

Abstract
Can economic forces be harnessed for biodiversity conservation? The answer hinges on characterizing the value of nature, a tricky business from biophysical, socioeconomic, and ethical perspectives. Although the societal benefits of native ecosystems are clearly immense, they remain largely unquantified for all but a few services. Here, we estimate the value of tropical forest in supplying pollination services to agriculture. We focus on coffee because it is one of the world's most valuable export commodities and is grown in many of the world's most biodiverse regions. Using pollination experiments along replicated distance gradients, we found that forest-based pollinators increased coffee yields by 20% within ≈1 km of forest. Pollination also improved coffee quality near forest by reducing the frequency of “peaberries” (i.e., small misshapen seeds) by 27%. During 2000–2003, pollination services from two forest fragments (46 and 111 hectares) translated into ≈$60,000 (U.S.) per year for one Costa Rican farm. This value is commensurate with expected revenues from competing land uses and far exceeds current conservation incentive payments. Conservation investments in human-dominated landscapes can therefore yield double benefits: for biodiversity and agriculture.
Keywords: bees, ecosystem service, landscape, pollination
Ecosystem services are those processes through which natural systems support and fulfill human life (1). Although the societal benefits of native ecosystems are clearly immense (2–5), they remain largely unquantified for all but a few services (e.g., carbon sequestration, water flow) (6, 7). Crop pollination is an ecosystem service of enormous economic value (8–11). Roughly two-thirds of the world's crop species include cultivars that require animal pollination (12, 13). Recent declines in both managed and wild bee populations have aroused global concern (14–17), prompting the United Nations Convention on Biological Diversity and Food and Agriculture Organization to create the International Pollinators Initiative to coordinate scientific investigation and pollinator conservation (18). Repeated studies have shown that wild bees pollinate many crops as effectively as do managed bees; however, maintaining wild bee populations requires conserving their habitats within agricultural landscapes (e.g., refs. 19–22).
Coffee (Coffea arabica and Coffea robusta) ranks among the five most valuable agricultural exports from developing nations (Food and Agriculture Organization, http://apps.fao.org), employs >25 million people worldwide (23), and is cultivated in many of the world's most biodiverse regions (24–26). C. arabica (the higher-quality, highland species studied here) is able to self-pollinate, but bee visitation can increase yields 15–50% over treatments with bees excluded (12, 27, 28). In coffee and other crops, pollinator diversity and visitation rate have been found to decline with increasing isolation from patches of native habitat, and this decline can affect yields (22, 29–31). Together, these studies suggest the importance of native habitats to agriculture, but none has yet combined ecological experiments and economic calculations to quantify the value of habitat conservation to farmers. Such quantification is key to informing land-use decisions in the face of difficult tradeoffs between conservation and agricultural production.
We address these issues in a Costa Rican landscape comprising coffee farms, forest fragments, and various other agricultural land uses. Our previous work in this landscape has shown that bee species richness and visitation rate decline significantly with distance from forest (31). Dominant visitors to coffee flowers include nonnative feral honey bees (Apis mellifera) and 10 native species of eusocial, “stingless” bees (Apidae: Meliponini). Here, we conduct pollination experiments to examine the effects of these patterns on coffee production and quality, and to estimate the resulting economic value of tropical forest fragments to coffee farms.
Methods
Landscape and Experimental Design. Our focal landscape is dominated by Finca Santa Fe, a large [1,065 hectare (ha)] coffee farm in the Valle General, Costa Rica (Fig. 1). Several forest fragments border the farm, which is managed similarly throughout its extent, including weed and pest control methods, planting and harvest practices, and shade tree species (Eucalyptus deglupta) and density. The expansive landscape thus suits our purposes well, allowing replicated distance gradients from forest into coffee while holding other important variables constant. In 2001, we placed 12 sites along gradients from two forest patches adjacent to the farm (Fig. 1, patches A and B) at three distance classes from forest: near (within 100 m); intermediate (700–800 m); and far (1,400–1,600 m). In 2002, farm managers considered abandoning the northern section of the farm, so we used only near and intermediate sites in the southern portion. We included one additional intermediate site and two additional near sites to maintain replication and placed the two additional near sites near a second southern forest patch (Fig. 1, patch C) to reduce pseudoreplication (see Supporting Text, which is published as supporting information on the PNAS web site). All study sites contained plants of the same variety (C. arabica, “Caturra” variety) and age (8–10 years). No honey bees are currently managed in the area, but feral, Africanized honey bees (A. mellifera) are abundant. As in many Costa Rican farms, shade trees were young and carefully managed, and therefore did not provide cavities preferred for nesting sites by common coffee-pollinating bees (32). Forest patches represented tropical/premontane moist forests (33) and had experienced modest selective harvest. (For additional information, see ref. 31.)
