

Abstract
Recent discoveries have revealed that microRNAs (miRNAs) play a key role in the regulation of gene expression. In this review, we summarize the rapidly evolving knowledge about liver miRNAs (including miR-33, -33*, miR-223, -30c, -144, -148a, -24, -29, and -122) and their link to hepatic lipid metabolism, atherosclerosis and cardiovascular disease, non-alcoholic fatty liver disease, metabolic syndrome, and type-2 diabetes. With regards to its biomarker potential, the main focus is on miR-122 as the most abundant liver miRNA with exquisite tissue specificity. MiR-122 has been proposed to play a central role in the maintenance of lipid and glucose homeostasis and is consistently detectable in serum and plasma. This miRNA may therefore constitute a novel biomarker for cardiovascular and metabolic diseases.
Keywords: MicroRNAs, Biomarkers, Lipid metabolism, Cardiovascular disease
Introduction
MicroRNAs (miRNAs, miRs) are small ∼22 nucleotides long non-coding regulatory molecules.1 They are generated from primary transcripts (pri-miR), which are processed by endonuclease complexes into miRNA precursors (pre-miR) and further into a duplex of two miRNA strands (5p and 3p strand).2 In most cases, only one of the two strands (termed the guide or mature strand) is stable and biologically active. In some cases, the other strand (termed passenger or star [*] strand) also regulates distinct targets. MiRNAs are able to modify gene expression at the post-transcriptional level by binding to the 3′-untranslated regions of target messenger RNAs and thereby inducing their degradation or repressing their translation.1 They often have multiple targets within the same biological pathway, closely interact with each other, and target both activators and inhibitors of a functional regulator.3 MiRNAs therefore constitute a layer of epigenetic regulation that provides additional control of intricate processes such as metabolism, cell growth, differentiation, stress response, and tissue remodelling, and safeguards the stability of biological systems. In 2015, the European Society of Cardiology Working Group Atherosclerosis and Vascular Biology has identified miRNAs as an area of major interest in the search for novel biomarkers.4
Over the past years, evidence has emerged that miRNAs involved in the regulation of cholesterol and lipid metabolism in the liver may contribute to the development of metabolic disturbances and cardiovascular disease. The present review (i) describes the mechanistic role of liver miRNAs in hepatic lipid metabolism; (ii) assesses the detectability of liver miRNAs in the circulation, a prerequisite for their exploitation as soluble biomarkers; (iii) summarizes the evidence from epidemiological studies on associations of liver miRNAs in circulation (i.e. those measured in acellular samples of serum or plasma) with metabolic and cardiovascular outcomes; and (iv) discusses further steps required for their use in clinical practise. The role of miRNAs in the regulation of lipid metabolism in non-hepatic tissues such as adipose tissue or in macrophages5 and their respective implications for obesity and atherosclerosis6 have been reviewed elsewhere.
Liver miRNAs as regulators of hepatic lipid metabolism
Most miRNAs involved in lipid homeostasis have been reported to modulate lipid transport processes and assembly rather than degradation and clearance. A key protein mediating the efflux of cholesterol to high-density lipoprotein (HDL)-forming apolipoprotein A I is the ATP-binding cassette transporter A1 (ABCA1). ABCA1 appears to be regulated by several miRNAs. This is due to the fact that ABCA1 has an exceptionally long 3′ UTR (3.3 kb, the average length of human 3′ UTRs is 0.8 kb), allowing the binding of multiple different miRNAs. MiRNAs directly targeting ABCA1 include miR-10b, miR-17, miR-19b, miR-26, miR-27a/b, miR-33a/b, miR-33a*/b*, miR-93, miR-101, miR-106b, miR-128, miR-144, miR-145, miR-148a, miR-302a, and miR-758,7–16 whereas miR-223 has been shown to regulate ABCA1 indirectly.17 This regulation of ABCA1 is a prime example of a miRNA regulatory network converging on one target, yet the physiological and pathological relevance of some of these miRNAs as regulators of cholesterol transport remains to be validated. In the following sections, we summarize the role of several miRNAs in hepatic lipid metabolism.
MiR-33a/b
In humans, miR-33a and miR-33b are encoded in introns of the genes coding sterol regulatory element-binding proteins (SREBP-2 and SREBP-1), while mice express only one miR-33 isoform, located in an intron of Srebp-2. MiR-33 is a key regulator of lipid metabolism and transport.18,19 MiR-33 targets ABCA1 and thus suppresses HDL synthesis by attenuation of cholesterol efflux to apolipoprotein A1 and nascent HDL.20 Loss of miR-33 in low-density lipoprotein receptor null (LDLR−/−) mice raises plasma HDL-cholesterol levels and promotes reverse cholesterol transport. As expected by these findings, anti-miR-33 therapy attenuates the progression and enhances the regression of atherosclerosis in LDLR−/− mice.21,22 Similarly, genetic ablation of miR-33 in ApoE−/− mice markedly reduces the progression of atherosclerosis.23 In contrast, a study has recently shown that anti-miR-33 therapy using different antisense oligonucleotides (ASOs) fails to prevent atherogenesis,24 suggesting that the chemical modification of the ASOs might influence their therapeutic efficacy. In addition to its prominent role in regulating lipid metabolism, a recent report revealed that miR-33 antagonism increased mitochondrial function and de-repressed ABCA1 in macrophages, which, in combination, resulted in an increased cholesterol efflux from macrophages.25 In the same study, anti-miR-33 treatment in atherosclerotic ApoE−/− mice reduced lesion size even though circulating lipid levels were unaffected. Moreover, elevated miR-33 and reduced expression of the mitochondria regulatory genes PGC-1α, SLC25A25, NRF1, and TFAM were observed in human atherosclerotic plaques.25 Another biological effect of miR-33 is the attenuation of hepatic secretion of very-low-density lipoprotein (VLDL)-triglycerides by targeting N-ethylmaleimide-sensitive factor, an essential component of the exocytic pathway.26
In addition to the guide strand miR-33 (miR-33-5p), the passenger strand miR-33* (miR-33-3p) has been implicated in the regulation of cholesterol and lipid metabolism.9 MiR-33* represses several genes encoding key enzymes and transcription factors involved in cholesterol efflux (including ABCA1), fatty acid (FA) metabolism, and insulin signalling.9 Overexpression of miR-33* in hepatic cells reduces FA oxidation. Thus, two primary transcripts (SREBP-1 and SREBP-2) give rise to six important regulators of lipid and cholesterol metabolism: SREBP-1, SREBP-2, miR-33a, miR-33a*, miR-33b, and miR-33b*. MiR-33, in turn, represses SREBP-1 thus fine-tuning cholesterol homeostasis by an auto-feedback loop.27 Because of its essential role in lipid metabolism, miR-33 has been considered and tested as a therapeutic target for metabolic disorders. Inhibition of miR-33 in non-human primates resulted in elevated plasma HDL-C and decreased VLDL-C levels.28 However, there are conflicting reports on the long-term inhibition of miR-33 in mice, which may or may not have detrimental effects such as elevated plasma triglyceride (TG) levels and moderate hepatic steatosis.29,30 Notably, the adverse effects were observed only upon feeding mice a high-fat diet.29 Nonetheless, the observation has clinical relevance given the potential target population for miR-33 therapy. Therefore, further studies are required to determine long-term effects and safety of miR-33 inhibition.
