

Abstract
The control of organ size presents a fundamental open problem in biology. A declining growth rate is observed in all studied higher animals, and the growth limiting mechanism may therefore be evolutionary conserved. Most studies of organ growth control have been carried out in Drosophila imaginal discs. We have previously shown that the area growth rate in the Drosophila eye primordium declines inversely proportional to the increase in its area, which is consistent with a dilution mechanism for growth control. Here, we show that a dilution mechanism cannot explain growth control in the Drosophila wing disc. We computationally evaluate a range of alternative candidate mechanisms and show that the experimental data can be best explained by a biphasic growth law. However, also logistic growth and an exponentially declining growth rate fit the data very well. The three growth laws correspond to fundamentally different growth mechanisms that we discuss. Since, as we show, a fit to the available experimental growth kinetics is insufficient to define the underlying mechanism of growth control, future experimental studies must focus on the molecular mechanisms to define the mechanism of growth control.
A fascinating aspect of embryonic development is how the growth of organ rudiments is globally coordinated such that all organs, tissues and the paired appendages grow to the correct (relative) size, even when final sizes differ between isogenic offspring because of external factors such as nutrition or temperature1. The mechanism by which growth is terminated has remained elusive.
Growth control has been studied extensively in Drosophila imaginal discs and can be separated into disc-intrinsic and disc-extrinsic mechanisms. Disc-extrinsic mechanisms ensure that body growth and organ growth are coupled in a way that correct size proportions are obtained, even when nutritional status and thus final size differs strongly during development, a phenomenon referred to as allometry2,3. Intriguingly however, similar final imaginal disc sizes are obtained even when imaginal discs are provided with additional developmental time4,5, when the frequency of cell divisions is perturbed6,7, after regeneration of lost tissue8,9, and when imaginal discs are cultured outside the larvae10,11. These results point to the existence of a disc autonomous mechanism of size control that ensures correct final sizes.
The morphogen Decapentaplegic (Dpp) affects growth in all fifteen imaginal discs in the Drosophila larvae, and models have been suggested how Dpp may result in a uniform declining growth rate12,13,14,15,16,17,18 in spite of its graded distribution in imaginal discs19,20,21. In the wing disc, in particular, Dpp forms a dynamically increasing gradient17 from the anterior-posterior boundary across the anterior and posterior sides22,23,24,25. In the eye disc, on the other hand, Dpp is secreted from the morphogenetic furrow (MF) that sweeps from the posterior to the anterior end of the eye disc26,27. Gonzalez-Gaitan and co-workers previously proposed that an exponential decline in the proliferation rate arises in both discs because cells sense the relative local change in the Dpp concentration and divide whenever they experience a fixed relative change of about 40%17,28. However, the area growth rate is unaltered in Mad-/Brk- clones that cannot sense Dpp28,29. Additionally, Dpp affects wing disc growth only in the first half of larval development30 and only in the medial part of the wing disc31. According to an alternative model, uniformly declining growth results from a combination of Dpp signaling and mechanical feedbacks12,15,32,33. Finally, we have recently shown that the growth rate of the apical area in the Drosophila eye imaginal disc declines inversely proportional to the total apical area, which is consistent with a dilution-based mechanism for growth control34.
Here, we use published quantitative growth data to carry out an unbiased evaluation of alternative candidate growth mechanisms. Based on the quantitative analysis, we rule out growth control by dilution in the wing disc. Evaluating alternative candidate growth mechanisms, we find that biphasic exponential growth best fits the wing disc data. An exponential decline of the growth rate with developmental time and logistic growth are also consistent with the data. These three growth laws correspond to fundamentally different growth mechanisms that we discuss. Given the wide range of growth laws that are consistent with the data, we emphasize the need to explore a wider range of growth limiting mechanisms and to carefully check the consistency of any proposed mechanism with additional experimental data – the ability to recapitulate the growth kinetics alone provides insufficient support for any mechanism.
Results and Discussion
Dilution of a cytokine cannot explain the Wing Disc Growth Kinetics
The growth of the apical area, A, of Drosophila imaginal discs over developmental time, t, can be described mathematically by
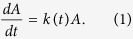 |
Here, k refers to the area growth rate. We have previously shown that the declining area growth rate in the eye disc can be described by an area-dependent decline34 of the form
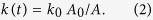 |
Here A0 is the initial area and k0 the maximal growth rate. We sought to test whether this mechanism would also apply to the wing disc. To this end, we used two independent, published datasets for wing disc area growth17,35 (Fig. 1A) to determine the area growth rate, k (Fig. 1B), and to fit the model (Equations 1 and 2) to these datasets (Fig. 1C,D). The poor fit to the data rejects this mechanism for growth control in the wing disc (Fig. 1C,D, Figs S1A–D and S2A–D).
