

Summary
Sox6 belongs to the Sox gene family and plays a pivotal role in fiber type differentiation, suppressing transcription of slow-fiber-specific genes during fetal development. Here, we show that Sox6 plays opposite roles in MyHC-I regulation, acting as a positive and negative regulator of MyHC-I expression during embryonic and fetal myogenesis, respectively. During embryonic myogenesis, Sox6 positively regulates MyHC-I via transcriptional activation of Mef2C, whereas during fetal myogenesis, Sox6 requires and cooperates with the transcription factor Nfix in repressing MyHC-I expression. Mechanistically, Nfix is necessary for Sox6 binding to the MyHC-I promoter and thus for Sox6 repressive function, revealing a key role for Nfix in driving Sox6 activity. This feature is evolutionarily conserved, since the orthologs Nfixa and Sox6 contribute to repression of the slow-twitch phenotype in zebrafish embryos. These data demonstrate functional cooperation between Sox6 and Nfix in regulating MyHC-I expression during prenatal muscle development.
Keywords: Sox6, Nfix, MyHC-I, myogenesis
Graphical Abstract
Highlights
-
•
Sox6 has opposite roles in MyHC-I regulation during embryonic and fetal myogenesis
-
•
In embryonic muscle, Sox6 enhances MyHC-I expression via regulation of Mef2C
-
•
In fetal muscle, Nfix is required for Sox6-mediated repression of MyHC-I
-
•
The Sox6 and Nfixa orthologs cooperate in repressing smyhc1 in zebrafish
Taglietti et al. reveal molecular mechanisms defining muscle fiber specification, driven by the key transcription factors Nfix and Sox6, during mouse and zebrafish development. They show that Nfix reverses Sox6 function between embryonic and fetal phases of myogenesis, allowing the proper expression of slow MyHC.
Introduction
In vertebrates, the process of skeletal muscle development occurs in subsequent steps that involve distinct populations of myogenic progenitors, the myoblasts, which arise from the dermomyotomal domain of somitic mesoderm (Christ and Ordahl, 1995). The process of myogenic differentiation is initiated in mesodermal cells by a family of basic-helix-loop-helix (bHLH) transcription factors, named muscle regulatory factors (MRFs), that are able to activate transcription of muscle-specific markers such as the myosin heavy chain (MyHC) isoforms (Pinney et al., 1988, Cao et al., 2010). Embryonic myoblasts sustain a first wave of myogenesis between embryonic day 10 (E10) and E12 in the mouse and give rise to primary myofibers that establish the primitive shape of muscle and express high levels of the slow MyHC isoform (MyHC-I, encoded by the Myh7 gene) and of the embryonic MyHC isoform (MyHC-emb, encoded by Myh3) (Schiaffino et al., 1986, Stockdale, 1992). A second wave of muscle differentiation takes place between E15 and E18, driven by fetal myoblasts that form secondary fibers, characterized by low levels of MyHC-I and high levels of neonatal MyHC (MyHC-neo, encoded by Myh8) (Eusebi et al., 1986, Lyons et al., 1990, Daou et al., 2013). Eventually, primary fibers conserve the slow-twitch phenotype typical of embryonic muscle, while secondary fibers lose the expression of several embryonic-specific markers such as MyHC-I and acquire expression of fast-twitch markers (Ferrari et al., 1997, Biressi et al., 2007). Embryonic and fetal myoblasts, once isolated from the embryo, are committed to a specific fiber type, suggesting the involvement of intrinsic factors rather than nerve activity in the establishment of fiber phenotype (Page et al., 1992). These observations suggest that the proper transition of skeletal muscle from the embryonic to the fetal/post-natal phenotype requires a switch in the transcriptional status of differentiating myoblasts.
In our previous study, we have shown that the transcription factor Nfix, a member of the nuclear factor I (Nfi) family, has a key role in the establishment of fetal muscle phenotype and in the downregulation of slow MyHC both in fetal and adult muscles (Messina et al., 2010, Rossi et al., 2016). We have shown that Nfix is strongly expressed in fetal myoblasts and indirectly represses MyHC-I expression via the transcription factor Nfatc4, a positive regulator of MyHC-I in skeletal muscle (Calabria et al., 2009, Messina et al., 2010). We have also reported that the zebrafish (Danio rerio) Nfix ortholog Nfixa has an evolutionarily conserved role in the transition from slow-twitch to fast-twitch myogenesis (Pistocchi et al., 2013). In the past few years, it has been shown that Sox6, a member of the Sry-related HMG box (Sox) factor family, which is highly conserved in vertebrates, plays a critical role in fetal fiber specification through direct repression of MyHC-I by binding to the 5′-upstream region in two different binding sites. The first is located −200 bp from the transcription start site (TSS) in the proximal promoter and is sufficient for Sox6-dependent MyHC-I repression in fetal myotubes (Hagiwara et al., 2007, An et al., 2011), and the second is located −2,900 bp from the TSS in a distal muscle enhancer that is required for full promoter activity (Giger et al., 2000, Blow et al., 2010). As a consequence, Sox6-null mouse muscle displays increased levels of MyHC-I and a general switch toward a slower phenotype (Hagiwara et al., 2007, An et al., 2011, Quiat et al., 2011). Of note, studies in zebrafish have shown that Sox6 is restricted to fast-twitch fibers during embryonic muscle development and that ectopic Sox6 expression in adaxial cells (the slow muscle progenitors in zebrafish) leads to the silencing of slow-twitch genes (such as the slow MyHC isoform smyhc1 and the transcription factor prox1a) (von Hofsten et al., 2008, Wang et al., 2011, Jackson et al., 2015).
Here, we observed that Sox6, at variance with Nfix, is expressed at comparable levels in embryonic and fetal myoblasts, despite its role of inhibitor of slow twitching program. Intriguingly, we demonstrated that Sox6 has opposite roles in regulating MyHC-I expression between embryonic and fetal myogenesis in mouse. Specifically, during embryonic myogenesis, Sox6 indirectly promotes MyHC-I expression via transcriptional activation of Mef2C. As a consequence, Sox6 deficiency in embryonic muscle leads to a strong downregulation of MyHC-I. On the contrary, during fetal myogenesis, Sox6 cooperates with Nfix to repress MyHC-I in a complex in which Nfix is necessary for the proper binding of Sox6 to the MyHC-I promoter in fetal myotubes. Finally, we show that Nfixa and Sox6 together regulate sMyHC in zebrafish embryos, revealing an evolutionarily conserved mechanism that is required for the acquisition of normal muscle phenotype.
Results
Sox6 Transcriptionally Promotes MyHC-I Expression during Embryonic Myogenesis
Sox6 has been intensively studied as an inhibitor of slow muscle phenotype during the fetal period. However, we observed that Sox6 is also expressed during embryonic myogenesis, which is mainly characterized by the expression of typical slow genes such as the slow MyHC isoform MyHC-I. Myf5GFP-P/+ embryos and fetuses were collected at E12.5 or E16.5, and GFP-positive myoblasts were isolated via fluorescence-activated cell sorting (FACS) as previously described (Messina et al., 2010). Using quantitative real-time PCR and western blot, we found that Sox6 levels do not significantly change, whereas Nfix, as known, is drastically upregulated in fetal progenitors (Figures 1A–1C). Importantly, these data show that Sox6 and Nfix proteins are co-expressed only during fetal myogenesis. We also performed extensive immunofluorescence analysis on frozen mouse embryo sections from E10.5 to E18.5 in order to follow Sox6 protein expression throughout development. Notably, Sox6 is first expressed between E11.5 and E12.5 in primary fibers that express high levels of MyHC-I (Figures S1A–S1H), whereas at E17.5, the localization of Sox6 is almost completely associated with secondary fibers that are negative for MyHC-I (Figures S1I–S1L), as previously described (An et al., 2011, Richard et al., 2011). We therefore decided to investigate the possible function of Sox6 during embryonic myogenesis, a function not apparently linked to repression of MyHC-I. To this aim, we performed immunofluorescence analysis on E12.5 muscle sections from homozygous mice carrying the Sox6lacZ allele (hence referred to as Sox6-null mice) (Smits et al., 2001). Surprisingly, the staining for MyHC-I is strongly decreased in Sox6 null in comparison to wild-type (WT) muscle (Figures 1D and 1E). Importantly, no differences in total MyHC content were assessed in embryonic muscle groups of Sox6-null embryos (Figures 1G and 1H), implying that the decrease in MyHC-I expression is not due to delayed or aberrant muscle differentiation. We also performed western blot and quantitative real-time PCR on embryonic muscle lysates to confirm the immunofluorescence data, and our results showed a decrease of MyHC-I protein in Sox6-null samples, without major changes in total MyHC content (Figures 1J and 1K). Interestingly, the phenotype of Sox6-null embryonic muscle is reminiscent of the Tg:Mlc1f-Nfix2 gain-of-function embryo (Figures 1F and 1I), in which the Nfix2 splice variant is ectopically expressed in muscle cells from E11.5, leading to a fetal-like muscle phenotype (Kelly et al., 1997, Messina et al., 2010). Importantly, Nfix expression in embryonic skeletal muscle is not altered in the absence of Sox6 at both the protein and mRNA levels (Figures S2A–S2D). Taken together, these results show that Sox6 is expressed at equal levels in skeletal muscle during embryonic and fetal myogenesis and that deletion of Sox6 during the embryonic period unexpectedly leads to downregulation of MyHC-I.
