

Abstract
Background
Streptococcus suis is an encapsulated Gram-positive bacterium and the leading cause of sepsis and meningitis in young pigs, resulting in considerable economic losses in the porcine industry. S. suis is considered an emerging zoonotic agent with increasing numbers of human cases over the last years. In the environment, both avirulent and virulent strains occur in pigs, with no evidence for consistent adapatation of virulent strains to the human host. Currently, there is an urgent need for a convenient, reliable and standardised animal model to rapidly assess S. suis virulence. Wax moth (Galleria mellonella) larvae have successfully been used in human and animal infectious disease studies. Here, we developed G. mellonella larvae as a model to assess virulence of S. suis strains.
Results
Fourteen isolates of S. suis belonging to different serotypes killed G. mellonella larvae in a dose-dependent manner. Larvae infected with the virulent serotype 2 strain, S. suis S3881/S10, were rescued by antibiotic therapy. Crucially, the observed virulence of the different serotypes and mutants was in agreement with virulence observed in piglets (Sus scrofa) and the zebrafish larval infection model. Infection with heat-inactivated bacteria or bacteria-free culture supernatants showed that in most cases live bacteria are needed to cause mortality in G. mellonella.
Conclusions
The G. mellonella model is simple, cost-efficient, and raises less ethical issues than experiments on vertebrates and reduces infrastructure requirements. Furthermore, it allows experiments to be performed at the host temperature (37 °C). The results reported here, indicate that the G. mellonella model may aid our understanding of veterinary microbial pathogens such as the emerging zoonotic pathogen S. suis and generate hypotheses for testing in the target animal host. Ultimately, this might lead to the timely introduction of new effective remedies for infectious diseases. Last but not least, use of the G. mellonella infection model to study S. suis virulence adheres to the principles of replacement, reduction and refinement (3Rs) and can potentially reduce the number of vertebrates used for experimental infection studies.
Electronic supplementary material
The online version of this article (doi:10.1186/s12866-016-0905-2) contains supplementary material, which is available to authorized users.
Keywords: Streptococcus suis, Galleria mellonella, Virulence, Reduction and refinement (3Rs), Infection model
Background
Streptococcus suis is an emerging zoonotic Gram-positive bacterial pathogen [1, 2] and the leading cause of sepsis and meningitis in piglets. Infections with S. suis have been reported worldwide throughout the pig production industry and have led to considerable economic losses [3, 4]. Recently, S. suis isolates causing more rapid and severe infections in both humans and pigs have been reported [5], raising concerns about the emergence of more virulent strains [6–10].
The most common symptoms caused by virulent S. suis in pigs and humans are meningitis, endocarditis and streptococcal toxic shock-like syndrome [11–13]. In spite of the importance of S. suis as a porcine pathogen and zoonotic agent, relatively little is known about epidemiology and virulence mechanisms leading to carriage and invasive disease. Genetic analysis of virulence and pathogenicity is not trivial as S. suis possesses multifactorial virulence factors [1]. The matter is further complicated because natural populations are characterized by high rates of recombination [14, 15], leading to many different genotypes [4]. A generally accepted virulence factor of S. suis is the capsular polysaccharide [16]. The critical role of the capsular polysaccharide in virulence and pathogenicity of S. suis has been demonstrated in multiple independent studies [17–19]. Of the 33 serotypes of the capsular polysaccharide identified so far, serotype 2 is the most frequently associated with invasive disease [6, 20]. Other serotypes often associated with porcine disease include serotype 1, 9, and 14 [21, 22]. S. suis colonises the upper respiratory tract of pigs and the intestine [23, 24]. Healthy pigs carrying S. suis are a source for S. suis transmission to the rest of the pigs in the herd [23]. Currently, it is not possible to distinguish a virulent strain from a non-pathogenic strain using molecular markers [1] highlighting the need for better genomic markers of virulence and/or animal models to rapidly assess the virulence of different S. suis isolates.
Pigs and mice have been successfully employed for S. suis virulence studies [5, 25]. However, pigs and mice have economical, logistic and ethical disadvantages over non-mammalian infection models. Recently, the zebrafish (Danio rerio) larval model has been validated for assessing virulence of S. suis strains [26]. G. mellonella larvae have been successfully utilised to study the virulence of other important pathogens, including Streptococcus spp., and to characterize existing and novel antibacterials [27–32]. G. mellonella larvae are exempt from ethical legislation. The larvae are cheap to purchase allowing experiments to be performed with a large number of samples. Furthermore, unlike other non-mammalian models, e.g. Caenorabditis elegans, Drosophila melanogaster, or zebrafish larvae, G. mellonella allows experiments to be performed at 37 °C, the body temperature of the natural hosts. Last but not least, the insect immune system exhibits both humoral and cellular components, and in some aspects the immune response is similar to the innate immune response of mammals [33]. For example, both insect hemocytes and mammalian neutrophils engulf and kill pathogens, and both types of cell produce superoxide, using similar p47 and p67 proteins [34]. Nevertheless, G. mellonella larvae are distantly related to pigs, and it is important to establish their predictive value for virulence in the natural host. A first step towards this goal is to compare the virulence of a range of clinical and non-clinical S. suis strains that differ in virulence in the natural host. Here, we evaluated the use of G. mellonella larvae as an in vivo model to assess virulence of S. suis isolates.
