

SUMMARY
Zika virus (ZKV) infection is a huge public health problem in Brazil because of the increased incidence of microcephaly in neonates from infected mothers. Detection of specific IgG antibodies in maternal serum samples constitutes an important approach for diagnosing ZKV infection and evaluating its relationship with neonatal microcephaly. However, as there is no serological test produced in Brazil to detect IgM and IgG antibodies against ZKV, we sought to examine specific IgG in serum samples from patients or suspected mothers to detect previous infection and to test for specificity with regard to flaviviral infections occurring in the same area. Brazilian Zika virus native antigens were obtained from infected Vero cell layers or free virions in the culture medium and then used in ELISA. We tested sera from eight ZKV RNA-diagnosed infected patients (ZKVR), seven neonates with microcephaly and their mothers after delivery (MM), 140 dengue virus IgM-positive (DM) and IgG (DG)-positive patients, and 100 yellow fever (YF)-vaccinated patients. According to the ELISA, ZKVR samples were mostly positive (7/8), and all the MM serum samples were positive for ZKV IgG (7/7). In contrast, cross-reactions for dengue or yellow fever-vaccinated patients were observed, including DM (48/95), DG (10/45) or YF (3/100) serum samples; however, these cross-reactions exhibited low antigen avidity so that 6 M urea largely removed this cross-reactivity, with only a few cross-reacting samples remaining (8/140). ELISA based on extracted virions was much more specific, with all ZKVR (8/8) and MM sera being positive for ZKV IgG (7/7) and only borderline cross-reactivity found for DM (6/95), DG (3/45) or YF (4/100)-vaccinated serum samples. This technique (ELISA) can identify specific IgG in ZKV-infected patients and may be helpful in diagnosing congenital infetions after maternal RNA virus clearance or in epidemiological studies.
KEYWORDS: Zika virus, Dengue virus, Congenital infection, IgG, Serology, Avidity.
INTRODUCTION
Congenital infections are acute infections whose etiological agents cross the placental barrier, infecting the fetuses, preventing maternal immune cells from controlling the infections 1 . Despite the control of acute infection in the mother, the agent usually causes several progressive lesions in the fetus because the fetal immune system is still developing 2 . Unknown fetal lesions are mainly diagnosed by ultrasound imaging, which is improving but fails to detect minute early lesions 3 . Some congenital infections are treatable, such as toxoplasmosis, and huge efforts have been made to detect fetal infections as early as possible by applying several serological approaches to maternal serum samples so as to accurately date the infection, and thus estimating the gestation age at which the vertical transmission has taken place 4 . Most causative agents of congenital infections have a high prevalence and low morbidity in the general population, constituting a health problem when a pregnant woman presents with acute infection 5 . Indeed, the immunity controlling these infections usually protects the mother and their fetuses, but this scenario typically requires screening of acute infections in a population with a large fraction of immune individuals, as for toxoplasmosis 6 . A specific IgG is a marker of a past contact with agents, and its detection only confirms a previous infection of the host. IgG avidity is a useful marker of recent acute infections such as rubella and toxoplasmosis, allowing the distinction between chronic non-pregnancy-related infections and acute infections. This approach is preferred over the specific IgM detection, which has lost efficiency due to the appearance on the market of highly sensitivity assays 7 .
Zika virus (ZKV) infection has become epidemic in Northeast Brazil 8 and in the rest of the world due to the spread of the virus and the presence of Aedes aegypti mosquitoes, the most probable culicine vector 9 . Since December 2015, an increasing number of neonates with microcephaly has been reported and attributed to ZKV maternal infection associated with an epidemic of ZKV in South America 10 . When suspected, ZKV detection in amniotic fluid samples is mandatory 11 . However, in cases of a negative result, the absence of fetal infection cannot be ruled out because fetal Central Nervous System (CNS) infection can progress despite the lack of evidence of the virus in fetal body compartments 12 . Detection of specific IgG in maternal serum samples after delivery would be a very specific approach for diagnosing ZKV infection and to determine its relationship with eventual microcephaly in the neonates 13 . The presence of IgG antibody in the serum samples of mothers of neonates with microcephaly would identify ZKV infection, regardless of viral particles clearance, as RT-PCR only detects acute infections. Furthermore, IgG avidity assays will help in the detection of recent acute infection 14 .
