

Abstract
Determining bacterial gene expression during infection is fundamental to understand pathogenesis. In this study, we used dual RNA-seq to simultaneously measure P. aeruginosa and the murine host’s gene expression and response to respiratory infection. Bacterial genes encoding products involved in metabolism and virulence were differentially expressed during infection and the type III and VI secretion systems were highly expressed in vivo. Strikingly, heme acquisition, ferric-enterobactin transport, and pyoverdine biosynthesis genes were found to be significantly up-regulated during infection. In the mouse, we profiled the acute immune response to P. aeruginosa and identified the pro-inflammatory cytokines involved in acute response to the bacterium in the lung. Additionally, we also identified numerous host iron sequestration systems upregulated during infection. Overall, this work sheds light on how P. aeruginosa triggers a pro-inflammatory response and competes for iron with the host during infection, as iron is one of the central elements for which both pathogen and host fight during acute pneumonia.
Pseudomonas aeruginosa is an opportunistic Gram-negative respiratory pathogen of major clinical significance causing acute and chronic respiratory infections as well as bloodstream, urinary tract, burn wound, and surgical site infections1. According to the Center for Disease Control (CDC), 51,000 healthcare-associated P. aeruginosa infections occur in the United States each year2. This bacterium is highly versatile, able to survive in harsh environments, and forms robust biofilms that are difficult to eradicate on medical devices. P. aeruginosa is inherently highly resistant to antibiotics, and effective antimicrobial treatment of infections caused by this bacterium is often challenging. Multidrug-resistant Pseudomonas cause 13% of all P. aeruginosa infections and were given a threat level of “serious” in the CDC Antibiotic Resistance Threat report in 20132. Overall, P. aeruginosa carries one of the highest case fatality rates of all Gram-negative infections3. The success of this microorganism is associated with its highly adaptive genome, ability to survive and thrive in diverse environments and the expression of a wide variety of virulence factors4,5,6. It is therefore important to develop new therapeutics for the treatment of P. aeruginosa infections.
To accomplish this objective, it is crucial to better understand the simultaneous pathogen and host transcriptional responses during infection. Several studies have attempted to define the interaction dynamics between the pathogen and the host by studying gene and protein expression of either one or the other7,8,9,10,11. These studies have identified P. aeruginosa genes and virulence factors involved in bacterial attachment during colonization, nutrient acquisition, biofilm formation, or resistance to the immune system to facilitate persistence. Numerous studies have also identified how the host responds to infection12,13,14 and shown that pathogen-host interaction drives the selection and co-evolution of the two organisms15,16,17. More recently, Westermann et al. performed the first dual-RNA sequencing during tissue culture infection, demonstrating that dual-RNA sequencing is possible and a great tool to study pathogen-host interactions18.
However, none of these studies have characterized simultaneously the bacterial and host molecular factors involved in acute lung infection. It has been hypothesized that sequencing the RNA of both the host and the pathogen during in vivo infection would be feasible19,20,21 and was recently accomplished during Toxoplasma gondii infections22. Advances in DNA and protein sequencing during the past decade have now made dual analysis of RNA and protein expression possible, even in complex tissues. To characterize the armamentarium used in the battle between P. aeruginosa and the host during acute lung infection, we developed a methodology using dual RNA-seq to identify the transcriptional response of the pathogen and the host during infection.
Results
Dual genome-wide gene expression analysis during infection
In this study, we hypothesized that in vivo conditions influence gene expression in P. aeruginosa during infection and that the bacterium elicits transcriptional changes in the host. To test this hypothesis, we used a murine model of acute pneumonia to characterize the differences in P. aeruginosa gene expression during respiratory infection compared to in vitro growth. Using dual RNA-seq, we also gained insights into the effects of the bacterium on the host transcriptome. P. aeruginosa PAO1 was grown on Pseudomonas isolation agar (PIA) at 37 °C and 2 × 108 colony forming units (CFU) were used to infect outbred CD-1 mice by intranasal administration. The P. aeruginosa PAO1 grown on PIA was used to purify RNA for in vitro growth control. Mice were infected with PAO1, or PBS as control. After 16 h, mice were euthanized, lungs were removed and total RNA was purified. rRNA was depleted, and three libraries were prepared and sequenced using the Illumina MiSeq platform. Read data were mapped to P. aeruginosa and Mus musculus genomes independently with CLC Genomics.
A total of 87 million 2 × 75 bp reads were obtained. In samples from in vitro culture of PAO1 on PIA, 83% of the reads mapped to the pathogen, with a total coverage of 99.1% of the open reading frames of P. aeruginosa PAO1. In infected mice, 11% of the reads mapped to P. aeruginosa, covering 98.2% of the transcriptome, and 64% of the reads mapped to the mouse transcriptome. A total of 78% of the reads mapped to the non-infected mouse control. These data indicate that the methodology used in this study provides quality RNA in sufficient quantity to study the transcriptomes of both the pathogen and the host in the same sample.
P. aeruginosa gene expression changes during infection
Global changes in P. aeruginosa gene expression
EDGE statistical analysis was used to measure differences in gene expression. A total of 1,887 P. aeruginosa genes were found to be up-regulated and 1,325 down-regulated in vivo compared to in vitro growth on PIA with a p value < 0.001 (Table S1). To define the systems differentially regulated in each experimental condition, data were analyzed using a wide range of metagenomics analysis tools including GO term analysis of the biological processes, hypergeometric tests on annotations, PseudoCAP, KEGG pathways and STRING analysis23,24,25,26.
Cellular localization analysis of the gene products expressed by P. aeruginosa indicates that the proportion of differentially regulated genes whose products are localized in the cytoplasm, cytoplasmic membrane, periplasm, outer-membrane, and extracellular compartment did not significantly change in vitro and during infection (Fig. 1a). However, genes whose products are associated with outer-membrane vesicles (OMV) were significantly up-regulated during infection (29 genes associated with OMV formation or OMV-localized products in vitro against 163 genes in vivo, Table S2). OMVs are produced under stress conditions27 and various authors suggested that OMVs are involved in resistance to the host immune system by absorbing antimicrobial compounds and complement28, providing a decoy membrane in the environment29 and delivering virulence factors to host cells during infection30. Our data seem to indicate that OMV-associated proteins are also important during acute respiratory infection.
Figure 1. Characterization of P. aeruginosa gene expression in vitro and in vivo.

(a) Primary cellular localization of the products of the genes of P. aeruginosa PAO1 differentially regulated in vivo and in vitro. The percentage of the total number of genes whose expression was significantly up-regulated in vivo compared to in vitro are represented on the left; the percentage of genes whose expression was significantly up-regulated in vitro compared to in vivo are represented on the right. Genes were classified according to their known and predicted cellular localization. (b) Hypergeometric test enrichment analysis on annotation of P. aeruginosa PAO1 genes differentially regulated in vitro and in vivo. Data are represented as the percentage of genes (hits) in each class which expression was significantly up-regulated in vitro (green) or in vivo (blue). The subsets in which the number of hits was significant are denoted with asterisks (***p < 0.001). STRING analysis representing the 300 most highly upregulated P. aeruginosa PAO1 genes expressed in vitro (c) or in vivo (d). Gene products are represented with circles and known associations between each gene or gene products are represented with a connecting line. Nodes of genes participating in similar functions are circled in green (expressed in vitro) or in blue (expressed in vivo).