MiR-223
MiR-223 was initially reported to be myeloid cell-specific, but it has recently been shown to be also expressed in hepatocytes where it regulates cholesterol homeostasis.17 Hepatic miR-223 reduces cholesterol biosynthesis by repressing 3-hydroxy-3-methylglutaryl-CoA synthase 1 and sterol-C4-methyloxidase-like protein, and inhibits HDL-C uptake by targeting the scavenger receptor class B member 1. Furthermore, miR-223 promotes cholesterol efflux by positively regulating ABCA1 expression via its direct target Sp3. Importantly, miR-223 levels are regulated by the cholesterol level; in a low-cholesterol state, miR-223 is suppressed. As a consequence, cholesterol synthesis and uptake are increased, whereas cholesterol efflux to HDL is attenuated in order to raise cellular cholesterol levels.17
MiR-30c
MiR-30c exerts lipid-lowering effects in the liver: It represses the microsomal triglyceride transfer protein (MTP), which is essential for lipoprotein assembly. Hepatic miR-30c reduces lipid synthesis and lipoprotein secretion and decreases atherosclerosis in ApoE−/− mice.31 In addition to its role in regulating lipoprotein metabolism and the progression of atherosclerosis, miR-30c overexpression in cardiomyocytes results in dilated cardiomyopathy.32
MiR-144
MiR-144 is an another regulator of cholesterol metabolism and transport in the liver33 and in macrophages.34 In the liver, the nuclear bile acid receptor farnesoid X receptor up-regulates miR-144, which, in turn, targets ABCA1. This results in lower plasma HDL-cholesterol levels.33 Similarly, in macrophages, activation of a related receptor, liver X receptor, leads to an up-regulation of miR-144 and a down-regulation of ABCA1. As a consequence, cholesterol efflux from macrophages is attenuated.34 Interestingly, overexpression of miR-144 increases atherogenesis in ApoE−/− mice.35
MiR-148a
Two independent studies have identified miR-148a-3p as a major regulator of lipoprotein metabolism using different experimental approaches.8,10 Hepatic miR-148a-3p negatively regulates LDL receptor (LDLR) and ABCA1 levels.8,10 In vivo inhibition of miR-148a-3p increases hepatic LDLR and ABCA1 expression lowering LDL-cholesterol and increasing HDL-cholesterol in plasma.8,10 MiR-148a-3p is also highly expressed in macrophages where it controls ABCA1 expression and cholesterol efflux.10 Importantly, the locus encoding miR-148a-3p is located in a genomic region enriched for single nucleotide polymorphisms associated with abnormal circulating total cholesterol, TG, or LDL-cholesterol levels.10,36,37 Thus, genetic determinants of lipoprotein metabolism in humans are also likely to affect the expression of miR-148a-3p in the liver and/or other tissues.
MiR-122
MiR-122 is the predominant miRNA in the liver and completely conserved in vertebrates. It accounts for >70% of the total hepatic miRNA expression38 and regulates a number of genes associated to cholesterol and FA metabolism (Figure 1).39 In mice, inhibition of miR-122 using ASO lowered circulating cholesterol by ∼25–35%, reduced hepatic lipid synthesis, and enhanced hepatic FA oxidation.40,41 Similarly, antagonism of miR-122 in non-human primates markedly lowers plasma cholesterol levels.42,43 Antisense targeting of miR-122 results in the down-regulation of a broad array of genes involved in lipid synthesis and lipoprotein assembly including 3-hydroxy-3-methylglutaryl-coenzyme A reductase, the rate-limiting enzyme in the cholesterol biosynthesis pathway, 3-hydroxy-3-methylglutaryl-CoA synthase 1, and MTP.44 All these proteins are no direct targets of miR-122, and the mechanisms by which miR-122 regulates their expression are still unknown.
Figure 1.

Role of miR-122 in lipid homeostasis. ASO, antisense oligonucleotide; FA, fatty acid; HMGCR, 3-hydroxy-3-methylglutaryl-CoA reductase; KO, knock-out; LDL, low-density lipoprotein; LPL, lipoprotein lipase; LDLR, low-density lipoprotein receptor; MTP, microsomal triglyceride transfer protein; TG, triglycerides; VLDL, very-low-density lipoprotein.
MiR-122 has also been implicated in systemic iron homeostasis by directly targeting mRNAs that encode activators of hepcidin expression (Hfe and Hjv),45 and in glucose homeostasis by indirect effects on AMP-activated kinase and glucose 6-phosphatase, a key regulatory enzyme of hepatic gluconeogenesis.40,44 Preliminary data from miR-122 null mice kept on high-fat diet for 3 months suggest improved insulin sensitivity compared with wild-type mice along with lower fasting glucose levels, enhanced thermogenesis, and lower body weight.46 However, genetic ablation of miR-122 in mice also causes a marked reduction of hepatic MTP expression and VLDL secretion leading to hepatosteatosis.47 Similar observations were found in a separate study, in which miR-122 deletion resulted in hepatic lipid accumulation, hepatitis, and the development of hepatocellular carcinoma-like tumours.48 There is accumulating evidence that miR-122 exerts tumour-suppressive effects49 but also inhibits collagen maturation in hepatic stellate cells.50
Detection of liver miRNAs in circulation
In circulation, miRNAs are packaged in membranous microvesicles and protein complexes,51 protecting them from enzymatic degradation.52 While their levels in serum and plasma are remarkably stable,38 miRNAs may be released into circulation by different tissues and upon different triggers (see Figure 2). For instance, we have previously shown that activation of platelets triggers the release of miRNAs, including miR-21, miR-24, miR-126, miR-191, and miR-223, into circulation, whereas therapeutic platelet inhibition reduces their levels. Measurement of circulating platelet miRNA levels could therefore be useful as an in vivo test for platelet activation.53–56 Another important trigger is tissue damage. MiRNAs released from the heart upon myocardial injury have been suggested as markers for diagnosis and prognosis of acute coronary syndrome but without clear evidence for superiority over troponins.57 Liver miRNAs, especially circulating miR-122, are sensitive markers of acute liver failure and acetaminophen toxicity58 and explored for in vitro assessment of drug-induced hepatocyte toxicity.59 Remarkably, even under normal conditions, miR-122 is released from the liver, mainly via hepatic exosomes,60,61 a mechanism that is modulated by statins.62 MiR-122 is, to our knowledge, the only tissue-specific miRNA that is released into the circulation in a constant manner. Due to its abundance in the liver, miR-122 is readily detectable in the circulation. Also, it deserves comment that circulating miR-122 level and hepatic miR-122 expression were highly correlated (r = 0.46) in a recent study of 67 patients with non-alcoholic fatty liver disease (NAFLD).63
Figure 2.
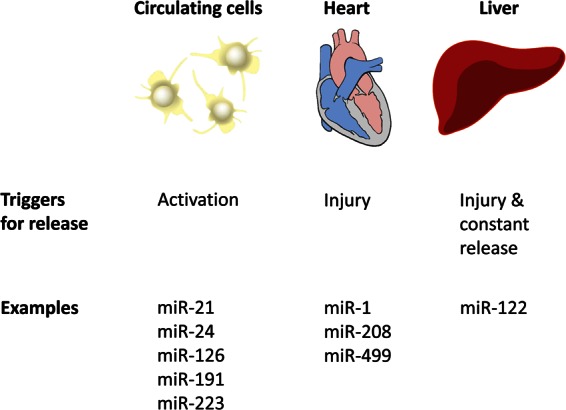
Sources of circulating miRNAs. Circulating cells, in particular platelets, release miRNAs upon activation (i.e. miR-21, miR-24, miR-126, miR-191, and miR-223). Tissues release miRNAs upon injury; i.e. miR-1, miR-208, and miR-499 are elevated after myocardial infarction. The liver, however, constantly secretes miR-122. Unlike miR-122, other liver miRNAs are not tissue specific and not as abundant. Thus, most of them cannot be reliably detected in the circulation (Table 1).
In comparison, we have assessed the ability to detect other liver miRNAs related to lipid homeostasis in plasma samples taken from healthy individuals (Table 1). Using quantitative real-time polymerase chain reactions (qPCR) and next generation sequencing, we found that apart from miR-122 only 8 out of 16 miRNAs could be reliably detected. However, many of these miRNAs are expressed in various tissues (Table 1) and thus lack specificity for the liver. Importantly, the high platelet contribution to circulating levels of miR-223, miR-320, miR-27b, miR-335, miR-148a, and miR-30c53–56 hampers their use as putative biomarkers for metabolic processes in the liver. As anticipated, the only miRNA with known impact on lipid metabolism that is liver-specific and reliably detectable in circulation was miR-122, making it a promising candidate biomarker.
Table 1.
Detectability of liver miRNAs implicated in lipid homeostasis in plasma samples taken from healthy individuals
| MiRNA | Average Ct | Average rpm | First author, reference |
|---|---|---|---|
| Detected reliably | |||
| 223 | 18.4 | 275.7 | Vickers17 |
| 320 | 23.8 | 214.5 | Chen91 |
| 122 | 25.3 | 60.8 | Esau40 |
| 27b | 25.4 | 938.0 | Vickers,92 Goedeke93 |
| 30c | 26.8 | 447.5 | Soh31 |
| 27a | 27.9 | 103.5 | Alvarez94 |
| 378 | 28.6 | 42.6 | Carrer95 |
| 335 | 28.9 | 36.5 | Nakanishi96 |
| 148a | 29.8 | 616.9 | Wagschal10 |
| Not detected reliably | |||
| 370 | 33 | 6.9 | Iliopoulos97 |
| 613 | 36.3 | 0.0 | Ou98 |
| 33a* | 37.5 | 0.1 | Goedeke9 |
| 378* | 38.6 | 0.1 | Carrer95 |
| 33a | 39.7 | 1.1 | Rayner28 |
| 33b | 40 | 1.2 | Rayner28 |
| 33b* | n.d. | 0.0 | Goedeke9 |
| 144 | n.d. | 28.6 | de Aguiar Vallim33 |
Shown is the average Ct value for individual miRNAs assessed by qPCR using Taqman assays in 12 random plasma samples from the Bruneck cohort and the average rpm (reads per million) from small RNA sequencing in 4 plasma samples. MiRNAs are categorised by the ability to reliably detect them in plasma samples using both methods (qPCR and sequencing). MiR-122 (in bold) is the only liver-specific miRNA reliably detectable in circulation.
n.d., not determined.