Figure 1.

Sox6 Acts a Positive Regulator of the Slow-Twitch Phenotype during Embryonic Myogenesis
(A) Quantitative real-time PCR analysis on freshly isolated Myf5GFP-P/+ embryonic (E12.5) and fetal (E16.5) myoblasts showing relative expression of Sox6 and Nfix transcripts in the two populations.
(B) Western blot on lysates from freshly isolated Myf5GFP-P/+ embryonic and fetal myoblasts. β-Tubulin was used to normalize the amount of protein loaded.
(C) Quantitative densitometry of the protein expression levels of Sox6 and Nfix at E12.5 and E16.5.
(D–I) Immunofluorescence on E12.5 muscle sections from WT (D and G), Sox6 null (E and H), and Tg:Mlc1f-Nfix2 (F and I) mice stained with anti-MyHC-I (D–F) or anti-MyHCs (MF20) antibodies (G-I). Dashed lines highlight the forelimb anlagen contour. Scale bars, 100 μm.
(J) Western blot on E12.5 muscle samples from WT, Sox6 null, and Tg:Mlc1f-Nfix2 mice. Vinculin was used to normalize the amount of loaded proteins.
(K) Quantitative real-time PCR on E12.5 muscle tissue from WT and Sox6-null mice (∗p < 0.05; ∗∗p < 0.01; n = 3).
Sox6 Positively Regulates the Slow-Twitch Phenotype of Embryonic Myoblasts by Binding to Mef2C Promoter
In order to define a possible mechanism by which Sox6 regulates the transcription of MyHC-I during embryonic myogenesis, we performed chromatin immunoprecipitation (ChIP) for Sox6 in differentiated embryonic myoblasts. We found that Sox6 does not significantly bind either to the proximal or to the distal regulative regions upstream of MyHC-I (Figure 2A), thus suggesting that Sox6 is not able to directly regulate MyHC-I transcription in embryonic muscle. To confirm this hypothesis, we performed luciferase assays on WT and Sox6-null embryonic differentiated myoblasts with vectors containing the 3,500-bp MyHC-I full 5′-upstream region (MyHC-I 3500), the 408-bp MyHC-I proximal promoter sequence (MyHC-I 408), or the mutated forms of the distal and proximal canonical Sox6 binding sites, MyHC-I 3500 m and MyHC-I 408 m (Figure S3A) (Hagiwara et al., 2007, An et al., 2011). As expected, in the absence of Sox6, we found a significant reduction of firefly luciferase activity in all the conditions with the only notable exception of the 408 WT construct (Figure 2B), suggesting that Sox6 is promoting MyHC-I expression in embryonic myocytes without direct binding to its canonical binding sites. In order to identify a possible indirect mechanism by which Sox6 enhances MyHC-I expression in embryonic muscle, we focused on the transcription factor Mef2C, a known positive regulator of the slow phenotype (Wu et al., 2000, Potthoff et al., 2007, Anderson et al., 2015). To verify the interaction between Mef2C and the MyHC-I promoter in embryonic myoblasts, we performed a ChIP assay, which showed direct binding of Mef2C on the proximal MyHC-I promoter (Figure S3B). Interestingly, Mef2C mRNA is downregulated in E12.5 Sox6-null muscle, at variance with the closely related Mef2A (Figure 2C). By ChIP on embryonic differentiated myoblasts, we found that Sox6 directly binds to a region located in the Mef2C promoter (Figure 2D), indicating that Sox6 is a direct activator of Mef2C during embryonic myogenesis. In order to validate a possible Mef2C-mediated mechanism of MyHC-I regulation, we transfected WT and Sox6-null unpurified embryonic myoblasts with a vector overexpressing Mef2C. Mef2C overexpression in Sox6-deficient cells (Figure S3C) leads to an increase of MyHC-I expression when normalized on the levels of MyHC-emb to account for the total number of myogenic cells (Figure 2E). To validate these data, we also transduced purified embryonic myoblasts with a lentiviral vector expressing small hairpin RNA (shRNA) against Sox6 to achieve in vitro Sox6 knockdown (Figure S3D). We then transfected cells with Mef2C-overexpression vector (Figure S3D) and assessed the levels of MyHC-I mRNA by quantitative real-time PCR (Figure 2F). Strikingly, overexpression of Mef2C was able to partially rescue MyHC-I expression in shSox6 cells (up to 30% of the levels of scramble-transduced cells). These results show that during the embryonic period, Sox6 acts indirectly as a positive regulator of MyHC-I via a Mef2C-dependent mechanism, in sharp contrast with its well-known and characterized function during fetal myogenesis (An et al., 2011, Quiat et al., 2011; see below).
Figure 2.

Sox6 Indirectly Activates MyHC-I Expression in Embryonic Myoblasts via a Mef2C-Dependent Mechanism
(A) ChIP assay with anti-Sox6 antibody on E12.5 differentiated myoblasts. Three different chromatin regions were tested: a negative control region (intergenic) located 15 kb upstream of MyHC-I TSS, the MyHC-I proximal promoter (−375 bp; MyHC-I promoter), and the MyHC-I distal enhancer (−2,900 bp; MyHC-I enhancer).
(B) Luciferase report assay on WT and Sox6-null E12.5 differentiated myoblasts transfected with control pGL3-basic, MyHC-I 408, MyHC-I 408 m, MyHC-I 3,500, and MyHC-I 3,500 m constructs (∗p < 0.05; ∗∗p < 0.01; n = 2).
(C) Quantitative real-time PCR for Mef2C and Mef2A on E12.5 muscle tissue from WT and Sox6-null mice (∗∗p < 0.01; n = 2).
(D) ChIP assay with anti-Sox6 antibody on E12.5 differentiated primary myoblasts showing binding of Sox6 to a region located −1.1 kb upstream of Mef2C TSS (∗∗∗p < 0.001; n = 3). As negative control, we used the MyHC-I −15 kb intergenic region.
(E) Quantitative real-time PCR for MyHC-I and MyHC-emb (MyHC-I/MyHC-emb ratio) on WT and Sox6-null differentiated embryonic myoblasts transfected with a Mef2C-overexpressing vector (∗p < 0.05; ∗∗p < 0.01; n = 2).
(F) Quantitative real-time PCR for MyHC-I on embryonic myoblasts purified from Myf5GFP-P/+ mice, infected with scramble or shSox6 virus. Myf5GFP-P/+ purified embryonic myoblasts infected by lentiviruses expressing the shRNA for Sox6 were transfected with Mef2C-overespressing vector (∗∗∗p < 0.001; n = 2).
