

Abstract
Plant growth and development can be influenced by mutualistic and non-mutualistic microorganisms. We investigated the ability of the ericoid endomycorrhizal fungus Oidiodendron maius to influence growth and development of the non-host plant Arabidopsis thaliana. Different experimental setups (non-compartmented and compartmented co-culture plates) were used to investigate the influence of both soluble and volatile fungal molecules on the plant phenotype. O. maius promoted growth of A. thaliana in all experimental setups. In addition, a peculiar clumped root phenotype, characterized by shortening of the primary root and by an increase of lateral root length and number, was observed in A. thaliana only in the non-compartmented plates, suggesting that soluble diffusible molecules are responsible for this root morphology. Fungal auxin does not seem to be involved in plant growth promotion and in the clumped root phenotype because co-cultivation with O. maius did not change auxin accumulation in plant tissues, as assessed in plants carrying the DR5::GUS reporter construct. In addition, no correlation between the amount of fungal auxin produced and the plant root phenotype was observed in an O. maius mutant unable to induce the clumped root phenotype in A. thaliana. Addition of active charcoal, a VOC absorbant, in the compartmented plates did not modify plant growth promotion, suggesting that VOCs are not involved in this phenomenon. The low VOCs emission measured for O. maius further corroborated this hypothesis. By contrast, the addition of CO2 traps in the compartmented plates drastically reduced plant growth, suggesting involvement of fungal CO2 in plant growth promotion. Other mycorrhizal fungi, as well as a saprotrophic and a pathogenic fungus, were also tested with the same experimental setups. In the non-compartmented plates, most fungi promoted A. thaliana growth and some could induce the clumped root phenotype. In the compartmented plate experiments, a general induction of plant growth was observed for most other fungi, especially those producing higher biomass, further strengthening the role of a nonspecific mechanism, such as CO2 emission.
Introduction
Plant-associated microorganisms are essential drivers of plant productivity because they can increase nutrient availability and uptake, enhance stress tolerance, provide disease resistance and expand plant metabolic pathways [1,2]. They also play important functions in plant development, and functional traits such as leaf development, shoot/root ratio and root architecture may undergo substantial changes following plant-microorganism interactions [2,3]. In particular, plant growth-promoting rhizobacteria (PGPR; [4]) and rhizospheric fungi (PGPF [5]) are able to promote plant growth and development thanks to direct and indirect mechanisms. Indirect mechanisms include improved mineral nutrition through mineral solubilisation or disease suppression, whereas direct mechanisms involve production of phytohormones and volatile organic compounds (VOCs) [6]. Many PGPR strains can release volatile mixtures that stimulate plant growth [7].
Plant growth and development may be influenced by the interaction with either beneficial or pathogenic microorganisms [3,6]. For example, several non-mutualistic PGPR modify plant root architecture by increasing primary root length, lateral root number, length and density, or root hair formation [8–12]. Several non-mutualistic fungi and root-associated endophytic fungi have been described in the literature as PGPF. Among them are strains belonging to the genera Penicillium, Fusarium, Phoma, Trichoderma, Ampelomyces, Coniothyrium, Aspergillus, Sarocladium, Ophiosphaerella, Piriformospora [6,13–19]. The mycorrhizal symbiosis facilitates water and nutrient absorption and positively affects plant growth [20]. For example, roots colonized by arbuscular mycorrhizal (AM) fungi display enhanced root biomass and increased lateral roots if compared to non-AM roots [21–24]. Ectomycorrhizal (ECM) fungi also increase root growth and lateral root formation in their host plants, which typically display numerous short lateral roots [25–30]. Several studies have shown stimulation of lateral root development in the host plant in the very early phase of the ECM interaction, prior to symbiosis establishment, suggesting that soluble diffusible/volatile signalling molecules are responsible for changes in root architecture in this early phase [29,31,32]. Some ECM fungi can also induce lateral root development in the non-host plant Arabidopsis thaliana, indicating that signalling is non-host specific and that the root phenotype does not depend on the plant’s ability to form ECM [29,31,32]. Felten et al. [29] and Splivallo et al. [31] demonstrated that fungal-derived indole-3-acetic acid (IAA) plays an important role in modifying root morphology, and that fungal-derived ethylene may influence plant lateral root development and branching in the early stages of interaction. ECM fungi may thus modify the endogenous hormonal plant balance, and Felten et al. [29] proposed a model where the fungus induces auxin accumulation at the root apex, thus stimulating lateral root formation. In addition to soluble phytohormones, fungi also emit species-specific blends of VOCs. The emission intensity as well as the VOCs composition depends strongly on physiological as well as environmental factors [33,34]. Recently, it was shown that plants could sense fungal emitted sesquiterpenes [32,35]; the airborne plant-fungal communication through the sesquiterpene thujopsene was suggested to prepare the plants for mycorrhizal symbiosis [32].
Ericoid mycorrhizal (ERM) fungi are soil-born fungi mostly belonging to Leotiomycetes (Ascomycetes) and form symbiotic associations with plants in the family Ericaceae [36], in which they induce plant growth under stressful conditions [37,38]. The influence of ERM fungi on the growth of other plant species has never been investigated, despite the identification of ERM-related fungi in association with the roots of non-ericaceous plants [39]. We have investigated the influence of the ERM fungus Oidiodendron maius [40] on growth and development of the non-mycorrhizal model plant A. thaliana. We used different experimental setups to investigate the role of both soluble and volatile molecules on the plant phenotype, and compared O. maius with nine other fungi comprising diverse ecological strategies, as well as with some O. maius mutants. O. maius strongly promoted plant growth in A. thaliana and induced a peculiar root phenotype that was also induced by other fungal species, but not by an O. maius mutant with altered nitrogen pathways. Unlike ECM fungi, O. maius does not seem to resort on IAA or VOCs to promote growth. Instead, the results underlined a major non-specific contribution of fungal emitted CO2 to plant growth promotion in our in vitro experimental setup.
Materials and Methods
Fungal strains and culture media
Oidiodendron maius strain Zn is deposited at the Mycotheca Universitatis Taurinensis collection (MUT1381) of the Department of Life Sciences and Systems Biology (University of Turin, Italy) and at the American Type Culture Collection (ATCC MYA-4765) of the University of Boulevard (Manassas, VA, US). This strain was isolated from the Niepolomice Forest (25 km northeast of Krakow, Poland) from the roots of Vaccinium myrtillus plants growing in experimental plots treated with dust containing high concentrations of heavy metals [41]. Nine fungal strains, obtained from the INRA fungal collection (Nancy, France), were used for comparison. They include three other ERM strains (Meliniomyces bicolor, M. variabilis and Rhizoscyphus ericae), three ECM strains (Cenococcum geophilum, Laccaria bicolor strain S238N and Suillus luteus), one orchid mycorrhizal strain (Tulasnella calospora), one white rot saprothrophic fungus (Trametes versicolor) and one pathogenic fungus (Cladosporium herbarum). Three characterized O. maius mutants were also used for comparative experiments: the O. maius SOD mutant [42], the O. maius mutant carrying a disruption on a gene belonging to the Major Facilitator Superfamily (MFS) transporter family (Abbà, unpublished) and the O. maius GOGAT mutant [43]. This last mutant carries a partial deletion of the glutamate synthase (NADH-GOGAT) gene [43]. The OmGOGAT disruption modifies the nitrogen pathway and this mutant has an altered nitrogen metabolism. All fungal strains were maintained on Czapek-glucose solid medium (NaNO3 3 g/L, K2HPO4*3H2O 1.31 g/L, MgSO4*7H2O 0.5 g/L, FeSO4*7H2O 0.01 g/L, KCl 0.5 g/L, glucose 20 g/L, agar 10 g/L). The medium was adjusted to pH 6 with the addition of 1 M HCl. All reagents were purchased from SIGMA.
Plant growth
Seeds of A. thaliana (the ecotype Col-0 and DR5::GUS transformant) were surface sterilized with a solution containing 70% ethanol for 10 minutes and 100% ethanol for few seconds. Then seeds were dried 2–3 h in the sterile hood and transferred on 1% agar medium containing 2.29 g/L half strength MS [44] medium (Murashige and Skoog Basal Salts Mixture including vitamins), 10 g/L sucrose, 1 g/L MES (2-N-morpholino ethane sulphonic acid) sodium salt (pH 5.8–6) for germination. Plates were kept at 4°C for 2 days and then they were placed vertically in a plant growth chamber (16-h photoperiod, light at 170 μmol m–2s–1, temperature at 23°C day and 21°C night) for 5–7 days.
Cellophane and cellulose nitrate membrane preparation
When necessary, prior to inoculation, sterile cellophane or cellulose nitrate membranes were placed aseptically on the agar surface to provide a convenient means of removing/transferring mycelia or plants. Cellophane membranes were prepared by first boiling for 30 min in 10 mM EDTA (disodium salt, dihydrate, SIGMA), rinsing and then autoclaving in ddH2O, while cellulose nitrate membranes were autoclaved and then dried over night at 50°C.