Liver miRNAs as biomarkers in epidemiological studies
We performed a literature search for case-control and prospective studies that reported on the potential association of miR-122 levels with metabolic and cardiovascular outcomes. The identified studies are summarized in Table 2.
Table 2.
Overview of published epidemiological studies on miR-122 and metabolic and cardiovascular outcomes
| First author | Study design | Sample type | Cases | Controls | No. of participants or cases/controls | Reported fold differences (FD) or odds ratios (OR) for circulating miR-122 with outcome |
|---|---|---|---|---|---|---|
| Metabolic outcomes | ||||||
| Becker67 | Case-control | Serum | NASH | Healthy controls | 87/61 | OR per 1 dCt decrease 1.69 (P < 0.001)a |
| NAFLD | Healthy controls | 50/61 | Higher miR-122 (P < 0.001) | |||
| Celikbilek68 | Case-control | Serum | NASH | Healthy controls | 20/20 | n.s.b |
| Cermelli64 | Case-control | Serum | NAFLD-SS | Healthy controls | 34/19 | FD 5.7 (P < 0.001) |
| NASH | Healthy controls | FD 11.4 (P < 0.001) | ||||
| Pirola66 | Case-control | Serum | NAFLD-SS | Healthy controls | 16/16 | FD 2.3 (P < 0.05)b |
| NASH | Healthy controls | 16/16 | FD 7.2 (P < 0.05)b | |||
| NAFLD-SS | Healthy controls | 30/19 | FD 1.7 (n.s.)b | |||
| NASH | Healthy controls | 47/19 | FD 3.3 (P = 0.006)b | |||
| Tan69 | Case-control | Serum | NAFLD | Healthy controls | 20/20 | FD 9.3 (P < 0.01)c |
| NAFLD | Healthy controls | 152/90 | FD 8.7 (P < 0.001)c | |||
| Yamada65 | Case-control | Serum | NAFLD | Healthy controls | 48/90 | Men: FD 3.1 (P < 0.001) |
| NAFLD | Healthy controls | 44/221 | Women: FD 2.6 (P < 0.001) | |||
| Gao76 | Case-control | Plasma | Hyperlipidaemia | Controls | 155/100 | FD 2.0 (P = 0.008) |
| Wang77 | Case-control | Serum | Insulin resistance | Controls | 123/107 | OR 3.4 (P = 0.028)d |
| Obese | Normal weight | 123/107 | FD 3.1 (P < 0.001) | |||
| Obese | Normal weight | 56/56 | FD 3.2 (P < 0.001) | |||
| Ortega78 | Intervention | Plasma | Surgery-induced weight loss | 22 | FD 0.05 (P < 0.001) | |
| Diet-induced weight loss | 9 | FD 0.6 (n.s.) | ||||
| Janssen79 | Phase-2 RCT | Not applicable | Miravirsen vs. placebo in HCV patients | 27/9 | Total cholesterol ↓ | |
| Willeit80 | Prospective | Serum and plasma | Incident metabolic syndrome in general population | 810 | OR per SD 1.60 (P < 0.001)e | |
| Incident type-2 diabetes in general population | 810 | OR per SD 1.37 (P = 0.021)e | ||||
| Cardiovascular outcomes | ||||||
| Li81 | Case-control | Plasma | AMI | Healthy controls | 115/16 | FD 4.0 (P = 0.036) |
| AMI | Non-CHD | 30/21 | FD 17.8 (P = 0.023) | |||
| AMI | Non-CHD | 77/51 | n.s. | |||
| D'Alessandra84 | Case-control | Serum | AMI | Healthy controls | 33/17 | FD 0.13 (P < 0.01) |
| Corsten83 | Case-control | Plasma | AMI | Normal angiogram | 32/36 | FD 0.83 (P = 0.35) |
| Gao76 | Case-control | Plasma | CAD | No CAD | 155/100 | OR 1.08 (P = 0.034)f |
| Corsten83 | Case-control | Plasma | Acute heart failure | Healthy controls | 33/34 | FD 2.5 (P < 0.05) |
| D'Alessandra82 | Case-control | Plasma | Unstable angina | Healthy controls | 19/20 | FD 4.0 (P = 0.011) |
| Jickling85 | Case-control | PBCs | Ischaemic stroke | Healthy controls | 24/24 | FD 0.44 (P = 0.047) |
| Willeit80 | Prospective | Serum and plasma | Incident CVD in general population | 810 | OR per SD 1.10 (P = 0.330)e | |
AMI, acute myocardial infarction; CAD, coronary artery disease; CHD, coronary heart disease; CVD, cardiovascular disease; FD, fold difference; NAFLD, non-alcoholic fatty liver disease; NASH, non-alcoholic steatohepatitis; n.s., not significant; OR, odds ratio; PBCs, peripheral blood cells; RCT, randomized controlled trial.
aAdjusted for age, sex, and body mass index.
bAge- and sex-matched.
cAge-, sex-, and ethnicity-matched.
dAdjusted for age, sex, body mass index, waist–hip ratio, fat percentage, systolic blood pressure, diastolic blood pressure, lipid profile and liver function.
eAdjusted for age, sex, socio-economic status, smoking, physical activity, and alcohol consumption.
fAdjusted for age, sex, BMI, smoking, hypertension, diabetes, and lipids.
Metabolic outcomes
Non-alcoholic fatty liver disease occurs when the accumulation of liver fat exceeds 5% and encompasses a broad spectrum of pathological conditions ranging from steatosis to non-alcoholic steatohepatitis (NASH), fibrosis and cirrhosis. NAFLD represents the most common liver disease in western countries with a common link to metabolic syndrome and type 2 diabetes. An invasive procedure (liver biopsy) is still required to confirm the diagnosis and assess the degree of fibrosis. Recently, liver miRNAs in the context of NAFLD have been implicated both as causal mediators and promising non-invasive biomarkers. Several studies strived for the identification of a serum miRNA signature of NAFLD,64–68 and one performed external validation in an independent cohort of patient (n = 152) and control (n = 90) samples with the final selection of miRNAs comprising miR-122, -1290, -27b, and -192 and the diagnostic yield surpassing that of standard transaminase levels (Table 2).69 Unsurprisingly, the common readout of the various studies was miR-122, the most abundant liver miRNA with exquisite tissue specificity, which was consistently elevated in all series of NAFLD patients and increased with disease severity.64,66,67 Large-scale epidemiological studies are now required to clarify whether high circulating miR-122 levels, alone or in concert with other miRNAs, offer sufficient diagnostic performance to limit future necessity of liver biopsies for NAFLD.