Members of the Flavivirus family, such as ZKV, dengue virus, Yellow Fever virus and West Nile encephalitis viruses, are closely related and exhibit vector and antigen similarities, which poses a difficult challenge for producing a specific IgG ELISA 15 . According to genomic studies, flaviviruses share similar sequences, vectors and hosts 9 . However, the occurrence of distinct human diseases suggests specific immunity, regarding both antibodies and cells, which must result in a possible specific immune-based diagnosis. The ZKV strain from South America has been recently isolated, and this strain is available for cell culture. We used a classical in vitro cell infection and antigen production, as has been previously established for measles virus 16 and rubella virus antigens 17 , in the Virology Laboratory of the Institute of Tropical Medicine, University of Sao Paulo, Brazil, to standardize and validate an IgG ELISA for diagnosing ZKV virus infection in samples positive for ZKV RNA detection in acute disease or in serum samples from mothers or neonates with microcephaly. We compared the results of positive ZKV samples with those from patients infected with other flaviviruses.
MATERIAL AND METHODS
Zika virus antigen production
Zika virus
The Brazilian Zika virus strain used was isolated and provided by the Evandro Chagas Institute in Belém, Pará State, to the Institute of Biological Sciences, University of Sao Paulo (ICB/USP) 14 .
Cell and viral culture
African green monkey kidney (Vero) cells were maintained at 37 °C in Eagle's Minimal Essential Medium (MEM) containing 5% fetal bovine serum-FBS (Gibco(tm), Life Technologies(r), Carlsbad, Califórnia, USA) supplemented with sodium pyruvate (Gibco(r)), non-essential amino acids (Gibco(r)), 10 µg/mL gentamicin and 1 µM amphotericin B. Confluent monolayers of Vero cells were inoculated with the Brazilian Zika virus strain followed by virus adsorption at 37 °C for 1 h. The cells were maintained in the medium described above, with 2% FBS (Gibco(r)), and incubated at 37 °C for four days; the cells were observed daily until the cytopathic effect (CPE) occurred. When the cells showed 90% CPE, the media and cell layers were harvested for antigen production.
Electron microscopy
The medium of ZKV strain-infected Vero cells at 50% CPE was removed, and the cells were fixed with 2 mL buffered 4% glutaraldehyde for 1 h in an ice bath. After fixation, the cell monolayer was scraped and transferred to 1.5 mL centrifuge tubes. The suspension was centrifuged to 10,000 x g for 2 min, and the supernatant was removed. The pellet was suspended in 1% osmium tetroxide and maintained in an ice bath for 1 hour. The fixed cells were recovered by centrifugation at 10,000 x g for 2 min; the pellet was overlaid with phosphate-buffered saline pH 7.4 (PBS) and centrifuged at 10,000 x g for 10 min at 4 °C. The supernatant was removed. The dense pellet was processed for Araldite(r) embedding and sectioning, and 0.2 µM slices were observed using a Jeol Ultramicroscope (JEOL Ltd. Tokyo, Japan) with digital imaging.
Protein antigen from the cell layer
After four days of infection, the cell layer and medium was harvested and centrifuged at 2,000 x g at 4 °C for 20 min. The pellet was washed twice in PBS and centrifuged under the same conditions. The washed pellet was suspended in 1:50 of the original volume using PBS containing 0.5% sodium deoxycholate. The sample was sonicated in an ice bath using several 1 min pulses until maximum cell disruption was observed under phase-contrast microscopy. The suspension of disrupted cells was cleared by centrifugation at 14,000 rpm and 4 °C for 15 min and stored at -80 °C until use.
Protein antigen from ultracentrifuged virions
The cell culture medium obtained from the 2,000 x g centrifugation in the previous step was stored at -20 °C. Pooled samples from several 75 cm2 cultures were thawed, pooled and cleared by centrifugation at 10,000 x g at 4 °C for 10 min. The cleared supernatant was concentrated by ultracentrifugation at 100,000 x g at 4 °C for 2 h. The supernatant was carefully and completely removed, and the antigen pellet was suspended in TNE buffer (0.05 M Tris/HCl, 0.075 M NaCl, 10 mM EDTA, 0.5% DOC and 1% formaldehyde). The protein content was determined by the Bradford method, aliquoted and stored at -80 °C until use.
SDS-PAGE and Western blot analysis
Antigens were analyzed using 12.5% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). Briefly, 10 µL of protein at 0.5 mg/mL were mixed with 5 µL loading buffer (pH 7.4) containing 1% SDS, 25 mM Tris-HCl, 0.5% β-mercaptoethanol and 0.001% bromophenol blue. The sample was heated at 95 °C for 5 min before loading onto the gel. Staining was performed using Coomassie blue G250 (Sigma(r), St. Louis, Missouri, USA) reagent, followed by electrophoretic transfer to nitrocellulose membranes. Western blotting was performed following the Towbin protocol18. The membranes were blocked with 5% fat-free milk in PBS with 0.01% Tween-20 (PBS-T). Convalescent serum samples obtained from RNA-positive ZKV patients were incubated with the antigens for 18 h at 4 °C. After careful washing with PBS-T, bound antibodies were detected by incubation with peroxidase-conjugated anti-IgG for 1 h, followed by washing, and then reacted with 4-chloronaphthol developing solution for 30 min with digital recording.