Co-culturing of fungi and A. thaliana plants
Co-culturing of fungi with A. thaliana plants included three different experimental setups. (1) Non-compartmented square plates. Twelve centimeters square petri dishes containing MS medium were used and five A. thaliana germinated seedlings prepared as described above were placed on the half-upper side of each petri dish. When the A. thaliana seedlings’ roots length reached 5 cm, the half-bottom side of the petri dish was inoculated with fungi. Two different systems were used for fungal inoculation: (a) three 0.5-cm mycelial plugs were placed directly on the MS solidified medium; (b) three 0.5-cm mycelial plugs of O. maius were previously grown for 15 days on cellulose nitrate membranes (Scheicher&Schuell, ME24ST, 0.2 μm, 47 mm) in MS containing petri plates. The membranes covered by the mycelium were then placed fungus side down (direct interaction) or fungus side up (indirect interaction) on the A. thaliana roots. Inoculated plates (including non-inoculated control plates) were placed vertically in a growth chamber (16-h photoperiod, light at 170 μmol m–2 s–1, temperature at 23°C day and 21°C night) for 30 days. (2) Bi-partite plates. Nine centimeters round bi-partite petri dishes containing MS medium were used. Three A. thaliana seedlings, prepared as described above, were placed in the upper part of the right plate compartment, while one fungal plug was inoculated in the middle of the left plate compartment previously covered with a sterile cellophane membrane prepared as described above. Control plates without fungi were also prepared. In this setup, the mycelial exudates do not diffuse to the plant compartment and only fungal volatiles can reach the seedlings. These plates were placed vertically in a growth chamber (16-h photoperiod, light at 170 μmol m–2 s–1, temperature at 23°C day and 21°C night) for two weeks. (3) Tri-partite plates with volatile and CO2 traps. Round nine centimeters tri-partite petri dishes were used. Three A. thaliana seedlings, prepared as described above, were placed in one compartment and one 0.5 cm fungal plug was inoculated in the second compartment previously covered with a sterile cellophane membrane prepared as described above. Both compartments contained MS medium. The third compartment was filled with volatile or CO2 traps. The volatile trap was made of 2 g of activated charcoal (untreated granular 8–20 mesh—C2889 Sigma-Aldrich). As CO2 trap, 7 ml of 0.1 M Ba(OH)2*8H20 were added together with two dental rolls to avoid barium hydroxide spillage in the neighboring plate compartments. 7 ml of 1 M Ca(OH)2 and 0.9 g of Ca(OH)2 (solid form) were also tested as CO2 traps. Control plates without fungi, control plates without the two trap compounds and control plates with a CO2 saturated barium hydroxide solution were also prepared. These plates were placed horizontally in a growth chamber (16-h photoperiod, light at 170 μmol m–2 s–1, temperature at 23°C day and 21°C night) for two weeks.
Five plates, each containing three to five plants, were prepared and analyzed for each treatment. Fresh and dry weights of mycelia, roots and aboveground (stem + leaves) plant portions from all the different co-culture systems described were determined. After harvest, mycelia and plants were blotted dry on a paper towel to remove agar and water excess, and fresh weights (FW) were measured. Plant dry weights (DW) were measured after drying plant material in a ventilated oven at 60°C to a constant weight. Plant images were recorded by using a Nikon eclipse E300 system.
Morphological analysis of A. thaliana root development
Twenty individual A. thaliana plants for each condition (five plants per Petri plate, 4 replicates) were observed every three days up to 12 days in a control experiment and after co-cultivation with O. maius WT or with the O. maius GOGAT mutant. For the quantification of root parameters, plants after 9 days of plant-fungus co-cultivation were considered because modifications of the root phenotype were already visible at this developmental stage, and roots had not touched the plate side either in the control plates or in the co-cultivation plates. Images of the whole plants were acquired with an Epson Perfection V300 scanner (Epson America, USA) at 600 dpi and saved in TIFF format. Primary and lateral roots (LRs) were counted and measured using the ImageJ plug in SmartRoot software [45].
Sampling and analysis of fungal VOCs
O. maius was grown at 25°C in plastic (6 replicates) and glass (3 replicates) petri plates containing MS medium covered with cellophane membranes. VOCs were collected in the cultures headspace after 15 and 30 days from fungal inoculum. Control plates without mycelium were sampled for background correction. VOCs were collected for 6 h from sealed Petri dishes by headspace sorptive extraction using the stir bar sorptive extraction method with Gerstel Twisters (Gerstel GmbH & Co. KG, Mülheim an der Ruhr, Germany) as described in [33]. The samples were analysed with a thermo-desorption unit (Gerstel GmbH & Co) coupled to a gas chromatograph-mass spectrometer (GC-MS; GC model: 7890A; MS model: 5975C; Agilent Technologies, Santa Clara, CA, USA) as described in [34]. The chromatograms were analyzed by the enhanced ChemStation software (MSD ChemStation E.02.01.1177, 1989–2010 Agilent Technologies, Santa Clara, CA, USA). The TIC (Total Inorganic Carbon) of each VOC in the final dataset was recalculated from the absolute abundance of the first representative m/z to eliminate noise. The calibration was done as described in [34]. The emission rates were calculated on fungal mycelium area (pmol cm-2 h-1) bases.
GUS assay
The GUS assay was done on aboveground (stems, leaves) and belowground portions of A. thaliana fresh tissues, sampled after 7 days of co-cultivation in non-compartmented plates. Each sample was incubated with the GUS-buffer (0.1 M sodium phosphate buffer pH 7; 5 mM K4Fe(CN)6; 5 mM K3Fe(CN)6), 0.1% Triton X100, 0.1% x-GlcA (5-Bromo-4-chloro-3-indolyl-ß-D-glucuronic acid) reagent (Duchefa Biochemie), 1 mM EDTA for 16h at 37°C in the dark and then washed with 70% ethanol. Tissues were observed and photographed using a Nikon Eclipse E400 optical microscope.
Auxin measurement using Salkowski reaction
Two mycelial plugs of O. maius WT, O. maius GOGAT mutant, O. maius MFS mutant and O. maius SOD mutant were used to inoculate three flasks each containing 40 ml of MS liquid medium. Three flasks containing the growth medium were used as a negative control. Flasks were kept on a shaking incubator at 120 rpm at 25°C in the dark. After twenty days of culture, fungal growing media were vacuum filtered through filter paper disks and concentrated through lyophilization. Three 1 ml aliquots of 8X concentrated samples were prepared and a colorimetric assay was used to estimate the concentration of indole compounds by mixing the supernatant with 1 ml of Salkowski reagent (0.138 M FeCl3 and 7.9 M H2SO4) [46]. Samples were incubated at room temperature for 30 min in the dark and analyzed at 530 nm on a spectrophotometer (Beckman DU®530). IAA levels were determined with an IAA standard curve using commercial IAA (Duchefa—Biochemie) and sterile medium as a blank. IAA levels were expressed as μg/g of dry mycelium.
Statistical analysis
The significance of differences among the different treatments was statistically evaluated by ANOVA with Tukey’s pairwise comparison as post hoc test for multiple comparisons for normal distributed data. Kruskall-Wallis with Bonferroni-corrected pairwise Mann-Whitney post-hoc adjustment were used as statistical test for non-normal distributed data. Pearson’s correlation test was performed to measure the strength of the association between plant and fungal biomass values obtained in the bi-partite plate assay, considering for the significance of the correlation a probability level <0.01. Statistical elaborations of growth and biomass data were performed using PAST statistical package, version 2.17 [47]. For statistical analysis of the VOC samples, Kruskall-Wallis test with Dunnet T3 post hoc test was used. For dependent samples Wilcoxons test was used. The differences were considered significant at a probability level of P<0.05.
Results
O. maius positively influences development of the non-host plant A. thaliana
When co-cultured with O. maius in Petri dishes, the biomass of both roots and aerial parts of A. thaliana significantly increased (Fig 1). In particular, a 4-fold biomass increase of the aerial parts and a 5-fold increase of the root biomass were measured after direct inoculation of O. maius on the solid medium (Fig 1a and 1b). If the fungus was inoculated on a cellulose nitrate membrane before its transfer to the co-culture plates, a significant biomass increase was observed in A. thaliana irrespective of the membrane side in contact with the roots (Fig 1c and 1d). In this last experiment, a significantly (P<0.05) higher biomass of the aerial portion was recorded when O. maius was in direct contact with the plant roots, as compared with the indirect interaction, while no significant differences were recorded for the root biomass (Fig 1d). Interestingly, the increase in root biomass induced by O. maius correlated with a pronounced shortening of the primary root and an increase of lateral root (LR) length and number, leading to a particular clumped root phenotype well visible in Fig 1a.
Fig 1. A. thaliana development in the presence of O. maius.