Fig. 1.

Map of study area and sites. Finca Santa Fe (1,065 ha) is in white; stippled area is a mix of coffee, pasture, and sugar cane; black areas are forests. The three focal forest patches are labeled A (46 ha), B (111 ha), and C (34 ha). Study sites are labeled n, i, and f for near, intermediate, and far distance classes.
Pollination Experiments. In each site, we chose five healthy plants, selected four branches on each that were well matched in length, shade, and vertical position, and divided the branches randomly between two treatments: hand-pollination (receiving augmented pollen to measure production with abundant cross-pollination) and control (unmanipulated to measure production under ambient pollination) (34). To hand-pollinate, we removed flowers from nearby individuals and gently dabbed their anthers directly onto the stigma of each flower on the branch, depositing an average of 3,570 pollen grains per stigma. We compared these treatments in terms of three response variables: seed mass, fruit set, and peaberry frequency. We measured seed mass by harvesting ripe fruits and weighing their wet seeds individually. We measured fruit set for each branch by dividing the number of fruits at harvest by the original number of flowers. We measured peaberry frequency for each branch by dividing the number of peaberries (i.e., small misshapen seeds resulting from the failure of one of the two ovules to develop) by the total number of harvested fruit. Farmers sell coffee in various units, including wet and dry seed mass, fruit mass, and fruit volume. Although we present only wet seed mass here, early analyses found this measure to be highly correlated with all others (Pearson correlations: dry seed mass, r = 0.95; fruit mass, r = 0.91; fruit volume, r = 0.84, all n = 187, P « 0.001). (See Supporting Text.)
Statistical Analyses. For all three response variables, we first calculated residual values for all branches around their plant means (i.e., we computed the mean of all branch values on the same plant, and subtracted this plant mean from each branch value). This technique controls for substantial interplant variation (caused by soil chemistry, moisture, microclimate, pruning history, etc.) and thus isolates the differences between treatments that are of interest here. Using residuals is equivalent to adding “PLANT” as a factor in the ANOVAs while avoiding overparameterization of the models and allowing key results to be presented graphically. We then pooled all branches (and both years, when applicable) within each site and used sites as appropriate replicates in ANOVA models. We calculated percentage yield effects as [(hand mean) – (control mean)]/(control mean). We also measured three potential covariates of productivity for each branch: length (number of nodes), number of leaves, and relative shade. None of the three response variables were significantly related to any of these covariates, so the covariates were not included in ANOVA models (Pearson correlations, all r < 0.19, n = 362, not significant, probabilities adjusted for multiple comparisons with Bonferroni method).
Economic Estimates. We used the results of our pollination experiments to estimate the value to Finca Santa Fe of the two major forest patches bordering the farm (Fig. 1, patches A and B). We ignored the effect on peaberry frequency because its value is context-dependent (see Results and Discussion). Previous work has shown that a 32-ha forest patch (Fig. 1, patch C) supplied abundant pollinators to coffee, but an 18-ha riparian strip did not (31). We therefore initially assumed that patches must be ≥20 ha to supply pollinators, but we explored this assumption by repeating the same calculations under six other patch-size thresholds. We rounded all results to the nearest $1,000 to reflect the precision of our assumptions and estimates.
Results and Discussion
Coffee Production and Quality. Pollination by wild bees increased coffee yields near forest patches (Fig. 2). Ambient pollination services were adequate in near and intermediate sites; neither seed mass nor fruit set increased with hand-pollination (Fig. 2 a and b). In far sites, however, adding pollen increased the mean mass of harvested seeds by 8.3% and fruit set by 11.5%, indicating that yields had been depressed by inadequate pollination (Fig. 2 a and b). Multiplying to calculate the combined effect of seed mass and fruit set (1.083 × 1.115 = 1.208), we estimate that adequate pollination in far sites would increase coffee yields by 20.8%.
Fig. 2.

Measures of coffee yield and quality along distance gradient from forest patches. (a) Residual seed mass (two-way ANOVA, distance × treatment interaction: F2,24 = 18.13, P < 0.0001). (b) Residual fruit set (distance × treatment interaction: F2,24 = 2.96, P = 0.0710). (c) Residual peaberry frequency (distance × treatment interaction: F2,24 = 7.28, P = 0.0034). Means ± 1 SE are shown. Asterisks denote a significant difference at that distance class (t tests with Bonferroni correction; *, P ≤ 0.05; **, P ≤ 0.01; ***, P ≤ 0.001). Residuals were obtained by calculating mean for each plant and subtracting it from each branch value, thereby controlling for variation among plant means to isolate treatment comparisons within plants. Because all plant means are thus standardized to zero, treatment means in each distance class are, by definition, symmetrical around zero. Therefore, only the interactions between hand-pollinated and control residuals are meaningful, not any trend in either treatment across distance classes.