Methods
Bacteria and growth conditions
S. suis strains used in this study are listed in Table 1. S. suis were stored at −80 °C in 20% (v/v) glycerol. S. suis strains were cultured routinely in Mueller Hinton broth (MHB) (Difco, USA) at 37 °C in the presence of 5% CO2 without shaking. For each strain studied, a calibration curve was generated for absorbance measurements of bacterial cultures at 600 nm (OD600) by plating aliquots on MH agar (1% v/v) to calculate the bacterial colony forming units per milliliter (CFU/ml) (data not shown).
Table 1.
Virulence of bacterial strains used in this study. HV stands for highly virulent, V stands for virulent, WV stands for weakly virulent, and AV stands for avirulent. The virulence in pigs is described in [35] and the scale of virulence in pig infection model is as described in [35]. Strains leading to more than 50% mortality of G. mellonella larvae post-infection with 107 CFU/ml were considered virulent, and strains leading to less than 50% mortality of G. mellonella larvae were considered weakly virulent
| Strain | Serotype | Virulence in pigs [35] | Virulence in G. mellonella | Clinical source | Reference/source |
|---|---|---|---|---|---|
| 6388 | 1 | HV | V | Organs | Laboratory collection [45] |
| 6555/NCTC 428 | 1 | V | WV | ||
| T15 | 2 | AV | WV | Tonsil | Laboratory collection [46] |
| S735R2 | 2 | WV | WV | Unknown | Laboratory collection [35] |
| P1/7 | 2 | V | V | Unknown | Laboratory collection [35] |
| 3881/S10 | 2 | V | V | [47] | |
| (J28) S10 cpsΔEF | 2 | AV | WV | [17] | |
| 15965 | 3 | ND | V | ||
| 5213 | 4 | ND | V | ||
| 8039 | 7 | ND | V | CNS | Laboratory collection [40] |
| 7709 | 9 | ND | V | Bacteraemia | Laboratory collection [40] |
| C132 | 9 | ND | WV | Brain/septicemia | Laboratory collection [40] |
| 7998 | 9 | ND | V | Joint | Laboratory collection [40] |
| 8186 | 9 | ND | WV | Tonsil | Laboratory collection [40] |
| 5128/22083 | 9 | ND | V |
V virulent, HV highly virulent, WV weakly virulent, AV avirulent, ND not determined
G. mellonella larvae
G. mellonella larvae in their final instar stage were purchased (UK Waxworms Ltd, Sheffield, UK), and stored in the dark at 15 °C, and used within 14 days. Larvae were separated by weight, and only larvae between 0.2 and 0.3 g were used for experiments. Unless otherwise stated, all experiments in G. mellonella were performed using 15 larvae per group and in most cases repeated using larvae from a different batch to give n = 30. Larvae were injected with 20 μL of bacterial suspension, antibiotic solution, or phosphate buffered saline (PBS) in the left posterior proleg using Terumo Myjector 29G insulin syringes (VWR International). The syringes were changed between treatments with different strains. Two negative control groups were included in every experiment; one group was not injected to control for background larval mortality (no manipulation control) and the other group (uninfected control) was injected with PBS to control for the possible effect of physical trauma on mortality. Larvae were stored in Petri dishes in the dark at 37 °C with 5% CO2 for up to 144 h post-infection (p.i.) and inspected every 24 h for survival; larvae were considered dead if they did not move after shaking of the petri dish.
Virulence of S. suis in G.mellonella
Infection of G. mellonalla larvae with S. suis strains was performed as described previously for other pathogens [27, 28]. Larvae were infected in the left posterior proleg with 20 μl inocula of different S. suis strains containing between 1 × 105 CFU/mL to 1 × 108 CFU/mL. Survival of larvae was recorded at 24 h intervals for 144 h p.i.. A heat-killed S. suis inoculum of > 1 × 108 CFU/ml incubated at 99 °C for 30 min was used as a negative control in the infection experiments. In some experiments S. suis culture supernatants were prepared by centrifugation of overnight cultures (i.e. > 1 × 108 CFU/ml) at 11 000 g for 30 min and injected into the posterior proleg of groups of larvae as described above. The absence of viable S. suis in culture supernatants and heat inactivated cultures was confirmed by plating on MH agar and incubation at 37 °C for 24 h.
Antibiotic treatment
Antibiotic stock solutions were prepared in 1× PBS. For studies of antibiotic efficacy, an inoculum of 1 × 107 CFU/mL of S. suis 3881/S10 was used. A single treatment with antibiotics was administered 2 h p.i. via the posterior proleg.
Statistical analysis
Survival data were plotted using the Kaplan–Meier method and comparisons between groups were made using the log-rank test using GraphPad Prism v 5.03. In all comparisons with the negative control, it was the uninfected control (rather than the un-manipulated control) that was used. In all tests, P ≤ 0.05 was considered significant. The lethal dose leading to 50% mortality (LD50) at different time points was determined by plotting the percentage of surviving larvae following infection with different inocula, and non-linear curve fitting using GraphPad Prism v 5.03.