(a) A. thaliana control plants (C) and A. thaliana-O. maius co-cultures (Om) 30 days after inoculation; (b) plant biomass measurements (roots—grey bars—and aboveground portions—open bars) in the presence (Om)/absence (C) of O. maius. Note the strong plant biomass increase in the presence of the fungus; (c) A. thaliana-O. maius co-cultures 30 days after inoculation: the fungus was previously grown on cellulose nitrate disks which were then placed on A. thaliana roots fungus side up (indirect contact—Om-IC) or fungus side down (direct contact—Om-DC); (d) plant biomass measurements (roots—grey bars—and aboveground portions—open bars) in the presence/absence of O. maius in indirect/direct contact with plant roots. Note the strong plant biomass increase in the presence of the fungus in both conditions and especially in the direct contact one. All pictures were taken at the same magnification. Bars represent the mean ±SD, n = 5 (each biological replicate represents the total biomass of 5 A. thaliana seedlings grown in an individual plate). Statistically significant differences (P<0.05) among treatments are indicated by asterisks or by different letters above the bars.
An O. maius GOGAT mutant could not induce the A. thaliana clumped root phenotype
The use of genetic mutants is a powerful tool to associate a particular phenotype to specific genes. Therefore, three available characterized O. maius mutants (see “Fungal strains and culture media” paragraph) were also tested: two of them induced in A. thaliana the same clumped root phenotype as the wild-type strain, whereas the O. maius GOGAT mutant, altered in N metabolism, did not (Fig 2a).
Fig 2. A. thaliana development in the presence of O. maius WT (Om) and of three O. maius mutants (OmΔGOGAT; OmΔMFS; OmΔSOD).

(a) Control plants (C) and plant-fungus co-cultures 30 days after inoculation (all pictures were taken at the same magnification) (b) Measurement of auxin quantity released in the culture medium by O. maius WT and by the three O. maius mutants, using the Salkowski reaction [46]. Auxin quantity measured was normalized to the mycelium biomass. Bars represent the mean ±SD, n = 3 (each biological replicate represents the total biomass of 5 A. thaliana seedlings grown in an individual plate). Statistically significant differences (P<0.05) among treatments are indicated by different letters above the bars.
Morphometric analysis of A. thaliana root development in the presence of the O. maius WT and of the O. maius GOGAT mutant
To better describe the two different A. thaliana root phenotypes observed after co-cultivation with the O. maius WT and the O. maius GOGAT mutant, plant root development was followed in a time course experiment (S4 Fig) and morphometric analyses were performed after 9 days of co-culture using the SmartRoot software tool. Average values of the number and length of three different root orders -primary (PR), secondary (SRs) and tertiary roots (TRs)- were calculated for each replicate (Fig 3). When compared to control plants, the root phenotype in the presence of the O. maius WT strain was characterized by PR shortening, an increase of SR and TR length and an increase of TR number (Fig 3). On the other hand, no significant PR shortening was observed in the presence of the O. maius GOGAT mutant. For these plants, an increase of SR and TR length and an increase of TR number were measured (Fig 3). If compared to control plants, the SR length increased more in the presence of the O. maius GOGAT mutant (3.2 times) than the WT strain (1.6 times) (Fig 3). On the other hand, TRs were 2.8 times longer in the WT strain than in the mutant strain (Fig 3).
Fig 3. Analysis of A. thaliana root development.

Average quantification of root parameters in 20 individual A. thaliana plants grown alone or co-cultured for 9 days with the O. maius WT strain (Om) or the O. maius GOGAT mutant (OmΔGOGAT) is plotted into charts. Primary (PR), secondary (SRs) and tertiary roots (TRs) were counted and measured using the ImageJ plug in SmartRoot software. A diagrammatic representation of A. thaliana root development in the different conditions tested is also shown.
Auxin is not involved in the A. thaliana—O. maius interaction
Felten et al. [29] showed that LR stimulation in A. thaliana by an ECM fungus paralleled an increase of auxin in the root apices. As alterations of nitrogen metabolism could affect auxin production [48], we tested whether the inability of the O. maius GOGAT mutant to induce the clumped root phenotype in A. thaliana could be related to a different production of fungal auxin. The amount of IAA produced by the wild type O. maius strain was significantly higher when compared to the amount produced by the three mutants tested in the plate assay (Fig 2b). However, since auxin production by the O. maius GOGAT mutant was similar to the other two mutants, this result would exclude a possible role of fungal auxin in the A. thaliana clumped root phenotype. We also verified possible induction of auxin accumulation in the plant tissues by using the A. thaliana DR5::GUS [49], that carries the promoter of the auxin-responsive DR5 gene fused with the GUS reporter gene, co-cultivated with O. maius WT and with the O. maius GOGAT mutant. After GUS staining, the blue dye was localized in the root apex and in the vascular tissues of the primary root (S1 Fig) as well as in the LR primordia, and in the secondary root tips (S1 Fig). Auxin was also detected in some areas of the leaf margin (S1 Fig). However, no differences were observed in the distribution and accumulation of GUS staining in A. thaliana roots and leaves, either in the absence or in the presence of the O. maius WT and the O. maius GOGAT mutant (S1 Fig). All together, these results suggest that auxin is probably not involved in the induction of the clumped root phenotype or in plant growth promotion by O. maius.
VOCs do not seem responsible for increased A. thaliana biomass
As previous papers reported a role of fungal-derived VOCs in root development induced by fungi [32], further experiments were performed to investigate a potential role of O. maius volatiles in A. thaliana growth promotion. Co-cultivation experiments were set up in tri-partite plates that only allowed air contact between A. thaliana and O. maius (Fig 4a). Plates containing a VOCs trap (activated charcoal) and a CO2 trap (barium hydroxide) were also used in parallel (Fig 4b and 4c). As the O. maius GOGAT mutant behaved differently from the O. maius WT in the square non-compartmented plates, this mutant strain was also tested in the tri-partite plate setup (S2 Fig). After 15 days of co-cultivation, A. thaliana roots and aerial portions were collected and fresh and dry weights were recorded (Figs 4d and S2d). Growth of O. maius mycelia was also recorded, and was similar in the different conditions tested (data not shown). When A. thaliana was grown in the absence of the fungus, no significant differences were measured between control plates and those added with activated charcoal. By contrast, A. thaliana biomass was significantly lower on plates containing the CO2 trap barium hydroxide (Figs 4c and 4d, S2c and S2d). To exclude possible phytotoxic effects of barium hydroxide, A. thaliana was grown in plates containing a CO2-saturated barium hydroxide solution. In this experiment, plant growth was not significantly different from control plates without barium hydroxide (S3 Fig). These results excluded a phytotoxic effect of barium hydroxide and confirmed that the dwarf plant phenotype was caused by CO2 depletion.
Fig 4. O. maius—A. thaliana co-cultivation experiments in the tripartite plate system.

(a) Control plants and plant-fungus co-culture 15 days after inoculation; (b) same as in (a) but plates were added with a VOC trap (activated charcoal, AC) in the third compartment; (c) same as in (a) but plates were added with a CO2 trap [Ba(OH)2*8H2O, B] together with two dental rolls in the third compartment; (d) plant biomass measurements (roots—grey bars—and aboveground portions—open bars) in the presence/absence of the fungus and of the trap compounds. Note the strong plant biomass increase in the presence of O. maius in all the conditions tested. Bars represent the mean ±SD, n = 5 (each biological replicate represents the total biomass of 3 A. thaliana seedlings grown in an individual plate). Statistically significant differences (P<0.05) among treatments are indicated by different letters above the bars.
Significant growth induction of both roots and aerial parts of A. thaliana was recorded in all treatments where the fungus, either O. maius WT or the GOGAT mutant, was present, indicating a release of growth-promoting volatile molecules. However, a similar A. thaliana biomass was recorded in the absence and in the presence of activated charcoal, an effective VOCs trap [50]. This result suggested that growth-promoting compounds were likely not VOCs (Figs 4b and 4d, S2b and S2d). Similar conclusion derived from direct measurement of VOCs emission by O. maius WT and GOGAT mutant. The results revealed in general a very low release of VOCs from both fungal strains (Fig 5). The typical fungal odor compound, 1-octen-3-ol, was emitted exclusively by 15 days old fungi, whereas emission of the sesquiterpene germacrene D was higher in the older fungal culture (Fig 5). Some of the compounds, such as the fungicide phenol,2,4-bis(1,1-dimethylethyl), showed higher emission for O. maius WT than for the O. maius GOGAT mutant (Fig 5), but it might be due to the smaller mycelium size of the mutant at the time of measurement.
Fig 5. VOC emission profiles of the O. maius WT and of the O. maius GOGAT mutant.