From a pathophysiological perspective, NAFLD, featured by excess accumulation of TGs, reflects an unbalance between de novo lipogenesis in the liver, FA and lipoprotein uptake, FA oxidation, and lipoprotein assembly and release (TG export). MiRNAs including miR-122 are crucially involved in all these processes as epigenetic regulators on a post-transcriptional level.70 MiR-122 is a key positive regulator of de novo lipogenesis, which accounts for roughly one-quarter of hepatic fat deposits in NAFLD patients and produces lipid species that confer a high cardiovascular disease risk.71 In line, silencing of miR-122 (by ASO) prevents hepatosteatosis in response to high-fat diet.40 Of note, some residual miR-122 activity is required for achieving favourable effects on liver fat content. MiR-122 null mice spontaneously develop severe hepatosteatosis,47 probably due to diminished expression of MTP and subsequent impairment of VLDL assembly and secretion.70 Remarkably, in NASH as well as in alcoholic hepatitis, packaging of miR-122 in exosomes is up-regulated leading to increased levels of circulating miR-122, increased miR-122 delivery to macrophages eliciting an inflammatory response,72 and decreased liver miR-122 levels potentially promoting fibrosis.73 Other miRNAs particularly relevant to NAFLD include (i) miR-33, down-regulation of which leads to severe liver steatosis29; (ii) miR-24, which targets insulin-induced gene 1 and inhibition of which blocks both hepatic steatosis and hyperlipidaemia74; and (iii) miR-29, which prevents lipoprotein lipase from being expressed in liver and inhibition of which promotes hepatic lipid accumulation in mice.75
The effects of miR-122 on de novo lipogenesis—and lipid metabolism in general—are also reflected in the cross-sectional correlations with major lipid species. Previous studies generally reported positive correlations of circulating miR-122 with total cholesterol, TG, LDL-cholesterol, and liver function enzymes, and inverse correlations with HDL-cholesterol. Participants with hyperlipidaemia and insulin resistance have markedly higher circulating miR-122 levels than healthy controls (Table 2).76,77
Circulating miR-122 levels are higher in obese people compared with people with normal weight (Table 2).77 In a study of morbidly obese patients, surgery-induced weight loss led to a major reduction of miR-122 levels.78 A Phase-2 clinical trial that aimed to assess the efficacy of the miR-122 inhibitor Miravirsen in reducing viral load in patients with hepatitis C reported a concurrent reduction in total cholesterol over 14 weeks of treatment, but no shift in the ratio of LDL-cholesterol to HDL-cholesterol.79 As the first prospective population-based study investigating the biomarker potential of miR-122 in the general population, the Bruneck study reported that miR-122 is significantly associated with incident metabolic syndrome and type 2 diabetes, even after adjustment for age, sex, socio-economic status, smoking, physical activity, and alcohol consumption.80
Cardiovascular outcomes
In contrast to metabolic outcomes, evidence available for cardiovascular outcomes is sparse (Table 2), with some studies reporting positive76,81,82 and some studies reporting inverse83–85 associations. The interpretation of these studies is further complicated by the reporting of crude fold differences (i.e. those unadjusted for potential confounding variables) and the notion that statins may influence the release of miR-122 from the liver into circulation.62 In the Bruneck study, circulating miR-122 was not significantly associated with the development of incident cardiovascular disease over a 15-year time frame.80 However, there is evidence that liver miRNAs may adversely influence plaque composition through their aforementioned effects on lipid metabolism or effects on other cell types.86 Antagonism of miR-33 in atherosclerosis-susceptible mice promoted accumulation of anti-inflammatory M2 macrophages and regulatory T-cells (FOXP3 + Tregs)87 and enhanced cholesterol efflux from lesional macrophages.21 Both effects are assumed to stabilize advanced plaques. MiR-24, in turn, targets pro-inflammatory chitinase-3-like protein 1, macrophage apoptosis, and matrix metalloproteinase-14 expression and activation in macrophages.88 Inhibition of miR-24 resulted in a vulnerable plaque phenotype with decreased collagen content and increased macrophage infiltration.89
Steps required for clinical translation
Epidemiological research into the role of miR-122 in metabolic and cardiovascular diseases is still in its infancy. Published studies on the cross-sectional correlates of miR-122 are limited to the major lipid classes (e.g. total cholesterol, HDL-cholesterol, and LDL-cholesterol), whereas a further breakdown of lipid classes would provide a better mechanistic understanding of the regulation of lipid homeostasis by miR-122. Furthermore, only one prospective study has investigated the association of baseline miR-122 levels with subsequent development of metabolic and cardiovascular diseases. Because prospective studies measure miR-122 well before the outcome of interest, they can establish a temporal relationship, are less prone to biases, and therefore are superior to cross-sectional and case-control studies.
Detection of miRNAs in plasma and serum by qPCR offers an opportunity for large-scale epidemiological studies into the role of miRNAs in human diseases. However, to prove clinical utility as a biomarker, more studies are needed that establish and define clinical cut-offs, quantify within-person variability over time and potential circadian fluctuations,90 provide assay standardization, and give more insights into the dependence of results on specimen type and storage requirements. Our in-house data show that, among a range of miRNAs, miR-122 showed the highest stability for repeat measurements taken 5 years apart. The pre-analytic requirements with regards to sample preparation that have become crucial in the study of circulating miRNAs, given the recent evidence for a predominant platelet origin,56 do not equally apply to miR-122, as this miRNA is liver specific and shows reproducible levels across different types of samples (e.g. serum, platelet-poor and platelet-rich plasma). Statins reduce the levels of miRNA-122 in circulation, whereas other types of medications such as platelet inhibitors do not show an effect on miR-122 but alter the levels of platelet-derived miRNAs in the circulation.54,56 Hence, further studies are needed to obtain a better understanding of the different factors that can affect the hepatic release of miR-122 into circulation.
As a drug target, it will be challenging to advance miR-122 to a licenced therapy given that duration, method, and degree of miR-122 lowering as well as the stage of liver pathology may all modify potential treatment effects. Also, the usefulness of miR-122 as a biomarker over and beyond the information provided by existing liver markers such as liver function enzymes (alanine aminotransferase, aspartate aminotransferase, or gamma-glutamyl transpeptidase) remains to be determined in detail.
Concluding remarks
Experimental studies and preliminary clinical data suggest that miR-122 may be relevant to common metabolic and cardiovascular diseases. Prospective epidemiological studies with long-term follow-up are required to confirm and further characterize this association in humans and to elaborate their predictive performance. If secreted liver miRNAs indeed emerge as useful biomarkers, this would offer the prospect of developing new diagnostics to complement existing markers in the early identification of people at risk of metabolic syndrome, type 2 diabetes, and associated complications.
Authors’ contributions
P.W. and P.S. performed statistical analyses. M.M. handled funding and supervision. P.W., P.S., S.K., and M.M. acquired the data. P.W. and M.M. conceived and designed the research. P.W., P.S., and M.M. drafted the manuscript. S.K. and C.F.-H. made critical revision of the manuscript for key intellectual content.
Funding
P.W. is an Erwin Schrödinger fellow in epidemiology of the Austrian Science Fund (J 3679-B13). M.M. is a senior fellow of the British Heart Foundation (FS/13/2/29892). The study was supported by a British Heart Foundation special project grant (SP/12/5/29574), the Fondation Leducq Transatlantic Network of Excellence in Cardiovascular Research (MIRVAD; 13 CVD, to M.M. and C.F-H.), Diabetes UK (12/0004530), and an excellence initiative [Competence Centers for Excellent Technologies (COMET)] of the Austrian Research Promotion Agency FFG: ‘Research Center of Excellence in Vascular Ageing – Tyrol, VASCage’ (K-Project number 843536). This work was also supported by the NIHR Biomedical Research Center based at Guy's and St Thomas’ National Health Service Foundation Trust and King's College London in partnership with King's College Hospital. This work was also supported by grants from the National Institutes of Health (R01HL107953, and R01HL106063 to C.F-H.). C.F-H. is supported by an Established Investigator Award from the American Heart Association (16EIA27550004).
Conflict of interest: King's College London and Medical University of Innsbruck have filed patent applications on miRNAs as biomarkers.
