

Abstract
Objective. Obesity is a well-known risk factor for thrombotic complications. The aim of the present study was to determine the frequency of thrombosis related ABO, F5, MTHFR, and FGG gene polymorphisms in morbidly obese patients and compare them with the group of nonobese individuals. Methods. Gene polymorphisms were analyzed in 320 morbidly obese patients (BMI > 40 kg/m2) and 303 control individuals (BMI < 30 kg/m2) of European descent. ABO C>T (rs505922), F5 C>G (rs6427196), MTHFR C>T (rs1801133), and FGG C>T (rs6536024) SNPs were genotyped by RT-PCR. Results. We observed a tendency for MTHFR rs1801133 TT genotype to be linked with morbid obesity when compared to CC genotype; however, the difference did not reach the significant P value (OR 1.84, 95% CI 0.83–4.05, P = 0.129). Overall, the genotypes and alleles of rs505922, rs6427196, rs1801133, and rs6536024 SNPs had similar distribution between morbidly obese and nonobese control individuals. Distribution of height and weight means among individuals carrying different rs505922, rs6427196, rs1801133, and rs6536024 genotypes did not differ significantly. Conclusions. Gene polymorphisms ABO C>T (rs505922), F5 C>G (rs6427196), MTHFR C>T (rs1801133), and FGG C>T (rs6536024) were not associated with height, weight, or morbid obesity among European subjects.
1. Introduction
Obesity has become one of the major health care challenges worldwide. It is linked with multiple medical conditions, significantly affecting longevity and quality of life [1]. Obesity is a well-known risk factor for developing deep venous thrombosis (DVT) [2]. Epidemiological studies show a clear link between body mass index (BMI) and the risk of DVT or related conditions [3, 4]. BMI is one of the major determinants of patients' outcomes in health care emergencies and elective surgery [5]. Development of thrombotic events in obese individuals is also associated with longer hospital admissions and mortality rates [6, 7]. More recently, a number of important research papers have linked the length of legs [8] and height [9] with recurrent venous thrombosis or pulmonary embolism [10]. To date, there are multiple prophylactic antithrombotic regiments; however, risk stratification strategies for prevention of DVT based on anthropometric data need to be improved [11].
Thrombosis and obesity are complex epidemiologically associated diseases, but the mechanism of this association is not yet understood [12]. Development of DVT in obese individuals is thought to result from a complex interaction of host and environmental factors [13]. The pathogenesis of DVT has been linked with chronic low grade inflammation, heritability, diet, physical activity, and other potential risk factors [13]. Recent advances in molecular genotyping techniques outlined the importance of genetic factors for development of thrombosis [14]. It is estimated that more than 60% of the variation in susceptibility to common thrombosis might be attributable to genetic factors [15].
To date, there are several genome wide association studies (GWAS) that have linked various genetic factors with the risk of developing thrombotic complications. Trégouët et al. conducted a GWAS by analyzing approximately 317,000 single nucleotide polymorphisms (SNPs) in 453 venous thromboembolism (VTE) cases and 1327 controls and found that three SNPs located in the F5 and ABO blood group genes were associated with VTE at a genome wide significant level [16]. Another comprehensive genome wide association analysis, testing 336,469 SNPs in 13,974 healthy Caucasian women, confirmed the association of MTHFR (rs1801133) and CBS (rs6586282) SNPs with homocysteine levels that have been linked with thrombotic events [17]. A large GWAS including nearly 45,000 individuals reported key genetic associations in F5, ABO, and FGG loci for VTE [18]. The same loci at F5, ABO, and FGG have been linked with VTE in another study including 1,542 cases and 1,110 controls [19]. The results of these GWAS studies have been replicated in smaller case-control studies [20]; however, the frequencies of these genetic variations have not been assessed in morbidly obese patients. Furthermore, they have not been previously analyzed in relation to height and weight.
Souto et al. have shown that BMI and thrombosis are genetically linked [12]. They showed that both venous and arterial thromboembolic disease and BMI had a significant genetic correlation. A Danish study observed a strong observational association between obesity and DVT with or without pulmonary embolism (PE), supported by a direct genetic association between the obesity-specific genetic loci and DVT with PE [21]. Studies discussed above clearly imply that obesity might likely be causally associated with DVT.
The aim of our present study was to determine the frequencies of thrombosis related ABO, F5, MTHFR, and FGG gene polymorphisms in morbidly obese patients and compare them with the group of nonobese individuals. Frequencies of ABO C>T (rs505922), F5 C>G (rs6427196), MTHFR C>T (rs1801133), and FGG C>T (rs6536024) gene polymorphisms have not been previously evaluated in morbidly obese patients. We also aimed to evaluate whether the genotypes of above-mentioned gene polymorphism are linked with height or weight of study individuals. Here, in this study we performed ABO, F5, MTHFR, and FGG SNP genotyping analysis in 320 morbidly obese patients (BMI > 40 kg/m2) and 303 control nonobese individuals (BMI < 30 kg/m2) of European descent.