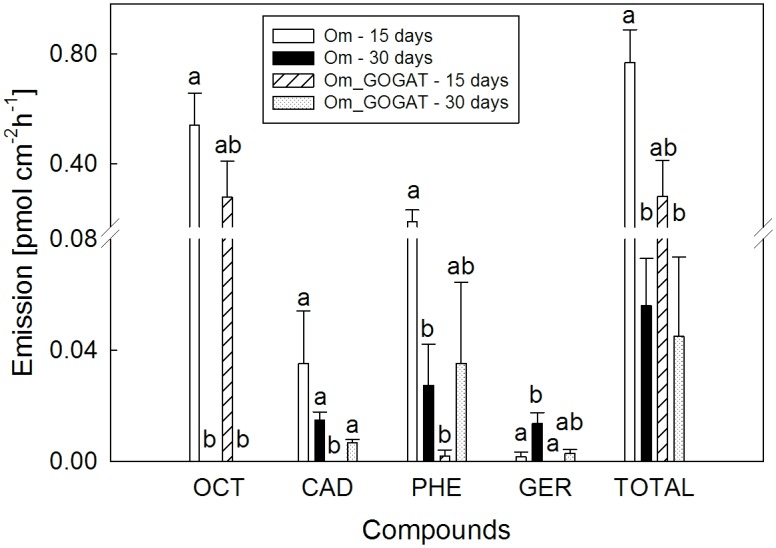
VOCs were collected in the headspace of culture plates 15 (open and hatched bars) and 30 (black and dotted bars) days after inoculation. Bars represent the mean ±SD as pmol cm-2 h-1, n = 6. (OCT) 1-octen-3-ol; (CAD) epsilon-cadinene; (PHE) phenol,2,4-bis(1,1-dimethylethyl); (GER) germacrene D.
In the presence of barium hydroxide, both O. maius WT and GOGAT mutant rescued the dwarf A. thaliana phenotype caused in the control plates by the presence of this CO2 trap. Growth of the aerial plant portion was slightly less when compared to the other co-culture conditions tested (Figs 4c and 4d, S2c and S2d), likely because part of the CO2 emitted by the fungus was adsorbed by, and saturated, the barium hydroxide trap.
Various fungal strains promoted A. thaliana growth in non-compartmented and compartmented plates
To investigate whether the clumped root phenotype and the biomass increase observed in A. thaliana were specifically induced by O. maius, different fungal strains were tested using the non-compartmented (Fig 6) and compartmented (Fig 7) plate setups. Among the fungal strains tested, two other ERM fungi (M. variabilis and R. ericae), the saprotrophic fungus T. versicolor and the pathogenic fungus C. herbarum induced the clumped root phenotype, while the ERM fungus M. bicolor, two ECM fungi (L. bicolor and C. geophilum) and the orchid mycorrhizal fungus T. calospora did not (Fig 6a). An intermediate situation was observed for S. luteus (Fig 6a). Irrespective of the root phenotype, most of these fungi significantly increased the biomass of both A. thaliana roots and aerial portions (Fig 6b), in particular the four ERM fungi, the ECM fungi S. luteus and C. geophylum and the saprotrophic fungus T. versicolor (Fig 6b). It should be also noted that, irrespective of the biomass increase and root phenotype, some other developmental traits of A. thaliana (e.g. flowering, leaf area) were depending on the fungus in the co-cultivation plates, despite the fact that seedlings were all at the same developmental stage at the beginning of the experiment (Fig 5b).
Fig 6. A. thaliana development in the presence of O. maius and of nine other fungi.

(a) Control plants (C) and plant-fungus co-cultures 30 days after inoculation; (b) plant biomass measurements (roots—grey bars—and aboveground portions—open bars) in the presence/absence of fungi. Note the strong plant biomass increase in the presence of some of the fungi tested. Bars represent the mean ±SD, n = 5 (each biological replicate represents the total biomass of 5 A. thaliana seedlings grown in an individual plate). Statistically significant differences (P<0.05) among treatments are indicated by different letters above the bars. Om, Oidiodendron maius; Mb, Meliniomyces bicolor; Mv, Meliniomyces variabilis; Re, Rhizoscyphus ericae; Lb, Laccaria bicolor; Sl, Suillus luteus; Cg, Cenococcum geophilum; Tc, Tulasnella calospora; Ch, Cladosporium herbarum; Tv, Trametes versicolor.
Fig 7. A. thaliana development in the presence of different fungi in the bipartite plate system.

(a) Control plants and plant-fungus co-cultures 15 days after inoculation; (b) plant biomass measurements (roots—grey bars—and aboveground portions—open bars) in the presence/absence of fungi. Note the strong plant biomass increase in the presence of some of the fungi tested. Bars represent the mean ±SD, n = 5 (each biological replicate represents the total biomass of 3 A. thaliana seedlings grown in an individual plate). Statistically significant differences (P<0.05) among treatments are indicated by different letters above the bars. Om, Oidiodendron maius; Mb, Meliniomyces bicolor; Mv, Meliniomyces variabilis; Re, Rhizoscyphus ericae; Lb, Laccaria bicolor; Cg, Cenococcum geophilum; Sl, Suillus luteus; Tc, Tulasnella calospora; Tv, Trametes versicolor; Ch, Cladosporium herbarum.
The same fungi tested in the non-compartmented square plates were also tested in bipartite plates, which allow only volatile molecules to be exchanged between the two partners (Fig 7). Similarly to O. maius WT, no clumped root phenotype was observed in this experimental setup for any of the fungi tested (Fig 7a), but a significant increase in plant biomass was recorded for most of them after 15 days of co-cultivation (though fungal growth promotion started to be visible after 7 days of co-cultivation), although to a different extent (Fig 7b). The most pronounced plant growth was induced by the four ericoid strains, by T. versicolor and by C. herbarum, whereas the three ECM fungi and the orchid mycorrhizal fungus were less effective in promoting plant growth (Fig 7b). These fungi were also those producing the lowest biomass in our experimental conditions (S1 Table). Pearson’s correlation test showed, with very few exceptions, a significant correlation (P<0.01) between plant and fungal biomass (S1 Table). As expected, a positive correlation was found between fungi producing a large biomass and the increase of above- and belowground plant portions, regardless of their ecological strategies. This experiment strengthened therefore the hypothesis that fungi induce plant growth in our in vitro experimental setup due to a nonspecific mechanism (i.e. CO2 emission).
Discussion
O. maius deeply influences development of a non-host plant
ERM fungi have been frequently found to associate with plant species outside the Ericaceae (see [36] and references therein), where they may form structures typical of fungal endophytes [39], but the role of these fungi when interacting with non-ericaceous plants is unclear. Here, we showed that O. maius significantly promotes growth of both aerial and root portions of A. thaliana in vitro. In addition, a peculiar clumped root phenotype was induced by this ericoid mycorrhizal fungus as a result of increased lateral root length and number associated to shortening of the primary root growth. Other fungi tested for comparison also promoted A. thaliana growth and influenced plant development, with some of them causing the peculiar clumped root phenotype, a behavior that may be quite widespread among soil fungi (this work; [19]). The ability to increase plant growth and to modify plant development and root architecture is largely documented for both PGPRs and PGPFs. The rhizobacterium Pseudomonas aeruginosa modifies root architecture in A. thaliana by increasing primary root length, number of lateral roots (LR) and root fresh weight [12] while Serratia marcescens, when co-cultured with A. thaliana, inhibited primary root elongation and induced LR in a distance-dependent manner [11]. Azospirillum brasilense inhibited root length while enhancing root hair formation in an inoculum concentration-dependent manner in Triticum aestivum [8]. Sirrenberg et al. [15] showed that the endophytic fungus Piriformospora indica increased root growth and branching in A. thaliana and Rai and Varma [51] showed that the same fungus increased root proliferation in Adhatoda visica. Enhanced A. thaliana root development and branching was reported also for three endophytic fungi isolated from Mentha aquatica [19]. Two free-living fungi, Trichoderma virens and Trichoderma atroviride, enhanced LR growth when co-cultured with A. thaliana [17]. Alteration of root morphology by mycorrhizal fungi is also well known, both in host and non-host plants. In Oryza sativa and Zea mays, fine and large lateral root formation is stimulated by Rhizophagus irregularis [23,52], while R. clarus and Gigaspora decipiens enhance root biomass in Aquilaria filaria and Dyera polyphylla [22]. ECM fungi also modified primary root growth and increased LR formation in plants like Populus, Picea abies, Pinus spp. and Cistus incanus, and they also modified root development and branching of the non-host A. thaliana [25–29,31]. Heller et al. [30] reported that LR production in Pinus sylvestris inoculated with Laccaria bicolor was faster than in non-inoculated plants. All these alterations of root architecture, due to interactions with soil microorganisms, could represent an advantage for plants by improving nutrient acquisition and tolerance to abiotic and biotic stress [53,54]. Ericoid mycorrhizal (ERM) fungi modify root development of their host plant and Villareal-Ruiz et al. [55] showed significant effects of a fungus of the Rhizoscyphus ericae aggregate on Vaccinium root development. The most profound effects were on total hair root length (eight-fold increase in the presence of the fungus) and the number of hair root tips (six-fold increase in the presence of the fungus). By contrast, the effect of ERM fungi on growth and development of non-host plants has never been investigated, to our knowledge.
Which molecules are involved in the interaction between O. maius and A. thaliana?
The results derived from plant-fungus co-cultures in compartmented and non-compartmented plates suggest that the two main phenotypes observed in A. thaliana, i.e. the increase in plant biomass and the clumped root phenotype, are induced by different compounds.