Figure 1. A dilution mechanism cannot explain growth control in the Drosophila wing disc.

(A) Drosophila wing disc area growth as reported in ref. 17 (black) and ref. 35 (red). Two spline fits (lines, see Materials and Methods for details) were fitted to each data set to estimate the slope at each data point. (B) The area growth rate k as determined from Eq. (8) declines over developmental time. Colour code corresponds to the data and fits in panel A. (C,D) A model based on the dilution mechanism (Equation 2) fits the two independent wing disc growth datasets by Wartlick et al.17 (C), and by Nienhaus et al.35 (D) badly; fits were obtained using the residuals given by Eq. (9) (solid lines) or Eq. (10) (dashed lines). Parameters are given in Table S1.
Unbiased Analysis of Candidate Growth Laws for Wing Disc Growth
We next tested which other candidate growth laws would allow us to reproduce the wing disc data (Fig. 2). To this end, we simulated the growth model in Eq. (1) with the growth laws that we had previously considered as candidates for growth control in the Drosophila eye disc34. In addition, we considered biphasic growth, which had previously been shown to describe wing disc growth2, and logistic growth, which had previously been shown to describe the regeneration kinetics of newt limbs14. In summary, in the simplest model we assumed a constant growth rate (CST, Fig. 2A,B)
Figure 2. Evaluation of Candidate Growth Laws for the Drosophila Wing Disc.

(A–J) Fits of the growth model in Eq. (1) with the different growth laws given by Eqs (3, 4, 5, 6) reveals best fit to the datasets obtained by Wartlick et al.17 (columns 1 and 3) and Nienhaus et al.35 (columns 2 and 4) with biphasic growth (C,D - green), logistic growth (I,J - red) and an exponentially declining growth law (E,F - yellow). The worst fit, but still better than the one based on the dilution mechanism (Fig. 1), is obtained with a constant growth law (A,B - black). A powerlaw decline (G,H - blue) provides an intermediate fit to the data. Fits were obtained using the residuals given by Eq. (9) (solid lines) or Eq. (10) (dashed lines). (K) Relative deviation of the resulting fits from the data (circles - Wartlick et al.17; squares - Nienhaus et al.35). The deviation was normalized with respect to the minimal value for each dataset and the residual definition (Equation 9 (closed symbols) & 10 (open symbols)). (L) Cell density in the wing disc over time as measured in Wartlick et al.17 (grey) and as inferred from the cell number and area data in Wartlick et al.17 (black). Vertical lines indicate the switch points that minimize the deviation between data and BPH model for the two different definitions of the residuals (Fig. S3A,B; solid line - Equation 9; dashed line - Eq. 10).
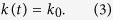 |
In biphasic growth (BPH), the value of k0 changes once, thus giving rise to two phases of different exponential growth (biphasic, BPH, Fig. 2C,D). The switch point from phase I to II was defined as the one providing the lowest deviation from the data (Fig. S3). In addition, we considered models with a continuously declining growth rate, either as exponential decaying growth rate (EXP, Fig. 2E,F) of the form
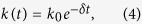 |
or by following a power law decay (POW, Fig. 2G,H) of the form
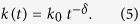 |
Finally, the growth rate could depend on the total wing disc area. Besides dilution (Equation 2, Fig. 1C,D), logistic growth (LOG, Fig. 2I,J) can result in an area-dependent growth rate
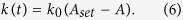 |
We note that the models with a constant or an area-dependent growth rate have two parameters, the biphasic exponential model (BHP) has four parameters, while all other models (EXP, POW, LOG) have three independent parameters. A full list for the parameter values can be found in the supplementary information (Tables S1 & S2).