2. Materials and Methods
2.1. Study Population
The group of morbidly obese subjects consisted of patients referred for elective bariatric surgery with a BMI >40 kg/m2. Control subjects were healthy individuals with BMI <30 kg/m2, who came from our previous genotyping studies [22, 23]. Morbidly obese patients and controls were recruited during the years 2011–2015 in the Departments of Surgery and Gastroenterology, Lithuanian University of Health Sciences (Kaunas, Lithuania). The inclusion criteria for control group were no previous history of malignancy, VTE, and BMI <30 kg/m2. In total, 623 individuals (303 controls and 320 morbidly obese patients) were included in the genotyping study. All patients were of European ethnicity. The study was approved by Kaunas Regional Ethics Committee (Protocol number BE-2-10). All patients have signed an informed consent form to participate in this genotyping case-control study.
2.2. DNA Extraction and Genotyping
Genomic DNA of study participants was obtained from peripheral blood mononuclear cells using phenol-chloroform extraction method as described previously [22]. DNA aliquots were stored at −20°C until RT-PCR analysis. SNPs of ABO T>C (rs505922), F5 C>G (rs6427196), MTHFR G>A (rs1801133), and FGG T>C (rs6536024) were genotyped using custom TaqMan® assays with a 7500™ real-time cycler according to manufacturer's instructions (Life Technologies, CA, USA). Thermal cycling conditions for polymerase chain reaction (PCR) were, first, denaturing at 95°C for 10 min, followed by 40 cycles of 95.5°C for 15 s and 60°C for 1 min. Alleles and genotypes of analyzed SNPs were determined with SDS 2.0.5 software compatible.
2.3. Genotyping Quality Control
5% of samples for each of the four SNPs were selected for repetitive analysis. Replication experiments revealed a 100% concordance rate of genotypes and alleles with the initial genotyping results.
2.4. Statistical Analysis
All study participants were stratified into two groups: 320 morbidly obese patients (BMI > 40 kg/m2) and 303 control nonobese individuals (BMI < 30 kg/m2). Age, height, weight, and BMI are presented as mean and standard deviations and was compared using unpaired Student's t-test. Categorical data (gender, distribution of genotypes or alleles) are presented as frequencies; comparisons were performed using the Chi-square test. Quality assessments and statistical analysis of genotyping data were carried out using free PLINK software (version 1.07) for genetic analysis [24]. Association of morbid obesity with gene polymorphisms was calculated using logistic regression analysis with adjustment for age and gender and presented as adjusted odds ratios (aOR) with 95% confidence intervals (CI). The relative risks for SNP genotypes and alleles were studied using recessive and dominant models that led to a comparison between wild type + heterozygous versus homozygous and wild type versus heterozygous + homozygous, respectively. Due to multiple association calculations we introduced an adjusted significance threshold for multiple comparisons α = 0.0125 (0.05/4). One way ANOVA or unpaired t-test was used to compare height and weight differences between different genotypes of each SNP.
3. Results
3.1. Characteristics of the Subjects
Characteristics of control and morbidly obese patient groups are presented in Table 1. In total 623 individuals participated in the study (320 morbidly obese and 303 control subjects). Individuals in the control group were significantly older than morbid obesity group subjects, 61.5 and 42.6 years, respectively (P < 0.001). Males accounted for 60.9% in a group of patients with BMI >40 kg/m2, while in the control group they constituted 42.6% (P < 0.001). Evidently, mean BMI in morbidly obese group was significantly higher (46.0 kg/m2) than compared to control group (25.1 kg/m2, P < 0.001). Since proportion of males and females as well as age were significantly different between the two groups, gender and age were included as confounding factors in further logistic regression analysis of genotyping results.
Table 1.
Characteristics of study groups.
| Morbid obesity group (n = 320) | Control group (n = 303) | P value | |
|---|---|---|---|
| Gender (n, %) | |||
| Males | 195 (60.9) | 129 (42.6) | <0.001 |
| Females | 125 (39.1) | 174 (57.4) | |
| Age (years) | |||
| Mean ± SD | 42.6 ± 11.2 | 61.5 ± 8.2 | <0.001 |
| BMI (kg/m2) | |||
| Mean ± SD | 46.0 ± 4.2 | 25.1 ± 2.7 | <0.001 |
BMI, body mass index.
3.2. Hardy-Weinberg Equilibrium
All four ABO T>C (rs505922), F5 C>G (rs6427196), MTHFR G>A (rs1801133), and FGG T>C (rs6536024) were tested for Hardy-Weinberg equilibrium. The results of the analysis are presented in Table 2. The conditions of Hardy-Weinberg equilibrium were fulfilled for all of investigated SNPs as the frequencies of observed and expected genotype and allele frequencies did not differ: rs505922, P = 0.241; rs6427196, P = 1; rs1801133, P = 0.235; rs6536024, P = 0.418 (Table 2).