Several molecules produced by PGPFs have been described as being involved in plant growth promotion, such as cytokinins and gibberellins, brassinosteroids, oligosaccharines, bioamines, salicylic acid, and jasmonic acid [13,17,56]. In addition, PGPFs can produce molecules affecting hormone homeostasis in plants [13,14,56]. Some microorganisms, including ECM fungi, recruit the auxin signaling pathway to change plant root architecture [29,31], and LR induction has been specifically attributed to redistribution of auxin transporters at the root apex [29]. The inhibition of primary root growth by Trichoderma spp. has been attributed to an increase of the auxin content and to a disruption of the auxin response gradients in root tips induced by peptaibols, a class of linear peptide antibiotics [57].
For ERM fungi, Berta and Gianinazzi-Pearson [58] suggested either fungal auxin production or stimulation of plant hormone production as a possible cause of the substantial change in root length and number of hair roots in Calluna vulgaris seedlings infected with R. ericae.
Auxin was identified in culture filtrates of R. ericae strains [59], and here we also showed auxin production by O. maius. This aspect will be further discussed in the following section, but our experiments with A. thaliana DR5::GUS suggest that the plant phenotype in the presence of O. maius cannot be ascribed to accumulation of this plant hormone.
Volatile compounds can also influence root development. Splivallo et al. [31] mentioned the influence of ethylene, a volatile hormone, on A. thaliana LR development caused by the ECM fungi Tuber borchii and T. melanosporum. Ditengou et al. [32] demonstrated that LR branching induced by L. bicolor in A. thaliana could be ascribed to a specific VOC, a sesquiterpene. Garnica-Vergara et al. [60] showed that the 6-pentyl-2H-pyran-2-one (6-PP), a major VOC biosynthesized by Trichoderma spp., promoted plant growth and regulated root architecture by modulating the expression of auxin-transport proteins. Sánchez-López et al. [61] suggested that VOCs emitted by different rhizospheric and non-rhizospheric bacteria and fungi enhanced plant growth, photosynthesis efficiency, cytokinin levels, sugars accumulation and flowering. We tested whether the increase in plant biomass and the induction of the clumped root phenotype may be due to soluble or volatile compounds by using compartmented plates, where only volatile compounds emitted by O. maius could be perceived by A. thaliana. Whereas a significant increase in plant biomass was still observed in the compartmented plates, the peculiar clumped root phenotype was only induced when O. maius and A. thaliana were co-cultured in non-compartmented plates, suggesting a soluble diffusible signal. The experiments in compartmented plates in the presence and absence of activated charcoal, a VOCs trap [50], also suggested that VOCs likely play no role in the plant growth promotion induced by O. maius. This conclusion is supported by the fact that VOCs emission by O. maius was low in the growth conditions and at the developmental stage chosen for measurements. Although it cannot be excluded that O. maius VOCs emission could change under different environmental or physiological conditions, similarly to what was recently shown for Alternaria and Fusarium spp. [34], it should be noted that only two genes putatively coding for terpene synthases were found in the O. maius genome. By contrast, eight putative terpene synthase genes were found in L. bicolor [32].
Similarly to O. maius, a general increase in A. thaliana biomass was observed in the bipartite plates, where only volatile molecules could reach the plant, with all other fungi tested, whereas the clumped root phenotype was only observed for some fungi, including all ERM fungi but M. bicolor, in the non-compartmented petri plates, where soluble molecules could diffuse from the fungus to the plant.
The results with the compartmented plates in the presence of CO2 traps support the involvement of fungal produced CO2 in plant growth promotion, at least in our experimental setup, either through increased photosynthetic carbon fixation or as a developmental signal. Increased root biomass may in fact indirectly derive from higher carbon translocation from the photosynthetic leaves, but carbon dioxide is also known to stimulate primary root elongation and root branching [62,31]. Similarly, Kai and Piechulla [63] suggested that A. thaliana growth promotion by the rhizobacterium Serratia odorifera was due to the microbial CO2 accumulation in the co-cultivation plates. Thus, fungal derived CO2 is likely responsible for the increased plant biomass observed in the bipartite plates, whereas an as yet unidentified soluble diffusing molecule must be responsible for the clumped root phenotype, characterized by primary root shortening and increased LR length and number.
The O. maius GOGAT mutant suggests possible relationships between nitrogen metabolism and the clumped root phenotype in A. thaliana
Previous authors have reported a clumped root phenotype in A. thaliana in the presence of both bacteria and fungi [6,11,19,31]. Although auxin can induce primary root growth inhibition and has been suggested to be responsible for this particular phenotype in A. thaliana [31], our results with the O. maius GOGAT mutant seem to exclude the involvement of fungal-derived auxin. The O. maius GOGAT mutant, recently characterized by Khouja et al. [43], carries a partial deletion of the glutamate synthase (NADH-GOGAT EC 1.4.1.14) gene. Glutamate (Glu) biosynthesis in this mutant is therefore only mediated by the NADP-dependent glutamate dehydrogenase (NADP-GDH, EC 1.4.1.4), in contrast with the WT strain, where two different metabolic pathways (GOGAT and GDH) for Glu biosynthesis are working [43]. Glu has been reported to be a signalling molecule in roots [64–67] and it is the amino acid whose effects on root development in several plant species are most distinctive [64–67]. In A. thaliana, where these effects have been studied, Glu inhibited primary root growth and stimulated the outgrowth of lateral roots, producing a shorter and more branched root system [68]. We obtained, in our experimental setup, the same Glu effect on A. thaliana seedlings, with shorter and more branched root (S5 Fig). Interestingly, this is the phenotype we observed in A. thaliana in the presence of the O. maius WT strain, and it is tempting to speculate that the O. maius GOGAT mutant may be unable to induce the clumped root phenotype in A. thaliana because, due to the OmGOGAT deletion, it produces and releases into the medium a lower amount of glutamate. Glutamate and glutamine are also very important for maintenance and promotion of cell function because they are substrates for protein synthesis, regulate acid-base balance, provide nitrogen transport, and act as precursors of nucleotide and nucleic acid synthesis and of glutathione production [69]. It is therefore highly possible that the GOGAT mutation, by modifying Glu synthesis, may indirectly affect the biosynthesis of other important molecules, which in turn could be involved in the modification of root architecture. Further investigations of the metabolites differentially released by O. maius WT and the GOGAT mutant would help to verify changes in Glu secretion, or to identify other potential diffusible signals capable of inducing this peculiar clumped root phenotype in A. thaliana.
Another possible explanation for the different root growth pattern observed in the presence of the O. maius WT and the O. maius GOGAT mutant could be related to their different exploitation of nitrogen sources in the growing medium. Both nitrate and ammonium were available in the growth substrate, but they could be differently accessed by the WT and the mutant strains due to their different nitrogen pathways. The inhibitory effect of ammonium on primary root growth has been well documented [70], as well as the stimulation of LR growth by nitrate [65]. A different exploitation rate of the nitrogen sources available in the culture media by the O. maius WT and by the O. maius GOGAT mutant could lead to a different concentration of these two nitrogen sources in the co-culture plates and therefore to a different plant root phenotype.
Conclusions
A. thaliana is becoming a recognized model to analyse both mutualistic and non-mutualistic plant-microbe interactions [71–74], and several PGPFs have been shown to promote growth of this plant in vitro [6,17,19]. We have confirmed this observation for ERM fungi, as well as for other mycorrhizal and non-mycorrhizal fungi. However, it seems that the general increase in plant biomass was mainly caused, in our experimental setup, by carbon dioxide produced by the fungal mycelium and accumulated in the in vitro conditions, rather than by specific VOC signals. This finding should raise awareness for the interpretation of the results of plant-fungus co-cultivation experiments. Indeed a very recent review [50] pointed out that microbial CO2 in sealed co-culture plates may lead to a general increase of plant growth similar to what can be observed in the presence of microbial volatiles, suggesting that care should be taken when ecologically relevant functions of microbial VOCs are studied. On the other hand, from an ecological point of view, soil respiration is a key ecosystem process that releases carbon from the soil organic matter, thus playing an important role in global carbon cycling [75–76]. Carbon dioxide released through respiration by fungi closely associated with plants can be used by plants covering the soil surface for photosynthesis, leading to healthier plants, with an increased nutrient uptake capacity due to a more developed root system and with a higher aboveground biomass.
In addition to the general biomass increase due to fungal respiration, O. maius as well as about half of the other fungi tested induced in A. thaliana a peculiar clumped root phenotype, likely caused by diffusible soluble fungal compounds. Although the nature of these fungal compounds is as yet unknown, the inability of the O. maius GOGAT mutant to induce this root phenotype should help us to elucidate the mechanisms involved.
Supporting Information
(a) A. thaliana DR5::GUS control plants, (b) A. thaliana DR5::GUS plants co-cultivated with the O. maius WT (b) and with the O. maius GOGAT mutant (c). Staining was performed on aboveground and belowground portions of A. thaliana fresh tissues and stained tissues were observed and photographed using a Nikon Eclipse E400 optical microscope. The staining was observed in the root apex, in the vascular tissues of the primary root, in the lateral root primordia, and in some areas of the leaf margin. No differences for dye distribution and accumulation in plant tissues were observed in the absence or in the presence of fungi. Bars = 100 μm.
