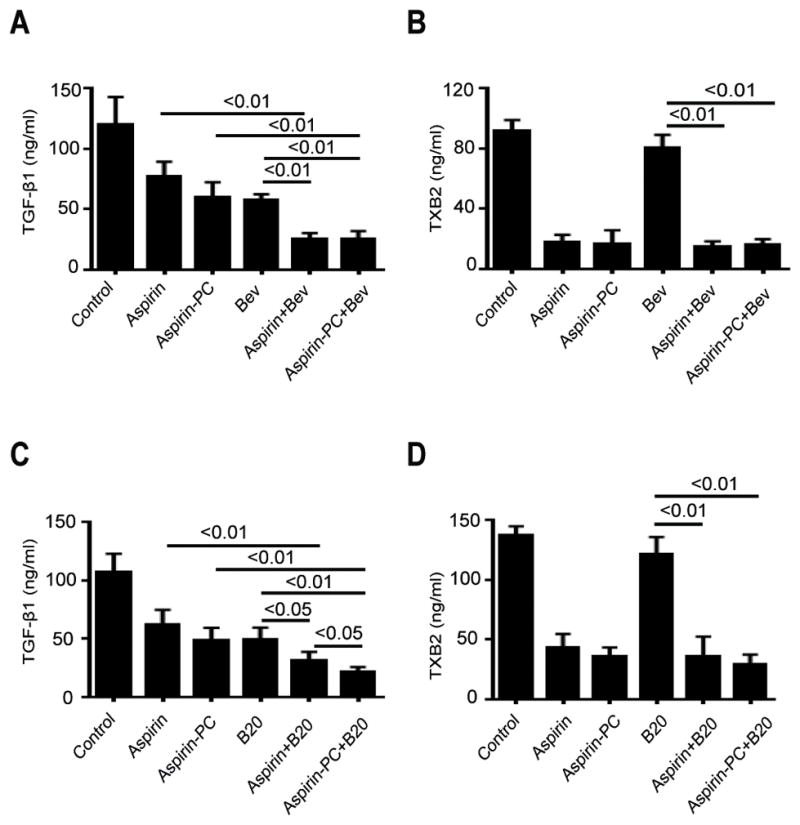
(A, C) Serum total TGF-β1 levels in each group in the HeyA8 model and in the ID8 model were measured by a sandwich ELISA. (B, D) Serum total TXB2 levels in each group in the HeyA8 model and in the ID8 model were measured by a competitive ELISA. * p<0.05, ** p<0.01.
We also determined that serum levels of total TGF-β1 in the ID8 mouse model were significantly decreased in the treatment groups over values of the control group (p<0.01). In addition, the combination of aspirin plus B20 resulted in a further reduction in serum levels of total TGF-β1 (31.2±3.4 vs. 62.1±5.6, p<0.01; 31.2±3.4 vs. 49.4±4.5, p<0.05) (Fig. 4C). Furthermore, it was determined that the combination of Aspirin-PC plus B20 resulted in a large reduction (21.6±1.8 vs. 48.8±4.7, p<0.01; 21.6±1.8 vs. 49.4±4.5, p<0.01) (Fig. 4C), and that there is a significant difference between aspirin plus B20 and aspirin-PC plus B20 on this growth factor (p<0.05).
It was determined that both aspirin and aspirin-PC significantly reduced (p<0.01) serum thromboxane (TXB2) levels >70% in the HeyA8 and ID8 mouse models of ovarian cancer and this profound ant-platelet effect was maintained when the aspirin test drugs were administered in combination with the two VEGF inhibitors (see Fig. 4B & D).
Effect of aspirin-based treatment on the induction of hypoxia in ovarian tumor tissue
Intratumoral hypoxia was assessed by immunohistochemical staining of HeyA8 intraperitoneal tumors using antibody against carbonic anhydrase-9 (CA-9), a downstream target of HIF1a and reliable indicator of hypoxia (31). The results depicted in Figure 5 indicate that tumors treated with aspirin had a significant increase in CA-9 staining compared to vehicle control.
Figure 5. Ovarian tumor tissue hypoxia as assessed by CA-9 immunoreactivity.