Table 2.
Analysis of Hardy-Weinberg equilibrium.
| SNP | Allele frequencies | Genotype distribution | Determined frequency of heterozygous allele | Expected frequency of heterozygous allele | P value | |
|---|---|---|---|---|---|---|
| rs1801133 | T (0.281) | C (0.719) | 43/264/315 | 0.424 | 0.404 | 0.235 |
| rs6427196 | C (0.052) | G (0.948) | 1/63/559 | 0.101 | 0.099 | 1 |
| rs6536024 | C (0.343) | T (0.657) | 20/319/184 | 0.512 | 0.495 | 0.418 |
| rs505922 | C (0.396) | T (0.604) | 90/313/220 | 0.502 | 0.478 | 0.241 |
SNP, single nucleotide polymorphism.
3.3. Frequencies of rs505922, rs6427196, rs1801133, and rs6536024 Genotypes and Allele
Genotype and allele distributions for ABO T>C (rs505922), F5 C>G (rs6427196), MTHFR G>A (rs1801133), and FGG T>C (rs6536024) in morbid obesity and control groups are presented in Table 3. All individuals were successfully genotyped for rs505922, rs6427196, and rs6536024 loci, while one individual in morbid obesity group failed genotyping for rs1801133. Overall, the genotypes and alleles of rs505922, rs6427196, rs1801133, and rs6536024 SNPs had similar distribution between morbidly obese and nonobese control individuals. We observed a tendency for MTHFR rs1801133 TT genotype to be linked with morbid obesity when compared to CC genotype; however, the difference did not reach the significant P value (odds ratio (OR), 1.84, 95% confidence interval (CI) 0.83–4.05, and P = 0.129, Table 3). Similar results were obtained in a recessive model for MTHFR SNP (TT versus CT + CC), but association remained beyond statistical significance (OR 1.81, 95% CI 0.83–3.90, and P = 0.131, Table 3).
Table 3.
Genotype and allele frequencies of ABO C>T (rs505922), F5 C>G (rs6427196), MTHFR C>T (rs1801133), and FGG C>T (rs6536024) SNPs in morbidly obese and nonobese control individuals.
| Alleles/genotypes | Morbid obesity group (n = 320) | Control group (n = 303) | aOR | 95% CI | P |
|---|---|---|---|---|---|
| n, % | n, % | ||||
| rs505922 ABO C>T |
|||||
| T | 380 (59.4) | 373 (61.6) | |||
| C | 260 (40.6) | 233 (38.4) | 1.10 | (0.877–1.38) | 0.406 |
| TT | 107 (35.3) | 113 (37.9) | — | ||
| TC | 166 (48.2) | 147 (47.3) | 1.26 | (0.834–1.90) | 0.273 |
| CC | 47 (16.5) | 43 (14.7) | 1.06 | (0.593–1.91) | 0.836 |
| CC + TC versus TT | 1.21 | (0.819–1.79) | 0.337 | ||
| CC versus TC + TT | 0.930 | (0.546–1.58) | 0.788 | ||
| rs6427196 F5 C>G |
|||||
| G | 610 (95.3) | 571 (94.2) | |||
| C | 30 (4.7) | 35 (5.8) | 0.805 | (0.488–1.32) | 0.395 |
| GG | 291 (90.8) | 268 (88.7) | — | ||
| GC | 28 (9.0) | 35 (10.9) | 0.605 | (0.325–1.13) | 0.112 |
| CC | 1 (0.2) | 0 (0) | 1.82 × 108 | (0–inf) | 0.999 |
| CC + GC versus GG | 0.613 | (0.330–1.14) | 0.120 | ||
| CC versus GC + GG | 1.95 × 108 | (0–inf) | 0.999 | ||
| rs1801133 MTHFR C>T |
|||||
| C | 447 (70.1) | 447 (73.8) | |||
| T | 191 (29.9) | 159 (26.2) | 1.201 | (0.938–1.54) | 0.147 |
| CC | 156 (49) | 159 (54.5) | — | ||
| CT | 135 (42) | 129 (38.7) | 1.043 | (0.706–1.54) | 0.832 |
| TT | 28 (8.9) | 15 (6.9) | 1.842 | (0.837–4.05) | 0.129 |
| TT + TC versus CC | 1.127 | (0.775–1.64) | 0.532 | ||
| TT versus CT + CC | 1.808 | (0.838–3.90) | 0.131 | ||
| rs6536024 FGG C>T |
|||||
| T | 355 (55.5) | 532 (66) | |||
| C | 285 (44.5) | 274 (34) | 0.972 | (0.777–1.22) | 0.804 |
| TT | 100 (30.8) | 184 (43.6) | — | ||
| TC | 155 (49.4) | 164 (44.9) | 0.796 | (0.516–1.23) | 0.303 |
| CC | 65 (19.8) | 55 (11.6) | 1.14 | (0.660–1.97) | 0.638 |
| CC + TC versus TT | 0.881 | (0.585–1.33) | 0.545 | ||
| CC versus TC + TT | 1.31 | (0.817–2.11) | 0.260 |
aOR, adjusted odds ratio by sex and age; CI, confidence interval.