Sox6 Is Necessary for Nfix Binding to the MyHC-I Promoter during the Fetal Period
Since Sox6 has opposite roles in MyHC-I regulation in embryonic and fetal muscle, and since Nfix is only expressed during fetal myogenesis (Messina et al., 2010), we decided to investigate the possible cooperation between Nfix and Sox6 during fetal myogenesis. We first investigated Nfix expression and function in Sox6-null fetuses (E16.5). As previously described in other Sox6-null mouse models (Hagiwara et al., 2007, An et al., 2011, Quiat et al., 2011), fetal fiber specification is completely disrupted in the absence of Sox6. Indeed, in contrast to WT, Sox6-null fetal muscle displays very high levels of MyHC-I by immunofluorescence, quantitative real-time PCR, and western blot (Figures S4A–S4D). We also looked at Nfix on sections from the Sox6-null mice and found that despite the dramatic increase in MyHC-I expression, Nfix is correctly expressed in the nuclei of muscle fibers in the absence of Sox6 (Figures 3A–3D), and western blot analysis did not reveal differences in Nfix protein content (Figure S4C). These data suggest that Nfix is normally expressed in fetal muscles lacking Sox6 but unable to properly repress MyHC-I. To verify whether Sox6 is required for Nfix function, we performed ChIP for Nfix in WT fetal myotubes, which revealed binding of Nfix to MyHC-I promoter in the same region of the proximal Sox6 binding site (−200 bp) (Figure 3E). Interestingly, we observed that in Sox6-null fetal myotubes, the binding of Nfix to the MyHC-I promoter is significantly reduced. Importantly, Nfix binding to Nfatc4 promoter is not impaired in the absence of Sox6 (Figure 3E), suggesting that Nfix is still able to bind to other transcriptional targets in the absence of Sox6. We further validated the repressive role of Nfix on the MyHC-I proximal promoter by performing a luciferase assay with MyHC-I 408 and MyHC-I 408 m vectors in WT and Nfix-null fetal myotubes (Figure 3F). Our results showed increased luciferase expression in the absence of Nfix with the vector carrying the WT sequence, suggesting that indeed Nfix represses MyHC-I by acting on the proximal promoter regulative region, in spite of the absence of any Nfix consensus sequences (data not shown). Importantly, Nfix-dependent negative regulation of MyHC-I is lost when the proximal Sox6 binding sequence is mutated, demonstrating that Sox6 is required for Nfix binding to the proximal MyHC-I promoter (Figure 3F). Overall, these results suggest a possible crosstalk between Nfix and Sox6 in regulating MyHC-I expression at the proximal promoter region.
Figure 3.

Sox6 Is Required for Normal Nfix Function in Fetal Muscle
(A–D) Immunofluorescence with anti-Nfix (green) and anti-MyHC-I (red) antibodies on E16.5 muscle sections from WT (A and B) or Sox6-null (C and D) mice. Arrows indicate Nfix-positive nuclei in secondary (MyHC-I negative) fibers. Arrowheads indicate Nfix-positive nuclei in MyHC-I-positive fibers. Nuclei are counterstained with Hoechst. Scale bars, 25 μm. Higher magnifications of Nfix-positive fibers are shown in the insets.
(E) ChIP assay with anti-Nfix antibody on WT and Sox6-null E16.5 myotubes on negative control region (intergenic) located 15 kb upstream of MyHC-I TSS, the MyHC-I proximal promoter (−375 bp; MyHC-I promoter), and the Nfatc4 promoter region (−1.2 kb upstream of the Nfatc4 TSS) (∗p < 0.05; ∗∗p < 0.01; n = 2).
(F) Luciferase report assay on WT and Nfix-null differentiated fetal myoblasts (E16.5) transfected with control pGL3-basic, MyHC-I 408, and MyHC-I 408 m vectors (∗∗p < 0.01; n = 2).
Nfix Is Necessary and Sufficient for Sox6 Regulation of MyHC-I Expression
To better investigate a possible reciprocal interplay between Nfix and Sox6, we performed immunofluorescence for Sox6 and total MyHC on frozen muscle sections from E16.5 WT and Nfix-null mice (Campbell et al., 2008). We found that Sox6 is expressed in both WT and Nfix-null muscles (Figures S4E and S4F), implying that Nfix is dispensable for normal Sox6 expression in fetal muscle. Interestingly, immunofluorescence for Sox6 and MyHC-I on sections from the same mice revealed that in the absence of Nfix, there is a marked increase in the number of Sox6-positive fibers that co-express MyHC-I, in contrast to WT muscles (Figures 4A–4D). We further validated these data by western blot and quantitative real-time PCR on differentiated fetal myotubes and observed that Nfix-null myotubes express higher levels of MyHC-I than WT cells (Figures 4E and 4F). These data show that in the absence of Nfix, the repressive activity of Sox6 on MyHC-I is partially impaired, even if the normal expression pattern of Sox6 is maintained. The results obtained in both Sox6 and Nfix-null fetuses led us to hypothesize a functional cooperation between Nfix and Sox6 in MyHC-I regulation. Therefore, we tested the ability of Nfix and Sox6 to bind to each other in a multi-protein complex. To this aim, we used fetal myotubes transiently transfected with an Nfix2 hemagglutinin (HA) vector (see Experimental Procedures) and performed a co-immunoprecipitation (coIP) assay from nuclear extracts for HA and Sox6. CoIP revealed the presence of a complex containing both Sox6 and Nfix2-HA, as shown in Figure 5A. Therefore, we wondered whether, as observed for Nfix fetuses in absence of Sox6, Nfix might be required for the binding of Sox6 to the MyHC-I promoter as well. Hence, we performed ChIP for Sox6 in WT and Nfix-null fetal myotubes. Our results showed a decrease in Sox6 binding to the two different sites in the MyHC-I 5′-upstream region (Figure 5B). Notably, the binding with the proximal promoter was completely lost in the absence of Nfix, whereas the binding with the distal enhancer was reduced by 50%. Interestingly, ChIP for Sox6 on the Mef2C promoter reveals that in WT fetal myotubes, Sox6 is not able to bind to the Mef2C promoter, at variance with what happens during the embryonic stage. On the contrary, Sox6 binding on the Mef2C promoter still occurs in Nfix-null fetal myotubes. These data indicate that Nfix is required for the proper binding of Sox6 to the MyHC-I promoter during fetal myogenesis. Finally, we performed ChIP for Sox6 on WT and Tg:Mlc1f-Nfix2 embryonic myoblasts (Figure 5C). Strikingly, Nfix overexpression leads to a switch in the binding properties of Sox6; indeed, we found that Sox6 is bound to the MyHC-I promoter, but not to the Mef2C promoter, in Tg:Mlc1f-Nfix2 embryonic myoblasts. These data demonstrate that Nfix is necessary and sufficient for the binding of Sox6 to the MyHC-I proximal promoter and therefore for Sox6 repressive activity on MyHC-I.
Figure 4.

Nfix Is Necessary for the Correct Function of Sox6 in Fetal Muscle
(A–D) Immunofluorescence with anti-Sox6 (green) and anti-MyHC-I (red) antibodies on fetal (E16.5) muscle sections from WT (A and B) and Nfix-null (C and D) mice. Arrows indicate secondary fibers, which present nuclear Sox6 expression and low or absent staining for MyHC-I. Arrowheads indicate fibers co-expressing nuclear Sox6 and MyHC-I. Nuclei are counterstained with Hoechst. Scale bars, 25 μm. Higher magnifications are shown in the insets.
(E) Western blot on lysates from WT and Nfix-null E16.5 myotubes. β-Tubulin was used to normalize the amount of proteins loaded.
(F) Quantitative real-time PCR on WT and Nfix-null E16.5 myotubes (∗∗p < 0.01; n = 3).
Figure 5.

Nfix Is Required for Fetal-Specific Binding of Sox6 to MyHC-I Regulative Regions
(A) Immunoprecipitation assay, from nuclear extracts, on fetal myoblasts transfected with Nfix2-HA vector, showing the immunoprecipitation of Sox6 (IP Sox6) and HA (IP HA). T, total lysate; IgG, negative control; IP, immunoprecipitated. The coIPs for Sox6 on Sox6-null myotubes and for HA on fetal myoblasts transfected with the HA-only expressing vector (pCH-HA) were used as negative controls.
(B) ChIP assay with anti-Sox6 antibody on WT and Nfix-null E16.5 myotubes on the same chromatin regions described in Figures 2A and 2D (∗p < 0.05; ∗∗p < 0.01; ∗∗∗p < 0.001; n = 2). As a negative control, we used only the MyHC-I −15 kb region.