Results
Galleria mellonella larvae are sensitive to infection by Streptococcus suis
The virulence of S. suis isolates (Table 1) that had been assessed for virulence in a piglet infection model [35] was assessed in G. mellonella larvae by inoculation with 20 μL of suspensions containing 1 × 105 to 1 × 108 CFU/ml and incubation at 37 °C. Notably, isolates reported as virulent in pigs (6388, 6555/NCTC 428, P1/7, 3881/S10, 7997) [35] and in a zebrafish larval model (3881/S10, 6555/NCTC 428) [26] showed dose-dependent virulence in G. mellonella larvae (Figs. 1 and 2, Table 1). Isolates reported as weakly virulent or avirulent in piglets (T15, S735R2) showed attenuated virulence in G. mellonella larvae compared to the virulent strains, e.g. 3881/S10 (Figs. 1 and 2, Table 1). One generally accepted virulence factor of S. suis is the capsular polysaccharide, and its critical role in virulence and pathogenicity has been demonstrated in multiple independent studies [17, 18, 26]. A mutant lacking the capsular polysaccharide, the serotype 2 mutant cpsΔEF [17], showed more than 20 times attenuated virulence to G. mellonella larvae compared to the wild type, 3881/S10 (Figs. 1 and 2, Additional file 1: Figure S1, and Table S1). After 24 h p.i. 5 × 106 CFU/ml of the wt 3881/S10 lead to 50% mortality, whereas more than 1×108 CFU/ml of the mutant lacking the capsular polysacharyde, cpsΔEF, are needed for 50% mortality 24 h p.i. These results confirm the applicability of the G. mellonella larvae infection model to study S. suis virulence factors. Furthermore, we studied isolates for which virulence data in piglet infection models has not been reported. G. mellonella larvae were infected with isolates from different clinical sources (central nervous system, bacteraemia, speticemia, joins, and tonsils, Table 1). Six of the studied isolates (15965, 5213, 8039, 7709, 7998, 5128/22083) showed dose-dependent virulence (Figs. 1 and 2). The serotype 9 isolates 5128/22083 and 7709 were among the most virulent isolates tested in G. mellonella larvae (Figs. 1 and 2 and Additional file 1: Figure S1, and Table 1 and Additional file 1: Table S1). Another serotype 9 isolate, 7998, showed virulence comparable to the virulence of the serotype 2 isolates 3881/S10 and P1/7. Isolates 5213 (serotype 4) and 8039 (serotype 7) showed weaker virulence, comparable to the non-capsulated serotype 2 mutant cpsΔEF (Figs. 1 and 2 and Additional file 1: Figure S1 and Table 1 and Additional file 1: Table S1). The serotype 3 isolate 15965 showed virulence comparable to the virulence of P1/7 (serotype 2). Isolates C132 and 8186 (both serotype 9) were weakly virulent (Figs. 1 and 2 and Additional file 1: Figure S1, and Table 1 and Additional file 1: Table S1).
Fig. 1.

Dose dependent virulence of different S. suis strains belonging to serotype 1 to 3 (Table 1). Survival data were plotted using the Kaplan–Meier method using GraphPad Prism v 5.03. The dotted lines represent the standard error. The observed virulence is in agreement with virulence observed in a piglet infection model (Table 1)
Fig. 2.
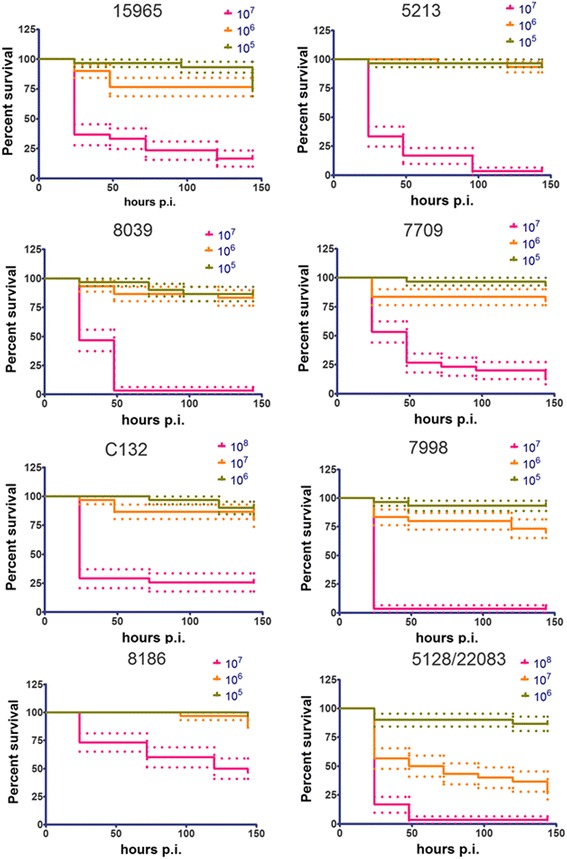
Dose dependent virulence of different S. suis strains belonging to serotype 4, 7 and 9 (Table 1). Survival data were plotted using the Kaplan–Meier method using GraphPad Prism v 5.03. The dotted lines represent the standard error. The observed virulence is in agreement with virulence observed in a piglet infection model (Table 1)
Melanisation in G. mellonella is thought to be a key part of the defence against a range of pathogens [29]. Melanin is deposited around microbes within the hemolymph where it is believed to facilitate pathogen killing [29]. Over the course of S. suis infection, G. mellonella larvae showed obvious signs of melanisation similar to melanisation induced by infection with other pathogens (Additional file 1: Figure S2).