Serum samples
The study population comprised 308 serum samples obtained from the biorepository collection of the Virology Laboratory (IMTSP, USP); the samples were stored at -80 °C until use. The serum samples were derived from four groups of patients: 95 samples from acute dengue patients with IgM anti-dengue virus (DM); 45 samples with isolated IgG anti-dengue virus (DG); 53 nonreactive samples, i.e., without dengue virus antibodies (N); 100 samples from yellow fever-vaccinated patients at 45 days after vaccination (YF); 7 samples from mothers (5) and neonates with microcephaly (2) with suspected ZKV infections, from São Paulo, Brazil, collected after birth (MM); and 8 samples collected from patients 15-20 days after the confirmation of diagnosis (Zika RNA detection) during an outbreak of ZIKV in Araraquara, Ribeirão Preto and São Paulo counties (São Paulo State). Dengue serology was performed using commercial IgM, IgG, and NS1 ELISA (Dengue Virus IgM and IgG DxSelect, Focus Diagnostics, Cypress, CA 90630 USA)
ELISA
We developed an in-house ELISA with high performance for detecting anti-Zika virus IgG antibodies. Briefly, a 384-well microplate (Corning(r), New York, USA), was coated with antigen obtained either from cell culture or ultracentrifuged virions at 5 µg /mL (30 µL) per well using ELISA plates coated with sodium carbonate buffer (Na2CO3-NaHCO3 0.1 M, pH 9.5); the plate was incubated overnight at 4 °C in a humid chamber. The plate was washed with PBS containing 0.05% Tween 20 (PBS-T). To avoid non-specific binding, the wells were blocked with 60 µL of 5% skim milk (Molico(r), Nestlé, Vevey, Switzerland) in PBS-T for 1 h at room temperature. Diluted serum samples (1:100, 30 µL) were added to the wells and incubated for 1 h at 37 °C. The microplate was washed four times with PBS-T, and 30 µL of an anti-human IgG peroxidase conjugate (DAKO A/S, Denmark(r)) at 1: 5,000 in 1% skim milk and PBS-T was added per well and incubated for 1 h at 37 °C. The microplate was again washed four times with PBS-T, and 30 µL TMB substrate solution (3,3',5,5'-tetramethylbenzidine, Dade Behring(r)) were added to each well. The microplate was incubated in the dark at room temperature for 15 min, and the reaction was stopped. The optical density at 450 nm was determined using a spectrofluorometer Multi-mode FilterMax F5 Microplate Reader (Molecular Devices(r), California, USA). To perform a high-avidity IgG immunoassay, an additional incubation step with 6 M urea (pH 7.4), for 15 minutes at 37 ºC (PBS-T in the control well) was performed after the serum samples incubation step; this was followed by washing with PBS-T and subsequent conjugate and chromogen incubations, as previously described.
Statistical analyses
The cut-off threshold (99%) was determined using the mean plus three standard deviations obtained from 53 serum samples that were negative for dengue virus. The receiver operating characteristic curve (ROC) analysis and serological indices (specificity, sensitivity, positive and negative predictive values and likelihood ratio) were performed using the GraphPad Prism 5.0 software (GraphPad Software, La Jolla California, USA) and RNA-positive serum samples. Significance was considered when the probability of equality was less than 5% (p < 0.05), with a power of at least 90%.
RESULTS
ZIKV Brazilian strain, electron microscopy and Western blotting
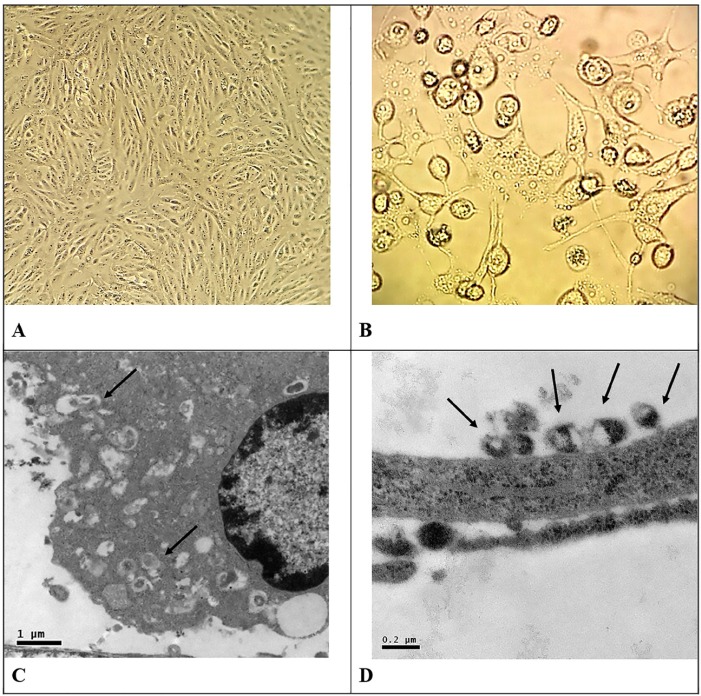
The Brazilian ZIKV strain was inoculated into monolayers of Vero cells in 150 mL plastic flasks, and cell infection was monitored daily under an inverted Leica microscope at 320 x magnification. Figure 1 shows the morphology of the Vero cell monolayer; Figure 1A shows Vero cell monolayers not infected with ZIKV, and Figure 1B shows the cytopathic effect of ZIKV in Vero cells. The electron microscopy shows that infected cells presented a large number of active ribosomes in the cytoplasm and viral particles budding (Fig. 1C). Complete virions with viral nucleic acid surrounded by a lipid envelop were also observed near the cells (Fig. 1D).