3.4. Distribution of Height and Weight Means for rs1801133, rs6427196, rs505922, and rs6536024 Genotypes
Distribution of height and weight means for ABO T>C (rs505922), F5 C>G (rs6427196), MTHFR G>A (rs1801133), and FGG T>C (rs6536024) genotypes for all subjects included in the study is presented in Figure 1. Additionally, one way ANOVA analysis was performed to compare height and weight means distribution between single SNP genotypes separately (Table 4). Rs6427196 analysis showed only one subject which was recognized as genotype CC; therefore, this group was excluded from analysis; height and weight means were compared only for CG and GG genotypes using t-test. The results show that height and weight means were similar when compared between different SNPs genotypes (Table 4; Figure 1).
Figure 1.
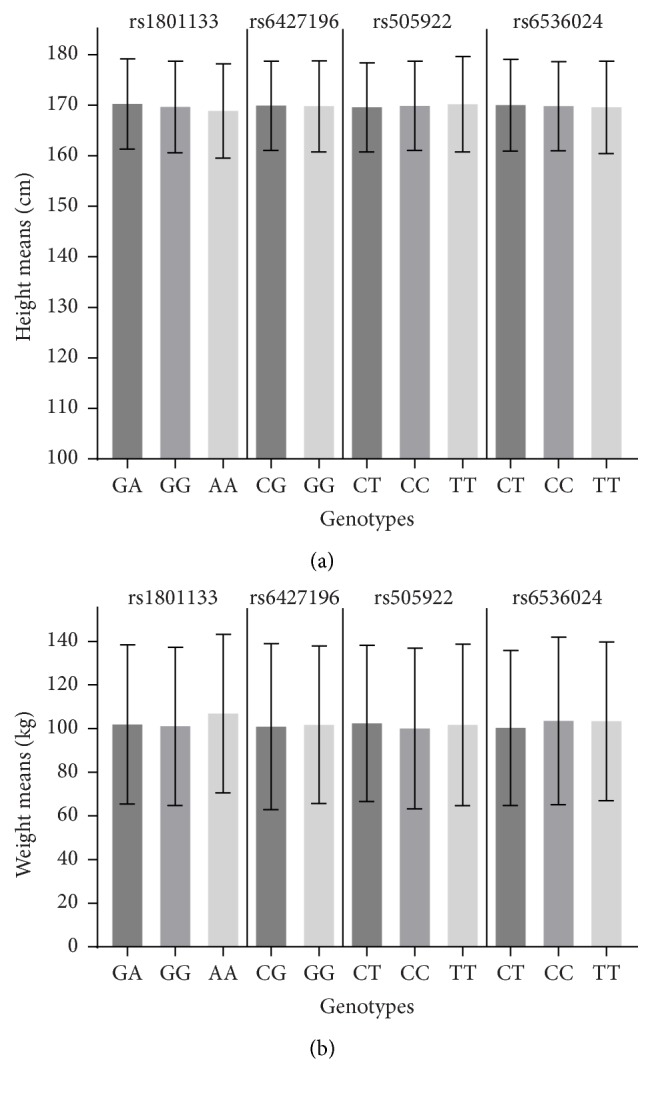
(a) Mean height by SNP (rs505922, rs6427196, rs1801133, and rs6536024) genotypes; (b) mean weight by SNP (rs505922, rs6427196, rs1801133, and rs6536024) genotypes. Error bars indicate the standard deviation in (a) cm and (b) kg in each group. In rs6427196 analysis only one subject was identified as having genotype CC and, therefore, was excluded from analysis.
Table 4.