To fit the models to the data, we defined the residuals in two independent ways: either by normalizing with respect to the standard error (dashed lines in Figs 1C,D & 2A–J & Fig. S1) or by taking the logarithm of the data (solid lines in Figs 1C,D & 2A–J & Fig. S2) to correct for the different orders of magnitudes in the data and thus be able to fit the data well (Fig. 2A–J) (see Material & Methods for details). To compare the model fits, the sum of squared residuals (RSS, Fig. 2K, Table S3) and coefficient of determination (R2, Table S4) were used. We note that fitting the logarithmic data provided the smaller deviation and better coefficient of determination (R2) in all cases (Tables S3 and S4). Using these measures, the biphasic model (BHP) performs best for almost all weightings and data sets (Fig. 2K). Interestingly, also the reported change in cell density17 correlates with the predicted switch point. Thus, the cell density increases until the inferred switch point (vertical lines) and subsequently stabilizes (Fig. 2L). Logarithmic growth (red) and to a slightly lesser extent an exponentially declining growth rate (yellow) also fit the data very well (Fig. 2K), and the difference to the biphasic model is only minor (Tables S3 & S4). The power law model fits the data slightly, but consistently, worse as judged by the R2 value (Table S4) and the RSS (Table S3).
The models with a constant (CST) or an area-dependent growth rate both have only two free parameters, and thus at least one parameter less than the models discussed above (BHP, EXP, POW, LOG). The model with the area-dependent growth rate performs considerably worse than the model with a constant (CST) growth rate, and can thus be rejected (Fig. 2K, Tables S3 & S4). To compare the model with a constant (CST) growth rate to the other models, the different number of free parameters needs to be taken into account. Unlike other commonly used model selection criterions, such as the Bayesian information criterion (BIC) or the Akaike information criterion (AIC), the F-test provides a p-value on the null hypothesis that the better fit can be solely explained by the increased number of parameters. Its use is, however, restricted to the case where the simpler model is nested within the more complex model, limiting it to the comparison of the biphasic (BHP) and single exponential model (CST). Conducting an F-test, we find that for both data sets and weightings, the biphasic exponential model fits the data significantly better than the model with a constant growth rate (Table S5).
Candidate Mechanisms for Growth Control in the Drosophila Wing Disc
While the biphasic growth law, logistic growth, and an exponentially declining growth rate all fit the data well, they point to very different underlying mechanisms. A biphasic growth law would require a sudden change in cell growth, potentially because of cell differentiation. It has been argued that larvae can monitor their size and trigger the switch to lower growth rates when reaching a ‘critical size’2. We note that in contrast to models with continuously declining growth rates, the biphasic growth law would still require an additional mechanism to ultimately stop growth. Otherwise, growth would continue with the speed from the second phase. Growth in the models with a continuously declining growth rate will eventually be so close to zero that the expansion becomes negligible.
An exponential decline in the growth rate could be achieved by the Dpp-dependent mechanism proposed by Gonzalez-Gaitan and co-workers17,28, or if a growth factor to which the system responded linearly, was degraded at a constant rate δ, i.e.
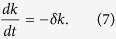 |
However, in both cases there are fundamental problems such as the reliable read-out at low concentrations and the robustness of the mechanism to changes in the total developmental time as observed in grafting experiments10,11. Thus, as the exponentially declining growth rate, k (Equation 4), declines 3-fold and 5-fold in the two different datasets from early to late stages17,35, cells would need to be able to linearly respond to 3–5-fold changes in this growth-controlling factor. Recent experiments indeed question a central role of the Dpp gradient in determining final disc size29,30,31.
Logistic growth requires a mechanism to set a final size and to reduce the growth rate as this final size is approached. The intercalation model results in logistic growth14. Here, the growth rate is postulated to be proportional to the positional difference between neighbouring cells, which is reduced as the structure grows out. How such positional differences would be measured by cells is not known. While loss of the cell-adhesion mediating cadherin Fat has been shown to enhance growth in the medial part of the wing disc, it has remained unclear whether the Fat-dependent growth limitation is central to growth termination33,36,37,38. Beyond its molecular implementation, an intercalation mechanism has a number of conceptual limitations. Thus, while the intercalation mechanism can explain restoration of missing positional values during regeneration and a logistic growth law would be robust to changes in developmental speed, it remains unclear how the positional identity would scale when tissues grow to different final sizes, for instance because of differences in nutrients available2. Finally, while it has been argued that larvae can sense a ‘critical size’2, it remains unclear how wing discs would achieve this on the molecular level in a disc-autonomous way and how disc size could then vary in response to changes in external conditions (nutrients, temperature etc.).