Micrographs of ovarian cancer tissue using a immunohistochemical stain for CA-9 from (A) saline-treated controls; (B) mice daily dosed with aspirin; or (C) aspirin-PC. CA-9 staining of HeyA8 tumors, 1:500 (4°C overnight, cat. no. NB100-417, Novus Biologicals). Scale bar: 200μm. (D) Bar graphs represent our quantitation of immunoreactive scores (IRS): of CA-9 staining multiplied by % of cells stained. Statistical analysis: one-way ANOVA with Tukey’s multiple comparison test.
GI injury/bleeding
We assessed the GI bleeding in the in vivo experiments by measuring fecal hemoglobin and confirmed that at the low dose of 20 mg/kg, neither aspirin nor aspirin-PC induced detectable evidence of GI bleeding (see Table 1). These results confirmed the macroscopic inspection of the entire GI tract at necropsy which similarly appeared normal in the aspirin-PC and aspirin treated mice. It should be noted that there was no notable difference in body weight among the test groups at the completion of the period of treatment, and that there was a clear trend that the body weight of the treatment groups were all less than the vehicle-treated control mice, which likely was primarily attributable to the reduction in ovarian tumor mass (see Table 1).
Table 1.
Body weight and fecal hemoglobin measurements from in vivo studies
| SKOV3ip1 cells/nude mice | Body weight (g) | Fecal Hb (mg/g feces) |
|---|---|---|
| Control | 23.25 ± 0.71 | 0.32 |
| Aspirin-PC | 22.74 ± 0.62 | 0.23 |
| Bev | 22.42 ± 0.87 | 0.24 |
| Aspirin-PC/Bev | 21.65 ± 0.45 | 0.28 |
| ID8 cells/C57BL-6 mice | ||
| Control | 23.65 ± 0.62 | 0.34 |
| Aspirin | 21.64 ± 0.32 | 0.34 |
| Aspirin-PC | 21.08 ± 0.52 | 0.34 |
| B20 | 20.00 ± 0.13 | 0.26 |
| Aspirin/B20 | 19.77 ± 0.16 | 0.26 |
| Aspirin-PC/B20 | 19.63 ± 0.22 | 0.29 |
| Hey A8 cells/nude mice | ||
| Control | 26.44 ± 0.53 | 0.95 |
| Aspirin | 25.15 ± 0.65 | 1.75 |
| Aspirin-PC | 24.99 ± 0.64 | 0.91 |
| Bev | 26.84 ± 0.39 | 0.91 |
| Aspirin/Bev | 25.15 ± 0.62 | 0.89 |
| Aspirin-PC/Bev | 24.40 ± 0.67 | 0.93 |
NOTE: Body weight in grams at the termination of the study is expressed as the mean ± standard error of the mean for N=10 mice per group. Fecal hemoglobin (Hb) was measured from pooled samples collected from group cages, and is expressed as mg Hb per gram of feces for each group. For comparison, these Hb values are considered baseline and are much lower than values typically obtained from animals chronically treated with a higher dose NSAID that induces GI bleeding (ie, 2.5 to 15 mg Hb/g feces).
DISCUSSION
One hundred and twenty years after its initial synthesis, the therapeutic utility of aspirin has expanded from reduction of fever, inflammation and pain to the treatment and/or reducing the risk of the development of cardiac disease, stroke thrombosis, diabetes, neurological disease and cancer afflicting numerous tissues (3, 32), which is the focus of the current study. As discussed earlier the evidence supporting the association of aspirin consumption and a reduction in the incidence and mortality is most compelling for colorectal cancer which has led the USPSTF to recommend long-term usage of low-dose aspirin to prevent this common form of cancer. Beyond its use as an effective chemopreventive agent, there is also evidence that aspirin usage may reduce the metastatic spread of the disease suggesting its possible adjunctive use in cancer chemotherapy (5, 12).
Although the evidence supporting the use of aspirin as a chemopreventive agent for cancer and specifically colorectal cancer is compelling, aspirin usage has not been recommended to the general public because of the drugs’ GI side effects, notably gastroduodenal ulcers and bleeding which can be life-threatening in susceptible individuals (33–35). This concern has led to the development of PC-associated aspirin that has been demonstrated to be less injurious to the gastroduodenal mucosa in both animal studies (16, 18–20) and short-term clinical trials (21, 22) due to its ability to preserve the surface hydrophobic barrier properties of the tissue. Animal studies also indicated that this novel family of lipidic NSAIDs maintained their therapeutic activity to reduce fever, inflammation and in the case of aspirin, platelet inhibition, with the suggestion that the PC-associated drug may have more pronounced therapeutic efficacy and potency than the parent NSAID (18, 30, 36). We have also reported that PC-associated aspirin is effective in reducing the growth rate of colon cancer cells in culture (23), and have recently completed a series of (unpublished observations) studies, demonstrating the efficacy of chronic dosing of aspirin-PC to reduce tumor mass in a rodent model of colon cancer. Because of the enormous clinical potential of a GI-safer aspirin, PLx Pharma has developed PL2200 Aspirin, in which aspirin is formulated with a PC-enriched soy lecithin, which has proven safe in long-term (3-month) preclinical rodent toxicological studies and effective in clinical trials (21). The opportunity to evaluate this formulation for cancer prevention/therapy, however remains.