References
- 1.Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell 2009;136:215–233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Meijer HA, Smith EM, Bushell M. Regulation of miRNA strand selection: follow the leader? Biochem Soc Trans 2014;42:1135–1140. [DOI] [PubMed] [Google Scholar]
- 3.Small EM, Olson EN. Pervasive roles of microRNAs in cardiovascular biology. Nature 2011;469:336–342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Hoefer IE, Steffens S, Ala-Korpela M, Bäck M, Badimon L, Bochaton-Piallat ML, Boulanger CM, Caligiuri G, Dimmeler S, Egido J, Evans PC, Guzik T, Kwak BR, Landmesser U, Mayr M, Monaco C, Pasterkamp G, Tuñón J, Weber C, ESC Working Group Atherosclerosis and Vascular Biology. Novel methodologies for biomarker discovery in atherosclerosis. Eur Heart J 2015;36:2635–2642. [DOI] [PubMed] [Google Scholar]
- 5.Arner P, Kulyté A. MicroRNA regulatory networks in human adipose tissue and obesity. Nat Rev Endocrinol 2015;11:276–288. [DOI] [PubMed] [Google Scholar]
- 6.Schober A, Nazari-Jahantigh M, Weber C. MicroRNA-mediated mechanisms of the cellular stress response in atherosclerosis. Nat Rev Cardiol 2015;12:361–374. [DOI] [PubMed] [Google Scholar]
- 7.Kang MH, Zhang LH, Wijesekara N, de Haan W, Butland S, Bhattacharjee A, Hayden MR. Regulation of ABCA1 protein expression and function in hepatic and pancreatic islet cells by miR-145. Arterioscler Thromb Vasc Biol 2013;33:2724–2732. [DOI] [PubMed] [Google Scholar]
- 8.Goedeke L, Rotllan N, Canfrán-Duque A, Aranda JF, Ramrez CM, Araldi E, Lin CS, Anderson NN, Wagschal A, de Cabo R, Horton JD, Lasunción MA, Näär AM, Suárez Y, Fernández-Hernando C. MicroRNA-148a regulates LDL receptor and ABCA1 expression to control circulating lipoprotein levels. Nat Med 2015;21:1280–1289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Goedeke L, Vales-Lara FM, Fenstermaker M, Cirera-Salinas D, Chamorro-Jorganes A, Ramrez CM, Mattison JA, de Cabo R, Suárez Y, Fernández-Hernando C. A regulatory role for microRNA 33* in controlling lipid metabolism gene expression. Mol Cell Biol 2013;33:2339–2352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Wagschal A, Najafi-Shoushtari SH, Wang L, Goedeke L, Sinha S, deLemos AS, Black JC, Ramrez CM, Li Y, Tewhey R, Hatoum I, Shah N, Lu Y, Kristo F, Psychogios N, Vrbanac V, Lu YC, Hla T, de Cabo R, Tsang JS, Schadt E, Sabeti PC, Kathiresan S, Cohen DE, Whetstine J, Chung RT, Fernández-Hernando C, Kaplan LM, Bernards A, Gerszten RE, Näär AM. Genome-wide identification of microRNAs regulating cholesterol and triglyceride homeostasis. Nat Med 2015;21:1290–1297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Sun D, Zhang J, Xie J, Wei W, Chen M, Zhao X. MiR-26 controls LXR-dependent cholesterol efflux by targeting ABCA1 and ARL7. FEBS Lett 2012;586:1472–1479. [DOI] [PubMed] [Google Scholar]
- 12.He Y, Lin L, Cao J, Mao X, Qu Y, Xi B. Up-regulated miR-93 contributes to coronary atherosclerosis pathogenesis through targeting ABCA1. Int J Clin Exp Med 2015;8:674–681. [PMC free article] [PubMed] [Google Scholar]
- 13.Lv YC, Tang YY, Peng J, Zhao GJ, Yang J, Yao F, Ouyang XP, He PP, Xie W, Tan YL, Zhang M, Liu D, Tang DP, Cayabyab FS, Zheng XL, Zhang DW, Tian GP, Tang CK. MicroRNA-19b promotes macrophage cholesterol accumulation and aortic atherosclerosis by targeting ATP-binding cassette transporter A1. Atherosclerosis 2014;236:215–226. [DOI] [PubMed] [Google Scholar]
- 14.Zhang M, Wu JF, Chen WJ, Tang SL, Mo ZC, Tang YY, Li Y, Wang JL, Liu XY, Peng J, Chen K, He PP, Lv YC, Ouyang XP, Yao F, Tang DP, Cayabyab FS, Zhang DW, Zheng XL, Tian GP, Tang CK. MicroRNA-27a/b regulates cellular cholesterol efflux, influx and esterification/hydrolysis in THP-1 macrophages. Atherosclerosis 2014;234:54–64. [DOI] [PubMed] [Google Scholar]
- 15.Zhang N, Lei J, Lei H, Ruan X, Liu Q, Chen Y, Huang W. MicroRNA-101 overexpression by IL-6 and TNF-α inhibits cholesterol efflux by suppressing ATP-binding cassette transporter A1 expression. Exp Cell Res 2015;336:33–42. [DOI] [PubMed] [Google Scholar]
- 16.Meiler S, Baumer Y, Toulmin E, Seng K, Boisvert WA. MicroRNA 302a is a novel modulator of cholesterol homeostasis and atherosclerosis. Arterioscler Thromb Vasc Biol 2015;35:323–331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Vickers KC, Landstreet SR, Levin MG, Shoucri BM, Toth CL, Taylor RC, Palmisano BT, Tabet F, Cui HL, Rye KA, Sethupathy P, Remaley AT. MicroRNA-223 coordinates cholesterol homeostasis. Proc Natl Acad Sci USA 2014;111:14518–14523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Rayner KJ, Suárez Y, Dávalos A, Parathath S, Fitzgerald ML, Tamehiro N, Fisher EA, Moore KJ, Fernández-Hernando C. MiR-33 contributes to the regulation of cholesterol homeostasis. Science 2010;328:1570–1573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Dávalos A, Goedeke L, Smibert P, Ramrez CM, Warrier NP, Andreo U, Cirera-Salinas D, Rayner K, Suresh U, Pastor-Pareja JC, Esplugues E, Fisher EA, Penalva LOF, Moore KJ, Suárez Y, Lai EC, Fernández-Hernando C. miR-33a/b contribute to the regulation of fatty acid metabolism and insulin signaling. Proc Natl Acad Sci USA 2011;108:9232–9237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Najafi-Shoushtari SH, Kristo F, Li Y, Shioda T, Cohen DE, Gerszten RE, Näär AM. MicroRNA-33 and the SREBP host genes cooperate to control cholesterol homeostasis. Science 2010;328:1566–1569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Rayner KJ, Sheedy FJ, Esau CC, Hussain FN, Temel RE, Parathath S, van Gils JM, Rayner AJ, Chang AN, Suarez Y, Fernandez-Hernando C, Fisher EA, Moore KJ. Antagonism of miR-33 in mice promotes reverse cholesterol transport and regression of atherosclerosis. J Clin Invest 2011;121:2921–2931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Rotllan N, Ramrez CM, Aryal B, Esau CC, Fernández-Hernando C. Therapeutic silencing of microRNA-33 inhibits the progression of atherosclerosis in Ldlr-/- mice—brief report. Arterioscler Thromb Vasc Biol 2013;33:1973–1977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Horie T, Baba O, Kuwabara Y, Chujo Y, Watanabe S, Kinoshita M, Horiguchi M, Nakamura T, Chonabayashi K, Hishizawa M, Hasegawa K, Kume N, Yokode M, Kita T, Kimura T, Ono K. MicroRNA-33 deficiency reduces the progression of atherosclerotic plaque in ApoE-/- mice. J Am Heart Assoc 2012;1:e003376. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Marquart TJ, Wu J, Lusis AJ, Baldán Á. Anti-miR-33 therapy does not alter the progression of atherosclerosis in low-density lipoprotein receptor-deficient mice. Arterioscler Thromb Vasc Biol 2013;33:455–458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Karunakaran D, Thrush AB, Nguyen MA, Richards L, Geoffrion M, Singaravelu R, Ramphos E, Shangari P, Ouimet M, Pezacki JP, Moore KJ, Perisic L, Maegdefessel L, Hedin U, Harper ME, Rayner KJ. Macrophage mitochondrial energy status regulates cholesterol efflux and is enhanced by anti-miR33 in atherosclerosis. Circ Res 2015;117:266–278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Allen RM, Marquart TJ, Jesse JJ, Baldán A. Control of very low-density lipoprotein secretion by N-ethylmaleimide-sensitive factor and miR-33. Circ Res 2014;115:10–22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Horie T, Nishino T, Baba O, Kuwabara Y, Nakao T, Nishiga M, Usami S, Izuhara M, Sowa N, Yahagi N, Shimano H, Matsumura S, Inoue K, Marusawa H, Nakamura T, Hasegawa K, Kume N, Yokode M, Kita T, Kimura T, Ono K. MicroRNA-33 regulates sterol regulatory element-binding protein 1 expression in mice. Nat Commun 2013;4:2883. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Rayner KJ, Esau CC, Hussain FN, McDaniel AL, Marshall SM, van Gils JM, Ray TD, Sheedy FJ, Goedeke L, Liu X, Khatsenko OG, Kaimal V, Lees CJ, Fernandez-Hernando C, Fisher EA, Temel RE, Moore KJ. Inhibition of miR-33a/b in non-human primates raises plasma HDL and lowers VLDL triglycerides. Nature 2011;478:404–407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Goedeke L, Salerno A, Ramrez CM, Guo L, Allen RM, Yin X, Langley SR, Esau C, Wanschel A, Fisher EA, Suárez Y, Baldán A, Mayr M, Fernández-Hernando C. Long-term therapeutic silencing of miR-33 increases circulating triglyceride levels and hepatic lipid accumulation in mice. EMBO Mol Med 2014;6:1133–1141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Karunakaran D, Richards L, Geoffrion M, Barrette D, Gotfrit RJ, Harper ME, Rayner KJ. Therapeutic inhibition of miR-33 promotes fatty acid oxidation but does not ameliorate metabolic dysfunction in diet-induced obesity. Arterioscler Thromb Vasc Biol 2015;35:2536–2543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Soh J, Iqbal J, Queiroz J, Fernandez-Hernando C, Hussain MM. MicroRNA-30c reduces hyperlipidemia and atherosclerosis in mice by decreasing lipid synthesis and lipoprotein secretion. Nat Med 2013;19:892–900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Wijnen WJ, van der Made I, van den Oever S, Hiller M, de Boer BA, Picavet DI, Chatzispyrou IA, Houtkooper RH, Tijsen AJ, Hagoort J, van Veen H, Everts V, Ruijter JM, Pinto YM, Creemers EE. Cardiomyocyte-specific miRNA-30c over-expression causes dilated cardiomyopathy. PLoS One 2014;9:e96290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.de Aguiar Vallim TQ, Tarling EJ, Kim T, Civelek M, Baldán Á, Esau C, Edwards PA. MicroRNA-144 regulates hepatic ATP binding cassette transporter A1 and plasma high-density lipoprotein after activation of the nuclear receptor farnesoid X receptor. Circ Res 2013;112:1602–1612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Ramrez CM, Rotllan N, Vlassov AV, Dávalos A, Li M, Goedeke L, Aranda JF, Cirera-Salinas D, Araldi E, Salerno A, Wanschel A, Zavadil J, Castrillo A, Kim J, Suárez Y, Fernández-Hernando C. Control of cholesterol metabolism and plasma high-density lipoprotein levels by microRNA-144. Circ Res 2013;112:1592–1601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Hu YW, Hu YR, Zhao JY, Li SF, Ma X, Wu SG, Lu JB, Qiu YR, Sha YH, Wang YC, Gao JJ, Zheng L, Wang Q. An agomir of miR-144-3p accelerates plaque formation through impairing reverse cholesterol transport and promoting pro-inflammatory cytokine production. PLoS One 2014;9:e94997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Global Lipids Genetics Consortium. Discovery and refinement of loci associated with lipid levels. Nat Genet 2013;45:1274–1283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Do R, Willer CJ, Schmidt EM, Sengupta S, Gao C, Peloso GM, Gustafsson S, Kanoni S, Ganna A, Chen J, Buchkovich ML, Mora S, Beckmann JS, Bragg-Gresham JL, Chang HY, Demirkan A, Den Hertog HM, Donnelly LA, Ehret GB, Esko T, Feitosa MF, Ferreira T, Fischer K, Fontanillas P, Fraser RM, Freitag DF, Gurdasani D, Heikkilä K, Hyppönen E, Isaacs A, Jackson AU, Johansson A, Johnson T, Kaakinen M, Kettunen J, Kleber ME, Li X, Luan J, Lyytikäinen LP, Magnusson PKE, Mangino M, Mihailov E, Montasser ME, Müller-Nurasyid M, Nolte IM, O'Connell JR, Palmer CD, Perola M, Petersen AK, Sanna S, Saxena R, Service SK, Shah S, Shungin D, Sidore C, Song C, Strawbridge RJ, Surakka I, Tanaka T, Teslovich TM, Thorleifsson G, Van den Herik EG, Voight BF, Volcik KA, Waite LL, Wong A, Wu Y, Zhang W, Absher D, Asiki G, Barroso I, Been LF, Bolton JL, Bonnycastle LL, Brambilla P, Burnett MS, Cesana G, Dimitriou M, Doney ASF, Döring A, Elliott P, Epstein SE, Eyjolfsson GI, Gigante B, Goodarzi MO, Grallert H, Gravito ML, Groves CJ, Hallmans G, Hartikainen AL, Hayward C, Hernandez D, Hicks AA, Holm H, Hung YJ, Illig T, Jones MR, Kaleebu P, Kastelein JJP, Khaw KT, Kim E, Klopp N, Komulainen P, Kumari M, Langenberg C, Lehtimäki T, Lin SY, Lindström J, Loos RJF, Mach F, McArdle WL, Meisinger C, Mitchell BD, Müller G, Nagaraja R, Narisu N, Nieminen TVM, Nsubuga RN, Olafsson I, Ong KK, Palotie A, Papamarkou T, Pomilla C, Pouta A, Rader DJ, Reilly MP, Ridker PM, Rivadeneira F, Rudan I, Ruokonen A, Samani N, Scharnagl H, Seeley J, Silander K, Stancáková A, Stirrups K, Swift AJ, Tiret L, Uitterlinden AG, van Pelt LJ, Vedantam S, Wainwright N, Wijmenga C, Wild SH, Willemsen G, Wilsgaard T, Wilson JF, Young EH, Zhao JH, Adair LS, Arveiler D, Assimes TL, Bandinelli S, Bennett F, Bochud M, Boehm BO, Boomsma DI, Borecki IB, Bornstein SR, Bovet P, Burnier M, Campbell H, Chakravarti A, Chambers JC, Chen YDI, Collins FS, Cooper RS, Danesh J, Dedoussis G, de Faire U, Feranil AB, Ferrières J, Ferrucci L, Freimer NB, Gieger C, Groop LC, Gudnason V, Gyllensten U, Hamsten A, Harris TB, Hingorani A, Hirschhorn JN, Hofman A, Hovingh GK, Hsiung CA, Humphries SE, Hunt SC, Hveem K, Iribarren C, Järvelin MR, Jula A, Kähönen M, Kaprio J, Kesäniemi A, Kivimaki M, Kooner JS, Koudstaal PJ, Krauss RM, Kuh D, Kuusisto J, Kyvik KO, Laakso M, Lakka TA, Lind L, Lindgren CM, Martin NG, März W, McCarthy MI, McKenzie CA, Meneton P, Metspalu A, Moilanen L, Morris AD, Munroe PB, Njølstad I, Pedersen NL, Power C, Pramstaller PP, Price JF, Psaty BM, Quertermous T, Rauramaa R, Saleheen D, Salomaa V, Sanghera DK, Saramies J, Schwarz PEH, Sheu WHH, Shuldiner AR, Siegbahn A, Spector TD, Stefansson K, Strachan DP, Tayo BO, Tremoli E, Tuomilehto J, Uusitupa M, van Duijn CM, Vollenweider P, Wallentin L, Wareham NJ, Whitfield JB, Wolffenbuttel BHR, Altshuler D, Ordovas JM, Boerwinkle E, Palmer CNA, Thorsteinsdottir U, Chasman DI, Rotter JI, Franks PW, Ripatti S, Cupples LA, Sandhu MS, Rich SS, Boehnke M, Deloukas P, Mohlke KL, Ingelsson E, Abecasis GR, Daly MJ, Neale BM, Kathiresan S. Common variants associated with plasma triglycerides and risk for coronary artery disease. Nat Genet 2013;45:1345–1352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Mitchell PS, Parkin RK, Kroh EM, Fritz BR, Wyman SK, Pogosova-Agadjanyan EL, Peterson A, Noteboom J, O'Briant KC, Allen A, Lin DW, Urban N, Drescher CW, Knudsen BS, Stirewalt DL, Gentleman R, Vessella RL, Nelson PS, Martin DB, Tewari M. Circulating microRNAs as stable blood-based markers for cancer detection. Proc Natl Acad Sci USA 2008;105:10513–10518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Rottiers V, Näär AM. MicroRNAs in metabolism and metabolic disorders. Nat Rev Mol Cell Biol 2012;13:239–250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Esau C, Davis S, Murray SF, Yu XX, Pandey SK, Pear M, Watts L, Booten SL, Graham M, McKay R, Subramaniam A, Propp S, Lollo BA, Freier S, Bennett CF, Bhanot S, Monia BP. miR-122 regulation of lipid metabolism revealed by in vivo antisense targeting. Cell Metab 2006;3:87–98. [DOI] [PubMed] [Google Scholar]
- 41.Krützfeldt J, Rajewsky N, Braich R, Rajeev KG, Tuschl T, Manoharan M, Stoffel M. Silencing of microRNAs in vivo with ‘antagomirs’. Nature 2005;438:685–689. [DOI] [PubMed] [Google Scholar]
- 42.Elmén J, Lindow M, Schütz S, Lawrence M, Petri A, Obad S, Lindholm M, Hedtjärn M, Hansen HF, Berger U, Gullans S, Kearney P, Sarnow P, Straarup EM, Kauppinen S. LNA-mediated microRNA silencing in non-human primates. Nature 2008;452:896–899. [DOI] [PubMed] [Google Scholar]