Changes in gene expression associated with bacterial metabolism
STRING and metabolic analyses indicate that DNA replication, arginine deiminase pathway, gluconeogenesis, cobalamin and choline metabolism, putrescine transport, and polysaccharide biosynthesis processes were significantly up-regulated during infection (Fig. 1b,d). These results suggest, as other have previously hypothesized11, that various amino-acids such as arginine, lysine and proline are available in vivo, and that P. aeruginosa also synthesizes many essential amino-acids during infection. Genes involved in phenylalanine and cellular amide catabolism, as well as cysteine and unsaturated fatty acid biosynthesis were significantly up-regulated in vitro. Expression of genes from the region PA21XX31, whose products are involved in carbohydrate metabolism, and alginate production, and of other genes involved in stress response such as hypochlorite, oxidative stress, and arsenic-containing substances, were significantly increased in vitro, potentially as a result of the stringent growth conditions encountered in PIA (Fig. 1b,c). Pyocyanin expression was also found to be up-regulated in vitro, likely as a result of the high magnesium chloride and potassium sulfate concentrations in PIA32. Overall these data suggest that P. aeruginosa adapts to changes in nutrient availability during infection, primarily as a result of its wide array of metabolic genes.
Non-coding RNA expression during infection
P. aeruginosa encodes within its genome numerous non-coding RNAs (ncRNA) involved in gene translation such as ribosomal RNAs and transfer RNAs. Other ncRNAs are involved in post-transcriptional regulation such as crcZ, nalA, phrS, prrF1, prrF2, rsmY, and rsmZ33,34,35. The expression of crcZ, prrF1, rsmY and rsmZ was detected both in vitro and during infection (RPKM values between 9 and 97,710) (Table S1). The largest number of transcripts detected for a ncRNA in both datasets belonged to crcZ. This ncRNA is involved in carbon catabolite repression33,36 and was significantly down-regulated during infection (15 fold). The expression of the gene encoding for the RNA chaperone Hfq (−1.4 fold) was also found to be significantly down-regulated during infection (Table S1). Altogether, these data suggest that decreased expression of hfq and crcZ might play a role in carbon utilization changes observed during infection. rsmZ was the smallest ncRNA detected (116 bp), suggesting that the method used in this study for RNA extraction and sequencing allows for detection of small ncRNA expression during infection. However, since the RNA extraction method used in this study is optimal for fragments over the size of 200 bp, specialized purification procedures designed for RNA fragments of smaller sizes would likely enhance the characterization of small ncRNAs in this type of samples.
Expression of virulence factors during infection
P. aeruginosa encodes a wide variety of virulence factors for host colonization and to establish infections. As expected, we observed changes in P. aeruginosa virulence gene expression during acute murine pneumonia. The expression of genes encoding P. aeruginosa type III (T3SS) and type VI (T6SS) secretion systems which translocate effectors across the bacterial cell envelope and the eukaryotic cell membrane was significantly increased in vivo (Fig. 2, Table S1). The expression of genes encoding the T3SS secreted effectors ExoS, ExoT, and ExoY was also up-regulated during infection. These two systems are involved in eukaryotic cell targeting and P. aeruginosa virulence in vivo37,38. It is important to note that T3SS and T6SS are usually known to be inversely controlled. Genes encoding the type I and type II secretion systems were more predominantly expressed in vitro (Fig. 2). Other genes encoding virulence factors such as the potent exotoxin A (toxA, 5.3 fold) or flagellar biosynthesis (1.62 to 3.55 fold) were up-regulated during infection. Unlike what was observed in burn wound infections11, the expression of the majority of the genes involved in LPS biosynthesis (PA3146-PA3159; 1.55 to 3.51 fold) were also up-regulated during infection. However, multiple genes associated with virulence, which would have been predicted to be expressed during infection, were found to be down-regulated in vivo. For instance, the genes encoding the LasR and RhlR systems involved in quorum sensing (QS) and regulation of numerous virulence factors were significantly down-regulated in vivo compared to in vitro conditions. Not surprisingly, 82 QS-regulated genes identified by Wagner et al.39 were also down-regulated, representing 46% of the QS regulon. This set includes genes involved in the biosynthesis of virulence factors such as alkaline protease (−1.67 to −4.70 fold), elastase (lasB, −154. fold) and protease IV (piv, −2.04 fold) (Table S1). This result might be due to differences in cell density between in vitro plate and in vivo growth conditions. Other non-QS regulated virulence factors were also down-regulated such as phospholipase C (plcB, −14.98 fold), alginate biosynthesis enzymes (−2.47 to −7.8 fold), and phenazines (−7.81 to −107.97 fold) (Table S1). These genes have been shown to be important for P. aeruginosa virulence in various models including keratitis40, burn wound infections41, or chronic respiratory infections42,43. Decreased expression of these virulence factors, together with the changes in T3SS and T6SS expression, could be associated with the down regulation during infection of the two-component system GacA/GacS (−2.0 and −1.5 fold respectively) which controls both acute and chronic virulence determinants such as biofilm formation, T6SS, or exopolysaccharide production44,45,46. The gene encoding RetS, which controls the planktonic lifestyle and coordinates the production of toxins and the T3SS through the small RNAs rsmZ and rsmY47, was significantly upregulated in vivo. Interestingly, the expression of the gene encoding for the lost adherence sensor LadS was not significantly changed during infection. Additionally, increases in expression of both T3SS and T6SS during infection could also be associated with population heterogeneity and changes in expression of other regulatory element, such as the up-regulation of the contact-dependent PpkA (3.13 fold) regulation of T6SS44. The results obtained here suggest that some virulence factors might not be required for acute pulmonary infections, but might be expressed in later stages, for instance, in association with biofilms in chronic lung infections44,48.
Figure 2. Differential expression of bacterial secretion systems in vitro and in vivo.

P. aeruginosa secretion systems expressed in vitro and in vivo were identified using the Kyoto Encyclopedia of Genes and Genomes (KEGG)24. Proteins encoded by genes up-regulated in vitro are indicated in green while proteins encoded by genes up-regulated in vivo are shown in blue.