In conclusion, we can rule out an area-dependent dilution mechanism for growth control in the wing disc. Rather the responsible mechanism must give rise to biphasic growth, logistic growth, or an exponentially declining growth rate. Given that several growth laws match the measured growth curves, it needs to be stressed that the reproduction of the growth kinetics alone is insufficient evidence for any proposed mechanism and mechanistic proof needs to be provided. At the same time, there could be other growth laws that also reproduce the data and that we have not yet identified. It will be interesting whether the same mechanism limits growth in all organs and species or whether distinct mechanisms have evolved17,28,34,39,40.
Materials and Methods
Software
All simulations and analysis were done using Matlab R2016A (The MathWorks, Natick, MA, USA) or the free software environment R in version 3.2.4.41.
Estimation of area growth rate
The area growth rate
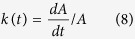 |
in Eq. (1) can be determined by estimating the slope 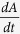 at all time points and diving this slope by the measured area A at each timepoint. To estimate the slope
at all time points and diving this slope by the measured area A at each timepoint. To estimate the slope 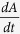 from the data without making any assumption about the (true) function underlying the growth dynamics, we used cubic smoothing splines as provided by the smooth.spline function in R41. As the exact shape of the decline of the area growth rate k depends on the spline fits, we used two different fits for each data set with a varying smoothing parameter. The resulting spline fits are shown in Fig. 1A, and the estimated area growth rate k(t) in Fig. 1B.
from the data without making any assumption about the (true) function underlying the growth dynamics, we used cubic smoothing splines as provided by the smooth.spline function in R41. As the exact shape of the decline of the area growth rate k depends on the spline fits, we used two different fits for each data set with a varying smoothing parameter. The resulting spline fits are shown in Fig. 1A, and the estimated area growth rate k(t) in Fig. 1B.
Model Fitting
The two available independent studies do not provide the variance for all data points17,35. We therefore quantified the deviation of the model from the data in form of the residual R at time point k as
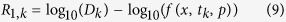 |
or by weighting the difference by the given standard error (SE) as
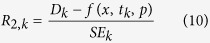 |
where Dk is the mean, SEk its standard error, and f(x,tk,p) refers to the model value at time point k. We took the logarithm of the data for the first measure since the data is spread over several orders of magnitude and larger values would therefore be weighted much higher otherwise. In the data set from Wartlick et al. no SE was given for the time point at approximately 80 hours and for the last time point (SE = 0)17. We therefore excluded these two time points from the analysis with Eq. 10.
The model parameters were then determined by solving the minimization
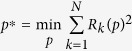 |
where N is the number of distinct time points. This was done independently for both definitions of the residuals using the trust-region reflective algorithm as implemented in lsqnonlin (Matlab R2016A, MathWorks, Natick, MA, USA), resulting in two parameter sets for each model.
Comparison of model fits and statistical analysis
Models were compared based on the residual sum of squares (RSS) and their R2 value. The R2 value of model j was calculated as
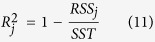 |
where RSSj is the residual sum of squares of model j calculated as explained above (Equations 9 or 10) and SST is the total sum of squares.
To evaluate whether there is a statistical significance in the difference how the models fit the data, the F-test was used. The F-test can only be used to compare a simple model which is nested within a more complex one. Thus, here it can only be used to compare the biphasic exponential model to the single exponential model, as the latter one is nested within the first one. The F-statistic was calculated as
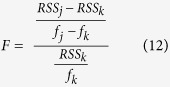 |
where model j is nested within model k, fj and fk are the degrees of freedom (number of data points minus number of parameters) for model j and k, respectively, and RSS their residual sum of squares. The respective p-value can then be calculated from the F cumulative distribution function. The degrees of freedom to be used in the cumulative distribution function are (fj − fk, fk).
Additional Information
How to cite this article: Vollmer, J. and Iber, D. An Unbiased Analysis of Candidate Mechanisms for the Regulation of Drosophila Wing Disc Growth. Sci. Rep. 6, 39228; doi: 10.1038/srep39228 (2016).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Material
Acknowledgments
The authors are grateful to Erkan Uenal, Patrick Fried, and Fernando Casares for discussions. This research was supported by a Swiss Institute of Bioinformatics (SIB) Fellowship to J.V.
Footnotes
Author Contributions D.I. conceived the study. J.V. performed the mathematical analysis of the different growth models. D.I., and J.V., wrote the manuscript.