In the current study we provide both in vitro and in vivo evidence that both aspirin and aspirin-PC have efficacy to significantly reduce the growth of human and mouse ovarian cancer cells. Similar to our earlier studies with colon cancer cells in culture (23), our aspirin-PC formulation appeared more potent/efficacious than the unmodified drug in inhibiting the growth of ovarian cancer cells in culture. It should be noted that because of the limitation in administering the orally active lecithin oil-based PL2200aspirin which detaches culture cells from plastic surfaces, the formulation used in the in vitro experiments utilized highly purified soy PC pre-associated with an equimolar mass of aspirin by sonication. This in vitro formulation, which is based upon the earlier PC-NSAID research by the Lichtenberger laboratory (16, 18–20), which did not affect cell attachment to the culture plates, thereby may possess different properties from the oral formulation.
To overcome this limitation, the composition of the Aspirin-PC formulation used in the animal studies was similar to PL2200 Aspirin. As demonstrated in Figures 2 & 3, low dose aspirin-PC proved to be highly effective in significantly reducing ovarian tumor growth in three independent experiments, and was as effective as the VEGF inhibitors. It should be emphasized that at the low dose used (20 mg/kg which corresponds to a clinical aspirin dose of ~100mg), neither aspirin nor aspirin-PC induced any detectable evidence of GI toxicity or bleeding in the treated mice. This observation may not conflict with the clinical experience where aspirin-induced toxicity in human subjects has been generally measured at doses >325 mg/day (35, 37). Remarkably, the combination of aspirin-PC with the VEGF inhibitors reduced the ovarian tumor tissue to levels barely detectable to the observers, “blinded” to the treatment groups. A similar but somewhat less robust anti-neoplastic effect was seen with unmodified aspirin alone and in combination with one of the VEGF blockers in two experiments, one with the human ovarian cell line HeyA8 and the second with the mouse ovarian cell line ID8. Future in vivo dose-response experiments are planned to determine if this robust growth inhibitory effect on ovarian cancer of administering aspirin-PC (or aspirin) in combination with a VEGF monoclonal antibody is an additive or synergistic interaction of these two classes of drugs. We determined that this anti-neoplastic effect was not due to a direct inhibitory effect of VEGF inhibitors on ovarian cancer cells, as we demonstrated that neither B20 nor Bev affected the growth of mouse (ID8) or human (HEYA8) cells within a broad dose range (0.01–125 μg/ml) either alone or in combination with our aspirin test formulations (see Supplemental Material, Figure S-3). Alternatively, we investigated the direct effect of aspirin and aspirin-PC at an ID50 dose of 0.4 mM on HUVEC tube formation and demonstrated that although no effect was recorded at 4-days, a modest but non-significant inhibitory effect was observed after 8-days of culture with our test agents (see Figure S-2). These aspirin drugs likely affect pathways that would indirectly or directly (in the presence of specific co-factors) affect angiogenesis. For example, it has been shown that aspirin may affect the expression of VEGF, Lipoxin A4, angiotensin type-1 receptor (AT1R), thrombospondin -1 (TSP-1) and PPARγ, all of which are regulators of angiogenesis (38).
Our in vivo studies also revealed that both aspirin and aspirin-PC inhibited cell growth and the growth of micro-vessels associated with the ovarian tumor tissue, with both inhibitory actions augmented by co-administration of a VEGF inhibitor. An unexpected but very interesting finding was that apoptotic activity which was modestly elevated by the aspirin test drugs was markedly increased when administered in combination with Bev, although the VEGF-A inhibitor had little effect in promoting cell death on its own.