Mean height and weight by SNPs genotypes (rs1801133, rs6427196, rs505922, and rs6536024).
| SNPs | Means ± SD by genotype | P |
|---|---|---|
| rs505922 ABO C>T |
||
| Height (CT/CC/TT) | 169.92 ± 9.29/170.67 ± 9.36/171.75 ± 9.05 | 0.7551 |
| Weight (CT/CC/TT) | 132.35 ± 24.97/132.95 ± 23.86/135.29 ± 24.67 | 0.8661 |
| rs64271963
F5 C>G |
||
| Height (CG/GG) | 170.04 ± 8.52/170.63 ± 9.22 | 0.9372 |
| Weight (CG/GG) | 137.08 ± 30.27/132.95 ± 24.02 | 0.8532 |
| rs1801133 MTHFR C>T |
||
| Height (GA/GG/AA) | 171.18 ± 9.14/170.41 ± 9.08/169.60 ± 10.63 | 0.5801 |
| Weight (GA/GG/AA) | 133.98 ± 26.28/133.77 ± 23.11/129.16 ± 26.35 | 0.6331 |
| rs6536024 FGG C>T |
||
| Height (CT/CC/TT) | 170.83 ± 9.38/171.35 ± 9.11/169.93 ± 9.11 | 0.8701 |
| Weight (CT/CC/TT) | 133.32 ± 23.72/135.53 ± 25.10/132.33 ± 26.04 | 0.5751 |
1One way ANOVA P value; 2unpaired t-test P value; 3In rs6427196 analysis only one subject was identified as having genotype CC and, therefore, was excluded from analysis.
4. Discussion
In our study we analyzed ABO C>T (rs505922), F5 C>G (rs6427196), MTHFR C>T (rs1801133), and FGG C>T (rs6536024) gene polymorphisms in a case-control study including 320 morbidly obese subjects and 303 controls of European descent. These genetic polymorphisms have been associated with the risk of DVT and PE; however, they have not been previously investigated in a group of patients with extreme overweight who are more prone to thrombotic complications. The results of our study show that gene polymorphisms of ABO (rs505922), F5 (rs6427196), MTHFR (rs1801133), and FGG (rs6536024) are not associated with the presence of morbid obesity among European subjects. We also did not determine differences in height or weight of individuals carrying different rs505922, rs6427196, rs1801133, or rs6536024 genotypes.
Multiple epidemiological and clinical studies showed a clear link between obesity and higher risk of thrombotic complications [2, 4]. The exact mechanisms of DVT development are still poorly understood [13]. Since genetic polymorphisms have been shown to have profound effect on the risk of thrombotic events [25], we expected that ABO, F5, MTHFR, and FGG genetic variations might be more prevalent in obese individuals who have higher risk of DVT than nonobese individuals. To date, 17 genes have been robustly demonstrated to harbor genetic variations associated with VT risk: ABO, F2, F5, F9, F11, FGG, GP6, KNG1, PROC, PROCR, PROS1, SERPINC1, SLC44A2, STXBP5, THBD, TSPAN15, and VWF [25]. Furthermore, genetic variations have been showing significant plasma levels of coagulation factors VII (FVII), VIII (FVIII), and von Willebrand factor (vWF) and thus mediate the risk of hemorrhage and thrombosis [26].
Previous studies have uncovered that ABO genetic variations are important risk factors for VTE [19, 27, 28]. A GWAS study identified ABO rs505922 as a risk factor for venous thrombosis [19]. Additionally, International Stroke Genetic Consortium revealed that SNP rs505922 was nominally associated with ischemic stroke (OR = 0.94, P = 0.023) [29]. Our genotyping analysis showed that T and C alleles of rs505922 and corresponding genotypes are distributed equally between the obese and control groups. The results of allele and genotype frequencies in our study are also comparable to the results obtained in previous studies with T allele ranging from 59 to 62% [30]. Interestingly, a recent meta-analysis provided some evidence that ABO rs505922 SNP C allele is a risk factor for cancer susceptibility, specifically for pancreatic cancer [30].
F5 is a protein of the coagulation system, which is not enzymatically active but functions as a cofactor [13]. Deficiency of F5 increases the risk of bleeding, while some mutations predispose to thrombosis [31]. Genetic variations in F5 region have been clearly linked with the risk of thrombosis with Leiden mutation rs6025 being the most well-known [25]. In order to identify additional novel genetic determinants of VTE, Tang et al. conducted a 2-stage GWAS among individuals of European ancestry [18]. This study showed two signals at the F5 region with an intronic variant rs2420370 and a coding variant rs6427196 in the 3′ untranslated region of F5. Within our study genotypes and alleles of rs6427196 were distributed equally between cases and controls.
MTHFR is the rate-limiting enzyme in the methyl cycle which is encoded by the MTHFR gene [32]. MTHFR gene polymorphism has been extensively studied in relation to different medical conditions. To date, there are reports that have linked rs1801133 polymorphism within MTHFR gene with arterial hypertension [33], cancer [34], diabetes [35], and many other diseases; however, the results of replication studies vary in between [36]. The homozygous mutated subjects for rs1801133 have higher homocysteine levels and hyperhomocysteinemia is an emerging risk factor for various thrombotic diseases [37]. A large number of studies has clearly shown that MTHFR gene polymorphism rs1801133 is a risk factor for thrombotic events [25]. In our study we observed a tendency for MTHFR rs1801133 TT genotype to be linked with morbid obesity when compared to CC genotype; however, the difference did not reach required significance. Similar results were obtained in a recessive model for MTHFR SNP (TT versus CT + CC), but association remained beyond statistical significance.