Fig. 1. Morphological aspects of Vero cells infected with Zika virus (ZKV). A- Vero cell monolayers not infected with ZKV (A). At four days after ZKV infection, showing the cytopathic effect of ZKV in Vero cells. (B). ZKV-infected Vero cell showing biosynthetic activity in the cytoplasm and virus particle processing (arrows in C). ZKV viral particles outside the monolayer (arrows in D).
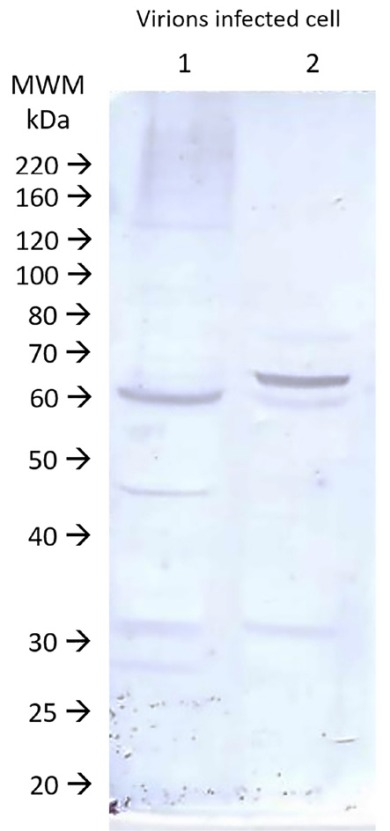
Viral antigens purified as described in Methods were subjected to SDS-PAGE and transferred to nitrocellulose membranes. Membranes containing antigens prepared by two methods reacted with serum samples from a convalescent patient, from a RNA-positive patient obtained at 21 days after the molecular diagnosis, as revealed using an anti-human IgG peroxidase conjugate and 4-chloronaphthol. The digitalized membrane is shown in Figure 2. Only a few bands in both antigen preparations were recognized by the convalescent serum, the most important of which is a band of 60 kDa corresponding to the viral capsid protein. Note that there is a small difference in molecular weight between the two fractions, most likely due to protein processing after particle formation.
Fig. 2. Protein patterns on immunoblots after SDS-PAGE of the two antigen fractions used in ELISA. Lane 1- Protein antigen of the cell layer. Lane 2- Protein antigen of ultracentrifuged virions. MWM: markers with known molecular weights. Digital imaging.
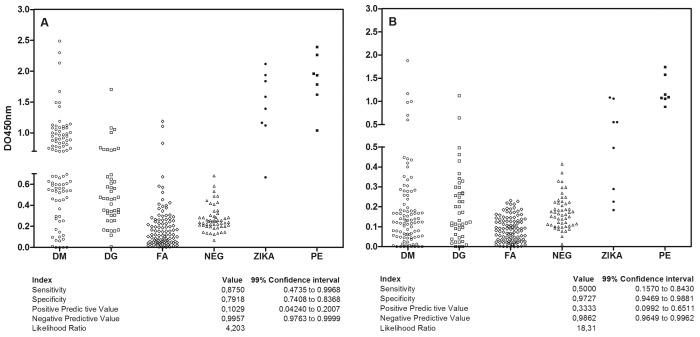
Detection of anti-Zika virus IgG using protein antigens from the cell layer in a classic solid-phase immunosorbent assay (IgG ELISA ZV) and after washing with 6 M urea (IgG ELISA ZVU)
We assayed 308 serum samples from positive and negative patients for dengue virus, yellow fever and Zika virus by classical ELISA, as well as ELISA followed by urea 6 M washing and compared the results to a reference panel of 8 serum samples from patients RT-PCR-positive for Zika virus. The reactivity of serum samples in the anti-Zika virus IgG detection by classical ELISA is shown in Figure 3A and by ELISA with 6 M urea is shown in Figure 3B. The cut-off value was established using 53 negative samples, as described in Methods, for each of the assays. Of the 95 serum samples obtained from dengue virus IgM-positive patients, 48 samples were positive by IgG ELISA ZV and 6 by IgG ELISA ZVU. Of the 45 serum samples obtained from dengue virus IgG-positive patients, 10 were positive by IgG ELISA ZV and two by IgG ELISA ZVU. Among the 100 serum samples obtained from yellow fever-vaccinated patients, three were positive by IgG ELISA ZV, and all samples were in concordance with the IgG ELISA ZVU reference test. Of the eight serum samples obtained from patients positive for Zika virus by RT-PCR, seven samples were positive by IgG ELISA ZV and four by IgG ELISA ZVU. Regarding the seven serum samples from mothers and neonates, all of them were positive by both IgG ELISA ZV and IgG ELISA ZVU. A high frequency of discordant results compared with the reference test was found for IgG ELISA ZV, though only one sample positive for Zika virus by RT-PCR was not identified by the classical ELISA, resulting in 87% sensitivity. The IgG ELISA ZVU showed lower sensitivity but high specificity of 97%, most likely due to the elimination of cross-reacting antibodies.