The culture supernatants or heat-treated extracts of specific strains induced mortality in G. mellonella larvae
To examine the effect of bacteria-free culture supernatants on larval survival, 20 μl was injected through the left posterior proleg. For most strains the supernatant of an overnight culture (more than 1 × 108 CFU/ml) did not significantly affect larval survival (Additional file 1: Figure S3) indicating that the exotoxin suilysin or endotoxins released into the growth medium were not causing mortality. The exception was strain 7709 for which the bacteria-free supernatant showed significantly increased mortality compared to the PBS control (p value 0.0085) indicating toxicity due to secreted factor(s). The heat inactivated inoculum (containing both the heat-treated cells and heat-treated supernatant) of 7709 did not lead to a significantly different effect than the PBS control. These indicate that the factor(s) in the 7709 supernatant causing mortality of the G. mellonella larvae is temperature sensitive.
The heat-inactivated inoculum of 8039 and 5213 significantly increased larval mortality compared to the PBS injected control group (p values of 0.0481 and < 0.0001, respectively). The virulence of the live bacteria of 8039 and 5213 is comparable to the non-capsulated serotype 2 mutant cpsΔEF and to the weakly virulent serotype 2 isolate S735 (Figs. 1 and 2 and Additional file 1: Figure S1, and Table 1 and Additional file 1: Table S1).
Antibiotic treatment rescues larvae infected with S. suis 3881/S10
G. mellenella larvae have been used to study antibiotic treatment for different bacterial and fungal infections [27, 35, 36]. To assess the utility of G. mellonella larvae to study antibacterial therapies against S. suis, we confirmed that antibiotic therapy could rescue larvae from S. suis infection. Larvae were infected with 1 × 107 CFU of the S. suis strain 3881/S10, a well characterised serotype 2 strain previously shown to be virulent in pigs and used in our previous research [37, 38, 39]. At 2 h p.i. the larvae were treated with doxycycline, amoxicillin and ampicillin (25 mg/kg). Additional control groups were assessed for the toxicity of the antibiotic treatment (data not shown). Treatment with 25 mg/kg of each of the antibiotics rescued the larvae, and 144 h p.i. there was 100% survival rate of the larvae treated with the three different antibiotics, compared to infected larvae that were not treated with antibiotics (data not shown).
Deletion of the two-component system CiaRH reduced S. suis virulence in G. mellonella larvae
The two component system (TCS) CiaRH contributes to the virulence of S. suis serotype 2 [40]. Deletion of the ciaRH operon results in a lower survival rate in a bactericidal assay compared to the wild-type. Furthermore, the mutant showed reduced virulence in mice and pigs in vivo, and in a zebrafish larval model of S. suis infection [26]. In order to assess whether reduced virulence of S. suis strain S10 ΔciaRH would also be observed in G. mellonella larvae, we compared the survival of larvae infected with 1 × 107 CFU/ml of the mutant strain S10 ΔciaRH or the wild type 3881/S10. Infection of larvae by the ΔciaRH strain resulted in a significantly higher survival rate compared to infection with the 3881/S10 wild-type strain, (p < 0.0001) (Fig. 3). Thus, the observed results in G. mellonella larvae are in agreement with results obtained with the same two isolates in mouse and pig infection studies, and in zebrafish larval infection model. Taken together these results indicate that G. mellonella larvae can be used to assess virulence of porcine S. suis strains reproducibly and in a biologically meaningful way.
Fig. 3.

a Survival rate of G. mellonela larvae injected with 105 – 107 CFU/ml of the S10 ΔciaRH mutant. b Comparison of the survival rate of larvae infected with 107 of S10 ΔciaRH mutant and the wild-type 3881/S10 strain. Survival data were plotted using the Kaplan–Meier method and comparisons between groups were made using the log-rank test using GraphPad Prism v 5.03. The dotted lines represent standard error
S. suis suilysin mutant shows reduced virulence in G. mellonella larvae
S. suis produces a secreted membrane damaging toxin, suilysin suggested which is to contribute to virulence [41]. An allelic replacement insertion mutant of S. suis strain P1/7 lacking functional sly, (Δsly), is not haemolytic and unlike the wild type P1/7 does not efficiently kill J774.2 cellsin vitro [40]. Furthermore, the deletion of suilysin mutation prevented S. suis from killing mice via the intraperitoneal route of infection [40]. In pig infection models, the mean survival times of the wild type P1/7 and the suilysin mutant Δsly were not significantly different [42]. In G. mellonella infection with 107 CFU/ml of the Δsly mutant showed slightly more than 2 times attenuated virulence 24 p.i. compared to the wild type P1/7 (Fig. 4). However, 48 to 144 h p.i. the virulence of the Δsly mutant was attenuated more than 1.5 fold compared to the wild-type strain P1/7. The attenuated virulence of the Δsly mutant in G. mellonella larvae compared to the wild-type stain P/7 is in agreement with the effects observed in J774.2 cells in vitro and in the interperitoneal mice infection model.