M. musculus host response during infection
During infection, various host factors come into play to control and eliminate bacterial pathogens. To better understand the expression of these factors, EDGE statistical analysis was used to measure differences in gene expression in the lung of non-infected mice and mice infected with P. aeruginosa. The expression of 551 M. musculus genes increased and the expression of 151 decreased during infection compared to non-infected animals with a p value < 0.001. Meta-analysis of the data using hyper-geometric tests indicated that 80% of the genes with increased expression during infection were associated with pro-inflammatory response, chemotaxis, response to antigens, stress response, and regulation of cell proliferation. Granulocyte colony-stimulating factor (G-CSF or CSF3) was the most up-regulated eukaryotic gene during infection with a fold change of 706.23 (p = 8.93 × 10−104). The expression of genes encoding other cytokines were also significantly increased during infection, including the pro-inflammatory cytokines IL1, IL8, tumor necrosis factor alfa (TNF-α), interferon gamma (IFN-γ), and CSF2. CXC-motif chemokines (CXCL1, 2, 5, 9, 10 and 16), CC-motif chemokines (CCL2, 3, 4, 5, 7, 9, 11, 17 and 20), and the chemokine receptors IL1R2, IL4R, IL17R, and IL18RAP. The genes encoding FAS and TNFR2 were also highly expressed during infection, together with genes whose products are involved in TNF and NFκB signaling (Fig. 3, Table S3). These changes in chemokine expression were likely due to LPS and flagellin recognition by the Toll-like receptors (TLR) 2 and 449,50,51,52,53. The genes encoding CD14 (42.55 fold) and MyD88 (5.54 fold), two proteins involved in signaling trough TLR4, had increased expression levels during infection (Fig. 3, Table S3). The expression of the gene encoding TLR2, associated with susceptibility to P. aeruginosa infections54, was also found to be up-regulated during infection (10.85 fold). These data are in agreement with findings from numerous studies highlighting the importance of Toll-like receptors (TLR) in the recognition of P. aeruginosa during infection55,56,57,58,59. Interestingly, while T3SS effectors, such as ExoS have been shown to inhibit the expression of IL-1β60, the expression of this chemokine was up-regulated in vivo (13.69 fold). Altogether, chemokine expression is responsible for leukocyte differentiation, activation and recruitment, the production and release of pro-inflammatory cytokines, and play a major role in host defense against P. aeruginosa9. These chemokines can also be sensed by P. aeruginosa proteins such as OprF and affect gene expression in response to proinflammatory signals61.
Figure 3. Comparison between host proteomes and transcriptomes in response to P. aeruginosa infection.

Ingenuity Pathway Analysis (IPA) of canonical pathways significantly up-regulated during P. aeruginosa infection detected by transcriptome analysis. Murine genes significantly up- or down-regulated during infection were analyzed using IPA and classified according to their known and predicted function.
Battle for iron during infection
P. aeruginosa iron acquisition during infection
Iron is a vital nutrient involved in a wide range of biological processes and enzymatic functions. In low iron conditions, P. aeruginosa uses siderophores such as pyoverdine and pyochelin to chelate free extracellular Fe3+ 62,63,64. Pyoverdine binds Fe3+ to form ferripyoverdine. The fpvA gene encodes the ferripyoverdine TonB-dependent receptor (TBDR) FpvA. This receptor is bound to the anti-σ factor FpvR that sequesters the σ factors PvdS and FpvI to the bacterial membrane. Binding of ferripyoverdine to FpvA triggers the proteolysis of FpvR, releasing PvdS and FpvI. These σ factors then associate with the RNA polymerase and drive the expression of fpvA and the genes responsible for pyoverdine biosynthesis65,66. In our RNA-seq analysis, the expression of fpvA was increased in vivo (7.41 fold), while the expression levels of fpvI, fpvR and pvdS were unchanged (Table S1). This result suggests that in vivo, ferripyoverdine binding to FpvA triggers the release of PvdS and FpvI. As a result, pyoverdine expression levels are significantly up-regulated during infection (pvdADEFGHJLNOPQRT), supporting established in vitro models65,66. The gene expression of the alternative ferripyoverdine receptor, encoding FpvB, was also up-regulated in vivo (4.86 fold) (Table S1). Previous studies have shown the importance of the expression of this system in vivo; pyoverdine and FpvA mutants show decreased virulence compared to the parental strain in establishing murine pulmonary infections67,68. The expression of genes used to produce pyochelin, which has a lower affinity for iron than pyoverdine63, were also up-regulated in vivo. The pchABCD operon was expressed 40 to 50 fold more in vivo than in vitro and the genes pchREFG were expressed over 20 fold more in vivo. pchA was found amongst the genes most highly expressed in vivo (Table S1).
P. aeruginosa is also able to obtain heme from hemoproteins using the Has and Phu sytems69,70. In the Has system, the heme is extracted by the hemophore HasAp, which delivers the iron through the TBDR HasR. In the Phu system, the TBDR PhuR directly extracts the heme. In both pathways, heme is then transported to the cytoplasm by an ABC transporter, bound to a heme chaperone, PhuS, before being delivered to the heme oxygenase HemO71. In this RNA-seq study, hasAp was the most highly expressed gene in vivo, with differences in expression of 317.43 fold compared to in vitro settings. The genes hasD (228.00 fold), hasE (177.53 fold), hasR (72.16 fold), phuR (27.42 fold), phuS (15.00 fold), and hemO (6.39 fold) were also highly expressed in vivo (Table S1). HemO expression was shown to be necessary to drive the metabolic flow of heme into the cell, with PhuS participating in heme sensing and acting as a control valve for heme intake and degradation72. Overall, the Has heme acquisition system was the most highly expressed set of genes in vivo, suggesting the importance of iron acquisition from heme during infection. To validate these findings and determine the contribution of the Has system to P. aeruginosa pathogenesis, 6 weeks old female CD1 mice were infected intranasally with 108 CFU of PAO1ΔhasR or the parental strain PAO1. Sixteen hours post infection, mice were euthanized by pentobarbital injection and the bacterial load in the respiratory tract was determined. In this model of acute respiratory infection, the bacterial load of PAO1ΔhasR was 18.89 fold lower in the lung, and 6.8 fold lower in the nasal washes than the parental strain (Fig. 4). These data indicate that a mutant lacking the heme receptor HasR was strongly attenuated during infection. Overall, both RNA-seq and animal infection data suggests, as other have hypothesized in the past73, that pyoverdine and the Has/Phu systems are involved in acute respiratory infections and play a major role in P. aeruginosa pathogenesis (Fig. 5).
Figure 4. Role of HasR on bacterial burden in mice during acute infection.
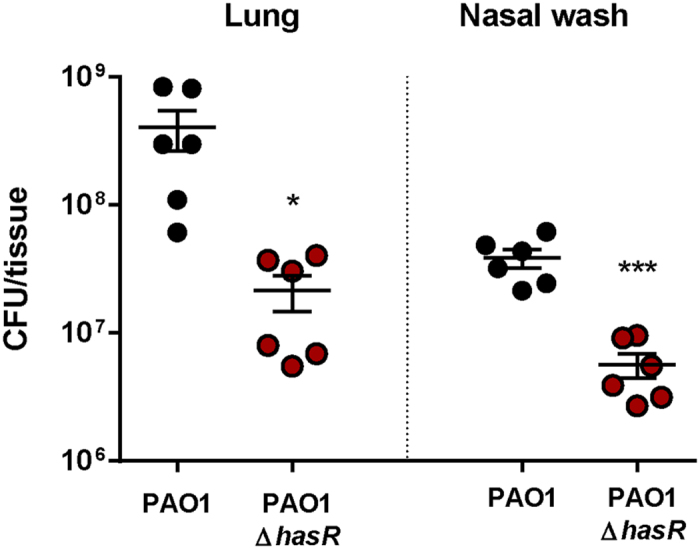
Mice were infected intranasally with 108 CFU of PAO1ΔhasR and the parental PAO1 strain. Bacterial burden were determined in the nares and lung 16 hours post infection. Lung were resuspended in 1 ml PBS and nasal washes were performed with 1 ml of PBS. Data are represented as number of CFU detected in the total volume of sample. Experiments were performed with 6 mice per group and four technical replicates. Data were analyzed using a two-tailed student t-test and the software package Graphpad Prism. Statistically significant differences are denoted with asterisks (*p < 0.05; ***p < 0.0001).
