

Abstract
This is a review of bioactive secondary metabolites isolated from conifer-associated endophytic fungi from 1990–2014. This includes compounds with antimicrobial, anti-inflammatory, anti-proliferative or cytotoxic activity towards human cancer cell lines, and activity against plant pathogens or plant insect pests. Compounds that were originally reported without associated activity were included if other studies ascribed activity to these compounds. Compounds were not included if they were exclusively phytotoxic or if they were isolated from active extracts but were not determined to be the active component of that extract.
Keywords: endophyte, endophytic fungus, secondary metabolites, natural products, bioactivity, conifer, cypress, spruce, pine
In 1991 we began studying the microbial endophytes of the Northwest Pacific yew tree Taxus brevifolia, in our search for a fungus or bacterium that could produce paclitaxel in de novo fashion [1]. At that time, there were few reports describing the chemistry of plant endophytes, although there was a rich literature cataloguing the secondary metabolism of plant pathogenic fungi and bacteria. Phytotoxins, secondary metabolites produced by plant pathogenic microorganisms, have been studied for almost a century as virulence factors and the initiators of diseases in susceptible plants. Three well-known examples are the host-specific toxins produced by three different Cochliobolus species, all of which caused severe blight diseases of economically important crops [2]. C. carbonum (Helminthosporium carbonum) produces host specific HC-toxin, which causes Northern leaf blight of maize and inhibits maize histone deacetylase [2,3]. C. heterostrophus (H. maydis) produces T-toxin which caused Southern Corn Leaf Blight, one of the worst plant disease epidemics in modern history, and which was especially virulent towards maize carrying Texas male sterile cytoplasm [3]. C. victoriae produces victorin which caused a devastating epidemic in the Victoria race of oats that was developed by plant breeders in an effort to produce oats that were resistant to crown rust [3].
Plant endophytes are more subtle, however, rarely causing problems and coexisting with their hosts under most circumstances. Hirsch and Braun provided an inclusive and widely accepted definition of endophytes: “microbes that colonize living, internal tissues of plants without causing any immediate, overt negative effects” [4]. They are generally nonpathogenic in nature, but may produce secondary metabolites that enable them to survive in the competitive world of plant interstitial space without harming their host. Microorganisms in most ecosystems establish and define their ecological niches by their ability to control fellow microbes with only their cell walls or membranes, and chemical arsenals to defend them. But these chemical arsenals have provided many of the important chemotherapeutics used to date. The potent anti-fungal agent griseofulvin is of fungal origin [5] and both the antibiotic streptomycin [6] and the anticancer agent calicheamycin are produced by actinomycetes [7].
Plant endophytes, however, received less attention - until the discovery of a taxol-producing fungus in the bark and needles of the Northwest pacific yew tree. In 2011 we published a review of cytotoxic or anticancer compounds produced by plant endophytes [8]. Over 100 compounds with demonstrated cytotoxicity or anticancer activity had been isolated from endophytic fungi- including several compounds originally isolated from higher plants [8]. Less than 10% of these compounds were isolated from coniferous species [8]. Our own work with the fungal endophytes of conifers has shown them to be rich producers of bioactive secondary metabolites. This review will highlight the bioactive compounds isolated from fungi associated with the bark or needles of various conifers.
Fungal endophytes are a diverse and versatile group of microorganisms that colonize plants in the arctic, antarctic, geothermal soils, deserts, oceans, rainforests, mangrove swamps, and coastal forests [8]. They have been isolated from the root complexes and aerial parts of a wide range of hosts including algae, bryophytes, pteridophytes, gymnosperms and angiosperms [8]. Endophytes have been found in every plant studied to date. There are over 300,000 higher plant species and it can be assumed that each of these species hosts a complex community of endophytic microbes [9]. Endophytes can infect virtually 100% of a host population or a small fraction of the population. Although the relationship between endophytes and their hosts varies from organism to organism, fungal endophytes are an important component of microbial biodiversity. These inconspicuous inhabitants of the tissues of higher plants may represent a rich source of as yet undiscovered and unexplored genera to contribute to fungal diversity and secondary metabolite investigations [9].
This review covers only those secondary metabolites that have been isolated from conifer-associated endophytic fungi and that have demonstrable bioactivity. This includes compounds with antimicrobial, anti-inflammatory, anti-proliferative or cytotoxic activity towards human cancer cell lines, and activity against plant pathogens or plant insect pests. Compounds that were originally reported without associated activity were included if other studies ascribed activity to these compounds. Compounds were not included if they were isolated from active extracts but were not determined to be the active component of that extract.
Anticancer Metabolites Produced by Both Higher Plants and Conifer Endophytes
Diterpenes: Paclitaxel (taxol®)
Our interest in the endophytic fungi of plants began in 1991 as we launched our search for a microorganism that could produce taxol®, (1) a highly functionalized diterpene with compelling anticancer activity, in de novo fashion. (When it was developed commercially by Bristol-Myers Squibb the generic name was changed to ‘paclitaxel’ and taxol® was trademarked.) Few plant secondary metabolites have had such a dramatic effect on cancer chemotherapy as taxol® (1) [10, 11]. It was the prototypical taxane isolated from the bark of the Northwest Pacific yew tree Taxus brevifolia by Wani et al. in 1971 [12, 13]. Paclitaxel showed early promise against a series of human solid tumor xenografts in nude mice including CX-1 colon and MX-1 breast xenografts [10]. Although these early results were encouraging, the real turning point in the paclitaxel saga, however, was the discovery of its unique activity as a promoter of tubulin polymerization [14]. Although other clinically useful drugs were known to act as anti-mitotic agents and inhibitors of tubulin polymerization, paclitaxel was the first compound to exhibit the opposite effect on tubulin – stabilization of its polymer [10,11,14].
Unfortunately as paclitaxel gained medicinal importance because of its unique mode of action and potential as a chemotherapeutic agent, it gained media attention because of problems associated with the supply issue. Early estimates suggested that the population of Northwest Pacific yew trees could not adequately supply the projected demands for paclitaxel. Alternative sources were considered for the compound including total synthesis, semi-synthesis and tissue culture [11]. We took another approach and in 1993 reported the isolation of a fungal endophyte from the needles of T. brevifolia that produced paclitaxel and baccatin III independently of the tree [1, 15, 16]. The fungus had not been previously described and was designated Taxomyces andreanae [17]. We later reported the production of paclitaxel by a second fungus, Penicillium raistrickii which was isolated from the inner bark of a yew tree [18]. The fungus also produced baccatin III (2) and 10-deacetyl baccatin III (3) in liquid culture.
Several other scientists have since reported the isolation of paclitaxel from different endophytic fungi associated not only with Taxus sp. but with unrelated host plants as well. Scientists have reported the production of paclitaxel by Pestalotiopsis microspora isolated from Taxus wallichiana [19] and by a second isolate of P. microspora from bald cypress, Taxodium distichum [20]. Although the focus of this review is the production of bioactive metabolites by coniferous endophytes, it is interesting to note that paclitaxel has also been reported from endophytes associated with various angiosperms including P. pausiceta isolated from Cardiospermum helicacabum [21]; P. terminaliae, an endophytic fungus of Terminalia arjuna [22]; Chaetomella raphigera, also associated with Terminalia arjuna [23]; and Bartalinia robillardoides an endophyte of Aegle marmelos [24]. This is not a comprehensive list of paclitaxel-producing endophytes and more producers are reported every year. Two excellent reviews of paclitaxel producers were published in 2012 [25, 26]. Over 20 genera of endophytic fungi (Alternaria, Aspergillus, Botryodiplodia, Botrytis, Cladosporium, Ectostroma, Fusarium, Metarhizium, Monochaetia, Mucor, Nigrospora, Ozonium, Papulaspora, Periconia, Pestalotia, Pestalotiopsis, Phoma, Phyllosticta, Pithomyces, Taxomyces, Tubercularia) have demonstrated the ability to produce paclitaxel or its analogues [25, 26].
Higher plants that have been reported as hosts of paclitaxel-producing fungi include the coniferous species mentioned above as well as Citrus medica, Ginkgo biloba, Hibiscus rosa-sinensis, Podocarpus sp., Torreya grandifolia, and Wollemia nobilis. As we had hoped in the early 1990’s, one of these paclitaxel-producing fungi could ultimately prove to be an alternative paclitaxel-producing resource. The discovery of a paclitaxel-producing fungus by a yew-associated endophyte spurred the search for other plant-derived compounds of interest that might also be produced by fungi. The focus of this review is the bioactive metabolites of compounds produced by conifer-associated endophytes. Although two of these compounds have been previously reported from higher plants, most of these compounds are uniquely fungal in origin.
Lignans: Podophyllotoxin
The aryltetralin lignan podophyllotoxin (4) is another important natural product that was originally isolated from the higher plants Podophyllum emodi from the Himalayas and Podophyllum peltatum Linnaeus, commonly known as the American mandrake or Mayapple [27]. Both of these plants had a history of medicinal use by indigenous peoples. Podophyllotoxin is a potent antimitotic agent and tubulin polymerase inhibitor. Unfortunately its toxicity and deleterious side effects have limited its clinical use - it is currently used primarily as a topical treatment for genital warts. Some of its semisynthetic derivatives, however, are effective anticancer drugs and include the topoisomerase II inhibitors etoposide, teniposide, and etoposide phosphate [28, 29]. Etoposide is used to treat a wide spectrum of human cancers. It has been in clinical use for more than two decades and remains one of the most highly prescribed anticancer drugs in the world [29].
As was true with paclitaxel, the difficulties involved in the total synthesis of podophyllotoxin and its analogues, as well as the destruction of wild populations of the primary source plant, led many researchers to search for alternative sources of these compounds [30]. Within months of each other two different laboratories reported the isolation of endophytes of Podophyllum capable of producing podophyllotoxin and related analogues [30, 31]. Puri isolated Trametes hirsuta from the dried rhizomes of Podophyllum hexandrum collected from the north-western Himalayan region of Jammu and Kashmir, India [30]. Podophyllotoxin was also purified from two strains of the Podophyllum peltatum-associated endophyte Phialocephala fortinii by Porter and his colleagues [31]. Podophyllotoxin is thus another example of a secondary metabolite produced by both an endophyte and its host.
Podophyllotoxin was also isolated from fungi associated with several non-host plants, in this case different species of junipers. It was isolated from an Alternaria sp. associated with Sabina (Juniperus) vulgaris [32]; from a Fusarium oxysporum associated with the Himalayan medicinal plant Juniperus recurva; [33] and from an Aspergillus fumigatus associated with Juniperus communis L. Horstmann [34].
These discoveries of parallel secondary metabolite production by both higher plants and plant-associated fungi are interesting and provide a potential source of procuring adequate compounds for commercial requirements. Unfortunately the discovery of compounds like paclitaxel or podophyllotoxin and its analogues – or of endophytic fungi that produce them - does not end the need for new effective anticancer agents. Cancer is still the second leading cause of death in the Western world, and even the most effective chemotherapeutic agents rarely constitute a cure. New drugs must be found to help in the fight against the complex diseases referred to as “cancers” [18]. There is also a critical need for new, selective antibacterial and antifungal agents to stem the rising tide of infectious diseases. Increased drug resistance of many common pathogenic bacteria as well as the growing number of immunocompromised individuals mandates an aggressive search for novel antibiotics and fungicidal drugs [18].
Microbes are not always the cause of infectious diseases: their metabolic products can also treat and often cure such infections. Fungi and bacteria establish and define their ecological niches by their ability to kill or control fellow microorganisms with only their cell walls or cell membranes and chemical arsenals to defend them. These chemical arsenals have provided many of the important chemotherapeutics used to date. The potent antifungal agent griseofulvin (5) is of fungal origin [35]; the antibiotics streptomycin and the anticancer agent calicheamycin are produced by actinomycetes [6]; and the anticancer drug taxol is produced by the fungus Taxomyces andreanae [1].
Compounds Isolated Exclusively from Fungi: Polyketides
Griseofulvin
Although this review covers only those bioactive secondary metabolites produced by conifer-associated fungi, many of these compounds were originally isolated from other producing organisms. Such is the case with the important antifungal agent griseofulvin (5), which was first isolated by Raistrick and his colleagues from Penicillium griseo-fulvum Dierckx in 1939 [35], and was subsequently re-isolated from Penicillium janczewskii by Brian and colleagues as the inducer of hyphal “curling-factor” against Botrytis allii, the fungal cause of neck-rot in stored onions [36]. The structure was determined several years later following a series of chemical studies by Grove, MacMillan and colleagues [37, 38]. Compound 5 is currently used as an orally administered antifungal agent in human and animal patients against dermatophytic fungi induced conditions of the skin and nails.