Fig. 4.

a Survival rate of G. mellonela larvae injected with 106 – 108 CFU/ml of the P1/7 Δsly mutant. b Comparison of the survival rate of larvae infected with 107 of P1/7 Δsly mutant and the wild-type P1/7. Survival data were plotted using the Kaplan–Meier method and comparisons between groups were made using the log-rank test using GraphPad Prism v 5.03
Discussion
The use of insect larvae for virulence testing of pathogens is not regulated by legislation and represents an alternative to experimental infection of rodents or pigs. Alternative infection models are important in order to comply with the efforts to reduce, refine, and replace animal experiments (the 3Rs). G. mellonella larvae have been used as infection models for other important pathogens [29] and to study the efficacy of antibacterial and antifungal therapies [28, 43, 38] due to the general similarities between the wax worm innate immune system and the innate immune response of higher organisms [33]. As an infection model G. mellonella offers a number of technical advantages over mammalian infection models (e.g. high-throughput, ease of handling, simple housing requirements, the possibility to purchase a large number of larvae at a relatively low price, no legal or ethical issues etc.). Importantly, experiments with G. mellonella larvae unlike experiments with other non-mammalian models (e.g. zebrafish larvae, C. elegans, or D. melanogaster) can be performed at the body temperature of the mammalian natural host, 37 °C. The immune system of insects, including G. mellonella larvae, is very similar to the innate immune response of mammals [33]. Therefore, results obtained using insects can serve as a starting point to study pathogenesis and generate hypotheses to be further tested in vertebrate models.
The present study aimed to validate G. mellonella larvae as an alternative model to investigate the virulence of S. suis. S. suis is an important zoonotic pathogen and significant cause of meningitis and sepsis in pigs and humans [6]. Clinical isolates of S. suis were shown to kill G. mellonella larvae in a dose-dependent manner. Furthermore, similar to infection with other pathogens, infection of G. mellonella larvae with virulent S. suis strains triggered melanisation (Additional file 1: Figure S2). Melanisaiton is part of the immune response in G. mellonella considered a key step of the defense against a range of pathogens [29]. Melanin is deposited around microbes within the haemolymph and is thought to facilitate pathogen killing. The strains with greater reported virulence in pigs were also more virulent in G. mellonella supporting the predictive value of the G. mellonella infection model. The correlation in virulence of S. suis isolates in pigs and G. mellonella was demonstrated using published virulence data, i.e. no pigs were required for the experiments reported here [35]. Additionally, we report on the virulence of several S. suis strains that have not been studied in pigs or rodent infection models. Live strains of S. suis were required to significantly increase larval mortality, the exception being strains 8039 and 5213 which still increased larval mortality when heat-killed prior to injection at the highest dose. Nevertheless, the effect of larval survival infection with 8039 or 5213 is comparable to the effect of the weakly virulent serotype 2 S735 or the non-capsulated serotype 2 mutant cpsΔEF.
The bacteria-free culture supernatants of S. suis did not induce significant death of larvae compared to the control except in the case of the serotype 9 strain 7709, which was among the most virulent strains tested in the Galleria infection model (Additional file 1: Figure S1, and Table S1). This might be due to higher expression of suilysin, the pore-forming toxin produced by most pathogenic strains of S. suis, which is carbon catabolite repressed by glucose when grown in vitro [44].
Conclusion
The use of invertebrate hosts such as G. mellonella, D. melanogaster and C. elegans to study microbial pathogenesis and host responses have contributed substantially to biomedical research over the last decades. In this study, we describe the utility of G. mellonella larvae to study the virulence of the zoonotic pathogen S. suis. Relatively little is known about the pathogenic mechanisms of S. suis. Therefore, the infection model reported here serves as a useful starting point for assessing the strain virulence or the impact of genetic mutations on virulence of S. suis. Interestingly, a good correlation between the pathogenicity of S. suis in G. mellonella and the virulence observed in pigs and humans, and in other infection models such as zebrafish larval infection model and mice infection model was observed. Furthermore, the infection model described here also has the potential to be used to evaluate novel therapies for in vivo activity against S. suis and help prioritise candidates for testing in mammalian models.
Acknowledgements
We thank the Department of Biology, Maynooth University, Co. Kildare, Ireland for the provided training in using G. mellonella larvae as an infection model.
Funding
This work was funded by H2020 IF AND – Antibacterial (Nano)medicines Development (AND 659–121, NV) and a travel grant by the Dutch Cystic Fibrosis Foundation (NV).
Availability of data and materials
All data supporting the findings in this manuscript is included here or in the Supplementary material.
Authors’ contribution
NV, KK, and JW designed the experiments. NV performed the experiments and analysed the data. NV wrote the manuscript. JW and KK corrected the manuscript. All authors have read and approved the manuscript.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
n/a.
Ethics approval and consent to participate
n/a.
Abbreviations
- 3Rs
Replacement, reduction, and refinement of animal experiments
- AV
Avirulent
- CFU
Colony forming units
- CNS
Central nervous system
- MHB
Mueller Hinton broth
- p.i.