References
- Azevedo R. B. R., French V. & Partridge L. Temperature modulates epidermal cell size in Drosophila melanogaster. J. Insect Physiol. 48, 231–237 (2002). [DOI] [PubMed] [Google Scholar]
- Shingleton A. W., Mirth C. K. & Bates P. W. Developmental model of static allometry in holometabolous insects. Proceedings. Biol. Sci. 275, 1875–85 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shingleton A. W., Frankino W. A., Flatt T., Nijhout H. F. & Emlen D. J. Size and shape: the developmental regulation of static allometry in insects. Bioessays 29, 536–48 (2007). [DOI] [PubMed] [Google Scholar]
- Colombani J., Andersen D. S. S. & Léopold P. Secreted Peptide Dilp8 Coordinates Drosophila Tissue Growth with Developmental Timing. Science 336, 582–585 (2012). [DOI] [PubMed] [Google Scholar]
- Garelli A., Gontijo A. M. M., Miguela V., Caparros E. & Dominguez M. Imaginal Discs Secrete Insulin-Like Peptide 8 to Mediate Plasticity of Growth and Maturation. Science 336, 579–82 (2012). [DOI] [PubMed] [Google Scholar]
- Neufeld T. P., de la Cruz A. F., Johnston L. A. & Edgar B. A. Coordination of growth and cell division in the Drosophila wing. Cell 93, 1183–1193 (1998). [DOI] [PubMed] [Google Scholar]
- Weigmann K., Cohen S. M. & Lehner C. F. Cell cycle progression, growth and patterning in imaginal discs despite inhibition of cell division after inactivation of Drosophila Cdc2 kinase. Development 124, 3555–3563 (1997). [DOI] [PubMed] [Google Scholar]
- Díaz-García S. & Baonza A. Pattern reorganization occurs independently of cell division during Drosophila wing disc regeneration in situ. Proc. Natl. Acad. Sci. USA 110, 13032–7 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun G. & Irvine K. D. Regulation of Hippo signaling by Jun kinase signaling during compensatory cell proliferation and regeneration, and in neoplastic tumors. Dev. Biol. 350, 139–151 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garcia-Bellido A. Larvalentwicklung transplantierter Organe von Drosophila melanogaster im Adultmilieu. J. Insect Physiol. 11, 1071–1078 (1965). [Google Scholar]
- Bryant P. J. & Levinson P. Intrinsic growth control in the imaginal primordia of Drosophila, and the autonomous action of a lethal mutation causing overgrowth. Dev. Biol. 107, 355–363 (1985). [DOI] [PubMed] [Google Scholar]
- Aegerter-Wilmsen T., Aegerter C. M., Hafen E. & Basler K. Model for the regulation of size in the wing imaginal disc of Drosophila. Mech. Dev. 124, 318–26 (2007). [DOI] [PubMed] [Google Scholar]
- Bittig T., Wartlick O., González-Gaitán M. & Jülicher F. Quantification of growth asymmetries in developing epithelia. Eur. Phys. J. E. Soft Matter 30, 93–9 (2009). [DOI] [PubMed] [Google Scholar]
- Bryant P. J. J. & Simpson P. Intrinsic and extrinsic control of growth in developing organs. Q. Rev. Biol. 59, 387–415 (1984). [DOI] [PubMed] [Google Scholar]
- Hufnagel L., Teleman A. A., Rouault H., Cohen S. M. & Shraiman B. I. On the mechanism of wing size determination in fly development. Proc. Natl. Acad. Sci. USA 104, 3835–40 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Milán M., Campuzano S. & García-Bellido A. Cell cycling and patterned cell proliferation in the wing primordium of Drosophila. Proc. Natl. Acad. Sci. USA 93, 640–5 (1996). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wartlick O. et al. Dynamics of Dpp signaling and proliferation control. Science 331, 1154–1159 (2011). [DOI] [PubMed] [Google Scholar]
- Day S. J. & Lawrence P. A. Measuring dimensions: the regulation of size and shape. Development 127, 2977–87 (2000). [DOI] [PubMed] [Google Scholar]
- Entchev E. V., Schwabedissen A. & González-Gaitán M. Gradient formation of the TGF-beta homolog Dpp. Cell 103, 981–991 (2000). [DOI] [PubMed] [Google Scholar]
- Teleman A. A. & Cohen S. M. Dpp gradient formation in the Drosophila wing imaginal disc. Cell 103, 971–980 (2000). [DOI] [PubMed] [Google Scholar]