Although it has been isolated primarily from Penicillium sp., griseofulvin was recently isolated for the first time from different strains of conifer-associated Xylaria sp. A Xylaria sp. isolated as a fungal endophyte of the Manchurian fir Abies holophylla produced both griseofulvin and dechlorogriseofulvin (6) in culture. The compounds were evaluated for their in vivo antifungal activity against several important plant pathogenic fungi. Griseofulvin (5) showed high in vivo and in vitro antifungal activity, and effectively controlled the development of rice blast (Magnaporthe grisea), rice sheath blight (Corticium sasaki), wheat leaf rust (Puccinia recondita), and barley powdery mildew (Blumeria graminis f. sp. hordei) [39].
More recently scientists reported the production of 5 by Xylaria sp. associated with both the needles of an eastern white pine (Pinus strobus) and the stems of lowbush blueberry shrubs (Vaccinium angustifolium) growing in close proximity in the Acadian forests of New Brunswick and Nova Scotia, Canada [40]. This study also highlighted the complexity of endophyte-host lifecycles and suggested an interesting connection between conifers and understory plant ecologies. This study highlights the complexity of endophyte-host lifecycles and points to the existence of a pine-blueberry ecotype [40].
Griseofulvin (5) was also isolated in our lab from a Penicillium sp. associated with the Northwest Pacific yew tree, Taxus brevifolia [15, 18]. In our study of taxol-producing endophytes we isolated over 300 different fungi from yew trees growing throughout the Pacific Northwest. After screening these fungi for taxol production we isolated other bioactive metabolites. One of the taxol-producing fungi, Penicillium raistrickii and three other endophytic Penicillium sp., produced other interesting secondary metabolites – some novel, some previously isolated from other fungi. These compounds will be described in this review.
Brefeldin A
Brefeldin A (7) is a compound with demonstrated antifungal, antiviral and anticancer activities, and that also acts as a protein-transport inhibitor [41]. Although it has been isolated from several fungal species including Curvularia, Alternaria, Ascochyta, Phyllosticta, Penicillium, and Cercospora [42], 7 was first reported as a secondary metabolite of Eupenicillium brefeldianum [43]. It is identical to decumbin, which was first reported in 1958 as a crystalline dihydroxy-α,β-unsaturated lactone produced by a Penicillium associated with spoiled corn [44]. It is also identical to cyanein, isolated from P. cyaneum [45] and ascotoxin, isolated from Ascochyta imperfecta [46].
One study showed that the incorporation of [1-14C]-, [2-14C]-malonate, and [1-14C]-acetate incorporation into brefeldin A (7) confirms its biosynthesis by the normal polyketide pathway [47]. Another study showed direct incorporation of [9-14C] palmitate into brefeldin A suggesting a biosynthetic pathway typical of the prostaglandins [48]. Recent studies have shown that brefeldin A biosynthesis is catalyzed by a specific highly reducing polyketide synthase (HRPKS) termed Bref-PKS [41]. This study demonstrated that the intrinsic programming rules of these enzymes determine the overall number of iterative cycles and the ultimate length of the polyketide chain. Bref-PKS demonstrated an NADPH-dependent reductive tailoring specificity that led to the synthesis of four different octaketide products with varying degrees of reduction. Although brefeldin A (7) is an octaketide, Bref-PKS is actually a nonaketide synthase in the absence of an associated thiohydrolase Bref-TH [41].
Brefeldin A (7) was isolated from two different endophytic fungi: Paecilomyces sp., which was isolated from the tissues of both Chinese Taxus mairei and Torreya grandis, and Aspergillus clavatus which was isolated from the phloem of T. grandis [42]. The authors tested 7 and compared the activity to that of paclitaxel (1) against the human cancer cell lines HL-60, KB, HeLa, MCF-7 and Spc-A-1 cell lines. The IC50 values of 7 were between 1.0 and 10.0 ng/mL with the greatest potency against human lung cancer cell line Spc-A-1. The IC50 values of 1 ranged between 0.16 and 5.0 ng/mL with greatest potency against KB, the keratin-forming subline of HeLa (cervical cancer) [42].
Sequoiatones
The two novel polyketides sequoiatones A (8) and B (9) were isolated from the endophyte Aspergillus parasiticus associated with the bark of Sequoia sempervirens. The compounds showed moderate and selective inhibition of human tumor cells, with greatest efficacy against breast cancer cell lines. Most of the GI50’s were between 4 – 10 μM with LC50’s > 100 μM [49]. Sequoiatones C–F (10–13) were isolated from the same fungus and were active in the brine shrimp lethality assay. The LD50 of 10–13 were 260, 1300, 640, and 260 μM, respectively [49b].
Sequoiamonascines
The same sequoia-associated A. parasiticus also yielded sequoiamonascins A–D (14–17) based on brine shrimp lethality. Compounds 14 and 15 were tested by the NCI Drug Therapeutic Program in their human cell line screen. The two compounds exhibited cytotoxic activity against MCF7 (breast), NCI-H460 (lung), and SF-268 (CNS) cell lines [50]. At that time, the NCI reported the activity in terms of percent of growth of treated cells compared to untreated cells. Values below 32% were considered active. At concentrations of 10 μM compound 14 allowed 1%, 1% and 2%, (percent of growth) respectively for each treated cell type, while compound 15 allowed 19%, 4% and 15% (percent of growth) of treated cancer cells respectively [50]. In the 60-human cell line assay, compound 14 had a median log GI50 of −5.00, below the potency threshold established by NCI to warrant further study. Compound 14 showed selective activity towards all six leukemia cell lines, one breast cancer cell line, and two melanoma cell lines, with median values approaching log GI50 of −6.00 [50].
Torreyanic acid
Torreyanic acid (18) is an unusual dimeric quinone isolated from Pestalotiopsis microspora, an endophyte of Torreya taxifolia [51]. In general, torreyanic acid was found to be 5–10 times more potent against cell lines that are sensitive to protein kinase C (PKC) agonists and it was suggested that it causes cell death by apoptosis. IC50 values for torreyanic acid were between 3.5 μg/mL for human colorectal neuroendocrine cell carcinoma (NEC) to 45 μg/mL for human adenocarcinomic alveolar basal epithelial cells (A549), with a mean value of 9.4 μg/mL for 25 different cell lines. Torreyanic acid also showed G1 arrest of G0 synchronized cells at the 1–5 μg/mL level depending on the cell line [51].
Nodulisporins
Nodulosporins A–C (19–21) were isolated from a fungal endophyte of the endangered plant Juniperus cedrus (Canary Island Juniper) identified as Nodulisporium sp. (Xylariaceae) [52]. The three compounds exhibited antifungal activity against Microbotryium violaceum. Three related compounds, nodulisporins D–F, were isolated from a Nodulisporium sp. isolated from an angiosperm shrub, Erica arborea, from the Canary Island of Gomera [53]. Nodulisporins D–F showed antibacterial activity against Bacillus megaterium, antifungal activity against Microbotryium violaceum (anther smut fungus), and anti-algal activity against Chlorella fusca, using agar diffusion assays.
Phomopsolides
A Penicillium sp. isolated from Taxus brevifolia produced the phomopsolides, a series of polyketides with a tiglic acid ester side chain. Phomopsolide A (22) and B (23) were originally isolated from Phomopsis oblonga, a fungus which apparently conferred some protection against elm bark beetle infestations [54]. In this study, several compounds, including the two phomopsolides, acted as boring/feeding deterrents to destructive Scolytid beetles. The yew-associated Penicillium sp. also yielded phomopsolides A and B, as well as the three new phomopsolides C–E (24–26), following antibacterial guided isolation protocols against Staphylococcus aureus and Vibrio harveyii rather than antifeeding activity [18, 55]. Phompsolides A–C had comparable activity to tetracycline, penicillin G and streptomycin against Staphylococcus aureus [55].
Rugulosin
Rugulosin (27) is a bright yellow bis-anthraquinoid pigment isolated from cultures of Hormonema dematioides, an endophytic fungus associated with Canadian balsam fir trees. It was isolated using bioassay-guided fractionation for inhibition of spruce budworm growth [56]. Conifer needles infected with the endophyte were associated with reduced weight gain of spruce budworm larvae when used as feed. Rugulosin has also been reported to be produced by various other fungal species including Penicillium [57]. Rugulosin is cytotoxic to both prokaryotes and eukaryotes and causes fatty degeneration, liver cell necrosis, and to a lesser extent, hepatocarcinogenesis to mice and rats [57].
Methyl (2Z,4E)-6(acetyloxy)-5-formyl-7-oxoocta-2,4-dienoate
The extracts of five foliar fungal endophytes isolated from Pinus strobus (eastern white pine) identified as Lophodermium sp. exhibited strong antifungal activity in disc diffusion assays and were selected for further study [58]. These five strains produced different combinations and concentrations of the aliphatic polyketide methyl (2Z,4E)-6(acetyloxy)-5-formyl-7-oxoocta-2,4-dienoate (28), the known macrolide pyrenophorol (29), and three related sesquiterpenes which will be described in a later section. Compounds 28 and 29 exhibited antifungal activity against the rust Microbotryum violaceum and Saccharomyces cerevisae [58].
Terpenes
5-(hydroxymethyl)-2-(20-trimethyltetrahydro-2H-pyran-2-yl)-phenol
The extracts of five foliar fungal Lophodermium sp. endophytes of Pinus stobus mentioned in the previous section also yielded three sesquiterpenes. One of these compounds, the sesquiterpene 5-(hydroxylmethyl)-2-(20-trimethyltetrahydro-2H-pyran-2-yl)phenol (30), also exhibited strong antifungal activity in the disc diffusion assay against the anther rust Microbotryum violaceum and the yeast Saccharomyces cerevisae [58].
Phomadecalins
Microdiplodia sp. KS 75-1 from the stem of Pinus sp produced the novel eremophilane sesquiterpenes 8α-acetoxyphomadecalin C (31) and phomadecalin E (32) [59]. Compounds 31 and 32, which were isolated for the first time from this fungus, showed moderate activity against Pseudomonas aeruginosa ATCC 15442, a standard strain used to evaluate bactericidal disinfectants, with inhibition zones of 12 mm and 11 mm in diameter at a test concentration of 200 μg/disk [59].
Xylarenones
Xylarenones A (33) and B (34), and xylarenic acid (35) were isolated from the endophytic fungus Xylaria sp. NCY2, obtained from Torreya jackii CHUN. These compounds were tested in vitro in antitumor and antimicrobial assays and exhibited moderate antitumor activities against HeLa cells (cervical cancer cell line) [60].
Tuberculariols
Tubercularia sp. TF5, an endophytic fungal strain isolated from the medicinal plant Taxus mairei, was submitted to protoplast mutations and genomic shuffling with UV and N-methyl-nitro-N-nitrosoguanidine (NTG). Two of the resultant mutant strains, M-741 and G-444, were selected for metabolite isolation and determination. Three new eremophilane-type sesquiterpenoids, tuberculariols A–C (36–38) were isolated from mutant strain M-741 and exhibited weak inhibitory activity against HeLa calls (cervical cancer cell line) [61].
Enfumafungin
Enfumafungin (39) is a new antifungal triterpenoid glycoside produced by Hormonema sp. (ATCC 74360), a fungal endophyte of Juniperus communis [62]. It exhibited highly potent antifungal activity (in vitro) against Candida albicans, C. tropicalis, Aspergillus fumigatus and Saccharomyces cerevisiae, and moderate efficacy in an in vivo mouse model of disseminated candidiasis [62]. In further studies, 39 specifically inhibited glucan synthesis in whole cells and in (1,3)-b-D-glucan synthase assays, altered the morphologies of yeasts and molds, and produced a unique response in Saccharomyces cerevisiae strains with point mutations in FKS1, the gene which encodes the large subunit of glucan synthase. Although enfumafungin 39 was active against several strains of fungi, it was particularly active against Aspergillus fumigatus and A. flavus (Table 1) [63].
Table 1.
Antifungal activity of enfumafungin reported as MIC (minimum inhibitory concentrations) in μg/mL
| Test Organism | MIC |
|---|---|
| Candida albicans MY1055 | 0.25 |
| Candida albicans MY2301 | 0.50 |
| Candida glabrata CLY574 | 1.00 |
| Candida glabrata MY1381 | 1.00 |
| Candida guilliermondii CLY308 | 1.00 |
| Candida guilliermondii CLY346 | 2.00 |
| Candida krusei CLY549 | 2.00 |
| Candida lusitaniae MY1396 | 1.00 |
| Candida parapsilosis ATCC 22019 | 0.50 |
| Candida tropicalis MY1012 | 0.50 |
| Aspergillus fumigatus MF5668b | ≤0.03 |
| Aspergillus flavus MF383 | ≤0.03 |
| Cryptococcus neoformans MY2061 | >64 |
| Cryptococcus neoformans MY2062 | >64 |
| Staphylococcus aureus MB2865 | >64 |
Periconicins
The fusicoccane diterpenes periconicins A (40) and B (41) were isolated by bioassay-guided fractionation from cultures of the endophytic fungus Periconia sp. associated with the inner bark of Taxus cuspidate [64]. Periconicin A (40) exhibited more potent antibiotic activity than B (41) against Bacillus subtilis, Klebsiella pneumoniae, and the opportunistic human pathogen Proteus vulgaris (ATCC 3851). Compound 40 also exhibited more potent antifungal activity against the human pathogens Candida albicans, Trichophyton mentagrophytes, the causative agent of cutaneous infections, and T. rubrum, the causative agent of jock itch, athlete’s foot and ringworm [65].
Heptelidic acid (konigic acid)
Heptelidic acid (42) and hydroheptelidic (43) acid are sesquiterpenes that were isolated from Phyllosticta sp., an endophytic fungus inhabiting the needles of Abies balsamea (balsam fir tree) from New Brunswick, Canada [66]. In this and many other regions, this tree species is affected by defoliating larvae of the spruce budworm (Choristoneura fumiferana). The endophyte-derived compounds were shown to be toxic to these larvae which have caused hundreds of millions of dollars in losses to the Canadian forestry sector.