Fig. 3. Quantitative reactivity of ELISA for anti-Zika virus IgG (A) and high-avidity anti-Zika virus IgG (B) in serum using protein antigen from the cell layer in solid support. Sample origin: DM: acute dengue virus infection with positive IgM serology. DG: dengue virus infection with positive IgG serology. YF: Yellow fever vaccination patients. N: negative controls from patients without history of travel to endemic areas. ZKVR: Acute patients with Zika virus with previous RNA detection. MM: Samples after delivery from mothers with microcephaly babies. Interruptions in the axes show the cut-off point of the assays. Serological indices are presented in the table below using RT-PCR-positive patient serum as a reference test.
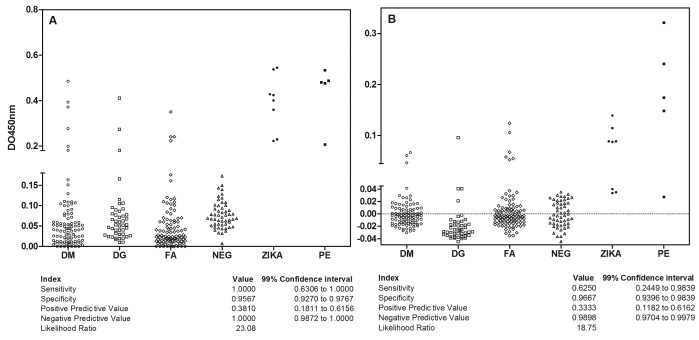
Detection of anti-Zika virus IgG using antigens from virions prepared by ultracentrifugation in a classical solid-phase immunosorbent assay (IgG ELISA ZVV) and after washing with 6 M urea (IgG ELISA ZVVU)
We also tested the samples using protein antigens from virions obtained by ultracentrifugation in solid-support IgG ELISA assays; the results are shown in Figure 4. The cut-off value was established using 53 negative samples, as described in Methods, for each of the assays. Of the 95 serum samples obtained from dengue virus IgM-positive patients, six samples were positive by IgG ELISA ZVV (Fig. 4A) but only three after the 6 M urea washing (Fig. 4B). Regarding the 45 samples from dengue virus IgG-positive patients, three were positive by IgG ELISA ZVV but only one after washing with urea. By IgG ELISA ZVV, four samples from yellow fever-vaccinated patients (100) were positive, which increased to six after urea washing, despite being in the grey zone, i.e., near the cut-off. Among the eight serum samples positive for Zika virus by RT-PCR, all were positive by IgG ELISA ZVV; however, three were below the cut-off after washing with urea, corroborating the low avidity of acute infection antibodies. The serum samples obtained from MM mothers (5) were positive by IgG ELISA ZVV. Virion antigens appear to be more sensitive and specific for detecting IgG anti-ZKV, resulting in 100% sensitivity and 95.6% specificity. Washing with urea resulted in reduced efficiency, which appears more related to the avidity than to the removal of cross-reacting antibodies.
Fig. 4. Quantitative reactivity of ELISA for anti-Zika virus IgG (A) and high-avidity anti-Zika virus IgG (B) in serum using protein antigen from ultracentrifuged virions in solid support. Sample origin: DM: acute dengue virus infection with positive IgM serology. DG: dengue virus infection with positive IgG serology. YF: Yellow fever vaccination patients. N: negative controls from patients without history of travel to endemic areas. ZKVR: Acute patients with Zika virus with previous RNA detection. MM: Samples after delivery from mothers with microcephaly babies. Interruptions in the axes show the cut-off point of the assays. Serological indices are presented in the table below using RT-PCR-positive patient serum as a reference test.