(TIF)
(a) Control plants and plant-fungus co-cultures 15 days after inoculation; (b) same as in (a) but plates were added with a VOCs trap compound (activated charcoal, AC) in the third compartment; (c) same as in (a) but plates were added with a CO2 trap compound [Ba(OH)2*8H2O, B] in the third compartment; (d) plant biomass measurements (roots—grey bars—and aboveground portions—open bars) in the presence/absence of the fungus and of the trap compounds. Note the strong plant biomass increase in the presence of the O. maius GOGAT mutant in all the conditions tested. Bars represent the mean ±SD, n = 5. Statistically significant differences (P<0.05) among treatments are indicated by different letters above the bars.
(TIF)
(a) A. thaliana plants growth in control plates (At) and in plates added with a CO2 trap compound, Ba(OH)2*8H2O (At-B) and with the same compound saturated with CO2 (At-B-CO2 saturated); (b) plant biomass measurements (roots—grey bars—and aboveground portions—open bars) in the presence/absence of the CO2 trap compound saturated or not with CO2. The saturation with CO2 of the barium hydroxide solution rescued the plant phenotype observed in the absence of CO2 trap compounds. Bars represent the mean ±SD, n = 5. Statistically significant differences (P<0.05) among treatments are indicated by different letters above the bars.
(TIF)
The clumped root phenotype started forming after 6 days of plant-fungus co-cultivation only in the presence of the O. maius WT strain.
(TIF)
Five days old A. thaliana plants grown for 4 days on the MS medium (C) and on the MS medium added with 25.6 mM Na-glutamate (Glu) using the non-compartmented square plate setup. Note the shorter and more branched root in the presence of glutamate.
(TIF)
Plant and fungal biomasses were measured in the bipartite plates and a correlation analysis was performed using the Pearson’s correlation test.
(DOCX)
Acknowledgments
We thank N. Colombano, A. Vayssières, L. Lanfranco, F. Venice and M. Mucciarelli for their help with some of the experiments and analysis; we also thank S. Wittulsky and R. Balestrini for helpful suggestions and A. Gillet, M. Buée and L. Fauchery for kindly providing some of the fungal strains used.
Data Availability
All relevant data are within the paper and its Supporting Information files. The sequence data from the O. maius Zn genome were produced by the US Department of Energy Joint Genome Institute and can be found at http://genome.jgi.doe.gov/Oidma1/Oidma1.home.html.
Funding Statement
S.C. was supported by a PhD fellowship from the Italian MIUR. The authors acknowledge financial support from local funding of the University of Turin and from the Laboratory of Excellence ARBRE (ANR-11-LABX-0002-01). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Berg G. Plant-microbe interactions promoting plant growth and health: perspectives for controlled use of microorganisms in agriculture. Appl Microbiol Biotechnol. 2009;84: 11–18. 10.1007/s00253-009-2092-7 [DOI] [PubMed] [Google Scholar]
- 2.Berg G, Grube M, Schloter M, Smalla K. The plant microbiome and its importance for plant and human health. Front Microbiol. 2014;5: 491 10.3389/fmicb.2014.00491 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Sukumar P, Legué V, VayssièRes A, Martin F, Tuskan GA, Kalluri UC. Involvement of auxin pathways in modulating root architecture during beneficial plant-microorganism interactions: Auxin, plant roots and plant-microbe interaction. Plant Cell Environ. 2013;36: 909–919. 10.1111/pce.12036 [DOI] [PubMed] [Google Scholar]
- 4.Lugtenberg B, Kamilova F. Plant-Growth-Promoting Rhizobacteria. Annu Rev Microbiol. 2009;63: 541–556. 10.1146/annurev.micro.62.081307.162918 [DOI] [PubMed] [Google Scholar]
- 5.Murali, Amruthesh KN, Sudisha J, Niranjana SR, Shetty HS. Screening for plant growth promoting fungi and their ability for growth promotion and induction of resistance in pearl millet against downy mildew disease. 2012;4(5): 30–36. [Google Scholar]
- 6.Salas-Marina MA, Silva-Flores MA, Cervantes-Badillo MG, Rosales-Saavedra MT, Islas-Osuna MA, Casas-Flores S. The Plant Growth-Promoting Fungus Aspergillus ustus Promotes Growth and Induces Resistance Against Different Lifestyle Pathogens in Arabidopsis thaliana. J Microbiol Biotechnol. 2011;21: 686–696. [DOI] [PubMed] [Google Scholar]
- 7.Ryu C-M, Hu C-H, Locy RD, Kloepper JW. Study of mechanisms for plant growth promotion elicited by rhizobacteria in Arabidopsis thaliana. Plant Soil. 2005;268: 285–292. [Google Scholar]
- 8.Dobbelaere S, Croonenborghs A, Thys A, Vande Broek A, Vanderleyden J. Phytostimulatory effect of Azospirillum brasilense wild type and mutant strains altered in IAA production on wheat. Plant Soil. 1999;212: 153–162. [Google Scholar]
- 9.López-Bucio J, Millán-Godínez M, Méndez-Bravo A, Morquecho-Contreras A, Ramírez-Chávez E, Molina-Torres J, et al. Cytokinin receptors are involved in alkamide regulation of root and shoot development in Arabidopsis. Plant Physiol. 2007;145: 1703–1713. 10.1104/pp.107.107953 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Zhang H, Kim M-S, Krishnamachari V, Payton P, Sun Y, Grimson M, et al. Rhizobacterial volatile emissions regulate auxin homeostasis and cell expansion in Arabidopsis. Planta. 2007;226: 839–851. 10.1007/s00425-007-0530-2 [DOI] [PubMed] [Google Scholar]
- 11.Shi C-L, Park H-B, Lee JS, Ryu S, Ryu C-M. Inhibition of primary roots and stimulation of lateral root development in Arabidopsis thaliana by the rhizobacterium Serratia marcescens 90–166 is through both auxin-dependent and -independent signaling pathways. Mol Cells. 2010;29: 251–258. 10.1007/s10059-010-0032-0 [DOI] [PubMed] [Google Scholar]
- 12.Ortiz-Castro R, Diaz-Perez C, Martinez-Trujillo M, del Rio RE, Campos-Garcia J, Lopez-Bucio J. Transkingdom signaling based on bacterial cyclodipeptides with auxin activity in plants. Proc Natl Acad Sci. 2011;108: 7253–7258. 10.1073/pnas.1006740108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Bent E. Induced Systemic Resistance Mediated by Plant Growth-Promoting Rhizobacteria (PGPR) and Fungi (PGPF) In: Tuzun S, Bent E, editors. Multigenic and Induced Systemic Resistance in Plants. Springer; US; 2006. pp. 225–258. http://link.springer.com/10.1007/0-387-23266-4_10 [Google Scholar]
- 14.Harman GE, Howell CR, Viterbo A, Chet I, Lorito M. Trichoderma species—opportunistic, avirulent plant symbionts. Nat Rev Microbiol. 2004;2: 43–56. 10.1038/nrmicro797 [DOI] [PubMed] [Google Scholar]
- 15.Sirrenberg A, Göbel C, Grond S, Czempinski N, Ratzinger A, Karlovsky P, et al. Piriformospora indica affects plant growth by auxin production. Physiol Plant. 2007;131: 581–589. 10.1111/j.1399-3054.2007.00983.x [DOI] [PubMed] [Google Scholar]
- 16.Kageyama SA, Mandyam KG, Jumpponen A. Diversity, Function and Potential Applications of the Root-Associated Endophytes In: Varma A, editor. Mycorrhiza. Berlin, Heidelberg: Springer Berlin Heidelberg; 2008. pp. 29–57. http://link.springer.com/10.1007/978-3-540-78826-3_2 [Google Scholar]
- 17.Contreras-Cornejo HA, Macías-Rodríguez L, Cortés-Penagos C, López-Bucio J. Trichoderma virens, a plant beneficial fungus, enhances biomass production and promotes lateral root growth through an auxin-dependent mechanism in Arabidopsis. Plant Physiol. 2009;149: 1579–1592. 10.1104/pp.108.130369 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Minerdi D, Bossi S, Maffei ME, Gullino ML, Garibaldi A. Fusarium oxysporum and its bacterial consortium promote lettuce growth and expansin A5 gene expression through microbial volatile organic compound (MVOC) emission. FEMS Microbiol Ecol. 2011;76: 342–351. 10.1111/j.1574-6941.2011.01051.x [DOI] [PubMed] [Google Scholar]
- 19.Dovana F, Mucciarelli M, Mascarello M, Fusconi A. In Vitro Morphogenesis of Arabidopsis to Search for Novel Endophytic Fungi Modulating Plant Growth. PloS One. 2015;10: e0143353 10.1371/journal.pone.0143353 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Smith SE. and Read DJ. Mycorrhizal Symbiosis. 2008. [Google Scholar]