Heptelidic acid (42) was originally isolated from a strain of Trichoderma virens. Studies have found that it is a potent and selective inhibitor of glyceraldehyde 3-phosphate dehydrogenase (GAPDH) and selectively kills high-glycolytic cancer cells through glucose-dependent active ATP deprivation [67]. Compound 42 selectively and competitively inhibited the activities of mammalian DNA polymerases (pol) beta and lambda, as well as terminal deoxynucleotidyl transferase (TdT) in family X of pol. It did not, however, influence the activities of other families (A-, B-, and Y-) of the mammalian pols tested, and showed no effect on the activities of plant pol alpha, fish pol delta, prokaryotic pol, or the other DNA metabolic enzymes tested.
Heptelidic acid (42) also inhibited etoposide-induced apoptosis in leukemia cells via down-regulation of specific caspases. Human leukemia U937 cells were treated with etoposide which induced apoptosis and its classic biological hallmarks which inlude DNA fragmentation and caspase-3 activation. [69].
Finally, 42 was studied along with six other compounds in an attempt to find new potent anti-malarial compounds [70]. It was found to inhibit the growth of indicator strains of bacteria in a heme-dependent manner typical of artemisinin. The potent growth inhibition of the test bacteria by 42 in the presence of hemin, and the loss of inhibition upon supplementation with the radical scavenger tocopherol, suggested that the inhibitory activity involved radical generation. Heptelidic acid (42) also exhibited moderate in vitro toxicity towards Plasmodium falciparum [70].
Isopimaratriene-dione and -trione analogues
Findlay’s lab focused on the discovery of insect toxins from endophytes of woody plants. Two novel diterpenoid toxins, 9α-hydroxy-l,8-(14),15-isopimaratriene-3,7,1l-trione (44) and 9α-hydroxy-1,8(14),15-isopimaratrien-3,11-dione (45) were isolated from the culture filtrates of an unidentified endophyte obtained from a needle of a balsam fir. Both 44 and 45 displayed comparable toxicity to spruce budworm cells and to larvae (Choristoneura fumiferana) in a feeding bioassay. The toxicity to budworm larvae appears to be substantially lower than that of azadirachtin [71].
Ramulosin and Mellein Analogues
In a continuing study directed at the discovery of insect toxins, Findlay et al. isolated several isocoumarin-producing endophyte strains, identified as Canoplea elegantula (Cooke) M.B. Ellis, from black spruce (Picea muriana BSP) needles obtained from trees in Parc Ashuapmushuan and Parc des Laurentides, Quebec, Canada. The culture filtrates of these strains showed toxicity to spruce budworm (Choristoneura fumifeana Clem.) cells and larvae [72]. In each case several isocoumarins related to ramulosin or mellein were isolated and bioassayed but only two exhibited cytotoxicity. Compound 46 was a new isocoumarin and was identified as (lS,3R,4S,5S)-4-hydroxy-3-methyl-2-oxobicyclo-[3.3.1]non-6-one, which is essentially a completely reduced mullein. Compound 47 was identified as (3R,4aS,8S,8aR)-8-hydroxy-3-methyl-3,4,4a,5,6,7,8,8a-octahydro-1H-2-benzopyran-1-one. Both 46 and 47 were toxic to spruce budworm larvae, and 46 was also toxic to budworm cells [72].
Mixed Biosynthetic Pathways
Mycophenolic acid
Mycophenolic acid is an amazing fungal metabolite with an acetate-mevalonate ring system and a terpenoid tail. Bioassay-guided fractionation (anti-Candida activity) of the organic extract of a liquid culture of a Taxus brevifolia associated Penicillium brevicompactum yielded mycophenolic acid (48) [18]. It was first isolated, however, from a Penicillium sp. by Gosio in 1896 [73] and was re-isolated in 1932 as the antifungal component of Penicillium brevicompactum [74]. Since that time several species of Penicillium, including P. stoloniferum, P. scabrum, P. nagemi, P. szaferi, P. patris-mei, P. griscobrunneum, and P. viridicatum, were reported to produce 48 [73]. This amazing fungal metabolite has been reported to possess the following diverse biological properties: antiviral [75], antifungal [76], antibacterial [76], antitumor [77], immunosuppressive [78], and anti-psoriatic [79].
Mycophenolic acid is currently used as an immunosuppressive drug used to prevent organ rejection and is marketed under the trade name Cell Cept (Roche) and the mycophenolate sodium salt as Myfortic (Novartis). The morpholinoethyl ester of mycophenolic acid (Mycophenolate mofetil) is increasingly being used as a corticosteroid-sparing agent in immunosuppressive regimens.
Much current attention has focused on the use of 48 in the treatment of dengue fever, a mosquito-borne viral disease of global importance with no available antiviral therapy [81]. Studies have shown that 48 could inhibit dengue virus (DV) antigen expression, RNA replication, and virus production. Pharmacological concentrations of 48 effectively blocked DV infection, decreasing the percentage of infected cells by 99% and the levels of secreted virus by up to a million-fold [81]. Results were reproduced with four hepatoma cell lines and different flaviviruses, including a recent West Nile virus isolate. Quantitative RT-PCR demonstrated that 48 prevented the accumulation of viral positive- and negative-strand RNA as the infection proceeded. It was concluded that 48 inhibited flavivirus infection by preventing synthesis and accumulation of viral RNA [81].
Pseurotin A
An unusual heterospirocyclic compound, pseurotin A (49), was isolated from Penicillium raistrickii associated with the inner bark of Taxus brevifolia [18] and was previously isolated from Pseudeurotium ovalis [82].
Pseurotin A (49) was also isolated from cultures of Penicillium janczewskii K.M. Zalessky, an endophytic fungus from the phloem of the Chilean plum yew (gymnosperm) Prumnopitys andina [83]. When grown in liquid yeast extract-malt extract-glucose broth, the fungus produced two main secondary metabolites: 49 and cycloaspeptide A, which will be described in the next section. Pseurotin A (49) presented low cytotoxicity towards human lung fibroblasts. It exhibited moderate antibacterial activity against the phytopathogenic bacteria Erwinia carotovora and Pseudomonas syringae, with IC50 values of 220 and 112 μg/mL, respectively [83].
Further studies found that 49 induced cell differentiation of PC12 neuronal cells, suggesting that it could be used as a tool for studying the mechanism of neurite formation [84]. It also suppressed the production of immunoglobulin E (IgE) suggesting its use as an interventional tool for studies of IgE-mediated systemic allergic response mechanisms [84].
Tauranin
Tauranin (50) was originally isolated from Oospora aurantia [85]. Compound 50 and a series of related sesquiterpene quinones were isolated from Phyllosticta spinarum, a fungal endophyte of the Sonoran desert cypress Platycladus orientalis [86]. Only 50 was active against several human cancer cell lines. These included NCI-H460 (non-small cell lung cancer), MCF-7 (breast cancer), SF-268 (CNS cancer-glioma), PC-3M (metastatic prostate cancer) and MIA Pa Ca-2 (pancreatic carcinoma) at values of 4.3, 1.5, 1.8, 3.5, and 2.8 μM, respectively. Taurinin (50) also inhibited the incorporation of acetate-1-14C into cholesterol. Three related compounds, 3-ketotauranin (51), 3-hydroxytauranin (52), and 12-hydroxytauranin (53), were inactive at all test concentrations despite the similarities among these closely related analogues [86].
Peptides
Leucinostatin A
Leucinostatin A (54) was isolated almost forty years ago from cultures of Penicillium lilacum [87]. It has received much attention over the years because of its potent biological activity against several different cancer cell lines. It inhibits prostate cancer cell growth through the reduction of insulin-like growth factor-I expression in prostate stromal cells [88]. Acremonium sp. isolated from Taxus baccata was also shown to produce 54 when grown in liquid culture [89]. The fungal endophyte also produced the new compound leucinostatin A β-di-0-glucoside 55, a glycosylated analogue of 54 which had a LD50 of > 25 nM against breast cancer cell line BT-20, compared to 54 which had a LD50 of 2 nM [89]. Compound 55 was also antifungal against the oomycete Pythium ultimum, an important plant pathogen that causes damping-off and root rot diseases [89].
Echinocandin
Echinocandin (56) was purified from cultures of the endophytic fungi Cryptosporiopsis sp. and Pezicula sp., associated with Pinus sylvestris (Scots pine) and Fagus sylvatica (European beech), respectively [90]. The compound showed antifungal activity against Candida albicans and Saccharomyces cerevisiae [90]. Mechanistically, echinocandins were found to inhibit the synthesis of cell wall glucans by inhibiting glucan synthase leading to cell lysis [91].
Cycloaspeptide A
The same fungus that produced paclitaxel, pseurotin, benzomalvin B, a Penicillium raistrickii associated with Taxus brevifolia, also produced cycloaspeptide A (57) [18]. The peptide was previously isolated from Aspergillus sp. NE-45 [92]. Cycloaspeptide A (57) was also isolated from the psychrotolerant fungus Penicillium algidum. It exhibited moderate activity (IC50 3.5 μg/mL) against Plasmodium falciparum and low level cytotoxicity towards human lung fibroblasts [93].
Nitrogen Containing Compounds
Benzomalvin C
A very interesting compound isolated from our yew-associated Penicillium raistrickii was the benzodiazepine benzomalvin C (58) [18]. It was originally isolated, along with benzomalvins A (59) and B (60), from a Penicillium sp. isolate SC67 by the Sterling Winthrop Pharmaceuticals Research Division in their search for substance P inhibitors [94]. Substance P is a neurokinin peptide whose NK1 receptor is involved in many physiological actions, particularly in pain transmission and activation of neurogenic inflammation. It induces a number of physiological responses including salivation, smooth muscle contraction and vasodilation. A selective substance P antagonist could prove to be a novel analgesic or anti-inflammatory agent. Compound 59 was the most active substance P inhibitor, while 58 and 60 were moderately active [94].
Fiscalin B
Fiscalin B (61) was isolated from the mycelial extract of Penicillium raistrickii associated with Taxus brevifolia [18]. It was spectroscopically identical to a compound originally isolated from Neosartorya fischeri by Sterling-Winthrop labs in their continuing search for substance P inhibitors [95]. Two related compounds, fiscalin A (62) and C (63) were also found in N. fischeri. These compounds inhibited the binding of radiolabeled substance P ligand to the human neurokinin (NK-1) receptor, with Ki values of 174, 57, and 68 μM, respectively. The fiscalins are structurally similar to the tremorgenic tryptoquivaline mycotoxins produced by Aspergillus clavatus and A. fumigatus [95].
Oxaline
A Penicillium sp. isolated from Taxus brevifolia synthesized several different classes of compound including the previously described phomopsolides 22–26 and an alkaloid that was spectroscopically identical to oxaline (64) [18]. Oxaline (64) was first isolated as a new alkaloid from a toxigenic strain of P. oxalicum M-555 [98].
Oxaline (64) was found to inhibit cell proliferation and to induce cell cycle arrest at the G2/M phase in Jurkat cells. It also inhibited tubulin polymerization resulting in cell cycle arrest at the M phase [99].
Roquefortine C
The Taxus brevifolia-associated Penicillium sp. that produced oxaline (64) also yielded the alkaloid roquefortine C which was isolated based on its brine shrimp lethality (65) [18], which was first isolated as a neurotoxic component of P. roqueforti [100] and P. crustosum [101]. Steyn and Vleggaar proved that roquefortine was an intermediate in oxaline biosynthesis and were able to induce production of radiolabeled oxaline in P. oxalicum by adding radiolabeled roquefortine to the fungal medium [102].
Conclusion
This review highlights the importance of endophytic fungi – those hidden, subtle inhabitants of the interstitial spaces in plants – as a source of secondary metabolites with promising biological activity. Even when the scope is narrowed to include only those compounds found in the fungal endophytes of coniferous plants, the catalogue of compounds is diverse both in terms of structural complexity and activity. There is a constant, critical need for new pharmaceutical agents to combat cancers, viral infections, infectious diseases, and autoimmune disorders. There is also a growing need to fight insect-borne diseases of both animals and plants as climatological changes provide conditions conducive to more intensive outbreaks of these events. The fight against any disease is a dynamic equilibrium between advances in chemotherapy and natural selection in infectious or invasive agents. If the scientific community is to maintain parity in this never-ending struggle, then new sources of novel, bioactive chemotherapeutic agents must be found. Endophytic microorganisms may prove such a source, as these fungi and bacteria wage constant chemical warfare with other microbes to establish an ecological niche. Secondary metabolites are on of their essential means of communication and defense, and these metabolites are readily available for discovery.
Endophytic fungi are proving to be prolific producers of bioactive compounds from many different chemical classes. If the focus is narrowed to anticancer compounds alone, over 100 compounds with significant cytotoxicity were reported from endophytic fungi from 2000 – 2010. Approximately 30% of these compounds were derived from conifer-associated endophytes. It is interesting to note from 1990–1995, only a single novel anticancer agent was reported from endophytic fungi. The most exciting discovery in this area of research during this period was a conifer associated fungus capable of producing paclitaxel, a new anticancer agent associated with a higher plant host, the Northwest Pacific yew tree. This discovery spurred interest not only in fungal endophytes as a source of novel anticancer agents, but also in endophytes as an alternative source of valuable higher-plant metabolites. A fungal source of any desirable metabolite is of particular value, as fungal fermentation provides a virtually inexhaustible source of desired metabolites. Novel compounds or previously isolated compounds are readily available and accessible to whatever specific biological screens researchers use for isolation and evaluation. As our understanding of the mechanisms associated with the onset and development of different diseases increases, our ability to use this knowledge to select for ever more potent and selective compounds should increase commensurately. Endophytic fungi will continue to provide a fertile arena for these quests.
Figure 1.