- 43.Lanford RE, Hildebrandt-Eriksen ES, Petri A, Persson R, Lindow M, Munk ME, Kauppinen S, Ørum H. Therapeutic silencing of microRNA-122 in primates with chronic hepatitis C virus infection. Science 2010;327:198–201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Fernández-Hernando C, Ramrez CM, Goedeke L, Suárez Y. MicroRNAs in metabolic disease. Arterioscler Thromb Vasc Biol 2013;33:178–185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Castoldi M, Vujic Spasic M, Altamura S, Elmén J, Lindow M, Kiss J, Stolte J, Sparla R, D'Alessandro LA, Klingmüller U, Fleming RE, Longerich T, Gröne HJ, Benes V, Kauppinen S, Hentze MW, Muckenthaler MU. The liver-specific microRNA miR-122 controls systemic iron homeostasis in mice. J Clin Invest 2011;121:1386–1396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Susanto H, Wang CH. Mice lacking miR-122 improve metabolic syndrome through regulating lipid and glucose metabolism. 104 NCTU-NTHU graduate students symposium, Hsinchu, Taiwan: 2015; https://www.researchgate.net/publication/280572199_Mice_Lacking_miR-122_improve_metabolic_syndrome_through_regulating_lipid_and_glucose_metabolism (14 December 2015). [Google Scholar]
- 47.Tsai WC, Hsu SD, Hsu CS, Lai TC, Chen SJ, Shen R, Huang Y, Chen HC, Lee CH, Tsai TF, Hsu MT, Wu JC, Huang HD, Shiao MS, Hsiao M, Tsou AP. MicroRNA-122 plays a critical role in liver homeostasis and hepatocarcinogenesis. J Clin Invest 2012;122:2884–2897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Hsu SH, Wang B, Kota J, Yu J, Costinean S, Kutay H, Yu L, Bai S, La Perle K, Chivukula RR, Mao H, Wei M, Clark KR, Mendell JR, Caligiuri MA, Jacob ST, Mendell JT, Ghoshal K. Essential metabolic, anti-inflammatory, and anti-tumorigenic functions of miR-122 in liver. J Clin Invest 2012;122:2871–2883. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Thakral S, Ghoshal K. miR-122 is a unique molecule with great potential in diagnosis, prognosis of liver disease, and therapy both as miRNA mimic and antimir. Curr Gene Ther 2015;15:142–150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Li J, Ghazwani M, Zhang Y, Lu J, Li J, Fan J, Gandhi CR, Li S. miR-122 regulates collagen production via targeting hepatic stellate cells and suppressing P4HA1 expression. J Hepatol 2013;58:522–528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Dignat-George F, Boulanger CM. The many faces of endothelial microparticles. Arterioscler Thromb Vasc Biol 2011;31:27–33. [DOI] [PubMed] [Google Scholar]
- 52.Zampetaki A, Willeit P, Drozdov I, Kiechl S, Mayr M. Profiling of circulating microRNAs: from single biomarkers to re-wired networks. Cardiovasc Res 2012;93:555–562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Zampetaki A, Willeit P, Tilling L, Drozdov I, Prokopi M, Renard JM, Mayr A, Weger S, Schett G, Shah A, Boulanger CM, Willeit J, Chowienczyk PJ, Kiechl S, Mayr M. Prospective study on circulating MicroRNAs and risk of myocardial infarction. J Am Coll Cardiol 2012;60:290–299. [DOI] [PubMed] [Google Scholar]
- 54.Willeit P, Zampetaki A, Dudek K, Kaudewitz D, King AS, Kirkby NS, Crosby-Nwaobi R, Prokopi M, Drozdov I, Langley S, Sivaprasad S, Markus HS, Mitchell JA, Warner T, Kiechl S, Mayr M. Circulating microRNAs as novel biomarkers for platelet activation. Circ Res 2013;112:595–600. [DOI] [PubMed] [Google Scholar]
- 55.Mayr M, Zampetaki A, Willeit P, Willeit J, Kiechl S. MicroRNAs within the continuum of postgenomics biomarker discovery. Arterioscler Thromb Vasc Biol 2013;33:206–214. [DOI] [PubMed] [Google Scholar]
- 56.Kaudewitz D, Skroblin P, Bender LH, Barwari T, Willeit P, Pechlaner R, Sunderland NP, Willeit K, Morton AC, Armstrong PC, Chan MV, Lu R, Yin X, Gracio F, Dudek K, Langley SR, Zampetaki A, de Rinaldis E, Ye S, Warner TD, Saxena A, Kiechl S, Storey RF, Mayr M. Association of microRNAs and YRNAs with platelet function. Circ Res 2016;118:420–432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Liu X, Fan Z, Zhao T, Cao W, Zhang L, Li H, Xie Q, Tian Y, Wang B. Plasma miR-1, miR-208, miR-499 as potential predictive biomarkers for acute myocardial infarction: an independent study of Han population. Exp Gerontol 2015;72:230–238. [DOI] [PubMed] [Google Scholar]
- 58.Vliegenthart ADB, Shaffer JM, Clarke JI, Peeters LEJ, Caporali A, Bateman DN, Wood DM, Dargan PI, Craig DG, Moore JK, Thompson AI, Henderson NC, Webb DJ, Sharkey J, Antoine DJ, Park BK, Bailey MA, Lader E, Simpson KJ, Dear JW. Comprehensive microRNA profiling in acetaminophen toxicity identifies novel circulating biomarkers for human liver and kidney injury. Sci Rep 2015;5:15501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Kia R, Kelly L, Sison-Young RLC, Zhang F, Pridgeon CS, Heslop JA, Metcalfe P, Kitteringham NR, Baxter M, Harrison S, Hanley NA, Burke ZD, Storm MP, Welham MJ, Tosh D, Küppers-Munther B, Edsbagge J, Starkey Lewis PJ, Bonner F, Harpur E, Sidaway J, Bowes J, Fenwick SW, Malik H, Goldring CEP, Park BK. MicroRNA-122: a novel hepatocyte-enriched in vitro marker of drug-induced cellular toxicity. Toxicol Sci 2015;144:173–185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Huang X, Yuan T, Tschannen M, Sun Z, Jacob H, Du M, Liang M, Dittmar RL, Liu Y, Liang M, Kohli M, Thibodeau SN, Boardman L, Wang L. Characterization of human plasma-derived exosomal RNAs by deep sequencing. BMC Genomics 2013;14:319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Gallo A, Tandon M, Alevizos I, Illei GG. The majority of microRNAs detectable in serum and saliva is concentrated in exosomes. PLoS One 2012;7:e30679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Wang S, Aurora AB, Johnson BA, Qi X, McAnally J, Hill JA, Richardson JA, Bassel-Duby R, Olson EN. The endothelial-specific microRNA miR-126 governs vascular integrity and angiogenesis. Dev Cell 2008;15:261–271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Miyaaki H, Ichikawa T, Kamo Y, Taura N, Honda T, Shibata H, Milazzo M, Fornari F, Gramantieri L, Bolondi L, Nakao K. Significance of serum and hepatic microRNA-122 levels in patients with non-alcoholic fatty liver disease. Liver Int 2014;34:e302–e307. [DOI] [PubMed] [Google Scholar]
- 64.Cermelli S, Ruggieri A, Marrero JA, Ioannou GN, Beretta L. Circulating microRNAs in patients with chronic hepatitis C and non-alcoholic fatty liver disease. PLoS One 2011;6:e23937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Yamada H, Suzuki K, Ichino N, Ando Y, Sawada A, Osakabe K, Sugimoto K, Ohashi K, Teradaira R, Inoue T, Hamajima N, Hashimoto S. Associations between circulating microRNAs (miR-21, miR-34a, miR-122 and miR-451) and non-alcoholic fatty liver. Clin Chim Acta 2013;424:99–103. [DOI] [PubMed] [Google Scholar]
- 66.Pirola CJ, Fernández Gianotti T, Castaño GO, Mallardi P, San Martino J, Mora Gonzalez Lopez Ledesma M, Flichman D, Mirshahi F, Sanyal AJ, Sookoian S. Circulating microRNA signature in non-alcoholic fatty liver disease: from serum non-coding RNAs to liver histology and disease pathogenesis. Gut 2015;64:800–812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Becker PP, Rau M, Schmitt J, Malsch C, Hammer C, Bantel H, Müllhaupt B, Geier A. Performance of serum microRNAs -122, -192 and -21 as biomarkers in patients with non-alcoholic steatohepatitis. PLoS One 2015;10:e0142661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Celikbilek M, Baskol M, Taheri S, Deniz K, Dogan S, Zararsiz G, Gursoy S, Guven K, Ozbakr O, Dundar M, Yucesoy M. Circulating microRNAs in patients with non-alcoholic fatty liver disease. World J Hepatol 2014;6:613–620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Tan Y, Ge G, Pan T, Wen D, Gan J. A pilot study of serum microRNAs panel as potential biomarkers for diagnosis of nonalcoholic fatty liver disease. PLoS One 2014;9:e105192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Liu W, Cao H, Yan J, Huang R, Ying H. ‘Micro-managers’ of hepatic lipid metabolism and NAFLD. Wiley Interdiscip Rev RNA 2015;6:581–593. [DOI] [PubMed] [Google Scholar]