Post-infection
- PBS
Phosphate buffered saline
- TCS
Two-component system
- V
Virulent
- WV
Weakly virulent
Additional file
LD50s of tested strains. Figure S2. Infection with S. suis triggers melanisation in G. mellonella larvae. Figure S3. Effect of cell-free supernatant and heat-inactivated inocula on G. mellonella larvae survivial. Table S1. LD50s of tested strains. (DOC 2250 kb).
Contributor Information
Nadya Velikova, Email: nadya.velikova@wur.nl.
Kevin Kavanagh, Email: kevin.kavanagh@nuim.ie.
Jerry M. Wells, Email: jerry.wells@wur.nl
References
- 1.Fittipaldi N, Segura M, Grenier D, Gottschalk M. Virulence factors involved in the pathogenesis of the infection caused by the swine pathogen and zoonotic agent Streptococcus suis. Future Microbiol. 2012;7(2):259–279. doi: 10.2217/fmb.11.149. [DOI] [PubMed] [Google Scholar]
- 2.Gottschalk M, Xu J, Calzas C, Segura M. Streptococcus suis: a new emerging or an old neglected zoonotic pathogen? Future Microbiol. 2010;5(3):371–391. doi: 10.2217/fmb.10.2. [DOI] [PubMed] [Google Scholar]
- 3.Staats JJ, Feder I, Okwumabua O, Chengappa MM. Streptococcus suis: past and present. Vet Res Commun. 1997;21(6):381–407. doi: 10.1023/A:1005870317757. [DOI] [PubMed] [Google Scholar]
- 4.Goyette-Desjardins G, Auger JP, Xu J, Segura M, Gottschalk M. Streptococcus suis, an important pig pathogen and emerging zoonotic agent-an update on the worldwide distribution based on serotyping and sequence typing. Emerg Microbes Infect. 2014;3(6):e45. doi: 10.1038/emi.2014.45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Dominguez-Punaro MC, Segura M, Plante MM, Lacouture S, Rivest S, Gottschalk M. Streptococcus suis serotype 2, an important swine and human pathogen, induces strong systemic and cerebral inflammatory responses in a mouse model of infection. J Immunol. 2007;179(3):1842–1854. doi: 10.4049/jimmunol.179.3.1842. [DOI] [PubMed] [Google Scholar]
- 6.Feng Y, Zhang H, Wu Z, Wang S, Cao M, Hu D, Wang C. Streptococcus suis infection: an emerging/reemerging challenge of bacterial infectious diseases? Virulence. 2014;5(4):477–497. doi: 10.4161/viru.28595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Mai NT, Hoa NT, Nga TV, le Linh D, Chau TT, Sinh DX, Phu NH, Chuong LV, Diep TS, Campbell J, et al. Streptococcus suis meningitis in adults in Vietnam. Clin. Infect. Dis. 2008;46(5):659–667. doi: 10.1086/527385. [DOI] [PubMed] [Google Scholar]
- 8.Suankratay C, Intalapaporn P, Nunthapisud P, Arunyingmongkol K, Wilde H. Streptococcus suis meningitis in Thailand. Southeast Asian J. Trop. Med. Public Health. 2004;35(4):868–876. [PubMed] [Google Scholar]
- 9.Tang J, Wang C, Feng Y, Yang W, Song H, Chen Z, Yu H, Pan X, Zhou X, Wang H, et al. Streptococcal toxic shock syndrome caused by Streptococcus suis serotype 2. PLoS Med. 2006;3(5) doi: 10.1371/journal.pmed.0030151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Yu H, Jing H, Chen Z, Zheng H, Zhu X, Wang H, Wang S, Liu L, Zu R, Luo L, et al. Human Streptococcus suis outbreak, Sichuan, China. Emerg Infect Dis. 2006;12(6):914–920. doi: 10.3201/eid1206.051194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Arends JP, Zanen HC. Meningitis caused by Streptococcus suis in humans. Rev Infect Dis. 1988;10(1):131–137. doi: 10.1093/clinids/10.1.131. [DOI] [PubMed] [Google Scholar]
- 12.Wangkaew S, Chaiwarith R, Tharavichitkul P, Supparatpinyo K. Streptococcus suis infection: a series of 41 cases from Chiang Mai University Hospital. J Infect. 2006;52(6):455–460. doi: 10.1016/j.jinf.2005.02.012. [DOI] [PubMed] [Google Scholar]
- 13.Watkins EJ, Brooksby P, Schweiger MS, Enright SM. Septicaemia in a pig-farm worker. Lancet. 2001;357(9249):38. doi: 10.1016/S0140-6736(00)03570-4. [DOI] [PubMed] [Google Scholar]
- 14.Weinert LA, Chaudhuri RR, Wang J, Peters SE, Corander J, Jombart T, Baig A, Howell KJ, Vehkala M, Valimaki N, et al. Genomic signatures of human and animal disease in the zoonotic pathogen Streptococcus suis. Nat Commun. 2015;6:6740. doi: 10.1038/ncomms7740. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Marini E, Palmieri C, Magi G, Facinelli B. Recombination between Streptococcus suis ICESsu32457 and Streptococcus agalactiae ICESa2603 yields a hybrid ICE transferable to Streptococcus pyogenes. Vet Microbiol. 2015;178(1–2):99–104. doi: 10.1016/j.vetmic.2015.04.013. [DOI] [PubMed] [Google Scholar]