(C) ChIP assay on unpurified embryonic myoblasts from WT or Tg:Mlc1f-Nfix2 to test Sox6 binding on MyHC-I and the Mef2C promoter. The intergenic region was used as a negative control and IgG as an unrelated antibody (∗∗p < 0.01; ∗∗∗p < 0.001; n = 2).
Functional Cooperation of Sox6 and Nfixa Is Evolutionarily Conserved in Zebrafish
It was previously demonstrated that both Sox6 and Nfixa have an evolutionarily conserved role in the repression of slow-twitch genes in zebrafish (von Hofsten et al., 2008, Pistocchi et al., 2013, Jackson et al., 2015). We thus wondered whether a functional interplay between Nfixa and Sox6 is conserved in zebrafish myogenesis. As a preliminary analysis, we performed quantitative real-time PCR on trunk and tail regions isolated from zebrafish embryos at 1, 2, and 3 days post-fertilization (dpf) and found that the sox6 transcript is expressed at high levels at 1 dpf and is steadily downregulated up to 3 dpf (Figure S5A), whereas the nfixa transcript peaks at 2 dpf (Figure S5B), as shown previously (Pistocchi et al., 2013). Additionally, we performed immunofluorescence for Sox6 and total MyHC (MF20 antibody) or the slow MyHC isoform sMyHC (F59 antibody) on 2 dpf embryos and found that Sox6 protein is specifically expressed in the nuclei of fast muscle fibers that are negative for sMyHC (Figures 6A and 6B), whereas the outer superficial fibers positive for sMyHC are mostly negative for Sox6 (data not shown). Interestingly, we observed that a minority of slow superficial fibers show cytoplasmic staining for Sox6 (Figures 6C–6J), suggesting that Sox6 subcellular localization might be spatially regulated in the different muscle domains. In order to elucidate the role of Sox6 in slow-twitch genes regulation, we performed morpholino (MO)-mediated knockdown of sox6 (von Hofsten et al., 2008). By quantitative real-time PCR analysis (Figures S5C and S5D), we found that the expression of smyhc1 is markedly increased in sox6 morphants at 2 dpf, whereas the fast-twitch gene mylpfa (fast myosin light chain isoform) is expressed at equal levels. Moreover, the nfixa transcript is drastically upregulated in sox6 morphants, suggesting that Sox6 might negatively regulate nfixa (Figure S5E). We conclude that Sox6 is a critical repressor of the slow-twitch phenotype in zebrafish and that Nfixa is not able to compensate for sox6 knockdown. To verify a possible cooperation between Nfixa and Sox6, we performed co-injections of morpholinos against sox6 and nfixa at lower doses with respect to those previously described (von Hofsten et al., 2008, Pistocchi et al., 2013) in order to minimize their effect when injected separately (see Experimental Procedures). Co-injection of these doses of MOs resulted in synergistic defects in motility in touch-response assays (Figure S6). Control embryos and the vast majority of sox6 or nfixa morphants readily swam away when touch-stimulated. On the contrary, 66% of double partial morphants were either shivering or bending their tails before eventually moving away but within a shorter distance (Figures S6A and S6C; Movie S1). The synergistic effect is more evident when lowering the doses (see Figures S6B and S6D). Strikingly, quantitative real-time PCR results show that smyhc1 is significantly upregulated only in the double morphants at 48 hpf, whereas the level of myl1, an early marker for differentiating fast muscle cells (Burguière et al., 2011), does not change (Figure 6K). Moreover, we validated the increased expression of sMyHC (F59 antibody) in sox6/nfixa double morphants by western blot (Figure 6L). We thus conclude that functional cooperation of Sox6 and Nfix is required for proper skeletal muscle development and that this cooperation is evolutionarily conserved in mouse and zebrafish.
Figure 6.

Functional Cooperation of Sox6 and Nfixa Is Conserved in Zebrafish
(A–J) Immunofluorescence with anti-Sox6 antibody (green) and anti-MyHC antibody (red) (A, B, and G–J) or with anti-Sox6 (green) and anti-sMyHC (F59, red) (C–F) on 2 dpf zebrafish muscle longitudinal sections. Arrowheads indicate Sox-positive nuclei in fast-twitch muscle fibers. Arrows indicate Sox6 staining in the cytoplasm of superficial slow fibers. Approximately one-fifth of the sMyHC-positive superficial cells displayed cytoplasmic Sox6 staining, whereas fast fibers negative for sMyHC only displayed nuclear Sox6 staining. Nuclei are counterstained with Hoechst. Scale bars, 25 μm.
(K) Quantitative real-time PCR analysis on trunk and tail regions at 48 hpf from embryos injected with std-MO or suboptimal doses of nfixa-MO (0.25 pmol), sox6-MO (0.1 pmol), or nfixa-MO + sox6-MO (∗∗p < 0.01; N = 2).
(L) Western blot for sMyHC at 52 hpf on trunk and tail regions of embryos injected with std-MO or suboptimal doses of nfixa-MO, sox6-MO, or nfixa-MO + sox6-MO. Vinculin was used to normalize the amount of loaded proteins.
Discussion
Adult skeletal muscle is composed of two major fiber types presenting a wide range of physiological and biochemical differences. Slow-twitch type I fibers use oxidative metabolism and express the slow MyHC isoform MyHC-I; in contrast, fast-twitch type II fibers present glycolytic or mixed metabolism and express three fast MyHC isoforms (MyHC-IIa, IIx/d, and IIb) (Peter et al., 1972, Schiaffino et al., 1988, Chakkalakal et al., 2012). The phenotype of post-natal muscle fibers is strictly regulated by extrinsic signals such as muscle activity and hormones (Butler-Browne et al., 1982, Gambke et al., 1983, Russell et al., 1988). Additionally, different factors controlling adult muscle plasticity have been identified, including the Nfatc (Calabria et al., 2009) and Mef2 (Wu et al., 2000, Potthoff et al., 2007, Anderson et al., 2015) transcription factor families and PGC-1α (Li et al., 2002). On the contrary, the molecular mechanisms by which muscle fiber diversity is achieved during pre-natal development are still poorly understood. It was shown that the intrinsic transcriptional properties of embryonic and fetal myogenic progenitors are important to set the fiber type in the absence of nerve activity (Cho et al., 1993, Cusella-De Angelis et al., 1994). Moreover, in recent years, several transcription factors contributing to developmental muscle fiber specification have been identified, including Sox6 (Hagiwara et al., 2007, An et al., 2011), Nfix (Messina et al., 2010), Six1/Six4 (Richard et al., 2011), and Nfatc2 (Daou et al., 2013). However, the network of transcription factors controlling fiber specification during embryogenesis is still far from being fully characterized, and until now, functional interactions among the different regulators were completely unknown.
In this work, we provided evidence for functional interplay between Nfix and Sox6 in controlling expression of the slow MyHC isoform during mouse pre-natal muscle development.