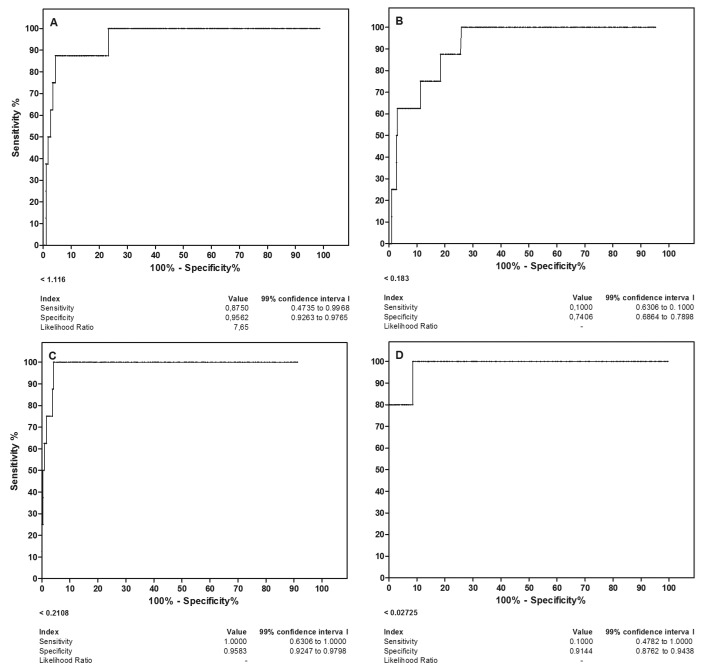
ROC curves and serological indices for ZIKV IgG ELISA assays
We performed a ROC analysis for all the IgG ELISA results, as shown in Figure 5 (assays are described in the legend). Four analyses were performed using samples from ZKV infections confirmed by RNA (as positive samples); all of the other Flavivirus models and negative samples were combined as the control pool. The best discriminating assay was achieved using the ultracentrifuged virion antigen-based assay: ELISA ZVV (Fig. 5C), without urea washing, which allowed for both high sensitivity and specificity. Although good serological indices were obtained for ELISA using the ZKV protein antigen obtained from the cell layer (Fig. 5A), this approach requires urea washing to remove cross-reacting antibodies from other flaviviral infections.
Fig. 5. ROC analysis for discrimination of anti-Zika virus IgG antibodies using protein antigen from the cell layer (A and B) or ultracentrifuged virions (C and D) in the IgG ELISA assay (A, C) and IgG avidity ELISA assay (B and D).
DISCUSSION
The model applied to produce antigen for Zika virus IgG detection showed that infection of Vero cells is a good approach for producing antigens for ZKV ELISA, both with regard to antigens obtained using the infected cell layer or virions concentrated from the supernatant. Our ultrastructural data are compatible with the few data available for ultrastructure of Zika virus in the literature 19 with typical cytoplasm involvement and virion production by infected cells. In addition, infective particles present in the medium can be concentrated by ultracentrifugation. This is a typical method of producing antigens for ELISA assays for viral infections, such as measles 16 or rubella 17 , and the low cytopathic effect of this RNA+ virus results in a higher production of viral particles that can be recovered from the medium. By examining the viral protein pattern recognized by confirmed-infection of a convalescent serum sample, we found slight differences in the molecular weight of the major ~60 kDa ZKV protein, which was smaller in the cell layer. This suggests that exit from the cell may be mediated via peptide removal, which is similar to the antigen presented to immune cells, resulting in a more specific IgG detection by ELISA. Our data using infected cell layer antigen may be due to non-structural proteins or unprocessed antigens, which are absent in virions 19 . In similar antigen preparations, such as for the preparation of measles antigen, the high cytopathic effect resulted in the accumulation of antigen in altered cells, which resulted in a high yield of specific viral protein in these models 16 . Nonetheless, similar proteins in other flaviviral infections could induce cross-reacting antigens production, resulting in a low efficiency of this antigen in our assays. Most viral antigens are currently produced by recombinant technology, both due to biosafety concerns or to obtain a higher yield of pure proteins. However, this is a complex problem in flaviviruses, which share similar cytoplasm processing and common sequences, resulting in similar proteins in several of these viruses. Although these similarities can result in cross-reactive antibodies production 20 , we cannot exclude the possibility of double infections in these cases, especially when there is no decay of the reaction after washing with urea.
We used classical approaches to obtain an antigen for ZKV and achieved good results using virions concentrated from the medium by ultracentrifugation. The antigen was readily recognized by a convalescent serum sample after SDS-PAGE and blotting, and it has also exhibited antigen processing after exiting the cell. The yield of the best preparation was relatively low, but five 75 cm2 culture flasks after four days of infection provided 100 mL of medium and approximately 1 mg of protein antigen after ultracentrifugation, resulting in an amount of protein sufficient for adsorbing 50 microplates or 4,500 assays. In contrast, antigen prepared from the cell layer yielded five times more protein (data not shown).