- 16.Segura M. Fisher scientific award lecture - the capsular polysaccharides of group b streptococcus and streptococcus suis differently modulate bacterial interactions with dendritic cells. Can J Microbiol. 2012;58(3):249–260. doi: 10.1139/w2012-003. [DOI] [PubMed] [Google Scholar]
- 17.Smith HE, Damman M, van der Velde J, Wagenaar F, Wisselink HJ, Stockhofe-Zurwieden N, Smits MA. Identification and characterization of the cps locus of Streptococcus suis serotype 2: the capsule protects against phagocytosis and is an important virulence factor. Infect Immun. 1999;67(4):1750–1756. doi: 10.1128/iai.67.4.1750-1756.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lecours MP, Segura M, Lachance C, Mussa T, Surprenant C, Montoya M, Gottschalk M. Characterization of porcine dendritic cell response to Streptococcus suis. Vet Res. 2011;42:72. doi: 10.1186/1297-9716-42-72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Meijerink M, Ferrando ML, Lammers G, Taverne N, Smith HE, Wells JM. Immunomodulatory effects of Streptococcus suis capsule type on human dendritic cell responses, phagocytosis and intracellular survival. PLoS One. 2012;7(4):e35849. doi: 10.1371/journal.pone.0035849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Wertheim HF, Nghia HD, Taylor W, Schultsz C. Streptococcus suis: an emerging human pathogen. Clin Infect Dis. 2009;48(5):617–625. doi: 10.1086/596763. [DOI] [PubMed] [Google Scholar]
- 21.Allgaier A, Goethe R, Wisselink HJ, Smith HE, Valentin-Weigand P. Relatedness of Streptococcus suis isolates of various serotypes and clinical backgrounds as evaluated by macrorestriction analysis and expression of potential virulence traits. J Clin Microbiol. 2001;39(2):445–453. doi: 10.1128/JCM.39.2.445-453.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Gottschalk M, Segura M, Xu J. Streptococcus suis infections in humans: the Chinese experience and the situation in North America. Anim Health Res Rev. 2007;8(1):29–45. doi: 10.1017/S1466252307001247. [DOI] [PubMed] [Google Scholar]
- 23.Marois C, Bougeard S, Gottschalk M, Kobisch M. Multiplex PCR assay for detection of Streptococcus suis species and serotypes 2 and 1/2 in tonsils of live and dead pigs. J Clin Microbiol. 2004;42(7):3169–3175. doi: 10.1128/JCM.42.7.3169-3175.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ferrando ML, de Greeff A, van Rooijen WJ, Stockhofe-Zurwieden N, Nielsen J, Wichgers Schreur PJ, Pannekoek Y, Heuvelink A, van der Ende A, Smith H, et al. Host-pathogen Interaction at the Intestinal Mucosa Correlates With Zoonotic Potential of Streptococcus suis. J Infect Dis. 2015;212(1):95–105. doi: 10.1093/infdis/jiu813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Madsen LW, Bak H, Nielsen B, Jensen HE, Aalbaek B, Riising HJ. Bacterial colonization and invasion in pigs experimentally exposed to Streptococcus suis serotype 2 in aerosol. J Vet Med B Infect Dis Vet Public Health. 2002;49(5):211–215. doi: 10.1046/j.1439-0450.2002.00491.x. [DOI] [PubMed] [Google Scholar]
- 26.Zaccaria E, Cao R, Wells JM, van Baarlen P. A zebrafish larval model to assess virulence of porcine streptococcus suis strains. PLoS One. 2016;11(3):e0151623. doi: 10.1371/journal.pone.0151623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hill L, Veli N, Coote PJ. Evaluation of Galleria mellonella larvae for measuring the efficacy and pharmacokinetics of antibiotic therapies against Pseudomonas aeruginosa infection. Int J Antimicrob Agents. 2014;43(3):254–261. doi: 10.1016/j.ijantimicag.2013.11.001. [DOI] [PubMed] [Google Scholar]
- 28.Desbois AP, Coote PJ. Wax moth larva (Galleria mellonella): an in vivo model for assessing the efficacy of antistaphylococcal agents. J Antimicrob Chemother. 2011;66(8):1785–1790. doi: 10.1093/jac/dkr198. [DOI] [PubMed] [Google Scholar]
- 29.Kavanagh K, Reeves EP. Exploiting the potential of insects for in vivo pathogenicity testing of microbial pathogens. FEMS Microbiol Rev. 2004;28(1):101–112. doi: 10.1016/j.femsre.2003.09.002. [DOI] [PubMed] [Google Scholar]
- 30.Loh JM, Adenwalla N, Wiles S, Proft T. Galleria mellonella larvae as an infection model for group A streptococcus. Virulence. 2013;4(5):419–428. doi: 10.4161/viru.24930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Evans BA, Rozen DE. A Streptococcus pneumoniae infection model in larvae of the wax moth Galleria mellonella. Eur J Clin Microbiol Infect Dis. 2012;31(10):2653–2660. doi: 10.1007/s10096-012-1609-7. [DOI] [PubMed] [Google Scholar]