The rationale for testing our aspirin test drugs in combination with anti-VEGF drugs was based upon our prior work that demonstrated a key role for platelets in stimulating tumor growth following anti-angiogenesis (8). In addition to the known effect of aspirin on activation of platelets, it has been also reported that most NSAIDs, including aspirin at doses above the COX-1 inhibitory anti-platelet dose have the capacity to inhibit COX-2-mediated prostacyclin production, which appears to be involved in the mechanism of hypoxia–induced angiogenic response in endothelial cells (39–41). We therefore evaluated local hypoxia in the tumor micro-environment by assessing CA-9 expression, an established marker of tissue hypoxia (29, 31) and demonstrated evidence of CA-9 expression in tumor bearing controls, which was further exacerbated by aspirin, an effect which was partially reversed in mice treated with aspirin-PC. Thus, our results suggest a COX-dependent inflammatory response in endothelial cells and associated cells supporting the tumor. Interestingly the hypoxic response was exacerbated by aspirin, possibly due to the drug’s ability to reduce micro-vessel growth, whereas aspirin-PC had a lesser effect on tissue hypoxia, which may suggest its use with other anti-neoplastic agents, as tissue hypoxia is thought to be a marker of resistance to chemotherapy (31).
Although we did not observe evidence of increased GI bleeding when aspirin and anti-VEGF agents were used in combination in our experiments, proper cautionary measures should be taken during clinical development.
In support of the possible role of the platelet in these growth responses, we also observed that both aspirin test drugs blocked platelet activation, as demonstrated by the drugs significantly inhibiting by >70% circulating thromboxane (presumably due to irreversible COX-1 inhibition) and by a comparable amount, release of the platelet growth factor, TGFβ1 into the medium. It also should be noted that the aspirin dose (20 mg/kg) administered to the mice in all three studies is comparable to low-dose (75–100 mg) aspirin, and neither aspirin-PC nor the unmodified NSAID induced evidence of GI injury and/or bleeding (see Table 1). Thus, these results suggest the clinical potential of using low-dose Aspirin-PC or aspirin as an adjunctive therapy with chemotherapeutic agents and notably VEGF inhibitors, which may be particularly useful for patients with later-stage ovarian cancer that are not candidates for surgical intervention and are refractory to most chemotherapeutic approaches.
The mechanism by which aspirin-PC and aspirin blocks the growth of ovarian cancer tissue, has yet to be elucidated. As briefly mentioned earlier the concept that COX-2 is over-expressed in most tumor tissue and drives un-regulated cell proliferation and reduced apoptotic activity (3, 42), clearly may not apply to the effect of our test drugs on ovarian cancer that preferentially over-express COX-1 (43–45). Nor does it explain why aspirin which is a weak COX-2 inhibitor has profound chemopreventive activity on ovarian cancer, while the more selective coxibs are less effective. Our cell culture results suggest a direct effect of aspirin on cell growth, likely due to COX-1 inhibition at doses of ≥0.4 mM. It is also possible that the aspirin test formulations may have promoted apoptotic activity of the ovarian cancer cells. However, our previous findings on colon cancer cell lines demonstrated that aspirin had no direct effect on promoting cell death at aspirin concentrations up to 2.5 mM (23). Under in vivo conditions, it is likely that aspirin will be rapidly converted to salicylic acid in circulation, due to the short half-life of the drug in circulation (which has been estimated at 7–12 min in rodents) as a consequence of the presence of de-acetyl hydrolase enzymatic pathway in plasma (46), giving the acetylated drug limited opportunity to inhibit ovarian tissue COX-1. In support of this we have (unpublished obsevations) evidence that aspirin levels in circulation in rodents ranged between 5–10 μM at 10–20 min after intragastric dosing of aspirin at a 5 x higher concentration (100 mg/kg) than the used in the current study, falling to undetectable levels thereafter.