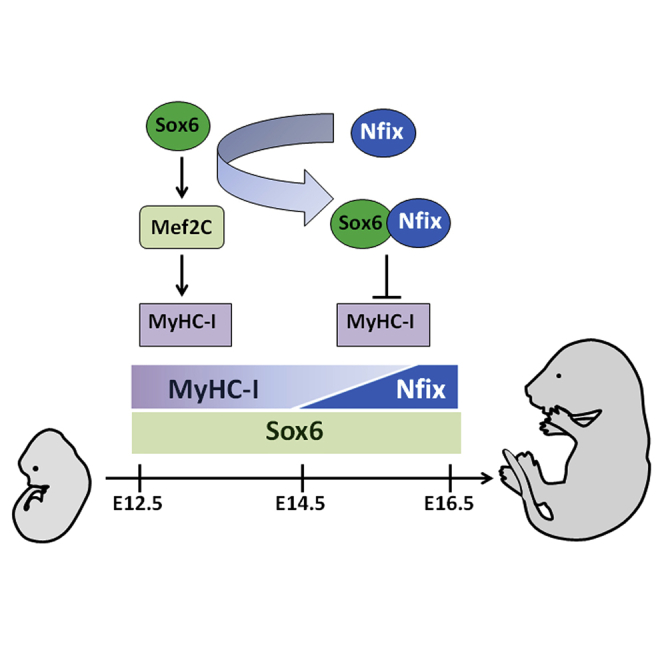
In contrast to Nfix, which is a specific marker of fetal myogenesis (Messina et al., 2010, Mourikis et al., 2012), Sox6 is expressed in both embryonic and fetal purified myoblasts at the mRNA and protein levels. Consistently, we found that Sox6 protein is expressed in skeletal muscle in vivo starting between E11.5 and E12.5. This was unexpected, since Sox6 is known to be a repressor of MyHC-I, which along with MyHC-emb is expressed in all embryonic fibers (Hagiwara et al., 2007, Hutcheson et al., 2009). It is known that Sox6 transcript is absent in mouse primary myotome between E9.5 and E10.75 (Vincent et al., 2012), suggesting that Sox6 is quickly activated in embryonic myoblasts at the beginning of primary myogenesis. Unexpectedly, Sox6 deficiency during primary myogenesis leads to a transient faster muscle phenotype with low levels of MyHC-I. This is followed in Sox6-null fetuses by dramatic upregulation of MyHC-I at the transcription level, consistently with previous characterizations of the Sox6-null phenotype (Hagiwara et al., 2007, An et al., 2011, Quiat et al., 2011). Our results demonstrate that Sox6 plays opposite roles in MyHC-I expression during development. Importantly, in embryonic myogenic cells, Sox6 does not bind to the two canonical MyHC-I binding sites, which are both dispensable for Sox6-dependent embryonic regulation of MyHC-I, at least in our culturing conditions. These data strongly suggest an indirect mechanism of regulation. We found that Sox6 directly binds to the promoter of Mef2C and activates its expression in primary myofibers. Mef2C is part of a transcription factor family that is constitutively expressed in muscle cells since early embryogenesis (Edmondson et al., 1994) and plays an important role in the specification and maintenance of type I fibers (Chin et al., 1998, Wu et al., 2000, Anderson et al., 2015). Importantly, conditional deletion of Mef2C in skeletal muscle leads to a drastic reduction in slow-twitch fibers (Potthoff et al., 2007). Our rescue experiment, although partial, clearly shows that one of the Sox6-dependent effects on MyHC-I transcription in embryonic myofibers is mediated by Mef2C, which is directly targeted by Sox6 in embryonic, but not fetal, muscle. This function led us to hypothesize that the binding ability of Sox6 is differentially regulated in discrete myogenic progenitor populations or at different times during development according to different co-factors, which contribute to the high versatility of Sox6 functions. It is known that SoxD factors, lacking trans-acting functional domains, have a critical requirement for co-factors in order to regulate transcription of target genes (reviewed in Hagiwara, 2011). Therefore, it is likely that the reversal in Sox6 function is due to different factors that are progressively recruited and activated during muscle development. Indeed, we found that during fetal myogenesis, Nfix acts as a fundamental co-factor of Sox6 and is able to form a complex with Sox6, which is no longer able to bind to the Mef2C promoter but can bind to MyHC-I regulative regions, in particular the proximal promoter that was shown to be critical for Sox6-dependent fetal repression of MyHC-I (Hagiwara et al., 2007). In our previous study, we showed that Nfix negatively regulates MyHC-I by repressing Nfatc4, a positive regulator of MyHC-I (Messina et al., 2010). Interestingly, we have now found that during fetal myogenesis, Nfix is also present at the MyHC-I proximal promoter along with Sox6, suggesting that a physical association between these two proteins may be required for proper MyHC-I downregulation. Since Nfix presents both a trans-repression domain and a trans-activation domain, we hypothesized that formation of a complex with Sox6 might provide the basis for the transcriptional repression at the MyHC-I locus (Figure 7). Indeed, Nfix and Sox6 were found to be part of the same complex in fetal myotubes and are both present at the MyHC-I proximal promoter. Moreover, our study on Nfix-null and Sox6-null fetuses clearly shows that Sox6 and Nfix are independently expressed during secondary myogenesis and that neither Sox6 nor Nfix is able to correctly downregulate MyHC-I expression when the other one is not present. A scheme of the scenario in which Sox6 and Nfix behave and cooperate during pre-natal muscle development is shown in Figure 7.
Figure 7.

Model of Sox6 and Nfix Interplay during Pre-natal Myogenesis
Scheme illustrating the functions of Sox6 and Nfix during embryonic (E12.5) and fetal (E16.5) myogenesis. Transcription factors are represented by circles, whereas the regulative region of MyHC-I is indicated by rectangles that are green when transcription is activated and red when transcription is repressed.
In this work, we have also provided evidence for a conserved transcriptional cooperation of Sox6 and Nfixa in zebrafish. We have shown that Sox6 is crucial for the regulation of smyhc1, in line with the findings of Jackson et al. (2015). It was shown that Sox6 expression is silenced in zebrafish slow fibers by the combined effects of Prdm1a and miR-499 (von Hofsten et al., 2008, Wang et al., 2011). We found that a minority of sMyHC-positive cells display cytoplasmic staining for Sox6, suggesting that Sox6 might also be regulated through subcellular localization in different fiber types. Notably, our double partial knockdown performed with suboptimal doses of sox6-MO (von Hofsten et al., 2008) and nfixa-MO (Pistocchi et al., 2013) caused a severe impairment in the touch-evoked escape response, suggesting that Sox6 and Nfixa cooperate in proper muscle function. Molecularly, our data suggest that the two transcription factors can act together to repress smyhc1 expression in zebrafish embryos, whereas the expression of a typical fast-twitch gene, such as myl1 (Burguière et al., 2011), was unaffected in double partial morphants.
In conclusion, we have presented a complex model of regulation of embryonic to fetal MyHC-I regulation that involves a functional interplay between Nfix and Sox6 that is conserved in mammals and teleosts.
Experimental Procedures
Mouse Strains and Fish Lines
The following murine lines were used: Myf5GFP-P/+ (Kassar-Duchossoy et al., 2004), Nfix null (Campbell et al., 2008), Sox6lacZ/+ (Smits et al., 2001), and Tg:Mlc1f-Nfix2 (Messina et al., 2010). For each of these lines, the genotyping strategy has been described in the references. CD1 WT mice (Jackson Laboratory) were used as well. Mice were kept in pathogen-free and controlled conditions, and all procedures conformed to Italian law (D. Lgs n° 2014/26, implementation of the 2010/63/UE) and were approved by the Animal Welfare Body of the University of Milan and the Italian Minister of Health (1212/2015PR). Zebrafish were raised and maintained according to established techniques. The following line was used: AB (from Carole Wilson, UCL, London, UK).
Immunofluorescence on Sections
Mouse or zebrafish embryos were fixed in 4% paraformaldehyde, extensively washed in PBS, and incubated overnight in PBS containing 15% sucrose. Samples were then frozen in nitrogen-chilled isopentane and kept at −80°C until use. Cryostat sections (8 μm thick) were permeabilized in 1% BSA, 0.2% Triton X-100 in PBS for 30 min at room temperature and then incubated for 1 hr in blocking solution (10% goat serum in PBS) and overnight with the primary antibody or with mock PBS. After incubation for 45 min with the fluorescent-conjugated secondary antibody (Jackson ImmunoResearch Laboratories), sections were washed in PBS 0.2% Triton X-100 and mounted, and fluorescent immunolabeling was recorded with a DM6000 Leica microscope. The following primary antibodies were used: rabbit anti-Sox6 (Abcam, 1:300), rabbit anti-Nfix (Novus Biological, 1:200), mouse anti-MyHC-I (Sigma, 1:200), BAD5 (monoclonal, 1:2), MF20 (monoclonal, 1:2), and F59 (monoclonal, 1:10). Nuclei were stained with Hoechst (1:1,000).