- 32.Buckley AA, Faustoferri RC, Quivey RG., Jr beta-Phosphoglucomutase contributes to aciduricity in Streptococcus mutans. Microbiology. 2014;160(Pt 4):818–827. doi: 10.1099/mic.0.075754-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Salzet M. Vertebrate innate immunity resembles a mosaic of invertebrate immune responses. Trends Immunol. 2001;22(6):285–288. doi: 10.1016/S1471-4906(01)01895-6. [DOI] [PubMed] [Google Scholar]
- 34.Browne N, Heelan M, Kavanagh K. An analysis of the structural and functional similarities of insect hemocytes and mammalian phagocytes. Virulence. 2013;4(7):597–603. doi: 10.4161/viru.25906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.de Greeff A, Wisselink HJ, de Bree FM, Schultsz C, Baums CG, Thi HN, Stockhofe-Zurwieden N, Smith HE. Genetic diversity of Streptococcus suis isolates as determined by comparative genome hybridization. BMC Microbiol. 2011;11(1):1–15. doi: 10.1186/1471-2180-11-161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Li DD, Deng L, Hu GH, Zhao LX, Hu DD, Jiang YY, Wang Y. Using Galleria mellonella-Candida albicans infection model to evaluate antifungal agents. Biol Pharm Bull. 2013;36(9):1482–1487. doi: 10.1248/bpb.b13-00270. [DOI] [PubMed] [Google Scholar]
- 37.Velikova N, Fulle S, Manso AS, Mechkarska M, Finn P, Conlon JM, Oggioni MR, Wells JM, Marina A. Putative histidine kinase inhibitors with antibacterial effect against multi-drug resistant clinical isolates identified by in vitro and in silico screens. Sci Rep. 2016;6:26085. doi: 10.1038/srep26085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Zaccaria E, van Baarlen P, de Greeff A, Morrison DA, Smith H, Wells JM. Control of competence for DNA transformation in streptococcus suis by genetically transferable pherotypes. PLoS One. 2014;9(6):e99394. doi: 10.1371/journal.pone.0099394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Astrid de Greeff, Herma Buys, Jerry M Wells, Hilde E Smith, Astrid de Greeff, Herma Buys, Jerry M Wells, Hilde E Smith, (2014) A naturally occurring nucleotide polymorphism in the orf2/folc promoter is associated with Streptococcus suis virulence. BMC Microbiology 14 (1). [DOI] [PMC free article] [PubMed]
- 40.Wisselink HJ, Smith HE, Stockhofe-Zurwieden N, Peperkamp K, Vecht U. Distribution of capsular types and production of muramidase-released protein (MRP) and extracellular factor (EF) of Streptococcus suis strains isolated from diseased pigs in seven European countries. Vet Microbiol. 2000;74(3):237–48. [DOI] [PubMed]
- 41.Feder I, Chengappa MM, Fenwick B, Rider M, Staats J. Partial characterization of Streptococcus suis type 2 hemolysin. J Clin Microbiol. 1994;32(5):1256–1260. doi: 10.1128/jcm.32.5.1256-1260.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Allen AG, Bolitho S, Lindsay H, Khan S, Bryant C, Norton P, Ward P, Leigh J, Morgan J, Riches H, et al. Generation and characterization of a defined mutant of streptococcus suis lacking suilysin. Infect Immun. 2001;69(4):2732–2735. doi: 10.1128/IAI.69.4.2732-2735.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Desbois AP, Coote PJ. Utility of greater wax moth larva (galleria mellonella) for evaluating the toxicity and efficacy of new antimicrobial agents. Adv Appl Microbiol. 2012;78:25–53. doi: 10.1016/B978-0-12-394805-2.00002-6. [DOI] [PubMed] [Google Scholar]
- 44.Ferrando ML, Van Baarlen P, Orrù G, Piga R, Bongers RS, Wels M, De Greeff A, Smith HE, Wells JM. Carbohydrate availability regulates virulence gene expression in Streptococcus suis. PloS one. 2014;9(3):e89334. doi: 10.1371/journal.pone.0089334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Stockhofe-Zurwieden N VU, Wisselink HJ, van Leshout H, Smith HE: Comparative studies on the pathogenicity of different Streptococcus suis type 1 strains. In: 14th IPVS: 1996; Bologna. 1996: 299.
- 46.Vecht U, Arends JP, van der Molen EJ, van Leengoed LA. Differences in virulence between two strains of Streptococcus suis type II after experimentally induced infection of newborn germ-free pigs. Am J Vet Res. 1989;50(7):1037–1043. [PubMed] [Google Scholar]
- 47.Vecht U, Wisselink HJ, Stockhofe-Zurwieden N, Smith HE. Characterization of virulence of the Streptococcus suis serotype 2 reference strain Henrichsen S 735 in newborn gnotobiotic pigs. Vet Microbiol. 1996;51(1–2):125–136. doi: 10.1016/0378-1135(96)00028-4. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All data supporting the findings in this manuscript is included here or in the Supplementary material.