Structures of compounds 1–3.
Figure 2.
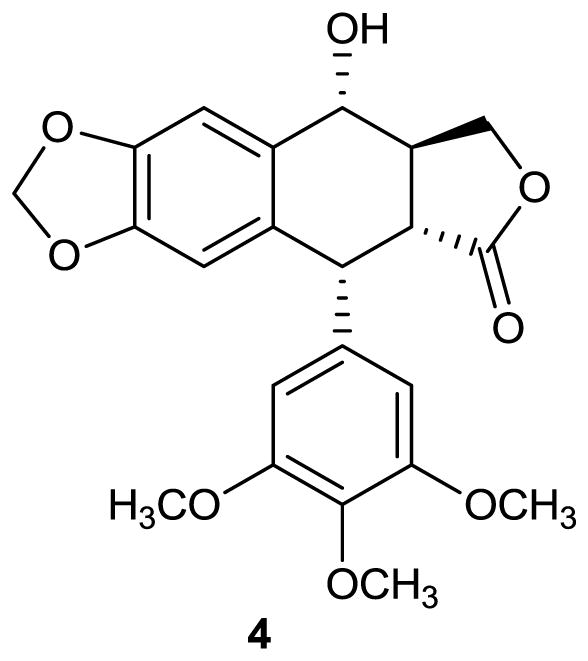
Structure of compound 4.
Figure 3.
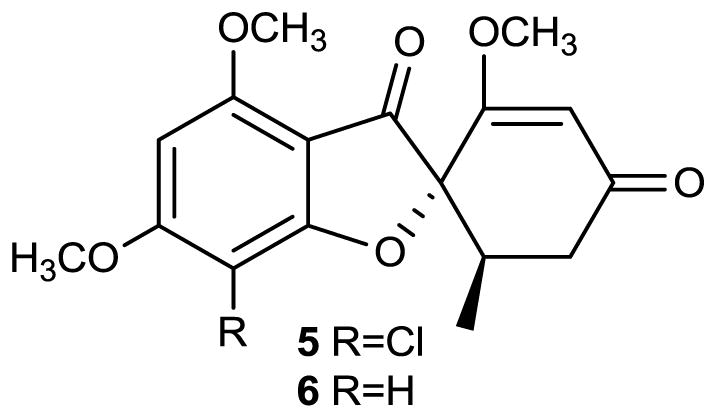
Structure of compounds 5–6.
Figure 4.
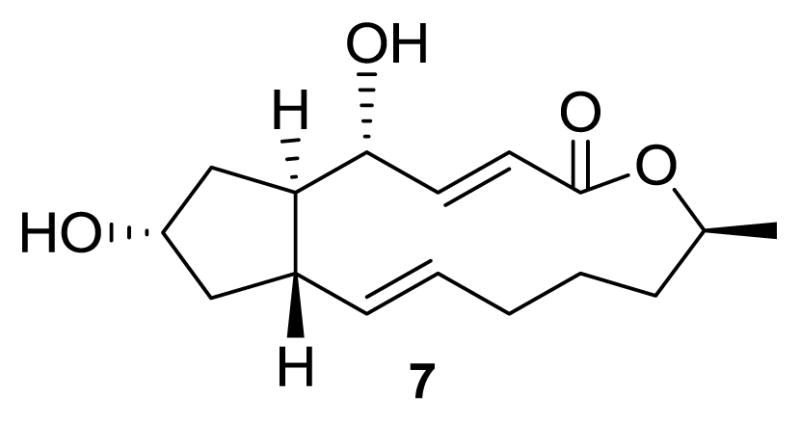
Structure of compound 7.
Figure 5.