Figure 5. Iron scavenging and regulation in vivo.

(a) Macrophages acquire iron via transferrin receptor (TRF)-mediated endocytosis of transferrin or via recycling of senescent erythrocytes. Macrophages degrade heme using heme oxygenase to generate iron, biliverdin and carbon monoxide. Iron is then transferred to the cytoplasm by DMT1 and either stored using ferritin or released using ferroportin (FPN). (b) During infection, bacterial recognition by pattern recognition receptors triggers activation of Toll-like receptors (TLR) and secretion of pro-inflammatory cytokines. As a result, cell release hepcidin to control iron release by FPN. Proinflammatory cyctokines are also involved in the regulation of Nramp1, which together with FPN deprives internalized bacteria of iron. TNF-α, IL-1 and IL-6 also stimulate the storage of iron using ferritin. (c) During infection, bacteria can lyse erythrocytes using various hemolysins and toxins to release heme and hemoglobin. The host re-captures these iron-rich proteins using haptoglobin and hemopexin, while P. aeruginosa secretes hemophores such as HasA for heme scavenging. (d) The host uses lactoferrin and transferrin to limit iron availability during infection. P. aeruginosa secretes siderophores with a high iron affinity such as pyoverdine to circumvent iron sequestration. In response, the host releases siderocalin/lipocalin-2 to neutralize bacterial siderophores. Elements that were found to be up-regulated during P. aeruginosa infection in the lung are indicated in bold black letters. Elements that were unchanged are shown in grey.
Iron sequestration by the host during infection
One of the first lines of defense against bacterial infection is the sequestration of nutrients such as iron to inhibit bacterial growth. During acute infection, the host decreases iron release into the circulation via hepcidin, increases intracellular iron storage using ferritin, decreases extracellular non-heme iron levels via ceruloplasmin and lactoferrin, and decreases extracellular-heme iron levels using hemopexin and haptoglobin. Vertebrates also produce lipocalin-2, or neutrophil gelatinase-associated lipocalin (NGAL), which is secreted by neutrophils in response to infection, and captures bacterial siderophores. In this study, we observed that RNA levels of genes encoding for the expression of ferritin, ceruloplasmin, lactoferrin and haptoglobin are increased 4 to 20 fold during P. aeruginosa acute pneumonia (Table 1). However, no changes were observed in ferroportin, hepcidin, hemopexin, transferrin, or transferrin receptor expression. Additionally, the expression of norepinephrine, which increases iron availability, was decreased. These results seem to indicate that during P. aeruginosa infection, the host attempts to sequester iron from the pathogen to limit growth and dissemination (Fig. 5).
Table 1. Host iron-associated genes differentially regulated during P. aeruginosa infection.
| Gene name | Description | Fold up-regulated during infection | p-value |
|---|---|---|---|
| Lcn2 | Lipocalin 2 | 30.85 | 8.34E-04 |
| Ltf | Lactotransferrin | 14.34 | 2.16E-05 |
| Fth1 | Ferritin heavy chain 1 | 6.27 | 2.33E-03 |
| Cp | Ceruloplasmin | 4.82 | 2.83E-05 |
| Hba-a1 | Hemoglobin alpha, adult chain 1 | 2.11 | 3.12E-03 |
| Hba-a2 | Hemoglobin alpha, adult chain 2 | 2.96 | 3.03E-04 |
| Hbb-bs | Hemoglobin beta adult s chain | 2.07 | 1.98E-03 |
| Hbb-bt | Hemoglobin beta adult t chain | 2.54 | 1.89E-04 |
| Steap4 | Metalloreductase able to reduce Fe3+ to Fe2+ using NAD+ as receptor | 20.64 | 4.59E-05 |
| Slc39a14 | Soluble metal carrier protein | 17.85 | 2.69E-05 |
| Fbxl5 | F-box and leucin-rich repeat protein 5, iron sensor | 5.72 | 1.65E-05 |
| Picalm | Phosphatidylinositol binding clathri assembly protein, transferrin receptor internalization | 3.74 | 3.45E-05 |
| Scd2 | Stearoyl-coenzyme A desaturase 2, iron binding and oxydoreductase activity | −1.9 | 5.54E-05 |
| Mmp8 | Metallopeptidase 8 | 34.28 | 2.57E-05 |
| Mmp9 | Metallopeptidase 9 | 13.82 | 2.28E-05 |
Fold changes were calculated by comparing infected to non-infected mice. Fold changes greater than 1 indicate that the gene was up-regulated during infection compared to in vitro growth conditions. Fold changes inferior to −1 indicate that the gene was down-regulated during infection compared to in vitro growth conditions.
Discussion
In this study, we determined the changes in P. aeruginosa and host gene expression in the context of an acute murine pneumonia. As expected, we observed that P. aeruginosa expressed numerous virulence factors during infection, including T3SS and T6SS. P. aeruginosa was found to up-regulate outer membrane vesicle formation and modify its metabolism in response to infection. In the past, we observed that arginine metabolism responded to changes of temperature corresponding to the transition from the environment to the host7. In this dataset, we were able to confirm that arginine metabolism gene expression is also increased during infection, suggesting that P. aeruginosa adapts its metabolism in response to nutrient availability such as arginine. Overall, there is an important overlap between the data obtained here and gene expression changes observed in acute and chronic burn wounds infections11 (T3SS, T6SS, iron acquisition, amino-acid metabolism). This overlap suggests, as others have hypothesized, that while a core set of virulence genes are required to cause infection, P. aeruginosa adaptation to the lung requires the coordinated expression of genes involved in both virulence and metabolism11,74.