Fibrinogen gamma (FGG) gene belongs to the fibrinolysis cascade [25]. The genotypes of FGG gene have provided robust positive findings with the risk of VTE [25]. The T allele of the rs2066865 polymorphism in the year 2005 was found to reduce gamma fibrinogen plasma levels and to increase thrombosis risk by a factor of ~1.50 [38]. A more recent large GWAS study confirmed the importance of another FGG locus rs6536024 for VTE development [18]. Within our study we observed equal distribution of rs6536024 alleles in control and morbidly obese patient groups. The frequencies of C and T alleles for rs6536024 in our study correspond to previously published data on Caucasian subjects [18].
Several studies have suggested that height or length of legs of individuals is linked with increased risk of thrombotic events [8–10]. Therefore, we performed additional analysis in order to compare means of height and weight according to individual genotype of each of the four SNPs analyzed. In our study the height and weight means between separate ABO (rs505922), F5 (rs6427196), MTHFR (rs1801133), and FGG (rs6536024) gene polymorphisms did not differ. Interestingly, combination of obesity and tall stature was associated with VTE in Norwegian study [39]. These findings might be due to greater venous surface area, a larger number of venous valves, or greater hydrostatic pressure with longer legs [8]; however, the role of genetic factors within this context remains to be established.
To our best knowledge, this is the first study which investigated the frequencies of rs505922, rs6427196, rs1801133, and rs6536024 in morbidly obese subjects. Nevertheless, we admit that there are several important limitations related to the design of our study. There were gender and age distribution differences between obese and control groups; however, when performing statistical analysis we included gender and age as covariates, thus reducing the potential influence of these factors for the results. We also included only four gene polymorphisms in our study, while other genetic variations might be more important in the pathogenesis of thrombosis in obese individuals. Further large scale studies including obese patients who have the history of thrombotic complications would be extremely valuable. We could only speculate that certain genetic polymorphisms might be included in multivariate risk models for predicting thrombosis in morbidly patients in the future.
5. Conclusions
Our study showed that gene polymorphisms ABO C>T (rs505922), F5 C>G (rs6427196), MTHFR C>T (rs1801133), and FGG C>T (rs6536024) were not associated with morbid obesity, height, or weight in European subjects.
Acknowledgments
The authors would like to thank Raminta Grigaliunaite for help with genotyping experiments in the lab.
Competing Interests
The authors declare that they have no conflict of interests.
References
- 1.Després J.-P., Lemieux I. Abdominal obesity and metabolic syndrome. Nature. 2006;444(7121):881–887. doi: 10.1038/nature05488. [DOI] [PubMed] [Google Scholar]
- 2.Bell E. J., Lutsey P. L., Basu S., et al. Lifetime risk of venous thromboembolism in two cohort studies. The American Journal of Medicine. 2016;129(3):339.e19–339.e26. doi: 10.1016/j.amjmed.2015.10.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Saab J., Salvatore S. P. Evaluating the cause of death in obese individuals: a ten-year medical autopsy study. Journal of Obesity. 2015;2015:7. doi: 10.1155/2015/695374.695374 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Olié V., Fuhrman C., Chin F., Lamarche-Vadel A., Scarabin P.-Y., de Peretti C. Time trends in pulmonary embolism mortality in France, 2000–2010. Thrombosis Research. 2015;135(2):334–338. doi: 10.1016/j.thromres.2014.12.002. [DOI] [PubMed] [Google Scholar]
- 5.Rocha A. T., de Vasconcellos Â. G., da Luz Neto E. R., Araújo D. M. A., Alves E. S., Lopes A. A. Risk of venous thromboembolism and efficacy of thromboprophylaxis in hospitalized obese medical patients and in obese patients undergoing bariatric surgery. Obesity Surgery. 2006;16(12):1645–1655. doi: 10.1381/096089206779319383. [DOI] [PubMed] [Google Scholar]
- 6.Childs B. R., Nahm N. J., Dolenc A. J., Vallier H. A. Obesity is associated with more complications and longer hospital stays after orthopaedic trauma. Journal of Orthopaedic Trauma. 2015;29(11):504–509. doi: 10.1097/bot.0000000000000324. [DOI] [PubMed] [Google Scholar]