Structures of compounds 8–13.
Figure 6.

Structures of compounds 14–17.
Figure 7.
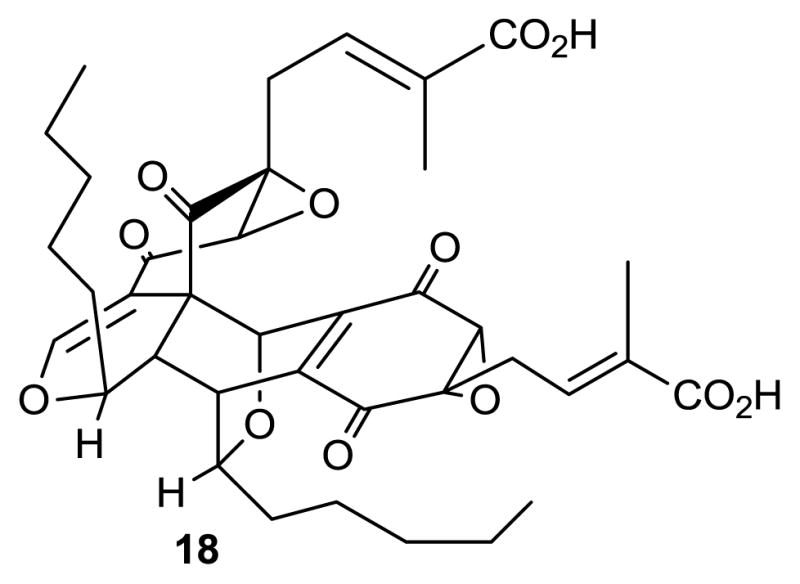
Structure of compound 18.
Figure 8.

Structures of compounds 19–21.
Figure 9.

Structures of compounds 22–26.
Figure 10.

Structure of compound 27.
Figure 11.

Structures of compounds 28–30.
Figure 12.

Structures of compounds 31 and 32.
Figure 13.
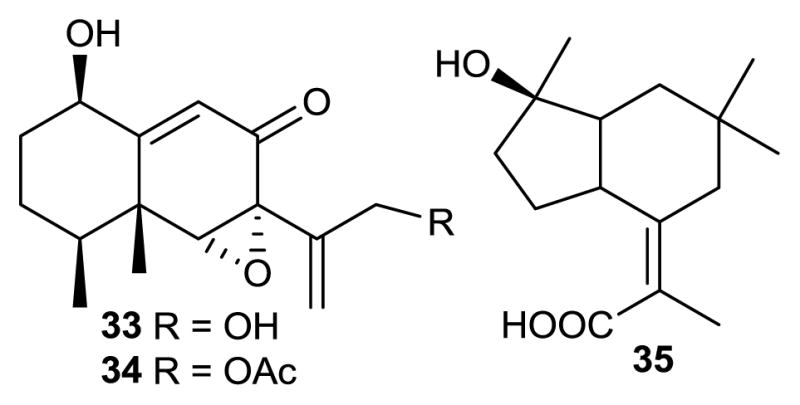
Structures of compounds 33–35.
Figure 14.

Structures of compounds 36–38.
Figure 15.

Structure of compound 39.
Figure 16.

Structures of compounds 40 and 41.
Figure 17.

Structures of compounds 42–43.
Figure 18.

Structures of compounds 44–47.
Figure 19.

Structures of compound 48 and 49.
Figure 20.
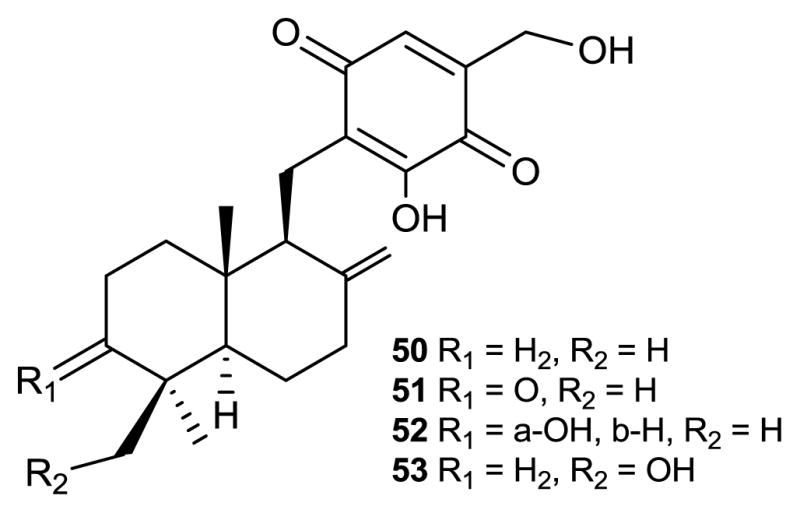
Structures of compounds 50–53.
Figure 21.

Structures of compounds 54 and 55
Figure 22.

Structure of compound 56.
Figure 23.

Structure of compound 57.
Figure 24.

Structures of compounds 58–60.
Figure 25.

Structures of compounds 61–63.
Figure 26.