Cell Sorting and Culturing
Dissected Myf5GFP-P/+ embryonic or fetal muscles were digested by 0.15 mg/mL Collagenase (Sigma), 1.5 mg/mL Dispase (GIBCO), and 0.1 mg/ml Dnase I (Sigma) for 30 min at 37°C in agitation as described in Biressi et al. (2007). After centrifugation and filtration, cells were collected in DMEM, 20% fetal bovine serum (FBS), 2 mM EDTA, and 20 mM HEPES. For cell sorting, we used the Sorter BD FACSAria. GFP-positive cells were collected for mRNA and protein extraction. For the preparation of unpurified fetal myoblasts after digestion of tissue, cells were pre-plated for 30 min on a plastic dish to lose fibroblasts, which normally adhere to plastic. Unpurified cells were kept in incubation at 37°C in 20% HS (20% horse serum in DMEM) and 24 hr later allowed to differentiate in DM (2% horse serum in DMEM) for 48 hr.
Lentivirus Production, Transduction, and Transfection
Preparation of viral particles were performed by co-transfecting pLKO.1-shSox6 vectors (Thermo Fisher Scientific) or non-targeting shRNA vectors (30 μg), together with the packaging plasmids pMDLg/p (16.25 μg), pCMV-VSVG (9 μg), and pRSV-REV (6.25 μg), in HEK293T cells. Transfection was performed using the calcium phosphate transfection method. Viral particles were collected 40 hr after transfection and subjected to ultracentrifugation at 20,000 rpm for 2 hr at 20°C (Beckman Coulter, Optima L-100 XP). The concentrated viral particles were re-suspended in PBS and stored in aliquots at −80°C until further use. Embryonic myoblasts were transduced by overnight incubation with viral preparation. These preparations were used to transduce embryonic myoblasts at an MOI of 10. The day after transduction, embryonic myoblasts were transfected with Mef2C (pCDNAI/A-Mef2C) or the empty vector as a negative control using Lipofectamine LTX (Invitrogen).
Protein Extraction and Western Blot
Cultured cells were washed twice in ice-cold PBS and then lysed (30 min in ice) with RIPA buffer (10 mM Tris-HCl [pH 8.0], 1 mM EDTA, 1% Triton-X, 0.1% sodium deoxycholate, 0.1% SDS, and 150 mM NaCl in deionized water) plus protease inhibitors (1 mM PMSF). For zebrafish, only trunk and tail regions were used, cutting out the head, and protein was extracted in Laemmli buffer (3 μL per embryo). Protein was harvested after centrifugation, quantified by absorbance reading at 750 nm, and stored at −80°C. 30 μg protein was resolved on 8%–12% polyacrylamide gels or on MiniProtean TDX Gels (Bio-Rad) after denaturation at 95°C for 5 min with SDS-PAGE Loading sample buffer 2X (100 mM Tris [pH 6.8], 4% SDS, 0.2% Bromophenol blue, 20% glycerol, and 10 mM dithiothreitol). For western blot analysis, proteins were transferred onto nitrocellulose with the iBlot Dry Blotting System (Invitrogen). Following transfer, membranes were blocked in 5% milk in Tris-buffered saline (TBS)-T (TBS plus 0.02% Tween20) for 1 hr at room temperature. The primary antibodies used were rabbit anti-Sox6 (Abcam; 1:1,000), mouse anti-MyHC-I (Sigma; 1:5,000), mouse anti-β-tubulin (Santa Cruz Biotechnology; 1:5,000), rabbit anti-Nfix (Novus Biologicals; 1:500), mouse anti-HA (Covance; 1:500), mouse anti-vinculin (Sigma; 1:1,000), and mouse anti-sMyHC (F59, Hybridoma Bank) anti-mouse sarcomeric MyHC (MF20, Hybridoma Bank). Horseradish peroxidase (HRP)-conjugated antibodies (Bio-Rad) were used as secondary antibodies, and the signal was revealed with the ChemiDoc MP System (Bio-Rad).
Transfection and Co-immunoprecipitation Assays
For transfection of fetal myoblasts, WT or Sox6-null cells were plated on 90-mm dishes and allowed to reach 80% confluence in proliferating conditions. Cells were transfected with pCH-Nfix2-HA or pCH-HA plasmids with Lipofectamine LTX (Invitrogen) overnight at 37°C. Cells were kept in DM for 36 hr after transfection, and then nuclear extracts were prepared by collecting 5 × 106 cells in 400 μL ice-cold hypotonic buffer A (10 mM HEPES [pH 7.9], 10 mM KCl, 0.1 mM EGTA, 1 mM DTT, protease inhibitors, 1 mM PMSF, and phosphatase inhibitors) for 15 min in ice. Then, NP40 was added to a final concentration of 0.625%, and cells were centrifuged at 5,000 rpm for 5 min at 4°C. The supernatant was discarded as cytoplasmatic extract, while the pelleted nuclei were resuspended in 100 μL immunoprecipitation lysis buffer containing 50 mM Tris-HCl (pH 7.5), 1% NP40, 1 mM EDTA, 100 mM NaCl, and protease and phosphatase inhibitors. Immunoprecipitation was performed using the immunoprecipitation kit Dynabeads Protein A (Life Technologies). Dynabeads Protein A (50 μL) was incubated for 2 hr with gentle rotation at room temperature with 5 μg rabbit anti-Sox6 antibody (Abcam), rabbit anti-HA antibody (Santa Cruz), or non-related rabbit immunoglobulin G (IgG) (Santa Cruz). Then, the bead-antibody complex was incubated with gentle rotation for 2 hr at 4°C with 1.5 mg total protein lysates per condition. The eluted proteins were denatured with non-reducing SDS-PAGE loading sample buffer (100 mM Tris [pH 6.8], 4% SDS, 0.2% bromophenol blue, and 20% glycerol) and loaded onto a gel.
Chromatin Immunoprecipitation and Reporter Assay
ChIP for Sox6 and luciferase assays were performed as previously published (An et al., 2011). For the detailed protocol, see Supplemental Experimental Procedures.
RNA Extraction and Analysis
RNA from homogenized mouse or zebrafish embryos and isolated cells was extracted with NucleoSpin RNA kits (Macherey-Nagel) following the manufacturer’s instructions. Eluted RNA was checked on agarose gels, quantified with a Nanodrop spectrophotomer, and stored at −80°C. Approximately 0.5 μg RNA was used with the ImProm-II Reverse Transcriptase kit (Promega). Real-time PCR was performed on cDNA using SYBR Green Supermix (Bio-Rad) and the CFX Connect Real Time System (Bio-Rad). After amplification, relative mRNA expression levels were calculated using standard curves from cDNA dilutions and normalized on the Gapdh expression levels. For quantitative real-time PCR in zebrafish, we used rpl8 to normalize the mRNA levels. The primers used are listed in Tables S1 and S2.
MO Microinjections
The antisense MOs (Gene Tools) used in this study, sox6-MO1 (von Hofsten et al., 2008) and nfixa-MO (Pistocchi et al., 2013), were previously described. MOs, diluted in Danieau buffer (Nasevicius and Ekker, 2000), were injected at the one- to two-cell stage. Escalating doses of each MO were tested for phenotypic effects; as control for unspecific effects, each experiment was performed in parallel with an std-MO (standard control oligo) with no target in zebrafish embryos. We usually injected 0.25 pmol sox6-MO per embryo. For combined knockdown experiments, we injected sox6-MO and nfixa-MO at 0.1 and 0.25 or 0.08 and 0.125 pmol per embryo, respectively.
Statistical Analysis
Values were expressed as means ± SD. Statistical significance was assessed by unpaired Student’s t test with Prism 5 software. Statistical significance with a probability of less than 5%, 1%, or 0.1% is indicated in each graph with a single, double, or triple asterisk, respectively, followed by the number of independent experiments (n).
Author Contributions
Conceptualization, G. Maroli, V.T., and G. Messina; Methodology, S.C., A.F., and S.M.; Investigation, V.T., G. Maroli, S.M., S.C., A.F., and G.R.; Writing – Original Draft, G. Maroli and V.T.; Writing – Review & Editing, V.T., S.C., G.R., G.C., M.B., and G. Messina; Supervision, M.B. and G. Messina; Funding Acquisition, G. Messina.
Acknowledgments
We are grateful to S. Antonini, C. Bonfanti, and C. Vezzali for technical assistance and S. Molinari for the Mef2C antibody. We thank Nobuko Hagiwara (UC Davis), Chung-Il An (UC Davis), and Antonella Ronchi (University of Milano-Bicocca) for helpful suggestions and Véronique Lefebvre (Cleveland Clinic) for providing the Sox6lacZ/+ mice. We thank Paolo Zuccolini for help in movie editing.