- Wartlick O., Kicheva A. & González-Gaitán M. Morphogen gradient formation. Cold Spring Harb. Perspect. Biol. 1, a001255 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raftery L. a, Sanicola M., Blackman R. K. & Gelbart W. M. The relationship of decapentaplegic and engrailed expression in Drosophila imaginal disks: do these genes mark the anterior-posterior compartment boundary? Development 113, 27–33 (1991). [DOI] [PubMed] [Google Scholar]
- Tabata T. & Kornberg T. B. Hedgehog is a signaling protein with a key role in patterning Drosophila imaginal discs. Cell 76, 89–102 (1994). [DOI] [PubMed] [Google Scholar]
- Basler K. & Struhl G. Compartment boundaries and the control of Drosophila limb pattern by hedgehog protein. Nature 368, 208–14 (1994). [DOI] [PubMed] [Google Scholar]
- Posakony L. G., Raftery L. A. & Gelbart W. M. Wing formation in Drosophila melanogaster requires decapentaplegic gene function along the anterior-posterior compartment boundary. Mech. Dev. 33, 69–82 (1990). [DOI] [PubMed] [Google Scholar]
- Heberlein U., Wolff T. & Rubin G. M. The TGF beta homolog dpp and the segment polarity gene hedgehog are required for propagation of a morphogenetic wave in the Drosophila retina. Cell 75, 913–926 (1993). [DOI] [PubMed] [Google Scholar]
- Firth L. C., Bhattacharya A. & Baker N. E. Cell cycle arrest by a gradient of Dpp signaling during Drosophila eye development. BMC Dev. Biol. 10, 28 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wartlick O., Jülicher F. & Gonzalez-Gaitan M. Growth control by a moving morphogen gradient during Drosophila eye development. Development 141, 1884–93 (2014). [DOI] [PubMed] [Google Scholar]
- Schwank G., Yang S.-F., Restrepo S. & Basler K. Comment on ‘Dynamics of Dpp Signaling and Proliferation Control’. Science 335, 401–401 (2012). [DOI] [PubMed] [Google Scholar]
- Akiyama T. & Gibson M. C. Decapentaplegic and growth control in the developing Drosophila wing. Nature 527, 375–8 (2015). [DOI] [PubMed] [Google Scholar]
- Harmansa S., Hamaratoglu F., Affolter M. & Caussinus E. Dpp spreading is required for medial but not for lateral wing disc growth. Nature 527, 317–22 (2015). [DOI] [PubMed] [Google Scholar]
- Shraiman B. I. Mechanical feedback as a possible regulator of tissue growth. Proc. Natl. Acad. Sci. USA 102, 3318–3323 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aegerter-Wilmsen T. et al. Integrating force-sensing and signaling pathways in a model for the regulation of wing imaginal disc size. Development 125, 3221–31 (2012). [DOI] [PubMed] [Google Scholar]
- Vollmer J. et al. A quantitative analysis of growth control in the Drosophila eye disc. Development 143, 1482–90 (2016). [DOI] [PubMed] [Google Scholar]
- Nienhaus U., Aegerter-Wilmsen T. & Aegerter C. M. In-vivo imaging of the Drosophila wing imaginal disc over time: novel insights on growth and boundary formation. PLoS One 7, e47594 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rogulja D. & Irvine K. D. Regulation of cell proliferation by a morphogen gradient. Cell 123, 449–61 (2005). [DOI] [PubMed] [Google Scholar]
- Rogulja D., Rauskolb C. & Irvine K. D. Morphogen Control of Wing Growth through the Fat Signaling Pathway. Dev. Cell 15, 309–321 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwank G. et al. Antagonistic growth regulation by Dpp and Fat drives uniform cell proliferation. Dev. Cell 20, 123–30 (2011). [DOI] [PubMed] [Google Scholar]
- Ricklefs R. E. Embryo growth rates in birds and mammals. Funct. Ecol. 24, 588–596 (2010). [Google Scholar]
- Grunert L. W., Clarke J. W., Ahuja C., Eswaran H. & Nijhout H. F. A Quantitative Analysis of Growth and Size Regulation in Manduca sexta: The Physiological Basis of Variation in Size and Age at Metamorphosis. PLoS One 10, e0127988 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Core Team R. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL http://www.R-project.org/ (2013).
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.