- 21.Kaldorf M, Ludwig-Muller J. AM fungi might affect the root morphology of maize by increasing indole-3-butyric acid biosynthesis. Physiol Plant. 2000;109: 58–67. [Google Scholar]
- 22.Turjaman M, Tamai Y, Santoso E, Osaki M, Tawaraya K. Arbuscular mycorrhizal fungi increased early growth of two nontimber forest product species Dyera polyphylla and Aquilaria filaria under greenhouse conditions. Mycorrhiza. 2006;16: 459–464. 10.1007/s00572-006-0059-4 [DOI] [PubMed] [Google Scholar]
- 23.Gutjahr C, Casieri L, Paszkowski U. Glomus intraradices induces changes in root system architecture of rice independently of common symbiosis signaling. New Phytol. 2009;182: 829–837 10.1111/j.1469-8137.2009.02839.x [DOI] [PubMed] [Google Scholar]
- 24.Orfanoudakis M, Wheeler CT, Hooker JE. Both the arbuscular mycorrhizal fungus Gigaspora rosea and Frankia increase root system branching and reduce root hair frequency in Alnus glutinosa. Mycorrhiza. 2010;20: 117–126. 10.1007/s00572-009-0271-0 [DOI] [PubMed] [Google Scholar]
- 25.Karabaghli-Degron C, Sotta B, Bonnet M, Gay G, Le Tacon F. The auxin transport inhibitor 2,3,5-triiodobenzoic acid (TIBA) inhibits the stimulation of in vitro lateral root formation and the colonization of the tap-root cortex of Norway spruce (Picea abies) seedlings by the ectomycorrhizal fungus Laccaria bicolor. New Phytol. 1998;140: 723–733. [DOI] [PubMed] [Google Scholar]
- 26.Tranvan H, Habricot Y, Jeannette E, Gay G, Sotta B. Dynamics of symbiotic establishment between an IAA-overproducing mutant of the ectomycorrhizal fungus Hebeloma cylindrosporum and Pinus pinaster. Tree Physiol. 2000;20: 123–129. [DOI] [PubMed] [Google Scholar]
- 27.Niemi K, Vuorinen T, Ernstsen A, Häggman H. Ectomycorrhizal fungi and exogenous auxins influence root and mycorrhiza formation of Scots pine hypocotyl cuttings in vitro. Tree Physiol. 2002;22: 1231–1239. [DOI] [PubMed] [Google Scholar]
- 28.Rincón A, Priha O, Sotta B, Bonnet M, Le Tacon F. Comparative effects of auxin transport inhibitors on rhizogenesis and mycorrhizal establishment of spruce seedlings inoculated with Laccaria bicolor. Tree Physiol. 2003;23: 785–791. [DOI] [PubMed] [Google Scholar]
- 29.Felten J, Kohler A, Morin E, Bhalerao RP, Palme K, Martin F, et al. The ectomycorrhizal fungus Laccaria bicolor stimulates lateral root formation in poplar and Arabidopsis through auxin transport and signaling. Plant Physiol. 2009;151: 1991–2005. 10.1104/pp.109.147231 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Heller G, Lundén K, Finlay RD, Asiegbu FO, Elfstrand M. Expression analysis of Clavata1-like and Nodulin21-like genes from Pinus sylvestris during ectomycorrhiza formation. Mycorrhiza. 2012;22: 271–277. 10.1007/s00572-011-0402-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Splivallo R, Fischer U, Göbel C, Feussner I, Karlovsky P. Truffles regulate plant root morphogenesis via the production of auxin and ethylene. Plant Physiol. 2009;150: 2018–2029. 10.1104/pp.109.141325 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Ditengou FA, Müller A, Rosenkranz M, Felten J, Lasok H, van Doorn MM, et al. Volatile signalling by sesquiterpenes from ectomycorrhizal fungi reprogrammes root architecture. Nat Commun. 2015;6: 6279 10.1038/ncomms7279 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Müller A, Faubert P, Hagen M, Zu Castell W, Polle A, Schnitzler J-P, et al. Volatile profiles of fungi—chemotyping of species and ecological functions. Fungal Genet Biol FG B. 2013;54: 25–33. 10.1016/j.fgb.2013.02.005 [DOI] [PubMed] [Google Scholar]
- 34.Weikl F, Ghirardo A, Schnitzler J-P, Pritsch K. Sesquiterpene emissions from Alternaria alternata and Fusarium oxysporum: Effects of age, nutrient availability, and co-cultivation. Sci Rep. 2016;6: 22152 10.1038/srep22152 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Hung R, Lee S, Bennett JW. Arabidopsis thaliana as a model system for testing the effect of Trichoderma volatile organic compounds. Fungal Ecol. 2013;6: 19–26. [Google Scholar]
- 36.Perotto S, Martino E, Abbà S, Vallino M. 14 Genetic Diversity and Functional Aspects of Ericoid Mycorrhizal Fungi In: Hock B, editor. Fungal Associations. Berlin, Heidelberg: Springer Berlin Heidelberg; 2012. pp. 255–285. http://link.springer.com/10.1007/978-3-642-30826-0_14 [Google Scholar]
- 37.Sharples JM, Meharg AA, Chambers SM, Cairney JW. Symbiotic solution to arsenic contamination. Nature. 2000;404: 951–952. [DOI] [PubMed] [Google Scholar]
- 38.Cairney JWG, Meharg AA. Ericoid mycorrhiza: a partnership that exploits harsh edaphic conditions. Eur J Soil Sci. 2003;54: 735–740. [Google Scholar]
- 39.Chambers SM, Curlevski NJA, Cairney JWG. Ericoid mycorrhizal fungi are common root inhabitants of non-Ericaceae plants in a south-eastern Australian sclerophyll forest: Ericoid mycorrhizal fungi in non-Ericaceae plants. FEMS Microbiol Ecol. 2008;65: 263–270. 10.1111/j.1574-6941.2008.00481.x [DOI] [PubMed] [Google Scholar]
- 40.Daghino S, Martino E, Perotto S. Model systems to unravel the molecular mechanisms of heavy metal tolerance in the ericoid mycorrhizal symbiosis. Mycorrhiza. 2016;26: 263–274. 10.1007/s00572-015-0675-y [DOI] [PubMed] [Google Scholar]
- 41.Martino E, Turnau T, Girlanda M, Bonfante P, Perotto S. Ericoid mycorrhizal fungi from heavy metal polluted soils: their identication and growth in the presence of zinc ions. Mycol Res. 2000;104: 338–344. [Google Scholar]
- 42.Abbà S, Khouja HR, Martino E, Archer DB, Perotto S. SOD1-targeted gene disruption in the ericoid mycorrhizal fungus Oidiodendron maius reduces conidiation and the capacity for mycorrhization. Mol Plant-Microbe Interact MPMI. 2009;22: 1412–1421. 10.1094/MPMI-22-11-1412 [DOI] [PubMed] [Google Scholar]
- 43.Khouja HR, Daghino S, Abbà S, Boutaraa F, Chalot M, Blaudez D, et al. OmGOGAT-disruption in the ericoid mycorrhizal fungus Oidiodendron maius induces reorganization of the N pathway and reduces tolerance to heavy-metals. Fungal Genet Biol FG B. 2014;71: 1–8. 10.1016/j.fgb.2014.08.003 [DOI] [PubMed] [Google Scholar]
- 44.Murashige T, Skoog F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol Plant. 1962;15: 473–497. [Google Scholar]
- 45.Lobet G, Pagès L, Draye X. A Novel Image-Analysis Toolbox Enabling Quantitative Analysis of Root System Architecture. Plant Physiology. September 2011; 157:129–39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Gordon SA, Weber RP. Colorimetric estimation of Indolaaceticacid. Plant Physiol. 1951;26: 192–195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Hammer Ø, Harper DAT, Ryan PD. PAST—PAleontological STatistics software. Package for education and data analysis Paleontologia Electronic. 2001. [Google Scholar]
- 48.Krouk G, Lacombe B, Bielach A, Perrine-Walker F, Malinska K, Mounier E, et al. Nitrate-Regulated Auxin Transport by NRT1.1 Defines a Mechanism for Nutrient Sensing in Plants. Dev Cell. 2010;18: 927–937. 10.1016/j.devcel.2010.05.008 [DOI] [PubMed] [Google Scholar]
- 49.Sabatini S, Beis D, Wolkenfelt H, Murfett J, Guilfoyle T, Malamy J, et al. An Auxin-Dependent Distal Organizer of Pattern and Polarity in the Arabidopsis Root. Cell. 1999;99: 463–472. [DOI] [PubMed] [Google Scholar]
- 50.Piechulla B, Schnitzler J-P. Circumvent CO2 effects in volatile- based microbe-plant interactions. Trends Pl Science. 2016;21: 541–543. [DOI] [PubMed] [Google Scholar]
- 51.Rai M, Varma A. Arbuscular mycorrhiza-like biotechnological potential of Piriformospora indica, which promotes the growth of Adhatoda vasica Nees. Electron J Biotechnol. 2005;8: 1–8. [Google Scholar]
- 52.Kaldorf M, Ludwig-Muller J. AM fungi might affect the root morphology of maize by increasing indole-3-butyric acid biosynthesis. Physiol Plant. 2000;109: 58–67. [Google Scholar]
- 53.Glick BR, Cheng Z, Czarny J, Duan J. Promotion of plant growth by ACC deaminase-producing soil bacteria. Eur J Plant Pathol. 2007;119: 329–339.4 [Google Scholar]
- 54.Rommert AK, Oros-Sichler M, Lange T, Aust HJ, Schulz B, Boyle C. Growth promoting effect of endophytic colonization of larch seedlings (Larix decidua) with Cryptosporiopsis sp. and Phialophora. Book of abstracts, the seventh international mycological congress, University of Oslo. 2002.