Pollination also improved coffee quality near forest by reducing the frequency of peaberries. Similar to the yield variables, treatments did not differ in near and intermediate sites, but in far sites hand-pollination decreased the frequency of peaberries by 27% (Fig. 2c). Uniform seed size and appearance is important for even roasting of coffee, so the markedly smaller peaberries are often thought to lower quality (T. Hamner, personal communication; R. A. Rice, personal communication; Coffee Research Institute, www.coffeeresearch.org). Some specialty coffee suppliers market pure peaberry coffee at a premium, touting its unique and powerful flavor profile. The market for peaberry coffee is small and unreliable, however, and accessing it requires costly separation of peaberries (T. Hamner, personal communication; R. A. Rice, personal communication).
The diverse, abundant, and active pollinator community supplied by native tropical forest (31) therefore increased both the quantity and quality of harvested coffee nearby. How? Pollinators probably provided higher rates of outcrossing among plants, leading to larger and more robust fruit (12, 28). Coffee flowers typically contain only two ovules (12), and stigmas received hundreds of pollen grains even in distant sites (31). Simple relief from pollen limitation is thus unlikely to account for the effects we found. Native bee species, by moving among plants more frequently, may cross-pollinate plants more effectively than honey bees, which often focus on single branches when flowers are dense (ref. 29 and personal observations). In addition, some native bees can deposit more pollen on stigmas per visit than honey bees (20). A diverse community of pollinators may provide greater and more stable pollination services through complementary foraging behaviors, greater pollination efficiencies, and broader climate tolerances, as well as asynchronous population dynamics (22, 29, 31).
Although both bee activity (31) and pollination adequacy (this study) declined significantly with increasing distance from forest, the scales of these two patterns differed somewhat. The major changes in bee activity occurred between the near and intermediate sites (31), but inadequate pollination was evident only in the far sites (Fig. 2). The reasons for this scale mismatch are difficult to determine, but three possibilities deserve mention. First, because bees must fly farther from forest patches to reach far sites, they may arrive later in the day, after many flowers have self-pollinated and are no longer receptive (12). Second, sites nearer to forest are also nearer to neighboring farms (Fig. 1). Bees visiting these sites may therefore visit other coffee plantations and carry more genetically heterogeneous pollen, resulting in wider outcrossing and more vigorous fruits. Third, results from near sites suggest that hand-pollination may slightly damage stigmas and result in yields below potential (34). In near sites, hand-pollinated branches showed lower production and quality than ambient controls (significant only in seed mass; Fig. 2). If hand-pollinated means were corrected so that treatments were equal in near sites, hand-pollination may exceed the control in intermediate sites as well. Such a correction would also result in a larger estimate of pollination effects in far sites, but to remain conservative we calculate only differences at far sites.
Economic Value of Forest. We combine the results of our experiments with data on farm yields and market prices to estimate the income contributed to Finca Santa Fe by two major neighboring forest patches (Fig. 1, patches A and B). To be conservative and simple, we assume that pollination effects from forest extend to 1 km, just beyond the intermediate sites (Fig. 2). Income resulting from these patches can be calculated as
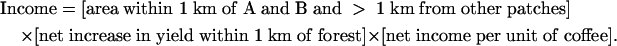 |
[1] |
In Finca Santa Fe, 480 ha are within 1 km of patches A and B and beyond 1 km from any other patch of significant size (i.e., ≥20 ha; see Methods). Mean reported yield for the entire farm in 2000–2003 was 20.0 fanegas (fa) per ha (M. Jimenez, farm manager, personal communication; 1 fa = 200 liters of berries). This farm-wide yield translates to 21.5 fa/ha within 1 km of forest and 17.8 fa/ha beyond 1 km of forest (i.e., an area-weighted difference of 20.8%, or 3.7 fa/ha). Net income per fa averaged $34.75 (range $25–43) for 2000–2003 (market price minus $22 harvest costs; M. Jimenez, personal communication). Plugging these estimates into Eq. 1, we calculate the income resulting from patches A and B to be
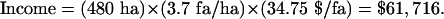 |
[2] |
Pollination services from these patches therefore contributed an average of U.S. $62,000 per year (i.e., 7% of total farm income) in 2000–2003, years of depressed coffee prices. This estimate incorporates both increased income from greater production and increased costs of harvesting the larger crop. Coffee production involves other costs (e.g., chemical inputs and pruning), but these do not typically change with yield, so they do not affect these estimates. Relaxing the minimum patch size assumption of 20 ha (see Methods) to 15 ha and even 10 ha produces similar results, and the estimated value remains substantial with no minimum size assumption at all (Table 1).