Infection with P. aeruginosa triggered a strong innate immune response and a cytokine storm mediated by TLR and MyD88 signaling. These data are in agreement with various studies showing that P. aeruginosa and outer-membrane vesicles are strong agonists of TLR pathogen-associated molecular pattern receptors75,76,77. Additionally, we observed that the expression of numerous host proteins involved nutritional immunity78 were increased during infection. In particular, proteins participating in iron sequestration were up-regulated during infection, including ferritin, ceruloplasmin, lactoferrin, and haptoglobin. Iron is an essential nutrient required by both mammalian hosts and pathogens, and the race for iron is undoubtedly crucial during infection. In the mammalian host, iron is not freely available and is bound to heme molecules found in hemoproteins or chelated by extracellular proteins such as lactoferrin and transferrin79. During infection, P. aeruginosa sequesters free iron using siderophores and scavenges iron from heme using the Has and Phu heme acquisition systems. One of the most striking observation in this study was the expression levels of the P. aeruginosa Has heme acquisition system during infection (over 300 fold up-regulated compared to in vitro conditions for hasAp). Pyoverdine and pyochelin siderophores, together with the Phu heme acquisition system were also up-regulated, suggesting that bacteria are starved for iron during infection. Up-regulation of both heme and siderophores systems in vivo is likely controlled by the master iron regulator Fur, as no significant changes in expression of the ncRNA prrF1,2 and prrH were observed during infection80. Additionally, both HasAp and PhuS could also be involved in heme sensing and might be up-regulated during infection in response to the presence of heme in the lung72. Numerous studies have demonstrated the importance of siderophores in bacterial virulence and colonization81, and authors have shown that pyoverdine production can be detected in the sputum of CF patients82. It has also been shown that P. aeruginosa interaction with epithelial cells in vitro in presence of iron is sufficient to up-regulate pyoverdine acquisition genes83. Siderophores have a high affinity for iron, can displace iron from host NGAL and transferrin for iron piracy, and have been shown to also act as toxins84. While others have suggested that other iron acquisition systems such as Has and Phu play a role in P. aeruginosa pathogenesis73, their use as an alternative iron acquisition system during infection has been underappreciated. The data presented here strongly suggests that heme acquisition by the Has/Phu systems play a crucial role for iron acquisition in vivo. Others have shown that iron and heme utilization by P. aeruginosa depends on disease progression, and suggested that heme acquisition is characteristic of chronic P. aeruginosa infections85,86. Isolates from chronic CF respiratory infections were shown to become more efficient in heme utilization by up-regulating PrrF and HemO expression86. In our dataset, heme acquisition also seems to play a major role in acute infection. For this reason, it would be interesting to further define the timing and synergy of heme and iron acquisition systems in P. aeruginosa during infection. While siderophores have been used as a therapeutic target87,88, it is crucial to determine the therapeutic potential of heme acquisition inhibitors in P. aeruginosa. Seminal studies have paved the way to understand the role of gene expression in P. aeruginosa pathogenesis11,41,74. Current advances in technology now allow us to study bacterial pathogenesis from a more systematic approach19. We anticipate that the methodology described in this study will further our understanding of pathogen-host interactions and be applicable in a wide range of systems to facilitate the development of novel therapeutics.
Experimental procedures
Bacterial strains and growth media
P. aeruginosa type strain PAO1 was originally obtained from Dr. Michael L. Vasil (University of Colorado). PAO1ΔhasR was obtained from Dr. Angela Wilks70 (University of Maryland). P. aeruginosa was grown in Pseudomonas isolation agar (Becton Dickinson, Franklin Lakes, NJ) for 16h at 37 °C before infection.
Infection model
P. aeruginosa PAO1 was grown as described above. Bacteria were swabbed off the plate and resuspended in phosphate buffered saline (PBS) at a concentration of 1010 colony forming units (CFU) per ml. A fraction of that suspension was centrifuged and resuspended in RNA protect to measure bacterial gene expression in vitro before infection. Twenty outbred six weeks old female CD1 mice (Charles River, Frederick, MD) were anesthetized by injection of ketamine and xylazine. Ten microliters of the bacterial suspension were administered to each nostril (108 CFU/mouse). Ten mice were infected with PAO1, and 10 mice were administered PBS vehicle control. Sixteen hours post-infection, mice were euthanized by exposure to carbon dioxide. Lungs were extracted from 4 mice per group and 3 immediately placed in bacterial RNA protect (Qiagen, Germantown, MD) to prevent RNA degradation. The remaining lung was homogenized in PBS and serial dilutions were plated to determine the load in the lung. Sixteen hours post infection, approximately 109 CFU were detected in the lung of each mouse (data not shown). To determine the role of hasR on P. aeruginosa pathogenesis, mice were infected as described above with 108 CFU of PAO1ΔhasR or the parental strain PAO1. After 16 hours of infection, mice were euthanized as described above, and nasal washes and lungs were obtained. Lungs were placed into 1 ml of PBS and homogenized. Nasal washes and lung homogenates were then diluted in PBS, and plated on Lysogeny agar (LA) to determine the number of viable bacteria. Data were analyzed using a two-tailed student t-test and the software package Prism 7 (GraphPad, La Jolla, CA). These experiments were performed in accordance with the National Institutes of Health guide for the care and use of laboratory animals. The protocols used were approved by the University of Virginia and West Virginia University Institutional Animal Care and Use Committees.
RNA purification and RNA-seq Illumina library preparation and sequencing
RNA was isolated from bacteria grown in vitro on PIA plates using RNeasy Mini Kit (Qiagen) as specified by the instructions of the manufacturer. Briefly, cells were pelleted by centrifugation at 8,000 g and RNA protect was pipetted off. Cell were lysed using 200 μl of TE lysozyme for 5 min at room temperature. Cells were then mixed with 700 μl of RLT and 500 μl of 70% ethanol. RNA was then applied to RNeasy purification columns and cleaned with RW1 and RPE buffers. RNA was eluted, treated with RNase-free DNase (Qiagen), and cleaned up on another RNeasy Mini column. RNA was isolated from the lung using the same kit with the following modifications: after removing the RNA protect, each lung was homogenized mechanically for 1 min in 667 μl of TE with 1 mg/ml of lysozyme. A total of 2.33 ml of RLT buffer was then added and tissue was homogenized again for 1 min. Samples were centrifuged at 12,000 g for 10 min and supernatant was transferred to a fresh tube. Five ml of 70% ethanol were added and RNA was purified using 12 RNeasy columns (2 columns per sample). Columns were cleaned with RW1 and RPE buffers, and RNA was eluted. The RNA extracted from the two columns per sample was pooled, treated with RNase-free DNase (Qiagen), and cleaned up using an RNeasy Mini column.
The resulting RNA was quantified on a Nanodrop ND-1000 (Nanodrop, Wilmington, DE). Next, the RNA integrity was assessed using Agilent BioAnalyzer RNA Pico chip (Agilent, Santa Clara, CA). All samples were then submitted to two rounds of eukaryotic and prokaryotic Ribo-zero rRNA depletion (Illumina, San Diego, CA) and reassessed for RNA integrity. rRNA depleted mRNA samples were then fragmented and prepared into libraries using Illumina TruSeq RNA library prep kit v2. Libraries were checked for quality control with KAPA qPCR QC assay (KAPA Biosystems, Wilmington, MA). The libraries were then sequenced on an Illumina MiSeq by the University of Virginia Department of Biology Genomics Core using Illumina MiSeq v3 2 × 75 bp reads. Three MiSeq lanes were used and each lane contained one biological sample of infected P. aeruginosa infected lung, one non-infected lung, and one P. aeruginosa in vitro sample. A total of three lanes were used to obtain for 81 million 2 × 75 bp reads for the three biological replicates of each group. Sequencing data were deposited to the Sequence Read Archive (SRA) and are available under the reference number SRP090213, BioProject number PRJNA343201.