One likely mechanism that aspirin could block the growth and potential metastatic spread of ovarian cancer cell, is by irreversibly blocking platelet activation via its well- known ability to acetylate and thereby inactivate platelet COX-1 (47, 48). Indeed, this would be consistent with the compelling evidence that late stage cancer patients, including those with ovarian cancer have elevated platelet counts (thrombocytosis) that results in a much higher risk for developing deep vein thrombosis than the general population (27, 49). As indicated earlier, we and others have indeed reported that platelets promote cancer cell proliferation, in part by promoting epithelial-mesenchymal transition (EMT), as well as promoting the translocation from the primary tumor into circulation and ultimately from the blood to tissue sites where metastatic growth and angiogenesis occurs, a process facilitated by the local release of platelet derived VEGF, TGFβ and PDGF (7, 25, 27). Thus, aspirin’s unique action to irreversibly inactivate platelets (via acetylation of COX-1) may be a very powerful mechanism to block the progression of cancer, a concept that has been considered by others (3, 5, 50). In support of this possibility, Cho and Stone et al (25, 27), used the orthotopic mouse model to demonstrate that the proliferation of human ovarian tissue can be promoted by the infusion of fresh platelets. Furthermore, mouse platelets have been reported to extravasate into the ovarian tumor tissue, as demonstrated with a specific immunohistochemical stain using an antibody directed against a platelet surface protein and genetically engineered fluorescently labelled platelets (27). Future studies will be directed at determining if the platelet indeed is the primary target of aspirin’s anti-neoplastic action and if PC-association provides an advantage for aspirin to target and inactivate the platelet.
Supplementary Material
Acknowledgments
This research was supported, in part, by grants from the NIH (CA109298, P50 CA083639, P50 CA098258, UH3 TR000943, and CA177909 to A.K.Sood; CA016672, to MD Anderson Cancer Center), the Ovarian Cancer Research Fund, Inc. (Program Project Development Grant; to A. K.Sood), the Blanton-Davis Ovarian Cancer Research Program and The Judi Rees Fund; to A.K.Sood, and the Frank McGraw Memorial Chair in Cancer Research; to A. K.Sood). RAP and JMH were supported by a T32 Training Grant (T32CA101642; to A.K.Sood). WMJ was supported by a T32 Training grant (T32 CA009666; to R. Champlin).
Footnotes
Disclosure of Potential Conflicts of Interest
L.M. Lichtenberger is the Scientific Founder and shareholder in PLx Pharma Inc, which is developing (Aspirin-PC) PL2200 Aspirin.
References
- 1.Cuzick J, Thorat MA, Bosetti C, Brown PH, Burn J, Cook NR, et al. Estimates of benefits and harms of prophylactic use of aspirin in the general population. Ann Oncol. 2015;26:47–57. doi: 10.1093/annonc/mdu225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Chan AT, Arber N, Burn J, Chia WK, Elwood P, Hull MA, et al. Aspirin in the chemoprevention of colorectal neoplasia: an overview. Cancer Prev Res (Phila) 2012;5:164–78. doi: 10.1158/1940-6207.CAPR-11-0391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Thun MJ, Jacobs EJ, Patrono C. The role of aspirin in cancer prevention. Nat Rev Clin Oncol. 2012;9:259–67. doi: 10.1038/nrclinonc.2011.199. [DOI] [PubMed] [Google Scholar]
- 4.Rothwell PM, Price JF, Fowkes FG, Zanchetti A, Roncaglioni MC, Tognoni G, et al. Short-term effects of daily aspirin on cancer incidence, mortality, and non-vascular death: analysis of the time course of risks and benefits in 51 randomised controlled trials. Lancet. 2012;379:1602–12. doi: 10.1016/S0140-6736(11)61720-0. [DOI] [PubMed] [Google Scholar]