Table 1.
Estimates of economic value of forest patches A and B (Fig. 1) to study farm, under seven different assumptions for minimum patch size required to sustain pollination services
| Patch size threshold, ha | Area near A and B and far from all other patches above threshold size, * ha | Income resulting from A and B, † $/yr |
|---|---|---|
| None | 235 | 30,000 |
| 5 | 270 | 35,000 |
| 10 | 363 | 47,000 |
| 15 | 450 | 58,000 |
| 20‡ | 480 | 62,000 |
| 25 | 480 | 62,000 |
| 30 | 480 | 62,000 |
Near area defined as within 1.0 km of forest.
Results rounded to the nearest $1,000 (see Methods).
Same as assuming threshold of 18 ha (the size of the riparian strip; see Methods), because there are no patches >18 and <20 ha.
Because Finca Santa Fe does not own these forest patches, the additional income constitutes a subsidy to the farm, for which the forest is not valued and its owners not compensated (4). For comparison, Costa Rica's innovative Environmental Service Payments Program would pay landowners roughly $6,600 ($42 per ha) annually to conserve these 157 ha of forest (35). Common nonforest land uses in the region earn between $24,000 per year (pasture for beef cattle, averaging $151 per ha per year) and $130,000 year (sugar cane, averaging $825 per ha per year) (36). (Land that still retains forest cover, however, is typically less suited to cattle or agricultural production than land already converted.) The value of forest in providing crop pollination services alone is, therefore, an order of magnitude greater than that recognized by Costa Rica for other forest ecosystem services, of at least the same order as major competing land uses, and infinitely greater than that recognized by most governments (i.e., zero).
Although simple, these calculations illustrate the potential economic value of forest conservation in agricultural landscapes, and they are likely underestimates for several reasons. First, we estimated benefits only to a single farm; several other coffee farms surround the same fragments and presumably benefit from pollination services they provide. Second, the fragments considered here may be larger than necessary to sustain pollinator communities, resulting in lower per-hectare estimates of value. Finally, including other services provided by these fragments (e.g., carbon storage and water purification) would increase estimates of their value (1). Policies that allow landowners to capture the value of pollination and other services could provide powerful incentives for forest conservation in some of the most biodiverse and threatened regions on Earth.
Supplementary Material
Acknowledgments
We thank B. Ettinger, J. Florez, D. Goehring, M. Hayden, A. Herra, A. Kane, P. Oliveira, B. Reed, J. Schwan, Finca Santa Fe, the Fallas family, and the Organization for Tropical Studies for field and laboratory assistance; R. Brooks for bee identifications; P. Armsworth, A. Balmford, C. Boggs, K. Carney, J. Clay, W. Falcon, J. Hellmann, J. Hughes, D. Inouye, P. Kevan, M. Mayfield, D. Roubik, N. Sanders, J. Vandermeer, and N. Williams for helpful comments and discussions; and the Summit Foundation for financial support.
Abbreviations: ha, hectare; fa, fanegas.
References
- 1.Daily, G. C. (1997) Nature's Services (Island, Washington, DC).
- 2.Daily, G. C., Soderqvist, T., Aniyar, S., Arrow, K., Dasgupta, P., Ehrlich, P. R., Folke, C., Jansson, A., Jansson, B. O., Kautsky, N., et al. (2000) Science 289, 395–396. [DOI] [PubMed] [Google Scholar]
- 3.Balmford, A., Bruner, A., Cooper, P., Costanza, R., Farber, S., Green, R. E., Jenkins, M., Jefferiss, P., Jessamy, V., Madden, J., et al. (2002) Science 297, 950–953. [DOI] [PubMed] [Google Scholar]
- 4.Heal, G. (2000) Nature and the Marketplace (Island, Washington, DC).
- 5.Alcamo, J., Ash, N. J., Butler, C. D., Callicot, J. B., Capistrano, D. & Carpenter, S. R. (2003) Ecosystems and Human Well-Being (Island, Washington, DC).