RNA-seq bioinformatics analyses
RNA-seq reads were analyzed using the software CLC Genomics workbench 7.5.1 (Qiagen). The P. aeruginosa PAO1 genome and annotations were downloaded from the Pseudomonas Genome Database and Mus musculus genome was downloaded from NCBI. Reads were mapped against the gene regions of the two genomes using the following settings for mapping: mismatch cost = 2, insertion cost = 3, deletion cost = 3, length fraction = 0.8, similarity fraction = 0.8. RPKM values were generated using default parameters for CLC Genomics. Fold changes in gene expression and statistical analyses were performed using an Extraction of Differential Gene Expression (EDGE) test with the Bonferroni correction89.
Metabolic and gene set enrichment analyses
Genes with differences in gene expression with a p value < 0.001 were mapped on the Kyoto Encyclopedia of Genes and Genomes using KEGG Mapper v.2.524. Gene set enrichment analysis (GSEA) tests were performed using CLC Genomic workbench 7.5.123. Analyses were performed using gene ontology (GO) terms for biological processes25, with a total of 10,000 permutations. Primary cellular localization of the gene products was obtained from the P. aeruginosa PAO1 genome annotation downloaded from the Pseudomonas Genome Database. The 300 genes most highly up and down regulated in the P. aeruginosa PAO1 gene expression dataset were analyzed and grouped using the STRING 9.1 database of known and predicted interactions26 (string-db.org).
Additional Information
How to cite this article: Damron, F. H. et al. Dual-seq transcriptomics reveals the battle for iron during Pseudomonas aeruginosa acute murine pneumonia. Sci. Rep. 6, 39172; doi: 10.1038/srep39172 (2016).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Material
Acknowledgments
The authors would like to acknowledge AnhThu Nguyen at University of Virginia for her invaluable help with the preparation of the cDNA library and sample sequencing. The authors would also like to thank Dr. Erik L. Hewlett for facilitating the implementation and development of this project. Special thanks to Dr. Joanna Goldberg, Dr. Michael Schurr, and Emel Sen-Kilic who reviewed and edited the manuscript.
Footnotes
Author Contributions M.B. and F.H.D. were equally involved in experimental design, mouse infection and cell harvesting, RNA purification, preparation and next generation sequencing transcriptome analysis. Both authors also participated evenly in the preparation of this manuscript. A.O.-S. and A.W. participated in experimental design and the preparation of this manuscript.
References
- Sadikot R. T., Blackwell T. S., Christman J. W. & Prince A. S. Pathogen-host interactions in Pseudomonas aeruginosa pneumonia. Am. J. Respir. Crit. Care Med. 171, 1209–23 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Centers for Disease Control and Prevention, C. D. C. Antibiotic resistance threats in the United States, 2013. Atlanta CDC (2013). [Google Scholar]
- Aliaga L., Mediavilla J. D. & Cobo F. A clinical index predicting mortality with Pseudomonas aeruginosa bacteraemia. J Med Microbiol 51, 615–619 (2002). [DOI] [PubMed] [Google Scholar]
- Behrends V. et al. Metabolic adaptations of Pseudomonas aeruginosa during cystic fibrosis chronic lung infections. Env. Microbiol 15, 398–408 (2013). [DOI] [PubMed] [Google Scholar]
- Mathee K. et al. Dynamics of Pseudomonas aeruginosa genome evolution. Proc. Natl. Acad. Sci. USA 105, 3100–5 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wolfgang M. C. et al. Conservation of genome content and virulence determinants among clinical and environmental isolates of Pseudomonas aeruginosa. Proc Natl Acad Sci USA 100, 8484–8489 (2003). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barbier M. et al. From the environment to the host: re-wiring of the transcriptome of Pseudomonas aeruginosa from 22 degrees C to 37 degrees C. PLoS One 9, e89941 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bielecki P. et al. In-vivo expression profiling of Pseudomonas aeruginosa infections reveals niche-specific and strain-independent transcriptional programs. PLoS One 6, e24235 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lavoie E. G., Wangdi T. & Kazmierczak B. I. Innate immune responses to Pseudomonas aeruginosa infection. Microbes Infect 13, 1133–1145 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Son M. S., Matthews W. J., Kang Y., Nguyen D. T. & Hoang T. T. In vivo evidence of Pseudomonas aeruginosa nutrient acquisition and pathogenesis in the lungs of cystic fibrosis patients. Infect. Immun. 75, 5313–24 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turner K. H., Everett J., Trivedi U., Rumbaugh K. P. & Whiteley M. Requirements for Pseudomonas aeruginosa acute burn and chronic surgical wound infection. PLoS Genet 10, e1004518 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Campodonico V. L., Gadjeva M., Paradis-Bleau C., Uluer A. & Pier G. B. Airway epithelial control of Pseudomonas aeruginosa infection in cystic fibrosis. Trends Mol Med 14, 120–133 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hartl D. et al. Innate immunity in cystic fibrosis lung disease. J Cyst Fibros 11, 363–382 (2012). [DOI] [PubMed] [Google Scholar]
- Williams B. J., Dehnbostel J. & Blackwell T. S. Pseudomonas aeruginosa: host defence in lung diseases. Respirology 15, 1037–1056 (2010). [DOI] [PubMed] [Google Scholar]
- Barber M. F. & Elde N. C. Nutritional immunity. Escape from bacterial iron piracy through rapid evolution of transferrin. Science. 346, 1362–1366 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marvig R. L., Sommer L. M., Molin S. & Johansen H. K. Convergent evolution and adaptation of Pseudomonas aeruginosa within patients with cystic fibrosis. Nat Genet 47, 57–64 (2015). [DOI] [PubMed] [Google Scholar]
- Marvig R. L. et al. Within-host evolution of Pseudomonas aeruginosa reveals adaptation toward iron acquisition from hemoglobin. MBio 5, e00966–14 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Westermann A. J. et al. Dual RNA-seq unveils noncoding RNA functions in host–pathogen interactions. Nature 529, 496–501 (2016). [DOI] [PubMed] [Google Scholar]
- Westermann A. J., Gorski S. A. & Vogel J. Dual RNA-seq of pathogen and host. Nat Rev Microbiol 10, 618–630 (2012). [DOI] [PubMed] [Google Scholar]
- Schulze S., Henkel S. G., Driesch D., Guthke R. & Linde J. Computational prediction of molecular pathogen-host interactions based on dual transcriptome data. Front. Microbiol. 6, 65 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cloney R. Microbial genetics: Dual RNA-seq for host-pathogen transcriptomics. Nat. Rev. Genet. 17, 126–7 (2016). [DOI] [PubMed] [Google Scholar]