- 7.Marquez-Lara A., Nandyala S. V., Sankaranarayanan S., Noureldin M., Singh K. Body mass index as a predictor of complications and mortality after lumbar spine surgery. Spine. 2014;39(10):798–804. doi: 10.1097/BRS.0000000000000232. [DOI] [PubMed] [Google Scholar]
- 8.Lutsey P. L., Cushman M., Heckbert S. R., Tang W., Folsom A. R. Longer legs are associated with greater risk of incident venous thromboembolism independent of total body height. The Longitudinal Study of Thromboembolism Etiology (LITE) Thrombosis and Haemostasis. 2011;106(1):113–120. doi: 10.1160/th11-02-0100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Flinterman L. E., van Hylckama Vlieg A., Rosendaal F. R., Cannegieter S. C. Body height, mobility, and risk of first and recurrent venous thrombosis. Journal of Thrombosis and Haemostasis. 2015;13(4):548–554. doi: 10.1111/jth.12860. [DOI] [PubMed] [Google Scholar]
- 10.Emerging Risk Factors Collaboration. Adult height and the risk of cause-specific death and vascular morbidity in 1 million people: individual participant meta-analysis. International Journal of Epidemiology. 2012;41(5):1419–1433. doi: 10.1093/ije/dys086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Bell B. R., Bastien P. E., Douketis J. D. Prevention of venous thromboembolism in the Enhanced Recovery After Surgery (ERAS) setting: an evidence-based review. Canadian Journal of Anesthesia. 2015;62(2):194–202. doi: 10.1007/s12630-014-0262-2. [DOI] [PubMed] [Google Scholar]
- 12.Souto J. C. A., Pena G., Ziyatdinov A., et al. A genomewide study of body mass index and its genetic correlation with thromboembolic risk. Results from the GAIT project. Thrombosis and haemostasis. 2014;112(5):1036–1043. doi: 10.1160/th14-03-0275. [DOI] [PubMed] [Google Scholar]
- 13.Samad F., Ruf W. Inflammation, obesity, and thrombosis. Blood. 2013;122(20):3415–3422. doi: 10.1182/blood-2013-05-427708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Standeven K. F., Uitte de Willige S., Carter A. M., Grant P. J. Heritability of clot formation. Seminars in Thrombosis and Hemostasis. 2009;35(5):458–467. doi: 10.1055/s-0029-1234141. [DOI] [PubMed] [Google Scholar]
- 15.Souto J. C., Almasy L., Borrell M., et al. Genetic susceptibility to thrombosis and its relationship to physiological risk factors: the GAIT study. Genetic Analysis of Idiopathic Thrombophilia. The American Journal of Human Genetics. 2000;67(6):1452–1459. doi: 10.1086/316903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Trégouët D.-A., Heath S., Saut N., et al. Common susceptibility alleles are unlikely to contribute as strongly as the FV and ABO loci to VTE risk: results from a GWAS approach. Blood. 2009;113(21):5298–5303. doi: 10.1182/blood-2008-11-190389. [DOI] [PubMed] [Google Scholar]
- 17.Paré G., Chasman D. I., Parker A. N., et al. Novel associations of CPS1, MUT, NOX4, and DPEP1 with plasma homocysteine in a healthy population: a genome-wide evaluation of 13 974 participants in the Women’s Genome Health Study. Circulation: Cardiovascular Genetics. 2009;2(2):142–150. doi: 10.1161/circgenetics.108.829804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Tang W., Teichert M., Chasman D. I., et al. A genome-wide association study for venous thromboembolism: the extended cohorts for heart and aging research in genomic epidemiology (CHARGE) consortium. Genetic Epidemiology. 2013;37(5):512–521. doi: 10.1002/gepi.21731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Germain M., Saut N., Greliche N., et al. Genetics of venous thrombosis: insights from a new genome wide association study. PLoS ONE. 2011;6(9) doi: 10.1371/journal.pone.0025581.e25581 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Folsom A. R., Tang W., Weng L.-C., et al. Replication of a genetic risk score for venous thromboembolism in whites but not in African Americans. Journal of Thrombosis and Haemostasis. 2016;14(1):83–88. doi: 10.1111/jth.13193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Klovaite J., Benn M., Nordestgaard B. G. Obesity as a causal risk factor for deep venous thrombosis: a Mendelian randomization study. Journal of Internal Medicine. 2015;277(5):573–584. doi: 10.1111/joim.12299. [DOI] [PubMed] [Google Scholar]
- 22.Kupcinskas J., Wex T., Link A., et al. PSCA and MUC1 gene polymorphisms are associated with gastric cancer and pre-malignant gastric conditions [corrected] Anticancer Research. 2014;34(12):7167–7175. [PubMed] [Google Scholar]