- 6.Fang, J., Chen, A., Peng, C., Zhao, S. & Ci, L. (2001) Science 292, 2320–2322. [DOI] [PubMed] [Google Scholar]
- 7.Guo, Z., Xiao, X. & Li, D. (2000) Ecol. Appl. 10, 925–936. [Google Scholar]
- 8.Kevan, P. G. & Phillips, T. P. (June 2001) Conserv. Ecol. 5, www.consecol.org/vol5/iss1/art8.
- 9.Southwick, E. E. & Southwick, L. (1992) J. Econ. Entomol. 85, 621–633. [Google Scholar]
- 10.Cane, J. H. (1997) Acta Horticulturae 446, 67–70. [Google Scholar]
- 11.Nabhan, G. P. & Buchmann, S. L. (1997) in Nature's Services, ed. Daily, G. C. (Island, Washington, DC), pp. 133–150.
- 12.Free, J. B. (1993) Insect Pollination of Crops (Academic, London).
- 13.Roubik, D. W. (1995) Pollination of Cultivated Plants in the Tropics (Food and Agriculture Organization, Rome).
- 14.Cane, J. H. & Tepedino, V. J. (June 2001) Conserv. Ecol. 5, www.consecol.org/vol5/iss1/art1.
- 15.Buchmann, S. L. & Nabhan, G. P. (1996) The Forgotten Pollinators (Island, Washington, DC).
- 16.Allen-Wardell, G., Bernhardt, P., Bitner, R., Burquez, A., Buchmann, S., Cane, J., Cox, P. A., Dalton, V., Feinsinger, P., Ingram, M., et al. (1998) Conserv. Biol. 12, 8–17. [Google Scholar]
- 17.Watanabe, M. E. (1994) Science 265, 1170–1170. [DOI] [PubMed] [Google Scholar]
- 18.Kevan, P. G., Frankie, G. W., O'Toole, C. & Vergara, C. H. (2002) Pollinating Bees: The Conservation Link Between Agriculture and Nature (Minesterio do Meio Ambiente, Brasilia, Brazil).
- 19.Heard, T. A. (1999) Annu. Rev. Entomol. 44, 183–206. [DOI] [PubMed] [Google Scholar]
- 20.Freitas, B. M. & Paxton, R. J. (1998) J. Appl. Ecol. 35, 109–121. [Google Scholar]
- 21.Richards, A. J. (2001) Ann. Bot. 88, 165–172. [Google Scholar]
- 22.Kremen, C., Williams, N. M. & Thorp, R. W. (2002) Proc. Natl. Acad. Sci. USA 99, 16812–16816. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.O'Brien, T. G. & Kinnaird, M. F. (2003) Science 300, 587. [DOI] [PubMed] [Google Scholar]
- 24.Clay, J. (2004) World Agriculture and the Environment (Island, Washington, DC).
- 25.Myers, N., Mittermeier, R. A., Mittermeier, C. G., daFonseca, G. A. B. & Kent, J. (2000) Nature 403, 853–858. [DOI] [PubMed] [Google Scholar]
- 26.Olson, D. M. & Dinerstein, E. (1998) Conserv. Biol. 12, 502–515. [Google Scholar]
- 27.Roubik, D. W. (2002) Nature 417, 708. [DOI] [PubMed] [Google Scholar]
- 28.Klein, A., Steffan-Dewenter, I. & Tscharntke, T. (2003) Am. J. Bot. 90, 153–157. [DOI] [PubMed] [Google Scholar]
- 29.Klein, A., Steffan-Dewenter, I. & Tscharntke, T. (2003) Proc. R. Soc. London Ser. B 270, 955–961. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Steffan-Dewenter, I. & Tscharntke, T. (1999) Oecologia 121, 432–440. [DOI] [PubMed] [Google Scholar]
- 31.Ricketts, T. H. (2004) Conserv. Biol. 18, 1–10. [Google Scholar]
- 32.Wille, A. & Michener, C. D. (1973) Rev. Biol. Trop. 21, Suppl. 1, 278. [Google Scholar]
- 33.Janzen, D. H. (1983) Costa Rican Natural History (Univ. of Chicago Press, Chicago).
- 34.Kearns, C. A. & Inouye, D. W. (1993) Techniques for Pollination Biologists (Univ. Press of Colorado, Niwot).
- 35.Pagiola, S. (2002) in Selling Forest Environmental Services, eds. Pagiola, S., Bishop, J. & Landell-Mills, N. (Earthscan, London).
- 36.Castro, R. (1999) Valuing the Environmental Service of Permanent Forest Stands to the Global Climate: The Case of Costa Rica (United Nations Development Program, Cambridge, MA).
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}