The aim of our study was to start the development of a useful serological approach for diagnosing congenital infections detected at birth. In general, the model of congenital infections is an association of a benign maternal infection with a few or uncharacteristic symptoms and the possibility of a severely compromised fetus, with a small proportion of infected neonates presenting with central nervous system impairment, as in toxoplasmosis 21 or rubella 22 . Zika virus infection causes a similar congenital infection; the acute clinical disease is short with few characteristic symptoms common to several other congenital diseases, and fetal involvement is also diagnosed very late. Neuronal death is difficult to evaluate in fetuses, requiring invasive sampling procedures and analysis of amniotic fluid samples by molecular techniques 11 . The abundant growth of the virus in the host results in effective immune clearing of the infection within few days, despite viral survival in some biological samples such as male semen 23 . However, blood clearance of the virus suggests the difficulty of diagnosis after a few days of acute infection and certainly at the end of pregnancy 11 .
We conclude that detection of ZKV IgG by ELISA constitute an adequate diagnostic tool due to its high sensitivity. The approach also allows to cople the ELISA technique to avidity assays, which could help to establish the timing of acute infection, similar to other congenital infections 24 . Our small sample precludes further analysis of the use of IgG avidity assays in ZKV infections. Nonetheless, some convalescent serum samples from RNA-confirmed patients become negative after urea washing, which is a promising result.
Our preliminary data show the usefulness of natural ZKV antigens in IgG ELISA assays, if adequate purification procedures are performed. These assays will help in the differential diagnosis of neonatal cases with microcephaly within the context of congenital infectious diseases.
ACKNOWLEDGEMENTS
We gratefully thank the expertise of Cleusa F. Takakura for the electron microscopy studies and Monica de Paula Leal for the Western blot studies. Claudio S. Pannuti and E.A. Luna provided the dengue serology samples. We thank the Pedro Vasconcelos group (I.E. Chagas) for isolating and E. Durigon (ICB/USP) for supplying the Brazilian ZVV strain. This work has no special funding but was indirectly supported by several grants to the senior authors (CMM and HFA Jr.) and LIMHCFMUSP.
REFERENCES
- 1.Racicot K, Kwon JY, Aldo P, Silasi M, Mor G. Understanding the complexity of the immune system during pregnancy. Am J Reprod Immunol. 2014;72:107–116. doi: 10.1111/aji.12289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Burt TD. Fetal regulatory T cells and peripheral immune tolerance in utero: implications for development and disease. Am J Reprod Immunol. 2013;69:346–358. doi: 10.1111/aji.12083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Gerber S, Hohlfeld P. Screening for infectious diseases. Childs Nerv Syst. 2003;19:429–432. doi: 10.1007/s00381-003-0766-5. [DOI] [PubMed] [Google Scholar]
- 4.Pomares C, Montoya JG. Laboratory diagnosis of congenital toxoplasmosis. J Clin Microbiol. In Press. 2016 doi: 10.1128/JCM.00487-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Neu N, Duchon J, Zachariah P. TORCH infections. Clin Perinatol. 2015;42:77–103. doi: 10.1016/j.clp.2014.11.001. [DOI] [PubMed] [Google Scholar]
- 6.Villard O, Cimon B, L'Ollivier C, Fricker-Hidalgo H, Godineau N, Houze S, et al. Serological diagnosis of Toxoplasma gondii infection: recommendations from the French National Reference Center for Toxoplasmosis. Diagn Microbiol Infect Dis. 2016;84:22–33. doi: 10.1016/j.diagmicrobio.2015.09.009. [DOI] [PubMed] [Google Scholar]
- 7.Sensini A. Toxoplasma gondii infection in pregnancy: opportunities and pitfalls of serological diagnosis. Clin Microbiol Infect. 2006;12:504–512. doi: 10.1111/j.1469-0691.2006.01444.x. [DOI] [PubMed] [Google Scholar]