- 5.Rothwell PM, Wilson M, Price JF, Belch JF, Meade TW, Mehta Z. Effect of daily aspirin on risk of cancer metastasis: a study of incident cancers during randomised controlled trials. Lancet. 2012;379:1591–601. doi: 10.1016/S0140-6736(12)60209-8. [DOI] [PubMed] [Google Scholar]
- 6.Huang L, Cronin KA, Johnson KA, Mariotto AB, Feuer EJ. Improved survival time: what can survival cure models tell us about population-based survival improvements in late-stage colorectal, ovarian, and testicular cancer? Cancer. 2008;112:2289–300. doi: 10.1002/cncr.23425. [DOI] [PubMed] [Google Scholar]
- 7.Davis AN, Afshar-Kharghan V, Sood AK. Platelet effects on ovarian cancer. Semin Oncol. 2014;41:378–84. doi: 10.1053/j.seminoncol.2014.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Haemmerle M, Bottsford-Miller J, Pradeep S, Taylor ML, Choi HJ, Hansen JM, et al. FAK regulates platelet extravasation and tumor growth after antiangiogenic therapy withdrawal. J Clin Invest. 2016;126:1885–96. doi: 10.1172/JCI85086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Trabert B, Ness RB, Lo-Ciganic WH, Murphy MA, Goode EL, Poole EM, et al. Aspirin, nonaspirin nonsteroidal anti-inflammatory drug, and acetaminophen use and risk of invasive epithelial ovarian cancer: a pooled analysis in the Ovarian Cancer Association Consortium. J Natl Cancer Inst. 2014;106:djt431. doi: 10.1093/jnci/djt431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Baandrup L, Kjaer SK, Olsen JH, Dehlendorff C, Friis S. Low-dose aspirin use and the risk of ovarian cancer in Denmark. Ann Oncol. 2015;26:787–92. doi: 10.1093/annonc/mdu578. [DOI] [PubMed] [Google Scholar]
- 11.Ni X, Ma J, Zhao Y, Wang Y, Wang S. Meta-analysis on the association between non-steroidal anti-inflammatory drug use and ovarian cancer. Br J Clin Pharmacol. 2013;75:26–35. doi: 10.1111/j.1365-2125.2012.04290.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Fraser DM, Sullivan FM, Thompson AM, McCowan C. Aspirin use and survival after the diagnosis of breast cancer: a population-based cohort study. Br J Cancer. 2014;111:623–7. doi: 10.1038/bjc.2014.264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Laine L, Takeuchi K, Tarnawski A. Gastric mucosal defense and cytoprotection: bench to bedside. Gastroenterology. 2008;135:41–60. doi: 10.1053/j.gastro.2008.05.030. [DOI] [PubMed] [Google Scholar]
- 14.Lichtenberger LM. Role of phospholipids in protection of the GI mucosa. Dig Dis Sci. 2013;58:891–3. doi: 10.1007/s10620-012-2530-8. [DOI] [PubMed] [Google Scholar]
- 15.Lichtenberger LM. The hydrophobic barrier properties of gastrointestinal mucus. Annu Rev Physiol. 1995;57:565–83. doi: 10.1146/annurev.ph.57.030195.003025. [DOI] [PubMed] [Google Scholar]
- 16.Lichtenberger LM, Wang ZM, Romero JJ, Ulloa C, Perez JC, Giraud MN, et al. Non-steroidal anti-inflammatory drugs (NSAIDs) associate with zwitterionic phospholipids: insight into the mechanism and reversal of NSAID-induced gastrointestinal injury. Nat Med. 1995;1:154–8. doi: 10.1038/nm0295-154. [DOI] [PubMed] [Google Scholar]
- 17.Lichtenberger LM, Zhou Y, Jayaraman V, Doyen JR, O’Neil RG, Dial EJ, et al. Insight into NSAID-induced membrane alterations, pathogenesis and therapeutics: characterization of interaction of NSAIDs with phosphatidylcholine. Biochim Biophys Acta. 2012;1821:994–1002. doi: 10.1016/j.bbalip.2012.04.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lichtenberger LM, Barron M, Marathi U. Association of phosphatidylcholine and NSAIDs as a novel strategy to reduce gastrointestinal toxicity. Drugs Today (Barc) 2009;45:877–90. doi: 10.1358/dot.2009.45.12.1441075. [DOI] [PubMed] [Google Scholar]