Structures of 64–65.
Acknowledgments
The Stierles acknowledge NSF grant #CHE-9977213 for acquisition of an NMR spectrometer; NIH grants # P20 RR-16455-02; # P20RR16455-06; 1R01CA139159-01; and 8P20GM103546-02.
References
- 1.Stierle A, Strobel G, Stierle D. Taxol and taxane production by Taxomyces andreanae an endophytic fungus of Pacific yew. Science. 1993;260:214–216. doi: 10.1126/science.8097061. [DOI] [PubMed] [Google Scholar]
- 2.Kimura M, Anzai H, Yamaguchi I. Microbial toxins in plant-pathogen interactions: Biosynthesis, resistance mechanisms, and significance. Journal of General and Applied Microbiology. 2001;47:149–160. doi: 10.2323/jgam.47.149. [DOI] [PubMed] [Google Scholar]
- 3.http://masters.agron.iastate.edu/classes/514/lesson06/6.2.1.html
- 4.Hirsch GU, Braun U. Communities of parasitic microfungi. In: Winterhoff W, editor. Handbook of Vegetation Science: Fungi in Vegetation Science. Vol. 19. Kluwer Academic; Dordrecht: 1992. pp. 225–250. [Google Scholar]
- 5.Grove JF, MacMillan J, Mulholland TPC, Rogers MAT. 762. Griseofulvin. Part IV. Structure. Journal of the Chemical Society. 1952:3977–3987. Resumed. [Google Scholar]
- 6.Schatz A, Bugie E, Waksman S. Streptomycin: a substance exhibiting antibiotic activity against Gram-positive and Gram-negative bacteria. Proceedings of the Society for Experimental and Biological Medicine. 1944;55:66–69. [Google Scholar]
- 7.Maiese WM, Lechevalier MP, Lechevalier HA, Korshalla J, Kuck N, Fantini A, Wildey MJ, Thomas J, Greenstein M. Calicheamicins, a novel family of antitumor antibiotics: taxonomy, fermentation and biological properties. Journal of Antibiotics. 1989;42:558–63. doi: 10.7164/antibiotics.42.558. [DOI] [PubMed] [Google Scholar]
- 8.Kharwar RN, Mishra A, Gond SK, Stierle A, Stierle D. Anticancer compounds derived from fungal endophytes: their importance and future challenges. Natural Product Reports. 2011;28:1208–1228. doi: 10.1039/c1np00008j. [DOI] [PubMed] [Google Scholar]
- 9.Saikkonen K, Faeth SH, Helander M, Sullivan TJ. Fungal Endophytes: A continuum of interactions with host plants. Annual Review of Ecology and Systematics. 1998;29:319–343. [Google Scholar]
- 10.Kingston DGI. Taxol and its analogues. In: Cragg GM, Kingston DGI, Newman DJ, editors. Anticancer Agents from Natural Products. Taylor and Francis Group, LLC, CRC Press; Boca Raton, Florida: 2005. pp. 89–120. [Google Scholar]
- 11.Cragg GM, Snader KM. Plants as a source of anti-cancer agents. Journal of Ethnopharmacology. 2005;100:72–79. doi: 10.1016/j.jep.2005.05.011. [DOI] [PubMed] [Google Scholar]
- 12.Wani MC, Taylor HL, Wall ME, Coggon P, McPhail AT. Plant Antitumor Agents VI. The isolation and structure of taxol, a novel antileukemic and antitumor agent from Taxus brevifolia. Journal of the American Chemical Society. 1971;93:2325–2327. doi: 10.1021/ja00738a045. [DOI] [PubMed] [Google Scholar]
- 13.Wall ME, Wani MC. Camptothecin and taxol: discovery to clinic--thirteenth Bruce F. Cain Memorial Award Lecture. Cancer Research. 1995;55:753–60. [PubMed] [Google Scholar]
- 14.Schiff PB, Fant J, Horwitz S. Promotion of microtubule assembly in vitro by taxol. Nature. 1979;277:665–667. doi: 10.1038/277665a0. [DOI] [PubMed] [Google Scholar]
- 15.Stierle A, Stierle D, Strobel G, Bignami G, Grothaus P. Bioactive metabolites of the endophytic fungi of Pacific yew, Taxus brevifolia: paclitaxel, taxanes, and other bioactve compounds. In: Georg GI, Chen TT, Ojima I, Vyas DM, editors. Taxane Anticancer Agents: Basic Science and Current Status. Washington, DC: 1995. pp. 81–97. ACS Symposium Series 583. [Google Scholar]
- 16.Stierle A, Stierle D, Strobel G, Grothaus P, Bignami G. The search for a taxol-producing microorganism among the endophytic fungi of the Pacific yew, Taxus brevifolia. Journal of Natural Products. 1995;58:1315–1324. doi: 10.1021/np50123a002. [DOI] [PubMed] [Google Scholar]
- 17.Strobel GA, Stierle A, Stierle D. Taxomyces andreanae, a proposed new taxon for a bulbilliferous hyphomycete associated with Pacific yew. Mycotaxon. 1993;47:71–78. [Google Scholar]
- 18.Stierle AA, Stierle DB. Bioactive compounds from four endophytic Penicillium sp. isolated from the Northwest Pacific yew tree. In: Atta-Ur-Rahman, editor. Bioactive Natural Products. Vol. 24. Elsevier Science Publishers; Amsterdam: 2000. pp. 933–978. [Google Scholar]
- 19.Strobel G, Yang X, Sears J, Kramer R, Sidhu RS, Hess WM. Taxol from Pestalotiopsis microspora, an endophytic fungus of Taxus wallachiana. Microbiology. 1996;142:435–440. doi: 10.1099/13500872-142-2-435. [DOI] [PubMed] [Google Scholar]
- 20.Li JY, Strobel G, Sidhu R, Hess WM, Ford EJ. Endophytic taxol-producing fungi from bald cypress, Taxodium distichum. Microbiology. 1996;142:2223–2226. doi: 10.1099/13500872-142-8-2223. [DOI] [PubMed] [Google Scholar]
- 21.Gangadevi V, Murugan M, Muthumary J. Taxol determination from Pestalotiopsis pauciseta, a fungal endophyte of a medicinal plant. Chinese Journal of Biotechnology. 2008;24:1433–1438. doi: 10.1016/s1872-2075(08)60065-5. [DOI] [PubMed] [Google Scholar]
- 22.Gangadevi V, Muthumary J. Taxol production by Pestalotiopsis terminaliae, an endophytic fungus of Terminalia arjuna (arjun tree) Biotechnology and Applied Biochemistry. 2009;158:675–684. doi: 10.1042/BA20070243. [DOI] [PubMed] [Google Scholar]
- 23.Gangadevi V, Muthumary J. A novel endophytic taxol-producing fungus Chaetomella raphigera isolated from a medicinal plant, Terminalia arjuna. Biotechnology and Applied Biochemistry. 2009;158:675–684. doi: 10.1007/s12010-009-8532-0. [DOI] [PubMed] [Google Scholar]
- 24.Gangadevi V, Muthumary J. Taxol, an anticancer drug produced by an endophytic fungus Bartalinia robillardoides Tassi, isolated from a medicinal plant, Aegle marmelos Correa ex Roxb. World Journal of Microbiology Biotechnology. 2008;24:717–724. [Google Scholar]
- 25.Zhao J, Zhou L, Wang J, Shan T, Zhong L, Liu X, Gao X. Endophytic fungi for producing bioactive compounds originally from their host plants. In: Mendez-Vilas A, editor. Current Research, Technology and Education Topics in Applied Microbiology and Microbial Biotechnology. Vol. 1. Formatex Research Center; Badajoz, Spain: 2012. pp. 567–576. [Google Scholar]
- 26.Chandra S. Endophytic fungi: novel sources of anticancer lead molecules. Appled Microbiology and Biotechnology. 2012;95:47–59. doi: 10.1007/s00253-012-4128-7. [DOI] [PubMed] [Google Scholar]
- 27.Leiter J, Downing V, Hartwell JL, Shear J. Damage induced in sarcoma 37 with podophyllin, podophyllotoxin alpha-peltatin, beta-peltatin, and quercetin. Journal of the National Cancer Institute. 1950;10:1273–1293. [PubMed] [Google Scholar]
- 28.Stahelinand HF, von Wartburg A. The chemical and biological route from podophyllotoxin glucoside to etoposide: ninth Cain Memorial Award Lecture. Cancer Research. 1991;51:5–15. [PubMed] [Google Scholar]
- 29.Baldwin EL, Osheroff N. Etoposide, topoisomerase II and cancer. Current Medicinal Chemistry Anticancer Agents. 2005;5:363–72. doi: 10.2174/1568011054222364. [DOI] [PubMed] [Google Scholar]
- 30.Puri SC, Nazir A, Chawla R, Arora R, Riyaz-ul-Hasan S, Amna T, Ahmed B, Verma V, Singh S, Sagar R, Sharma A, Kumarc R, Sharma RK, Qazi GN. The endophytic fungus Trametes hirsuta as a novel alternative source of podophyllotoxin and related aryl tetralin lignans. Journal of Biotechnology. 2006;122:494–510. doi: 10.1016/j.jbiotec.2005.10.015. [DOI] [PubMed] [Google Scholar]
- 31.Eyberger AL, Dondapati R, Porter JR. Endophyte fungal isolates from Podophyllum peltatum produce podophyllotoxin. Journal of Natural Products. 2006;69:1121–1124. doi: 10.1021/np060174f. [DOI] [PubMed] [Google Scholar]
- 32.Lu L, He J, Yu X, Li G, Zhang X. Studies on isolation and identification of endophytic fungi strain SC13 from pharmaceutical plant Sabina vulgaris Ant. and metabolites. Acta Botany Boreal-Occident Sinica. 2006;15:85–89. [Google Scholar]
- 33.Kour A, Shawl AS, Rehman S, Sultan P, Qazi PH, Suden P, Khajuria RK, Verma V. Isolation and identification of an endophytic strain of Fusarium oxysporum producing podophyllotoxin from Juniperus recurva. World Journal of Microbiology and Biotechnology. 2008;24:1115–1121. [Google Scholar]