- 71.Stegemann C, Pechlaner R, Willeit P, Langley SR, Mangino M, Mayr U, Menni C, Moayyeri A, Santer P, Rungger G, Spector TD, Willeit J, Kiechl S, Mayr M. Lipidomics profiling and risk of cardiovascular disease in the prospective population-based Bruneck study. Circulation 2014;129:1821–1831. [DOI] [PubMed] [Google Scholar]
- 72.Momen-Heravi F, Saha B, Kodys K, Catalano D, Satishchandran A, Szabo G. Increased number of circulating exosomes and their microRNA cargos are potential novel biomarkers in alcoholic hepatitis. J Transl Med 2015;13:261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Szabo G, Csak T. Role of microRNAs in NAFLD/NASH. Dig Dis Sci 2016. [Epub ahead of print]. [DOI] [PubMed] [Google Scholar]
- 74.Ng R, Wu H, Xiao H, Chen X, Willenbring H, Steer CJ, Song G. Inhibition of microRNA-24 expression in liver prevents hepatic lipid accumulation and hyperlipidemia. Hepatology 2014;60:554–564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Mattis AN, Song G, Hitchner K, Kim RY, Lee AY, Sharma AD, Malato Y, McManus MT, Esau CC, Koller E, Koliwad S, Lim LP, Maher JJ, Raffai RL, Willenbring H. A screen in mice uncovers repression of lipoprotein lipase by microRNA-29a as a mechanism for lipid distribution away from the liver. Hepatology 2015;61:141–152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Gao W, He HW, Wang ZM, Zhao H, Lian XQ, Wang YS, Zhu J, Yan JJ, Zhang DG, Yang ZJ, Wang LS. Plasma levels of lipometabolism-related miR-122 and miR-370 are increased in patients with hyperlipidemia and associated with coronary artery disease. Lipids Health Dis 2012;11:55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Wang R, Hong J, Cao Y, Shi J, Gu W, Ning G, Zhang Y, Wang W. Elevated circulating microRNA-122 is associated with obesity and insulin resistance in young adults. Eur J Endocrinol 2015;172:291–300. [DOI] [PubMed] [Google Scholar]
- 78.Ortega FJ, Mercader JM, Catalán V, Moreno-Navarrete JM, Pueyo N, Sabater M, Gómez-Ambrosi J, Anglada R, Fernández-Formoso JA, Ricart W, Frühbeck G, Fernández-Real JM. Targeting the circulating microRNA signature of obesity. Clin Chem 2013;59:781–792. [DOI] [PubMed] [Google Scholar]
- 79.Janssen HLA, Reesink HW, Lawitz EJ, Zeuzem S, Rodriguez-Torres M, Patel K, van der Meer AJ, Patick AK, Chen A, Zhou Y, Persson R, King BD, Kauppinen S, Levin AA, Hodges MR. Treatment of HCV infection by targeting microRNA. N Engl J Med 2013;368:1685–1694. [DOI] [PubMed] [Google Scholar]
- 80.Willeit P, Yin X, Kaudewitz D, Skroblin P, Zampetaki A, Moschen AR, Ramirez CM, Goedeke L, Rotllan N, Bonora E, Hughes AD, Weger S, Fernandez-Hernando C, Tilg H, Willeit J, Kiechl S, Mayr M. Abstract 17961: circulating MicroRNA-122 is associated with incident metabolic syndrome and type-2-diabetes. Circulation 2015;132:A17961. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Li X, Yang Y, Wang L, Qiao S, Lu X, Wu Y, Xu B, Li H, Gu D. Plasma miR-122 and miR-3149 potentially novel biomarkers for acute coronary syndrome. PLoS One 2015;10:e0125430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.D'Alessandra Y, Carena MC, Spazzafumo L, Martinelli F, Bassetti B, Devanna P, Rubino M, Marenzi G, Colombo GI, Achilli F, Maggiolini S, Capogrossi MC, Pompilio G. Diagnostic potential of plasmatic microRNA signatures in stable and unstable angina. PLoS One 2013;8:e80345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Corsten MF, Dennert R, Jochems S, Kuznetsova T, Devaux Y, Hofstra L, Wagner DR, Staessen JA, Heymans S, Schroen B. Circulating microRNA-208b and microRNA-499 reflect myocardial damage in cardiovascular disease. Circ Cardiovasc Genet 2010;3:499–506. [DOI] [PubMed] [Google Scholar]
- 84.D'Alessandra Y, Devanna P, Limana F, Straino S, Di Carlo A, Brambilla PG, Rubino M, Carena MC, Spazzafumo L, De Simone M, Micheli B, Biglioli P, Achilli F, Martelli F, Maggiolini S, Marenzi G, Pompilio G, Capogrossi MC. Circulating microRNAs are new and sensitive biomarkers of myocardial infarction. Eur Heart J 2010;31:2765–2773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Jickling GC, Ander BP, Zhan X, Noblett D, Stamova B, Liu D. microRNA expression in peripheral blood cells following acute ischemic stroke and their predicted gene targets. PLoS One 2014;9:e99283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Novák J, Olejncková V, Tkácová N, Santulli G. Mechanistic role of microRNAs in coupling lipid metabolism and atherosclerosis. Adv Exp Med Biol 2015;887:79–100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Ouimet M, Ediriweera HN, Gundra UM, Sheedy FJ, Ramkhelawon B, Hutchison SB, Rinehold K, van Solingen C, Fullerton MD, Cecchini K, Rayner KJ, Steinberg GR, Zamore PD, Fisher EA, Loke P, Moore KJ. MicroRNA-33-dependent regulation of macrophage metabolism directs immune cell polarization in atherosclerosis. J Clin Invest 2015;125:4334–4348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Santovito D, Egea V, Weber C. Small but smart: microRNAs orchestrate atherosclerosis development and progression. Biochim Biophys Acta 2015. [Epub ahead of print]. [DOI] [PubMed] [Google Scholar]
- 89.Di Gregoli K, Jenkins N, Salter R, White S, Newby AC, Johnson JL. MicroRNA-24 regulates macrophage behavior and retards atherosclerosis. Arterioscler Thromb Vasc Biol 2014;34:1990–2000. [DOI] [PubMed] [Google Scholar]
- 90.Gatfield D, Le Martelot G, Vejnar CE, Gerlach D, Schaad O, Fleury-Olela F, Ruskeepää AL, Oresic M, Esau CC, Zdobnov EM, Schibler U. Integration of microRNA miR-122 in hepatic circadian gene expression. Genes Dev 2009;23:1313–1326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Chen C, Wang Y, Yang S, Li H, Zhao G, Wang F, Yang L, Wang DW. MiR-320a contributes to atherogenesis by augmenting multiple risk factors and down-regulating SRF. J Cell Mol Med 2015;19:970–985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Vickers KC, Shoucri BM, Levin MG, Wu H, Pearson DS, Osei-Hwedieh D, Collins FS, Remaley AT, Sethupathy P. MicroRNA-27b is a regulatory hub in lipid metabolism and is altered in dyslipidemia. Hepatology 2013;57:533–542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Goedeke L, Rotllan N, Ramrez CM, Aranda JF, Canfrán-Duque A, Araldi E, Fernández-Hernando A, Langhi C, de Cabo R, Baldán Á, Suárez Y, Fernández-Hernando C. miR-27b inhibits LDLR and ABCA1 expression but does not influence plasma and hepatic lipid levels in mice. Atherosclerosis 2015;243:499–509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Alvarez ML, Khosroheidari M, Eddy E, Done SC. MicroRNA-27a decreases the level and efficiency of the LDL receptor and contributes to the dysregulation of cholesterol homeostasis. Atherosclerosis 2015;242:595–604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Carrer M, Liu N, Grueter CE, Williams AH, Frisard MI, Hulver MW, Bassel-Duby R, Olson EN. Control of mitochondrial metabolism and systemic energy homeostasis by microRNAs 378 and 378*. Proc Natl Acad Sci USA 2012;109:15330–15335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Nakanishi N, Nakagawa Y, Tokushige N, Aoki N, Matsuzaka T, Ishii K, Yahagi N, Kobayashi K, Yatoh S, Takahashi A, Suzuki H, Urayama O, Yamada N, Shimano H. The up-regulation of microRNA-335 is associated with lipid metabolism in liver and white adipose tissue of genetically obese mice. Biochem Biophys Res Commun 2009;385:492–496. [DOI] [PubMed] [Google Scholar]
- 97.Iliopoulos D, Drosatos K, Hiyama Y, Goldberg IJ, Zannis VI. MicroRNA-370 controls the expression of microRNA-122 and Cpt1alpha and affects lipid metabolism. J Lipid Res 2010;51:1513–1523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Ou Z, Wada T, Gramignoli R, Li S, Strom SC, Huang M, Xie W. MicroRNA hsa-miR-613 targets the human LXRα gene and mediates a feedback loop of LXRα autoregulation. Mol Endocrinol 2011;25:584–596. [DOI] [PMC free article] [PubMed] [Google Scholar]