- 19.Lichtenberger LM, Romero JJ, Dial EJ. Surface phospholipids in gastric injury and protection when a selective cyclooxygenase-2 inhibitor (Coxib) is used in combination with aspirin. Br J Pharmacol. 2007;150:913–9. doi: 10.1038/sj.bjp.0707176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Lim YJ, Dial EJ, Lichtenberger LM. Advent of novel phosphatidylcholine-associated nonsteroidal anti-inflammatory drugs with improved gastrointestinal safety. Gut Liver. 2013;7:7–15. doi: 10.5009/gnl.2013.7.1.7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Cryer B, Bhatt DL, Lanza FL, Dong JF, Lichtenberger LM, Marathi UK. Low-dose aspirin-induced ulceration is attenuated by aspirin-phosphatidylcholine: a randomized clinical trial. Am J Gastroenterol. 2011;106:272–7. doi: 10.1038/ajg.2010.436. [DOI] [PubMed] [Google Scholar]
- 22.Lanza FL, Marathi UK, Anand BS, Lichtenberger LM. Clinical trial: comparison of ibuprofen-phosphatidylcholine and ibuprofen on the gastrointestinal safety and analgesic efficacy in osteoarthritic patients. Aliment Pharmacol Ther. 2008;28:431–42. doi: 10.1111/j.1365-2036.2008.03765.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Dial EJ, Doyen JR, Lichtenberger LM. Phosphatidylcholine-associated nonsteroidal anti-inflammatory drugs (NSAIDs) inhibit DNA synthesis and the growth of colon cancer cells in vitro. Cancer Chemother Pharmacol. 2006;57:295–300. doi: 10.1007/s00280-005-0048-x. [DOI] [PubMed] [Google Scholar]
- 24.Pecot CV, Rupaimoole R, Yang D, Akbani R, Ivan C, Lu C, et al. Tumour angiogenesis regulation by the miR-200 family. Nat Commun. 2013;4:2427. doi: 10.1038/ncomms3427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Cho MS, Bottsford-Miller J, Vasquez HG, Stone R, Zand B, Kroll MH, et al. Platelets increase the proliferation of ovarian cancer cells. Blood. 2012;120:4869–72. doi: 10.1182/blood-2012-06-438598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kim TJ, Landen CN, Lin YG, Mangala LS, Lu C, Nick AM, et al. Combined anti-angiogenic therapy against VEGF and integrin alphaVbeta3 in an orthotopic model of ovarian cancer. Cancer Biol Ther. 2009;8:2263–72. doi: 10.4161/cbt.8.23.10134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Stone RL, Nick AM, McNeish IA, Balkwill F, Han HD, Bottsford-Miller J, et al. Paraneoplastic thrombocytosis in ovarian cancer. N Engl J Med. 2012;366:610–8. doi: 10.1056/NEJMoa1110352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Huang J, Hu W, Bottsford-Miller J, Liu T, Han HD, Zand B, et al. Cross-talk between EphA2 and BRaf/CRaf is a key determinant of response to Dasatinib. Clin Cancer Res. 2014;20:1846–55. doi: 10.1158/1078-0432.CCR-13-2141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Tanaka N, Kato H, Inose T, Kimura H, Faried A, Sohda M, et al. Expression of carbonic anhydrase 9, a potential intrinsic marker of hypoxia, is associated with poor prognosis in oesophageal squamous cell carcinoma. Br J Cancer. 2008;99:1468–75. doi: 10.1038/sj.bjc.6604719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Lichtenberger LM, Romero JJ, de Ruijter WM, Behbod F, Darling R, Ashraf AQ, et al. Phosphatidylcholine association increases the anti-inflammatory and analgesic activity of ibuprofen in acute and chronic rodent models of joint inflammation: relationship to alterations in bioavailability and cyclooxygenase-inhibitory potency. J Pharmacol Exp Ther. 2001;298:279–87. [PubMed] [Google Scholar]
- 31.Rupaimoole R, Wu SY, Pradeep S, Ivan C, Pecot CV, Gharpure KM, et al. Hypoxia-mediated downregulation of miRNA biogenesis promotes tumour progression. Nat Commun. 2014;5:5202. doi: 10.1038/ncomms6202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Cuzick J, Otto F, Baron JA, Brown PH, Burn J, Greenwald P, et al. Aspirin and non-steroidal anti-inflammatory drugs for cancer prevention: an international consensus statement. Lancet Oncol. 2009;10:501–7. doi: 10.1016/S1470-2045(09)70035-X. [DOI] [PubMed] [Google Scholar]
- 33.Barkin RL, Beckerman M, Blum SL, Clark FM, Koh EK, Wu DS. Should nonsteroidal anti-inflammatory drugs (NSAIDs) be prescribed to the older adult? Drugs Aging. 2010;27:775–89. [Google Scholar]