- 55.Villarreal-Ruiz L, Anderson IC, Alexander IJ. Interaction between an isolate from the Hymenoscyphus ericae aggregate and roots of Pinus and Vaccinium. New Phytol. 2004;164: 183–192. [DOI] [PubMed] [Google Scholar]
- 56.Tsavkelova EA, Klimova SY, Cherdyntseva TA, Netrusov AI. Hormones and hormone-like substances of microorganisms: A review. Appl Biochem Microbiol. 2006;42: 229–235. [PubMed] [Google Scholar]
- 57.Shi W-L, Chen X-L, Wang L-X, Gong Z-T, Li S, Li C-L, et al. Cellular and molecular insight into the inhibition of primary root growth of Arabidopsis induced by peptaibols, a class of linear peptide antibiotics mainly produced by Trichoderma spp. J Exp Bot. 2016;. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Berta G, Gianinazzi-Pearson V. Influence of mycorrhizal infection on root development in Calluna vulgaris (L.) Hull seedlings [Pezizella ericae]. Mycorrhizae Physiol Genet Eds VGianinazzi-Pearson SGianinazzi. 1987; 673–676.
- 59.Gay G, Debaud JC. N Mycorrhizae Physiol Genet Eds VGianinazzi-Pearson SGianinazzi. 1986.
- 60.Garnica-Vergara A, Barrera-Ortiz S, Muñoz-Parra E, Raya-Gonzalez J, Mendez-Bravo A, Macıas-Rodrıguez L, et al. The volatile 6-pentyl-2H-pyran-2-one from Trichoderma atroviride regulates Arabidopsis thaliana root morphogenesis via auxin signaling and ETHYLENE INSENSITIVE 2 functioning. New Phytol. 2016;209: 1496–1512 10.1111/nph.13725 [DOI] [PubMed] [Google Scholar]
- 61.Sánchez-López AM, Baslam M, De Diego N, Muñoz FJ, Bahaji A, Almagro G, et al. Volatile compounds emitted by diverse phytopathogenic microorganisms promote plant growth and flowering through cytokinin action. Plant, Cell. Env. 2016. [DOI] [PubMed] [Google Scholar]
- 62.Crookshanks M, Taylor G, Dolan L. A model system to study the effects of elevated CO2 on the developmental physiology of roots: the use of Arabidopsis thaliana. J Exp Bot. 1998;49: 593–597. [Google Scholar]
- 63.Kai M, Piechulla B. Plant growth promotion due to rhizobacterial volatiles—An effect of CO2? FEBS Lett. 2009;583: 3473–3477. 10.1016/j.febslet.2009.09.053 [DOI] [PubMed] [Google Scholar]
- 64.Walch-Liu P, Liu L-H, Remans T, Tester M, Forde BG. Evidence that L-glutamate can act as an exogenous signal to modulate root growth and branching in Arabidopsis thaliana. Plant Cell Physiol. 2006;47: 1045–1057. 10.1093/pcp/pcj075 [DOI] [PubMed] [Google Scholar]
- 65.Walch-Liu P, Ivanov II, Filleur S, Gan Y, Remans T, Forde BG. Nitrogen Regulation of Root Branching. Ann Bot. 2006;97: 875–881. 10.1093/aob/mcj601 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Forde BG, Lea PJ. Glutamate in plants: metabolism, regulation, and signalling. J Exp Bot. 2007;58: 2339–2358. 10.1093/jxb/erm121 [DOI] [PubMed] [Google Scholar]
- 67.Forde BG. Glutamate signalling in roots. J Exp Bot. 2014;65: 779–787. 10.1093/jxb/ert335 [DOI] [PubMed] [Google Scholar]
- 68.Walch-Liu P, Forde BG. L-Glutamate as a Novel Modifier of Root Growth and Branching: What’s the Sensor? Plant Signal Behav. 2007;2: 284–286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Newsholme P, Procopio J, Lima MM, Pithon-Curi TC, Curi R. Glutamine and glutamate-their central role in cell metabolism and function. Cell Biochem Funct. 2003;21: 1–9. 10.1002/cbf.1003 [DOI] [PubMed] [Google Scholar]
- 70.Liu Y, Lai N, Gao K, Chen F, Yuan L, Mi G. Ammonium Inhibits Primary Root Growth by Reducing the Length of Meristem and Elongation Zone and Decreasing Elemental Expansion Rate in the Root Apex in Arabidopsis thaliana. PLoS One. 2013;8: e61031 10.1371/journal.pone.0061031 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Bulgarelli D, Rott M, Schlaeppi K, Ver Loren van Themaat E, Ahmadinejad N, Assenza F, et al. Revealing structure and assembly cues for Arabidopsis root-inhabiting bacterial microbiota. Nature. 2012;488: 91–95. 10.1038/nature11336 [DOI] [PubMed] [Google Scholar]
- 72.Lundberg DS, Lebeis SL, Paredes SH, Yourstone S, Gehring J, Malfatti S, et al. Defining the core Arabidopsis thaliana root microbiome. Nature. 2012;488: 86–90. 10.1038/nature11237 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Philippot L, Raaijmakers JM, Lemanceau P, van der Putten WH. Going back to the roots: the microbial ecology of the rhizosphere. Nat Rev Microbiol. 2013;11: 789–799. 10.1038/nrmicro3109 [DOI] [PubMed] [Google Scholar]
- 74.Martinuz A, Zewdu G, Ludwig N, Grundler F, Sikora RA, Schouten A. The application of Arabidopsis thaliana in studying tripartite interactions among plants, beneficial fungal endophytes and biotrophic plant-parasitic nematodes. Planta. 2015;241: 1015–1025. 10.1007/s00425-014-2237-5 [DOI] [PubMed] [Google Scholar]
- 75.Schlesinger WH, Andrews JA. Soil respiration and the global carbon cycle. Biogeochemistry. 2000;48: 7–20. [Google Scholar]
- 76.Gougoulias C, Clark JM, Shaw LJ. The role of soil microbes in the global carbon cycle: tracking the below-ground microbial processing of plant-derived carbon for manipulating carbon dynamics in agricultural systems. J Sci Food Agric. 2014;94: 2362–2371. 10.1002/jsfa.6577 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(a) A. thaliana DR5::GUS control plants, (b) A. thaliana DR5::GUS plants co-cultivated with the O. maius WT (b) and with the O. maius GOGAT mutant (c). Staining was performed on aboveground and belowground portions of A. thaliana fresh tissues and stained tissues were observed and photographed using a Nikon Eclipse E400 optical microscope. The staining was observed in the root apex, in the vascular tissues of the primary root, in the lateral root primordia, and in some areas of the leaf margin. No differences for dye distribution and accumulation in plant tissues were observed in the absence or in the presence of fungi. Bars = 100 μm.
(TIF)
(a) Control plants and plant-fungus co-cultures 15 days after inoculation; (b) same as in (a) but plates were added with a VOCs trap compound (activated charcoal, AC) in the third compartment; (c) same as in (a) but plates were added with a CO2 trap compound [Ba(OH)2*8H2O, B] in the third compartment; (d) plant biomass measurements (roots—grey bars—and aboveground portions—open bars) in the presence/absence of the fungus and of the trap compounds. Note the strong plant biomass increase in the presence of the O. maius GOGAT mutant in all the conditions tested. Bars represent the mean ±SD, n = 5. Statistically significant differences (P<0.05) among treatments are indicated by different letters above the bars.
(TIF)
(a) A. thaliana plants growth in control plates (At) and in plates added with a CO2 trap compound, Ba(OH)2*8H2O (At-B) and with the same compound saturated with CO2 (At-B-CO2 saturated); (b) plant biomass measurements (roots—grey bars—and aboveground portions—open bars) in the presence/absence of the CO2 trap compound saturated or not with CO2. The saturation with CO2 of the barium hydroxide solution rescued the plant phenotype observed in the absence of CO2 trap compounds. Bars represent the mean ±SD, n = 5. Statistically significant differences (P<0.05) among treatments are indicated by different letters above the bars.
(TIF)
The clumped root phenotype started forming after 6 days of plant-fungus co-cultivation only in the presence of the O. maius WT strain.
(TIF)
Five days old A. thaliana plants grown for 4 days on the MS medium (C) and on the MS medium added with 25.6 mM Na-glutamate (Glu) using the non-compartmented square plate setup. Note the shorter and more branched root in the presence of glutamate.
(TIF)
Plant and fungal biomasses were measured in the bipartite plates and a correlation analysis was performed using the Pearson’s correlation test.
(DOCX)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files. The sequence data from the O. maius Zn genome were produced by the US Department of Energy Joint Genome Institute and can be found at http://genome.jgi.doe.gov/Oidma1/Oidma1.home.html.