- 34.Kusari S, Lamshöft M, Spiteller M. Aspergillus fumigatus Fresenius, an endophytic fungus from Juniperus communis L. Horstmann as a novel source of the anticancer pro-drug deoxypodophyllotoxin. Journal of Applied Microbiology. 2009;107:1019–1030. doi: 10.1111/j.1365-2672.2009.04285.x. [DOI] [PubMed] [Google Scholar]
- 35.Oxford AE, Raistrick H, Simonart P. XXIX. Studies in the biochemistry of microorganisms. Griseofulvin, C17H17O6Cl, a metabolic product of Penicillium griseo-fulvum Dierckx. Biochemistry Journal. 1939;33:240–248. doi: 10.1042/bj0330240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Brian PW, Curtis PJ, Hemming HG. A substance causing abnormal development of fungal hyphae produced by Penicillium janczewskii Zal. I. Biological assay, production and isolation of “curling factor”. Trans Brit Mycol Soc. 1946;29:173–187. [Google Scholar]
- 37.Grove JF, Macmillan J, Mulholland TPC, Zealley J. Griseofulvin. Part III. The structures of the oxidation products C9H9O5Cl and C14H15O7Cl. Journal of the Chemical Society. 1952:3967–3977. [Google Scholar]
- 38.Grove JF, MacMillan J, Mulholland TPC, Thorold Rogers MA. Griseofulvin. Part IV. Structure. Journal of the Chemical Society. 1952:3977–3987. [Google Scholar]
- 39.Park JH, Choi GJ, Lee SW, Lee HB, Kim KM, Jung HS, Jang KS, Cho KY, Kim JC. Griseofulvin from Xylaria sp. strain F0010, an endophytic fungus of Abies holophylla and its antifungal activity against plant pathogenic fungi. Journal of Microbiology and Biotechnology. 2005;15:112–117. [Google Scholar]
- 40.Richardson SN, Walker AK, Nsiama TK, McFarlane J, Sumarah MW, Ibrahim A, Miller JD. Griseofulvin-producing Xylaria endophytes of Pinus strobus and Vaccinium angustifolium: evidence for a conifer-understory species endophyte ecology. Fungal Ecology. 2014;11:107–113. [Google Scholar]
- 41.Zabala AO, Chooi YH, Choi MS, Lin HC, Tang Y. ACS Chemical Biology. 2014;9:1576–1586. doi: 10.1021/cb500284t. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Wang J, Huang Y, Fang M, Zhang Y, Zheng Z, Zhao Y, Su W. Brefeldin A, a cytotoxin produced by Paecilomyces sp. and Aspergillus clavatus isolated from Taxus mairei and Torreya grandis. FEMS Immunology and Medical Microbiology. 2002;34:51–57. doi: 10.1111/j.1574-695X.2002.tb00602.x. [DOI] [PubMed] [Google Scholar]
- 43.Harri E, Loeffler W, Sigg HP, Staehelin S, Tamm C. Uber die isolierung der stoffwech selprodukte aus Penicillium brefel-dianum Dodge. Helvetica Chimica Acta. 1963;46:1235–1244. [Google Scholar]
- 44.Singleton VL, Bohonos N, Ullstrup AJ. Decumbin, a new compound from a species of Penicillium. Nature. 1958;181:1072–1073. doi: 10.1038/1811072a0. [DOI] [PubMed] [Google Scholar]
- 45.Betina V, Barath Z, Nemec P, Dobias J. Cyanein, a new antibiotic from Penicillium cyaneum. Folia Biologica. 1962;7:353–357. doi: 10.1007/BF02928123. [DOI] [PubMed] [Google Scholar]
- 46.Suzuki Y, Tanaka H, Aoki H, Tamura T. Ascotoxin (decumbin), a metabolite of Ascochyta imperfecta Peck. Agricultural and Biological Chemistry. 1970;34:395–413. [Google Scholar]
- 47.Coombe RG, Foss PS, Jacobs JJ, Watson TR. The biosynthesis of brefeldin A. Australian Journal of Chemistry. 1969;22:1943–50. [Google Scholar]
- 48.Bu’Lock JD, Clay PT. Fatty acid cyclization in the biosynthesis of brefeldin-A: a new route to some fungal metabolites. Journal of the Chemical Society. D: Chemical Communications. 1969:237–238. [Google Scholar]
- 49.a) Stierle AA, Stierle DB, Bugni T. Sequoiatones A and B: novel antitumor metabolites isolated from a redwood endophyte. Journal of Organic Chemistry. 1999;64:5479–5484. doi: 10.1021/jo990277l. [DOI] [PubMed] [Google Scholar]; b) Stierle A, Stierle DB, Bugni T. Journal of Natural Products. 2001;64:1350–1353. doi: 10.1021/np010022e. [DOI] [PubMed] [Google Scholar]
- 50.Stierle DB, Stierle AA, Bugni T. Sequoiamonascins A–D: novel anticancer metabolites isolated from a redwood endophyte. Journal of Organic Chemistry. 2003;68:4966–49699. doi: 10.1021/jo0340253. [DOI] [PubMed] [Google Scholar]
- 51.Lee JC, Strobel GA, Lobkovsky E, Clardy J. Torreyanic acid: a selectively cytotoxic quinone dimer from the endophytic fungus. Pestalotiopsis microspora Journal of Organic Chemistry. 1996;61:3232–3233. [Google Scholar]
- 52.Dai J, Krohn K, Flörke U, Draeger S, Schulz B, Kiss-Szikszai A. Metabolites from the endophytic fungus Nodulisporium sp. from Juniperus cedrus. European Journal of Organic Chemistry. 2006:3498–3506. [Google Scholar]
- 53.Dai J, Krohn K, Draeger S, Schulz B. New naphthalenechroman coupling products from the endophytic fungus, Nodulisporium sp. from Erica arborea. European Journal of Organic Chemistry. 2009:1564–1569. [Google Scholar]
- 54.Grove JFJ. Metabolic products of Phomopsis Oblonga. Part 2. Phomopsolides A and B, tiglic esters of two 6-substituted 5,6-dihydo-5-hydroxypyran-2-ones. Chemical Society Perkin Transactions. 1985;1:865–869. [Google Scholar]
- 55.Stierle DB, Stierle AA, Ganser B. New phomopsolides from a Penicillium sp. Journal of Natural Products. 1997;60:1207–1209. doi: 10.1021/np970338f. [DOI] [PubMed] [Google Scholar]
- 56.Calhoun LA, Findlay JA, Miller DJ, Whitney NJ. Metabolites toxic to spruce budworm from balsam fir needle endophytes. Mycological Research. 1992;96:281–286. [Google Scholar]
- 57.Ueno Y, Sato N, Ito T, Ueno I, Enomoto M, Tsunoda H. Chronic toxicity and hepatocarcinogenicity of (+) rugulosin, an anthraquinoid mycotoxin from Penicillium species: preliminary surveys in mice. Journal of Toxicological Sciences. 1980;5:295–302. doi: 10.2131/jts.5.295. [DOI] [PubMed] [Google Scholar]
- 58.Sumarah MW, Kesting JR, Sorensen D, Miller JD. Antifungal metabolites from fungal endophytes of Pinus strobus. Phytochemistry. 2011;72:1833–1837. doi: 10.1016/j.phytochem.2011.05.003. [DOI] [PubMed] [Google Scholar]
- 59.Hatakeyama T, Koseki T, Murayama T, Shiono Y. Eremophilane sesquiterpenes from the endophyte Microdiplodia sp. KS 75-1 and revision of the stereochemistries of phomadecalins C and D. Phytochemistry Letters. 2010;3:148–151. [Google Scholar]
- 60.Hu ZY, Li YY, Huang YJ, Su WJ, Shen YM. Three new sesquiterpenoids from Xylaria sp. NCY2. Helvetica Chimica Acta. 2008;91:46–52. [Google Scholar]
- 61.Xu R, Wang MZ, Lu CH, Zheng ZH, Shen YM. Tuberculariols A–C, new sesquiterpenes from the mutant strain M-741 of Tubercularia sp. TF 5. Helvetica Chimica Acta. 2009;92:1514–1519. [Google Scholar]
- 62.Pelaez F, Cabello A, Platas G, Diez MT, Gonzalez del Val A, Basilio A, Martan I, Vicente F, Bills GF, Giacobbe RA, Schwartz RE, Onishi JC, Meinz MS, Abruzzo GK, Flattery AM, Kong L, Kurtz MB. The discovery of enfumafungin, a novel antifungal compound produced by an endophytic Hormonema species biological activity and taxonomy of the producing organisms. Systematic and Applied Microbiology. 2000;23:333–343. doi: 10.1016/s0723-2020(00)80062-4. [DOI] [PubMed] [Google Scholar]
- 63.Onishi J, Meinz M, Thompson J, Curotto J, Dreikorn S, Rosenbach M, Douglas C, Abruzzo G, Flattery A, Kong L, Cabello A, Vicente F, Pelaez F, Diez MT, Martin I, Bills G, Giacobbe R, Dombrowski A, Schwartz R, Morris S, Harris G, Tsipouras A, Wilson K, Kurtz MB. Discovery of novel antifungal (1,3)-b-D-glucan synthase inhibitors. Antimicrobial Agents and Chemotherapy. 2000:368–377. doi: 10.1128/aac.44.2.368-377.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Kim S, Shin DS, Lee T, Oh KB. Periconicins, two new fusicoccane diterpenes produced by an endophytic fungus Periconia sp. with antibacterial activity. Journal of Natural Products. 2004;67:448–450. doi: 10.1021/np030384h. [DOI] [PubMed] [Google Scholar]
- 65.Shin DS, Oh MN, Yang HC, Oh KB. Biological characterization of periconicins, bioactive secondary metabolites, produced by Periconia sp. OBW-15. Journal of Microbiology and Biotechnology. 2005;15:216–219. [Google Scholar]
- 66.Calhoun LA, Findlay JA, Miller J, Whitney NJ. Metabolites toxic to spruce budworm from balsam fir needle endophytes. Mycological Research. 1992;96:281–286. [Google Scholar]
- 67.Endo A, Hasumi K, Sakai K, Kanbe T. Specific inhibition of glyceraldehyde-3-phosphate dehydrogenase by konigic acid (heptelidic acid) Journal of Antibiotics. 1985;38:920–925. doi: 10.7164/antibiotics.38.920. [DOI] [PubMed] [Google Scholar]