- 34.Wallace JL. Nonsteroidal anti-inflammatory drugs and gastroenteropathy: the second hundred years. Gastroenterology. 1997;112:1000–16. doi: 10.1053/gast.1997.v112.pm9041264. [DOI] [PubMed] [Google Scholar]
- 35.Wolfe MM, Lichtenstein DR, Singh G. Gastrointestinal toxicity of nonsteroidal antiinflammatory drugs. N Engl J Med. 1999;340:1888–99. doi: 10.1056/NEJM199906173402407. [DOI] [PubMed] [Google Scholar]
- 36.Lichtenberger LM, Ulloa C, Vanous AL, Romero JJ, Dial EJ, Illich PA, et al. Zwitterionic phospholipids enhance aspirin’s therapeutic activity, as demonstrated in rodent model systems. J Pharmacol Exp Ther. 1996;277:1221–7. [PubMed] [Google Scholar]
- 37.Goldstein JL, Lowry SC, Lanza FL, Schwartz HI, Dodge WE. The impact of low-dose aspirin on endoscopic gastric and duodenal ulcer rates in users of a non-selective non-steroidal anti-inflammatory drug or a cyclo-oxygenase-2-selective inhibitor. Aliment Pharmacol Ther. 2006;23:1489–98. doi: 10.1111/j.1365-2036.2006.02912.x. [DOI] [PubMed] [Google Scholar]
- 38.Usman MW, Luo F, Cheng H, Zhao JJ, Liu P. Chemopreventive effects of aspirin at a glance. Biochim Biophys Acta. 2015;1855:254–63. doi: 10.1016/j.bbcan.2015.03.007. [DOI] [PubMed] [Google Scholar]
- 39.Cheng Y, Austin SC, Rocca B, Koller BH, Coffman TM, Grosser T, et al. Role of prostacyclin in the cardiovascular response to thromboxane A2. Science. 2002;296:539–41. doi: 10.1126/science.1068711. [DOI] [PubMed] [Google Scholar]
- 40.Fitzgerald GA. Coxibs and cardiovascular disease. N Engl J Med. 2004;351:1709–11. doi: 10.1056/NEJMp048288. [DOI] [PubMed] [Google Scholar]
- 41.Mitchell JA, Warner TD. COX isoforms in the cardiovascular system: understanding the activities of non-steroidal anti-inflammatory drugs. Nat Rev Drug Discov. 2006;5:75–86. doi: 10.1038/nrd1929. [DOI] [PubMed] [Google Scholar]
- 42.Cha YI, DuBois RN. NSAIDs and cancer prevention: targets downstream of COX-2. Annu Rev Med. 2007;58:239–52. doi: 10.1146/annurev.med.57.121304.131253. [DOI] [PubMed] [Google Scholar]
- 43.Gupta RA, Tejada LV, Tong BJ, Das SK, Morrow JD, Dey SK, et al. Cyclooxygenase-1 is overexpressed and promotes angiogenic growth factor production in ovarian cancer. Cancer Res. 2003;63:906–11. [PubMed] [Google Scholar]
- 44.Daikoku T, Wang D, Tranguch S, Morrow JD, Orsulic S, DuBois RN, et al. Cyclooxygenase-1 is a potential target for prevention and treatment of ovarian epithelial cancer. Cancer Res. 2005;65:3735–44. doi: 10.1158/0008-5472.CAN-04-3814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Cho M, Kabir SM, Dong Y, Lee E, Rice VM, Khabele D, et al. Aspirin Blocks EGF-stimulated Cell Viability in a COX-1 Dependent Manner in Ovarian Cancer Cells. J Cancer. 2013;4:671–8. doi: 10.7150/jca.7118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Kapetanovic IM, Bauer KS, Tessier DM, Lindeblad MO, Zakharov AD, Lubet R, et al. Comparison of pharmacokinetic and pharmacodynamic profiles of aspirin following oral gavage and diet dosing in rats. Chem Biol Interact. 2009;179:233–9. doi: 10.1016/j.cbi.2008.10.008. [DOI] [PubMed] [Google Scholar]
- 47.Vane J. Towards a better aspirin. Nature. 1994;367:215–6. doi: 10.1038/367215a0. [DOI] [PubMed] [Google Scholar]
- 48.Vane JR. Inhibition of prostaglandin synthesis as a mechanism of action for aspirin-like drugs. Nat New Biol. 1971;231:232–5. doi: 10.1038/newbio231232a0. [DOI] [PubMed] [Google Scholar]
- 49.Timp JF, Braekkan SK, Versteeg HH, Cannegieter SC. Epidemiology of cancer-associated venous thrombosis. Blood. 2013;122:1712–23. doi: 10.1182/blood-2013-04-460121. [DOI] [PubMed] [Google Scholar]
- 50.Dovizio M, Tacconelli S, Sostres C, Ricciotti E, Patrignani P. Mechanistic and pharmacological issues of aspirin as an anticancer agent. Pharmaceuticals (Basel) 2012;5:1346–71. doi: 10.3390/ph5121346. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.