This work received funding from the ERC StG2011 (RegeneratioNfix 280611) and the Italian Ministry of University and Research (MIUR-Futuro in Ricerca 2010 RBFR 10YNGH-001). M.B. thanks Fondazione Cariplo for support (grant 2011-0555).
Published: November 22, 2016
Footnotes
Supplemental Information includes Supplemental Experimental Procedures, six figures, two tables, and one movie and can be found with this article online at http://dx.doi.org/10.1016/j.celrep.2016.10.082.
Supplemental Information
{kind=link}
References
- An C.I., Dong Y., Hagiwara N. Genome-wide mapping of Sox6 binding sites in skeletal muscle reveals both direct and indirect regulation of muscle terminal differentiation by Sox6. BMC Dev. Biol. 2011;11:59. doi: 10.1186/1471-213X-11-59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson C.M., Hu J., Barnes R.M., Heidt A.B., Cornelissen I., Black B.L. Myocyte enhancer factor 2C function in skeletal muscle is required for normal growth and glucose metabolism in mice. Skelet. Muscle. 2015;5:7. doi: 10.1186/s13395-015-0031-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Biressi S., Tagliafico E., Lamorte G., Monteverde S., Tenedini E., Roncaglia E., Ferrari S., Ferrari S., Cusella-De Angelis M.G., Tajbakhsh S., Cossu G. Intrinsic phenotypic diversity of embryonic and fetal myoblasts is revealed by genome-wide gene expression analysis on purified cells. Dev. Biol. 2007;304:633–651. doi: 10.1016/j.ydbio.2007.01.016. [DOI] [PubMed] [Google Scholar]
- Blow M.J., McCulley D.J., Li Z., Zhang T., Akiyama J.A., Holt A., Plajzer-Frick I., Shoukry M., Wright C., Chen F. ChIP-Seq identification of weakly conserved heart enhancers. Nat. Genet. 2010;42:806–810. doi: 10.1038/ng.650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burguière A.C., Nord H., von Hofsten J. Alkali-like myosin light chain-1 (myl1) is an early marker for differentiating fast muscle cells in zebrafish. Dev. Dyn. 2011;240:1856–1863. doi: 10.1002/dvdy.22677. [DOI] [PubMed] [Google Scholar]
- Butler-Browne G.S., Bugaisky L.B., Cuénoud S., Schwartz K., Whalen R.G. Denervation of newborn rat muscle does not block the appearance of adult fast myosin heavy chain. Nature. 1982;299:830–833. doi: 10.1038/299830a0. [DOI] [PubMed] [Google Scholar]
- Calabria E., Ciciliot S., Moretti I., Garcia M., Picard A., Dyar K.A., Pallafacchina G., Tothova J., Schiaffino S., Murgia M. NFAT isoforms control activity-dependent muscle fiber type specification. Proc. Natl. Acad. Sci. USA. 2009;106:13335–13340. doi: 10.1073/pnas.0812911106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Campbell C.E., Piper M., Plachez C., Yeh Y.T., Baizer J.S., Osinski J.M., Litwack E.D., Richards L.J., Gronostajski R.M. The transcription factor Nfix is essential for normal brain development. BMC Dev. Biol. 2008;8:52. doi: 10.1186/1471-213X-8-52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cao Y., Yao Z., Sarkar D., Lawrence M., Sanchez G.J., Parker M.H., MacQuarrie K.L., Davison J., Morgan M.T., Ruzzo W.L. Genome-wide MyoD binding in skeletal muscle cells: a potential for broad cellular reprogramming. Dev. Cell. 2010;18:662–674. doi: 10.1016/j.devcel.2010.02.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chakkalakal J.V., Kuang S., Buffelli M., Lichtman J.W., Sanes J.R. Mouse transgenic lines that selectively label Type I, Type IIA, and Types IIX+B skeletal muscle fibers. Genesis. 2012;50:50–58. doi: 10.1002/dvg.20794. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chin E.R., Olson E.N., Richardson J.A., Yang Q., Humphries C., Shelton J.M., Wu H., Zhu W., Bassel-Duby R., Williams R.S. A calcineurin-dependent transcriptional pathway controls skeletal muscle fiber type. Genes Dev. 1998;12:2499–2509. doi: 10.1101/gad.12.16.2499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cho M., Webster S.G., Blau H.M. Evidence for myoblast-extrinsic regulation of slow myosin heavy chain expression during muscle fiber formation in embryonic development. J. Cell Biol. 1993;121:795–810. doi: 10.1083/jcb.121.4.795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Christ B., Ordahl C.P. Early stages of chick somite development. Anat. Embryol. (Berl.) 1995;191:381–396. doi: 10.1007/BF00304424. [DOI] [PubMed] [Google Scholar]
- Cusella-De Angelis M.G., Molinari S., Le Donne A., Coletta M., Vivarelli E., Bouche M., Molinaro M., Ferrari S., Cossu G. Differential response of embryonic and fetal myoblasts to TGF beta: a possible regulatory mechanism of skeletal muscle histogenesis. Development. 1994;120:925–933. doi: 10.1242/dev.120.4.925. [DOI] [PubMed] [Google Scholar]
- Daou N., Lecolle S., Lefebvre S., della Gaspera B., Charbonnier F., Chanoine C., Armand A.S. A new role for the calcineurin/NFAT pathway in neonatal myosin heavy chain expression via the NFATc2/MyoD complex during mouse myogenesis. Development. 2013;140:4914–4925. doi: 10.1242/dev.097428. [DOI] [PubMed] [Google Scholar]
- Edmondson D.G., Lyons G.E., Martin J.F., Olson E.N. Mef2 gene expression marks the cardiac and skeletal muscle lineages during mouse embryogenesis. Development. 1994;120:1251–1263. doi: 10.1242/dev.120.5.1251. [DOI] [PubMed] [Google Scholar]
- Eusebi V., Rilke F., Ceccarelli C., Fedeli F., Schiaffino S., Bussolati G. Fetal heavy chain skeletal myosin. An oncofetal antigen expressed by rhabdomyosarcoma. Am. J. Surg. Pathol. 1986;10:680–686. doi: 10.1097/00000478-198610000-00003. [DOI] [PubMed] [Google Scholar]
- Ferrari S., Molinari S., Melchionna R., Cusella-De Angelis M.G., Battini R., De Angelis L., Kelly R., Cossu G. Absence of MEF2 binding to the A/T-rich element in the muscle creatine kinase (MCK) enhancer correlates with lack of early expression of the MCK gene in embryonic mammalian muscle. Cell Growth Differ. 1997;8:23–34. [PubMed] [Google Scholar]
- Gambke B., Lyons G.E., Haselgrove J., Kelly A.M., Rubinstein N.A. Thyroidal and neural control of myosin transitions during development of rat fast and slow muscles. FEBS Lett. 1983;156:335–339. doi: 10.1016/0014-5793(83)80524-9. [DOI] [PubMed] [Google Scholar]
- Giger J.M., Haddad F., Qin A.X., Baldwin K.M. In vivo regulation of the beta-myosin heavy chain gene in soleus muscle of suspended and weight-bearing rats. Am. J. Physiol. Cell Physiol. 2000;278:C1153–C1161. doi: 10.1152/ajpcell.2000.278.6.C1153. [DOI] [PubMed] [Google Scholar]
- Hagiwara N. Sox6, jack of all trades: a versatile regulatory protein in vertebrate development. Dev. Dyn. 2011;240:1311–1321. doi: 10.1002/dvdy.22639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hagiwara N., Yeh M., Liu A. Sox6 is required for normal fiber type differentiation of fetal skeletal muscle in mice. Dev. Dyn. 2007;236:2062–2076. doi: 10.1002/dvdy.21223. [DOI] [PubMed] [Google Scholar]