- Pittman K. J., Aliota M. T. & Knoll L. J. Dual transcriptional profiling of mice and Toxoplasma gondii during acute and chronic infection. BMC Genomics 15, 806 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tian L. et al. Discovering statistically significant pathways in expression profiling studies. Proc Natl Acad Sci USA 102, 13544–13549 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kanehisa M., Sato Y., Kawashima M., Furumichi M. & Tanabe M. KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res. 44, D457–62 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ashburner M. et al. Gene ontology: tool for the unification of biology. The Gene Ontology Consortium. Nat Genet 25, 25–29 (2000). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Franceschini A. et al. STRING v9.1: protein-protein interaction networks, with increased coverage and integration. Nucleic Acids Res 41, D808–15 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Macdonald I. A. & Kuehn M. J. Stress-induced outer membrane vesicle production by Pseudomonas aeruginosa. J Bacteriol 195, 2971–2981 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan T. T., Morgelin M., Forsgren A. & Riesbeck K. Haemophilus influenzae survival during complement-mediated attacks is promoted by Moraxella catarrhalis outer membrane vesicles. J Infect Dis 195, 1661–1670 (2007). [DOI] [PubMed] [Google Scholar]
- Vidakovics M. L. et al. B cell activation by outer membrane vesicles-a novel virulence mechanism. PLoS Pathog 6, e1000724 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kulp A. & Kuehn M. J. Biological functions and biogenesis of secreted bacterial outer membrane vesicles. Annu Rev Microbiol 64, 163–184 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Damron F. H. et al. Analysis of the Pseudomonas aeruginosa regulon controlled by the sensor kinase KinB and sigma factor RpoN. J Bacteriol 194, 1317–1330 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- KING E. O., WARD M. K. & RANEY D. E. Two simple media for the demonstration of pyocyanin and fluorescin. J. Lab. Clin. Med. 44, 301–7 (1954). [PubMed] [Google Scholar]
- Sonnleitner E., Abdou L. & Haas D. Small RNA as global regulator of carbon catabolite repression in Pseudomonas aeruginosa. Proc Natl Acad Sci USA 106, 21866–21871 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sonnleitner E. & Haas D. Small RNAs as regulators of primary and secondary metabolism in Pseudomonas species. Appl. Microbiol. Biotechnol. 91, 63–79 (2011). [DOI] [PubMed] [Google Scholar]
- Sonnleitner E., Romeo A. & Bläsi U. Small regulatory RNAs in Pseudomonas aeruginosa. RNA Biol. 9, 364–71 (2012). [DOI] [PubMed] [Google Scholar]
- Sonnleitner E. & Bläsi U. Regulation of Hfq by the RNA CrcZ in Pseudomonas aeruginosa carbon catabolite repression. PLoS Genet. 10, e1004440 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shaver C. M. & Hauser A. R. Relative contributions of Pseudomonas aeruginosa ExoU, ExoS, and ExoT to virulence in the lung. Infect Immun 72, 6969–6977 (2004). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mougous J. D. et al. A virulence locus of Pseudomonas aeruginosa encodes a protein secretion apparatus. Science 312, 1526–30 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wagner V. E., Bushnell D., Passador L., Brooks A. I. & Iglewski B. H. Microarray analysis of Pseudomonas aeruginosa quorum-sensing regulons: effects of growth phase and environment. J Bacteriol 185, 2080–2095 (2003). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caballero A., Thibodeaux B., Marquart M., Traidej M. & O’Callaghan R. Pseudomonas keratitis: protease IV gene conservation, distribution, and production relative to virulence and other Pseudomonas proteases. Invest Ophthalmol Vis Sci 45, 522–530 (2004). [DOI] [PubMed] [Google Scholar]
- Bielecki P., Glik J., Kawecki M. & Martins dos Santos V. a P. Towards understanding Pseudomonas aeruginosa burn wound infections by profiling gene expression. Biotechnol. Lett. 30, 777–90 (2008). [DOI] [PubMed] [Google Scholar]
- Jaffar-Bandjee M. C. et al. Production of elastase, exotoxin A, and alkaline protease in sputa during pulmonary exacerbation of cystic fibrosis in patients chronically infected by Pseudomonas aeruginosa. J. Clin. Microbiol. 33, 924–9 (1995). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hogardt M. & Heesemann J. Microevolution of Pseudomonas aeruginosa to a chronic pathogen of the cystic fibrosis lung. Curr. Top. Microbiol. Immunol. 358, 91–118 (2013). [DOI] [PubMed] [Google Scholar]
- Goodman A. L. et al. A signaling network reciprocally regulates genes associated with acute infection and chronic persistence in Pseudomonas aeruginosa. Dev. Cell 7, 745–54 (2004). [DOI] [PubMed] [Google Scholar]
- Lapouge K., Schubert M., Allain F. H.-T. & Haas D. Gac/Rsm signal transduction pathway of gamma-proteobacteria: from RNA recognition to regulation of social behaviour. Mol. Microbiol. 67, 241–53 (2008). [DOI] [PubMed] [Google Scholar]
- Lory S., Merighi M. & Hyodo M. Multiple activities of c-di-GMP in Pseudomonas aeruginosa. Nucleic Acids Symp. Ser. (Oxf). 51–2 (2009). [DOI] [PubMed] [Google Scholar]
- Bordi C. et al. Regulatory RNAs and the HptB/RetS signalling pathways fine-tune Pseudomonas aeruginosa pathogenesis. Mol Microbiol 76, 1427–1443 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goodman A. L. et al. Direct interaction between sensor kinase proteins mediates acute and chronic disease phenotypes in a bacterial pathogen. Genes Dev. 23, 249–59 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramphal R. et al. Control of Pseudomonas aeruginosa in the lung requires the recognition of either lipopolysaccharide or flagellin. J Immunol 181, 586–592 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raoust E. et al. Pseudomonas aeruginosa LPS or flagellin are sufficient to activate TLR-dependent signaling in murine alveolar macrophages and airway epithelial cells. PLoS One 4, e7259 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Skerrett S. J., Wilson C. B., Liggitt H. D. & Hajjar A. M. Redundant Toll-like receptor signaling in the pulmonary host response to Pseudomonas aeruginosa. Am. J. Physiol. Lung Cell. Mol. Physiol. 292, L312–22 (2007). [DOI] [PubMed] [Google Scholar]
- Ernst R. K. et al. Pseudomonas aeruginosa lipid A diversity and its recognition by Toll-like receptor 4. J. Endotoxin Res. 9, 395–400 (2003). [DOI] [PubMed] [Google Scholar]
- Erridge C., Pridmore A., Eley A., Stewart J. & Poxton I. R. Lipopolysaccharides of Bacteroides fragilis, Chlamydia trachomatis and Pseudomonas aeruginosa signal via toll-like receptor 2. J. Med. Microbiol. 53, 735–40 (2004). [DOI] [PubMed] [Google Scholar]
- Pene F. et al. Toll-like receptor 2 deficiency increases resistance to Pseudomonas aeruginosa pneumonia in the setting of sepsis-induced immune dysfunction. J Infect Dis 206, 932–942 (2012). [DOI] [PubMed] [Google Scholar]
- Faure K. et al. TLR4 signaling is essential for survival in acute lung injury induced by virulent Pseudomonas aeruginosa secreting type III secretory toxins. Respir. Res. 5, 1 (2004). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Adamo R., Sokol S., Soong G., Gomez M. I. & Prince A. Pseudomonas aeruginosa flagella activate airway epithelial cells through asialoGM1 and toll-like receptor 2 as well as toll-like receptor 5. Am J Respir Cell Mol Biol 30, 627–634 (2004). [DOI] [PubMed] [Google Scholar]