- 68.Yamaguchi Y, Manita D, Takeuchi T, Kuramochi K, Kuriyama I, Sugawara F, Yoshida H, Mizushina Y. Novel terpenoids, trichoderonic acids A and B isolated from Trichoderma virens, are selective inhibitors of family X DNA polymerases. Bioscience Biotechnology and Biochemistry. 2010;74:793–801. doi: 10.1271/bbb.90843. [DOI] [PubMed] [Google Scholar]
- 69.Kim JH, Choong HL. Heptelidic acid, a sesquiterpene lactone, inhibits etoposide-induced apoptosis in human leukemia U937 Cells. Journal of Microbiology and Biotechnology. 2009;19:787–791. [PubMed] [Google Scholar]
- 70.Tanak Y, Shiomi K, Kamaei K, Sugoh-Hagino M, Enomoto Y, Fang F, Yamaguchi Y, Masuma R, Zhang CG, Zhang XW, Omura S. Antimalarial activity of radicicol, heptelidic acid and other fungal metabolites. Journal of Antibiotics. 1998;51:153–160. doi: 10.7164/antibiotics.51.153. [DOI] [PubMed] [Google Scholar]
- 71.Findlay JA, Li G, Penner PE. Novel diterpenoid insect toxins from a conifer endophyte. Journal of Natural Products. 1995;58:197–200. [Google Scholar]
- 72.Findlay JA, Buthelezi S, Lavoie R, Rodriguez L. Bioactive isocoumarins and related metabolites from conifer endophytes. Journal of Natural Products. 1995;58:1759–1766. doi: 10.1021/np50125a021. [DOI] [PubMed] [Google Scholar]
- 73.Muth W, Nash CH. Biosynthesis of mycophenolic acid: purification and characterization of s-adenosyl-l-methionine: demethylmycophenolic acid o-methyltransferase. Antimicrobial Agents and Chemotherapy. 1975:321–327. doi: 10.1128/aac.8.3.321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Clutterbuck PW, Oxford AE, Raistrick H, Smith G. CLXXI. Studies in the biochemistry of microorganisms. XXIV. The metabolic products of the Penicillium brevi-compactum series. Biochemistry Journal. 1932;26:1442–1458. doi: 10.1042/bj0261441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Ando K, Suzuki S, Tamura G, Auma K. Antiviral activity of mycophenolic acid. Journal of Antibiotics. 1968;21:649–652. doi: 10.7164/antibiotics.21.649. [DOI] [PubMed] [Google Scholar]
- 76.Gilliver K. The inhibitory action of antibiotics on plant pathogenic bacteria and fungi. Annals of Botany. 1946;10:271–282. [Google Scholar]
- 77.Williams RH, Lively DH, Delong DC, Cline JC, Sweeney J, Poore GA, Parson SH. Mycophenolic acid: antiviral and antitumor properties. Journal of Antibiotics. 1968;21:463–464. doi: 10.7164/antibiotics.21.463. [DOI] [PubMed] [Google Scholar]
- 78.Mitsui A, Suzuki S. Immunosuppressive effect of mycophenolic acid. Journal of Antibiotics. 1969;22:358–363. doi: 10.7164/antibiotics.22.358. [DOI] [PubMed] [Google Scholar]
- 79.Johnson IS. Pharmaceuticals containing mycophenolic acid for treatment of psoriasis. Chemical Abstracts. 1972;77:92853. [Google Scholar]
- 80.Mimouni D, Anhalt GJ, Cummins DL, Kouba DJ, Thorne JE, Nousari HC. Treatment of pemphigus vulgaris and pemphigus foliaceus with mycophenolate mofetil. Archives of Dermatology. 2003;139:739–742. doi: 10.1001/archderm.139.6.739. [DOI] [PubMed] [Google Scholar]
- 81.Diamond MS, Zachariah M, Harris E. Mycophenolic acid inhibits dengue virus infection by preventing replication of viral RNA. Virology. 2002;304:211–221. doi: 10.1006/viro.2002.1685. [DOI] [PubMed] [Google Scholar]
- 82.Bloch P, Tamm C, Bollinger P, Petcher T, Weber HP. Pseurotin, a new metabolite of Pseudeurotium ovalis Stolk having an unusual heterospirocyclic system. Helvetica Chimica Acta. 1976;59:133–137. doi: 10.1002/hlca.19760590114. [DOI] [PubMed] [Google Scholar]
- 83.Schmeda-Hirschmann G, Hormazabal E, Rodriguez JA, Theoduloz C. Cycloaspeptide A and pseurotin A from the endophytic fungus Penicillium janczewskii. Zeitschrift für Naturforschung C. 2008;63:383–388. doi: 10.1515/znc-2008-5-612. [DOI] [PubMed] [Google Scholar]
- 84.Ishikawa M, Ninomiya T, Akaban H, Kushida N, Tsujiuchi G, Ohyama M, Gomi S, Shito K, Murata T. Pseurotin A and its analogues as inhibitors of immunoglobuline E production. Bioorganic & Medicinal Chemistry Letters. 2009;19:1457–1460. doi: 10.1016/j.bmcl.2009.01.029. [DOI] [PubMed] [Google Scholar]
- 85.Kawashima K, Nakanishi K, Nishikawa H. Structure of tauranin and a note on the “C-16-Acids” obtained from di- and triterpenoids. Chemical and Pharmaceutical Bulletin. 1964;12:796–803. doi: 10.1248/cpb.12.796. [DOI] [PubMed] [Google Scholar]
- 86.Wijeratne EMK, Paranagama PA, Marron MT, Gunatilaka MK, Arnold AE, Gunatilaka AAL. Sesquiterpene quinones and related metabolites from Phyllosticta spinarum, a fungal strain endophytic in Platycladus orientalis, of the sonoran desert. Journal of Natural Products. 2008;71:218–222. doi: 10.1021/np070600c. [DOI] [PubMed] [Google Scholar]
- 87.Arai T, Mikami Y, Fushima K, Utsumi T, Yazawa K. A new antibiotic, leucinostatin, derived from Penicillium lilacinum. Journal of Antibiotics. 1973;26:157–161. doi: 10.7164/antibiotics.26.157. [DOI] [PubMed] [Google Scholar]
- 88.Kawada M, Inoue H, Ohba SI, Masuda T, Momose I, Ikeda D. Leucinostatin A inhibits prostate cancer growth through reduction of insulin-like growth factor-I expression in prostate stromal cells. International Journal of Cancer. 2010;126:810–818. doi: 10.1002/ijc.24915. [DOI] [PubMed] [Google Scholar]
- 89.Strobel GA, Hess WM. Glucosylation of the peptide leucinostatin A, produced by an endophytic fungus of European yew, may protect the host from leucinostatin toxicity. Chemistry and Biology. 1997;4:529–536. doi: 10.1016/s1074-5521(97)90325-2. [DOI] [PubMed] [Google Scholar]
- 90.Noble HM, Langley D, Sidebottom PJ, Lane SJ, Fisher PJ. An echinocandin from an endophytic Cryptosporiopsis sp. and Pezicula sp. in Pinus sylvestris and Fagus sylvatica. Mycological Research. 1991;95:1439–1440. [Google Scholar]
- 91.Chapman SW, Sullivan DC, Cleary JD. In search of the Holy Grail of antifungal therapy. Transactions of the American Clinical and Climatological Association. 2008;119:197–216. [PMC free article] [PubMed] [Google Scholar]
- 92.Kobayashi R, Samejina Y, Nakajima S, Kawai K, Udagawa S. Studies on fungal products XI. Isolation and structures of novel cyclic pentapeptides from Aspergillus sp. NE-45. Chemical and Pharmaceutical Bulletin. 1987;35:1347–1352. doi: 10.1248/cpb.35.1347. [DOI] [PubMed] [Google Scholar]
- 93.Dalsgaard PW, Larsen TO, Christophersen C. Bioactive cyclic peptides from the psychrotolerant fungus Penicillium algidum. Journal of Antibiotics. 2005;58:141–144. doi: 10.1038/ja.2005.16. [DOI] [PubMed] [Google Scholar]
- 94.Sun HH, Barrow CJ, Sedlock DM, Gillum AM, Cooper R. Benzomalvins, new substance P inhibitors from a Penicillium sp. Journal of Antibiotics. 1994;47:515–522. doi: 10.7164/antibiotics.47.515. [DOI] [PubMed] [Google Scholar]
- 95.Wong SM, Musza LL, Kydd GC, Kullnig R, Gillum AM, Cooper RJ. Fiscalins: new substance P inhibitors produced by the fungus Neosartorya fischeri. Taxonomy, fermentation, structures, and biological properties. Journal of Antibiotics. 1993;46:545–53. doi: 10.7164/antibiotics.46.545. [DOI] [PubMed] [Google Scholar]
- 96.Regoli D, Drapeau G, Dion S. New selective agonists for neurokinin receptors: pharmacological tools for receptor characterization. Trends in Pharmacological Sciences. 1988;9:290–295. doi: 10.1016/0165-6147(88)90013-2. [DOI] [PubMed] [Google Scholar]
- 97.Pernow B. Substance P. Pharmacology Reviews. 1983;35:85–141. [PubMed] [Google Scholar]
- 98.Nagel DW, Pachler KGR, Steyn PS, Vleggaar R, Wessels PL. The chemistry and 13C NMR assignments of oxaline, a novel alkaloid from Penicillium oxalicum. Tetrahedron. 1976;32:2625–2631. [Google Scholar]
- 99.Koizumi Y, Arai M, Tomoda H, Omura S. Oxaline, a fungal alkaloid, arrests the cell cycle in M phase by inhibition of tubulin polymerization. Biochimica et Biophysica Acta. 2004;1693:47–55. doi: 10.1016/j.bbamcr.2004.04.013. [DOI] [PubMed] [Google Scholar]
- 100.Polonsky J, Merrien MA, Scott PM. Roquefortine and isofumigaclavine A, alkaloids from Penicillium roqueforti. Annales de la nutrition et de l’alimentation. 1977;31:963–8. [PubMed] [Google Scholar]
- 101.Mantle PG, Perera KP, Maishman NJ, Mundy GR. Biosynthesis of penitrems and roquefortine by Penicillium crustosum. Applied Environmental Microbiology. 1983;45:1486–1490. doi: 10.1128/aem.45.5.1486-1490.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Steyn PS, Vleggaar R. Roquefortine, an intermediate in the biosynthesis of oxaline in cultures of Penicillium oxalicum. Journal of the Chemical Society: Chemical Communications. 1983:560–561. [Google Scholar]