- Hutcheson D.A., Zhao J., Merrell A., Haldar M., Kardon G. Embryonic and fetal limb myogenic cells are derived from developmentally distinct progenitors and have different requirements for beta-catenin. Genes Dev. 2009;23:997–1013. doi: 10.1101/gad.1769009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jackson H.E., Ono Y., Wang X., Elworthy S., Cunliffe V.T., Ingham P.W. The role of Sox6 in zebrafish muscle fiber type specification. Skelet. Muscle. 2015;5:2. doi: 10.1186/s13395-014-0026-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kassar-Duchossoy L., Gayraud-Morel B., Gomès D., Rocancourt D., Buckingham M., Shinin V., Tajbakhsh S. Mrf4 determines skeletal muscle identity in Myf5:Myod double-mutant mice. Nature. 2004;431:466–471. doi: 10.1038/nature02876. [DOI] [PubMed] [Google Scholar]
- Kelly R.G., Zammit P.S., Schneider A., Alonso S., Biben C., Buckingham M.E. Embryonic and fetal myogenic programs act through separate enhancers at the MLC1F/3F locus. Dev. Biol. 1997;187:183–199. doi: 10.1006/dbio.1997.8577. [DOI] [PubMed] [Google Scholar]
- Li J., Kinoshita T., Pandey S., Ng C.K., Gygi S.P., Shimazaki K., Assmann S.M. Modulation of an RNA-binding protein by ascisic-acid-activated protein kinase. Nature. 2002;418:793–797. doi: 10.1038/nature00936. [DOI] [PubMed] [Google Scholar]
- Lyons G.E., Ontell M., Cox R., Sassoon D., Buckingham M. The expression of myosin genes in developing skeletal muscle in the mouse embryo. J. Cell Biol. 1990;111:1465–1476. doi: 10.1083/jcb.111.4.1465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Messina G., Biressi S., Monteverde S., Magli A., Cassano M., Perani L., Roncaglia E., Tagliafico E., Starnes L., Campbell C.E. Nfix regulates fetal-specific transcription in developing skeletal muscle. Cell. 2010;140:554–566. doi: 10.1016/j.cell.2010.01.027. [DOI] [PubMed] [Google Scholar]
- Mourikis P., Gopalakrishnan S., Sambasivan R., Tajbakhsh S. Cell-autonomous Notch activity maintains the temporal specification potential of skeletal muscle stem cells. Development. 2012;139:4536–4548. doi: 10.1242/dev.084756. [DOI] [PubMed] [Google Scholar]
- Nasevicius A., Ekker S.C. Effective targeted gene ‘knockdown’ in zebrafish. Nat. Genet. 2000;26:216–220. doi: 10.1038/79951. [DOI] [PubMed] [Google Scholar]
- Page S., Miller J.B., DiMario J.X., Hager E.J., Moser A., Stockdale F.E. Developmentally regulated expression of three slow isoforms of myosin heavy chain: diversity among the first fibers to form in avian muscle. Dev. Biol. 1992;154:118–128. doi: 10.1016/0012-1606(92)90053-j. [DOI] [PubMed] [Google Scholar]
- Peter J.B., Barnard R.J., Edgerton V.R., Gillespie C.A., Stempel K.E. Metabolic profiles of three fiber types of skeletal muscle in guinea pigs and rabbits. Biochemistry. 1972;11:2627–2633. doi: 10.1021/bi00764a013. [DOI] [PubMed] [Google Scholar]
- Pinney D.F., Pearson-White S.H., Konieczny S.F., Latham K.E., Emerson C.P., Jr. Myogenic lineage determination and differentiation: evidence for a regulatory gene pathway. Cell. 1988;53:781–793. doi: 10.1016/0092-8674(88)90095-5. [DOI] [PubMed] [Google Scholar]
- Pistocchi A., Gaudenzi G., Foglia E., Monteverde S., Moreno-Fortuny A., Pianca A., Cossu G., Cotelli F., Messina G. Conserved and divergent functions of Nfix in skeletal muscle development during vertebrate evolution. Development. 2013;140:1528–1536. doi: 10.1242/dev.076315. [DOI] [PubMed] [Google Scholar]
- Potthoff M.J., Arnold M.A., McAnally J., Richardson J.A., Bassel-Duby R., Olson E.N. Regulation of skeletal muscle sarcomere integrity and postnatal muscle function by Mef2c. Mol. Cell. Biol. 2007;27:8143–8151. doi: 10.1128/MCB.01187-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Quiat D., Voelker K.A., Pei J., Grishin N.V., Grange R.W., Bassel-Duby R., Olson E.N. Concerted regulation of myofiber-specific gene expression and muscle performance by the transcriptional repressor Sox6. Proc. Natl. Acad. Sci. USA. 2011;108:10196–10201. doi: 10.1073/pnas.1107413108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richard A.F., Demignon J., Sakakibara I., Pujol J., Favier M., Strochlic L., Le Grand F., Sgarioto N., Guernec A., Schmitt A. Genesis of muscle fiber-type diversity during mouse embryogenesis relies on Six1 and Six4 gene expression. Dev. Biol. 2011;359:303–320. doi: 10.1016/j.ydbio.2011.08.010. [DOI] [PubMed] [Google Scholar]
- Rossi G., Antonini S., Bonfanti C., Monteverde S., Vezzali C., Tajbakhsh S., Cossu G., Messina G. Nfix regulates temporal progression of muscle regeneration through modulation of myostatin expression. Cell Rep. 2016;14:2238–2249. doi: 10.1016/j.celrep.2016.02.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Russell S.D., Cambon N., Nadal-Ginard B., Whalen R.G. Thyroid hormone induces a nerve-independent precocious expression of fast myosin heavy chain mRNA in rat hindlimb skeletal muscle. J. Biol. Chem. 1988;263:6370–6374. [PubMed] [Google Scholar]
- Schiaffino S., Gorza L., Sartore S., Saggin L., Carli M. Embryonic myosin heavy chain as a differentiation marker of developing human skeletal muscle and rhabdomyosarcoma. A monoclonal antibody study. Exp. Cell Res. 1986;163:211–220. doi: 10.1016/0014-4827(86)90574-4. [DOI] [PubMed] [Google Scholar]
- Schiaffino S., Ausoni S., Gorza L., Saggin L., Gundersen K., Lomo T. Myosin heavy chain isoforms and velocity of shortening of type 2 skeletal muscle fibres. Acta Physiol. Scand. 1988;134:575–576. doi: 10.1111/j.1748-1716.1998.tb08539.x. [DOI] [PubMed] [Google Scholar]
- Smits P., Li P., Mandel J., Zhang Z., Deng J.M., Behringer R.R., de Crombrugghe B., Lefebvre V. The transcription factors L-Sox5 and Sox6 are essential for cartilage formation. Dev. Cell. 2001;1:277–290. doi: 10.1016/s1534-5807(01)00003-x. [DOI] [PubMed] [Google Scholar]
- Stockdale F.E. Myogenic cell lineages. Dev. Biol. 1992;154:284–298. doi: 10.1016/0012-1606(92)90068-r. [DOI] [PubMed] [Google Scholar]
- Vincent S.D., Mayeuf A., Niro C., Saitou M., Buckingham M. Non conservation of function for the evolutionarily conserved prdm1 protein in the control of the slow twitch myogenic program in the mouse embryo. Mol. Biol. Evol. 2012;29:3181–3191. doi: 10.1093/molbev/mss125. [DOI] [PubMed] [Google Scholar]
- von Hofsten J., Elworthy S., Gilchrist M.J., Smith J.C., Wardle F.C., Ingham P.W. Prdm1- and Sox6-mediated transcriptional repression specifies muscle fibre type in the zebrafish embryo. EMBO Rep. 2008;9:683–689. doi: 10.1038/embor.2008.73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang X., Ono Y., Tan S.C., Chai R.J., Parkin C., Ingham P.W. Prdm1a and miR-499 act sequentially to restrict Sox6 activity to the fast-twitch muscle lineage in the zebrafish embryo. Development. 2011;138:4399–4404. doi: 10.1242/dev.070516. [DOI] [PubMed] [Google Scholar]
- Wu H., Naya F.J., McKinsey T.A., Mercer B., Shelton J.M., Chin E.R., Simard A.R., Michel R.N., Bassel-Duby R., Olson E.N., Williams R.S. MEF2 responds to multiple calcium-regulated signals in the control of skeletal muscle fiber type. EMBO J. 2000;19:1963–1973. doi: 10.1093/emboj/19.9.1963. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.