- Soong G., Reddy B., Sokol S., Adamo R. & Prince A. TLR2 is mobilized into an apical lipid raft receptor complex to signal infection in airway epithelial cells. J Clin Invest 113, 1482–1489 (2004). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lau G. W. et al. The Drosophila melanogaster toll pathway participates in resistance to infection by the gram-negative human pathogen Pseudomonas aeruginosa. Infect. Immun. 71, 4059–66 (2003). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hajjar A. M., Ernst R. K., Tsai J. H., Wilson C. B. & Miller S. I. Human Toll-like receptor 4 recognizes host-specific LPS modifications. Nat. Immunol. 3, 354–9 (2002). [DOI] [PubMed] [Google Scholar]
- Sutterwala F. S. et al. Immune recognition of Pseudomonas aeruginosa mediated by the IPAF/NLRC4 inflammasome. J Exp Med 204, 3235–3245 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu L. et al. Recognition of host immune activation by Pseudomonas aeruginosa. Science 309, 774–7 (2005). [DOI] [PubMed] [Google Scholar]
- Peek M. E., Bhatnagar A., McCarty N. A. & Zughaier S. M. Pyoverdine, the Major Siderophore in Pseudomonas aeruginosa, Evades NGAL Recognition. Interdiscip. Perspect. Infect. Dis. 2012, 843509 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brandel J. et al. Pyochelin, a siderophore of Pseudomonas aeruginosa: physicochemical characterization of the iron(III), copper(II) and zinc(II) complexes. Dalton Trans. 41, 2820–34 (2012). [DOI] [PubMed] [Google Scholar]
- Cornelis P. Iron uptake and metabolism in pseudomonads. Appl Microbiol Biotechnol 86, 1637–1645 (2010). [DOI] [PubMed] [Google Scholar]
- Redly G. A. & Poole K. Pyoverdine-mediated regulation of FpvA synthesis in Pseudomonas aeruginosa: involvement of a probable extracytoplasmic-function sigma factor, FpvI. J Bacteriol 185, 1261–1265 (2003). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Redly G. A. & Poole K. FpvIR control of fpvA ferric pyoverdine receptor gene expression in Pseudomonas aeruginosa: demonstration of an interaction between FpvI and FpvR and identification of mutations in each compromising this interaction. J Bacteriol 187, 5648–5657 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meyer J. M., Neely A., Stintzi A., Georges C. & Holder I. A. Pyoverdin is essential for virulence of Pseudomonas aeruginosa. Infect Immun 64, 518–523 (1996). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takase H., Nitanai H., Hoshino K. & Otani T. Impact of siderophore production on Pseudomonas aeruginosa infections in immunosuppressed mice. Infect Immun 68, 1834–1839 (2000). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ochsner U. A., Johnson Z. & Vasil M. L. Genetics and regulation of two distinct haem-uptake systems, phu and has, In Pseudomonas aeruginosa. Microbiology 146 (Pt 1), 185–198 (2000). [DOI] [PubMed] [Google Scholar]
- Smith A. D. & Wilks A. Differential contributions of the outer membrane receptors PhuR and HasR to heme acquisition in Pseudomonas aeruginosa. J Biol Chem 290, 7756–7766 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lansky I. B. et al. The cytoplasmic heme-binding protein (PhuS) from the heme uptake system of Pseudomonas aeruginosa is an intracellular heme-trafficking protein to the delta-regioselective heme oxygenase. J Biol Chem 281, 13652–13662 (2006). [DOI] [PubMed] [Google Scholar]
- O’Neill M. J. & Wilks A. The P. aeruginosa heme binding protein PhuS is a heme oxygenase titratable regulator of heme uptake. ACS Chem. Biol. 8, 1794–802 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cornelis P. & Dingemans J. Pseudomonas aeruginosa adapts its iron uptake strategies in function of the type of infections. Front Cell Infect Microbiol 3, 75 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turner K. H., Wessel A. K., Palmer G. C., Murray J. L. & Whiteley M. Essential genome of Pseudomonas aeruginosa in cystic fibrosis sputum. Proc. Natl. Acad. Sci. 112, 4110–4115 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao K., Deng X., He C., Yue B. & Wu M. Pseudomonas aeruginosa outer membrane vesicles modulate host immune responses by targeting the Toll-like receptor 4 signaling pathway. Infect Immun 81, 4509–4518 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- Blohmke C. J. et al. Innate immunity mediated by TLR5 as a novel antiinflammatory target for cystic fibrosis lung disease. J Immunol 180, 7764–7773 (2008). [DOI] [PubMed] [Google Scholar]
- Schnare M., Rollinghoff M. & Qureshi S. Toll-like receptors: sentinels of host defence against bacterial infection. Int Arch Allergy Immunol 139, 75–85 (2006). [DOI] [PubMed] [Google Scholar]
- Hood M. I. & Skaar E. P. Nutritional immunity: transition metals at the pathogen-host interface. Nat. Rev. Microbiol. 10, 525–37 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cornelissen C. N. & Sparling P. F. Iron piracy: acquisition of transferrin-bound iron by bacterial pathogens. Mol Microbiol 14, 843–850 (1994). [DOI] [PubMed] [Google Scholar]
- Oglesby-Sherrouse A. G. & Vasil M. L. Characterization of a Heme-Regulated Non-Coding RNA Encoded by the prrF Locus of. PLoS One 5, e9930 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Vos D. et al. Study of pyoverdine type and production by Pseudomonas aeruginosa isolated from cystic fibrosis patients: prevalence of type II pyoverdine isolates and accumulation of pyoverdine-negative mutations. Arch Microbiol 175, 384–388 (2001). [DOI] [PubMed] [Google Scholar]
- Martin L. W., Reid D. W., Sharples K. J. & Lamont I. L. Pseudomonas siderophores in the sputum of patients with cystic fibrosis. Biometals 24, 1059–1067 (2011). [DOI] [PubMed] [Google Scholar]
- Frisk A. et al. Transcriptome analysis of Pseudomonas aeruginosa after interaction with human airway epithelial cells. Infect. Immun. 72, 5433–8 (2004). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kirienko N. V., Ausubel F. M. & Ruvkun G. Mitophagy confers resistance to siderophore-mediated killing by Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 112, 1821–6 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Konings A. F. et al. Pseudomonas aeruginosa uses multiple pathways to acquire iron during chronic infection in cystic fibrosis lungs. Infect Immun 81, 2697–2704 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nguyen A. T. et al. Adaptation of iron homeostasis pathways by a Pseudomonas aeruginosa pyoverdine mutant in the cystic fibrosis lung. J Bacteriol 196, 2265–2276 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wurst J. M. et al. Identification of inhibitors of PvdQ, an enzyme involved in the synthesis of the siderophore pyoverdine. ACS Chem Biol 9, 1536–1544 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Möllmann U., Heinisch L., Bauernfeind A., Köhler T. & Ankel-Fuchs D. Siderophores as drug delivery agents: application of the “Trojan Horse” strategy. Biometals 22, 615–24 (2009). [DOI] [PubMed] [Google Scholar]
- Robinson M. D. & Smyth G. K. Small-sample estimation of negative binomial dispersion, with applications to SAGE data. Biostatistics 9, 321–32 (2008). [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.