- 8.Campos GS, Bandeira AC, Sardi SI. Zika virus outbreak, Bahia, Brazil. Emerg Infect Dis. 2015;21:1885–1886. doi: 10.3201/eid2110.150847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Faria NR, Azevedo RS, Kraemer MU, Souza R, Cunha MS, Hill SC, et al. Zika virus in the Americas: early epidemiological and genetic findings. Science. 2016;352:345–349. doi: 10.1126/science.aaf5036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Schuler-Faccini L, Ribeiro EM, Feitosa IM, Horovitz DD, Cavalcanti DP, Pessoa A, et al. Possible association between Zika virus infection and microcephaly: Brazil, 2015. MMWR Morb Mortal Wkly Rep. 2016;65:59–62. doi: 10.15585/mmwr.mm6503e2. [DOI] [PubMed] [Google Scholar]
- 11.Calvet G, Aguiar RS, Melo AS, Sampaio SA, de Filippis I, Fabri A, et al. Detection and sequencing of Zika virus from amniotic fluid of fetuses with microcephaly in Brazil: a case study. Lancet Infect Dis. 2016;16:653–660. doi: 10.1016/S1473-3099(16)00095-5. [DOI] [PubMed] [Google Scholar]
- 12.Mlakar J, Korva M, Tul N, Popović M, Poljšak-Prijatelj M, Mraz J, et al. Zika virus associated with microcephaly. N Engl J Med. 2016;374:951–958. doi: 10.1056/NEJMoa1600651. [DOI] [PubMed] [Google Scholar]
- 13.Cordeiro MT, Pena LJ, Brito CA, Gil LH, Marques ET. Positive IgM for Zika virus in the cerebrospinal fluid of 30 neonates with microcephaly in Brazil. Lancet. 2016;387(10030):1811–1812. doi: 10.1016/S0140-6736(16)30253-7. [DOI] [PubMed] [Google Scholar]
- 14.Cugola FR, Fernandes IR, Russo FB, Freitas BC, Dias JL, Guimarães KP, et al. The Brazilian Zika virus strain causes birth defects in experimental models. Nature. 2016;534:267–271. doi: 10.1038/nature18296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Beck C, Jimenez-Clavero MA, Leblond A, Durand B, Nowotny N, Leparc-Goffart I, et al. Flaviviruses in Europe: complex circulation patterns and their consequences for the diagnosis and control of West Nile disease. Int J Environ Res Public Health. 2013;10:6049–6083. doi: 10.3390/ijerph10116049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Souza VA, Pannuti CS, Sumita LM, Andrade HF., Júnior Enzyme-linked immunosorbent assay-IgG antibody avidity test for single sample serologic evaluation of measles vaccines. J Med Virol. 1997;52:275–279. doi: 10.1002/(sici)1096-9071(199707)52:3<275::aid-jmv7>3.0.co;2-#. [DOI] [PubMed] [Google Scholar]
- 17.Souza VA, Moraes JC, Sumita LM, Camargo MC, Fink MC, Hidalgo NT, et al. Prevalence of rubella antibodies in a non-immunized urban population, São Paulo, Brazil. The Division of Immunization, CVE. Rev Inst Med Trop Sao Paulo. 1994;36:373–376. doi: 10.1590/s0036-46651994000400011. [DOI] [PubMed] [Google Scholar]
- 18.Towbin H, Staehelin T, Gordon J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc Natl Acad Sci USA. 1979;76:4350–4354. doi: 10.1073/pnas.76.9.4350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hamel R, Dejarnac O, Wichit S, Ekchariyawat P, Neyret A, Luplertlop N, et al. Biology of Zika virus infection in human skin cells. J Virol. 2015;89:8880–8896. doi: 10.1128/JVI.00354-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Gyurech D, Schilling J, Schmidt-Chanasit J, Cassinotti P, Kaeppeli F, Dobec M. False positive dengue NS1 antigen test in a traveller with an acute Zika virus infection imported into. Switzerland. Swiss Med Wkly. 2016;146:w14296–w14296. doi: 10.4414/smw.2016.14296. [DOI] [PubMed] [Google Scholar]
- 21.Daffos F, Forestier F, Capella-Pavlovsky M, Thulliez P, Aufrant C, Valenti D, et al. Prenatal management of 746 pregnancies at risk for congenital toxoplasmosis. N Engl J Med. 1988;318:271–275. doi: 10.1056/NEJM198802043180502. [DOI] [PubMed] [Google Scholar]
- 22.Andrade JQ, Bunduki V, Curti SP, Figueiredo CA, de Oliveira MI, Zugaib M. Rubella in pregnancy: intrauterine transmission and perinatal outcome during a Brazilian epidemic. J Clin Virol. 2006;35:285–291. doi: 10.1016/j.jcv.2005.09.007. [DOI] [PubMed] [Google Scholar]
- 23.Musso D, Roche C, Robin E, Nhan T, Teissier A, Cao-Lormeau VM. Potential sexual transmission of Zika virus. Emerg Infect Dis. 2015;21:359–361. doi: 10.3201/eid2102.141363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Prince HE, Lapé-Nixon M. Role of cytomegalovirus (CMV) IgG avidity testing in diagnosing primary CMV infection during pregnancy. Clin Vaccine Immunol. 2014;21:1377–1384. doi: 10.1128/CVI.00487-14. [DOI] [PMC free article] [PubMed] [Google Scholar]