- 23.Kupcinskas J., Bruzaite I., Juzenas S., et al. Lack of association between miR-27a, miR-146a, miR-196a-2, miR-492 and miR-608 gene polymorphisms and colorectal cancer. Scientific Reports. 2014;4, article 5993 doi: 10.1038/srep05993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Purcell S., Neale B., Todd-Brown K., et al. PLINK: a tool set for whole-genome association and population-based linkage analyses. The American Journal of Human Genetics. 2007;81(3):559–575. doi: 10.1086/519795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Morange P.-E., Suchon P., Trégouët D.-A. Genetics of venous thrombosis: update in 2015. Thrombosis and Haemostasis. 2015;114(5):910–919. doi: 10.1160/th15-05-0410. [DOI] [PubMed] [Google Scholar]
- 26.Smith N. L., Chen M.-H., Dehghan A., et al. Novel associations of multiple genetic loci with plasma levels of factor VII, factor VIII, and von Willebrand factor, The CHARGE (Cohorts for Heart and Aging Research in Genome Epidemiology) Consortium. Circulation. 2010;121(12):1382–1392. doi: 10.1161/circulationaha.109.869156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Germain M., Chasman D. I., de Haan H., et al. Meta-analysis of 65,734 individuals identifies TSPAN15 and SLC44A2 as two susceptibility loci for venous thromboembolism. The American Journal of Human Genetics. 2015;96(4):532–542. doi: 10.1016/j.ajhg.2015.01.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Bruzelius M., Bottai M., Sabater-Lleal M., et al. Predicting venous thrombosis in women using a combination of genetic markers and clinical risk factors. Journal of Thrombosis and Haemostasis. 2015;13(2):219–227. doi: 10.1111/jth.12808. [DOI] [PubMed] [Google Scholar]
- 29.Williams F. M. K., Carter A. M., Hysi P. G., et al. Ischemic stroke is associated with the ABO locus: the EuroCLOT study. Annals of Neurology. 2013;73(1):16–31. doi: 10.1002/ana.23838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Duan Y.-F., Zhu F., Li X.-D., et al. Association between ABO gene polymorphism (rs505922) and cancer risk: a meta-analysis. Tumor Biology. 2015;36(7):5081–5087. doi: 10.1007/s13277-015-3159-y. [DOI] [PubMed] [Google Scholar]
- 31.Segers K., Dahlbäck B., Nicolaes G. A. F. Coagulation factor V and thrombophilia: background and mechanisms. Thrombosis and Haemostasis. 2007;98(3):530–542. doi: 10.1160/th07-02-0150. [DOI] [PubMed] [Google Scholar]
- 32.Goyette P., Sumner J. S., Milos R., et al. Human methylenetetrahydrofolate reductase: isolation of cDNA, mapping and mutation identification. Nature Genetics. 1994;7(2):195–200. doi: 10.1038/ng0694-195. [DOI] [PubMed] [Google Scholar]
- 33.Wu Y.-L., Hu C.-Y., Lu S.-S., et al. Association between methylenetetrahydrofolate reductase (MTHFR) C677T/A1298C polymorphisms and essential hypertension: a systematic review and meta-analysis. Metabolism: Clinical and Experimental. 2014;63(12):1503–1511. doi: 10.1016/j.metabol.2014.10.001. [DOI] [PubMed] [Google Scholar]
- 34.Rai V. Folate pathway gene MTHFR C677T polymorphism and risk of lung cancer in Asian populations. Asian Pacific Journal of Cancer Prevention. 2014;15(21):9259–9264. doi: 10.7314/APJCP.2014.15.21.9259. [DOI] [PubMed] [Google Scholar]
- 35.Abbas S., Raza S. T., Ahmed F., Ahmad A., Rizvi S., Mahdi F. Association of Genetic polymorphism of PPARγ-2, ACE, MTHFR, FABP-2 and FTO genes in risk prediction of type 2 diabetes mellitus. Journal of Biomedical Science. 2013;20, article 80 doi: 10.1186/1423-0127-20-80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Liew S.-C., Das Gupta E. Methylenetetrahydrofolate reductase (MTHFR) C677T polymorphism: epidemiology, metabolism and the associated diseases. European Journal of Medical Genetics. 2015;58(1):1–10. doi: 10.1016/j.ejmg.2014.10.004. [DOI] [PubMed] [Google Scholar]
- 37.Park W.-C., Chang J.-H. Clinical implications of methylenetetrahydrofolate reductase mutations and plasma homocysteine levels in patients with thromboembolic occlusion. Vascular Specialist International. 2014;30(4):113–119. doi: 10.5758/vsi.2014.30.4.113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.de Willige S. U., de Visser M. C. H., Houwing-Duistermaat J. J., Rosendaal F. R., Vos H. L., Bertina R. M. Genetic variation in the fibrinogen gamma gene increases the risk for deep venous thrombosis by reducing plasma fibrinogen γ′ levels. Blood. 2005;106(13):4176–4183. doi: 10.1182/blood-2005-05-2180. [DOI] [PubMed] [Google Scholar]
- 39.Borch K. H., Nyegaard C., Hansen J.-B., et al. Joint effects of obesity and body height on the risk of venous thromboembolism: the Tromsø Study. Arteriosclerosis, Thrombosis, and Vascular Biology. 2011;31(6):1439–1444. doi: 10.1161/atvbaha.110.218925. [DOI] [PubMed] [Google Scholar]