

Abstract
Recent articles have highlighted numerous additional functions of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) that are independent of its well-documented glycolytic function. One of the most intriguing of these functions is as an initiator of programmed cell death cascades. This activity involves a nuclear appearance of GAPDH, a considerable proportion of which requires synthesis of new GAPDH protein and has characteristics suggesting the involvement of a novel isozyme. The relevance of such findings to human neurodegenerative conditions is emphasized by the increased nuclear GAPDH observed in postmortem samples from patients with Parkinson's disease, Alzheimer's disease, Huntington's disease and glaucoma, among others. A number of small-molecule compounds have now been identified that show anti-apoptotic activity because of their ability to interact with GAPDH and prevent its nuclear accumulation. These compounds, one of which is currently being tested in late-stage Phase II clinical trials as a disease-modifying therapy for Parkinson's disease, have potential utility in the treatment of human neurodegenerative conditions.
Medical subject headings: apoptosis, drug therapy, glyceraldehyde-3-phosphate dehydrogenase, neurodegenerative diseases
Abstract
Des articles récents ont décrit de nombreuses fonctions supplémentaires de la glycéraldéhyde-3-phosphate déshydrogénase (GAPDH) indépendantes de sa fonction glycolytique bien documentée. Une des plus intrigantes de ces fonctions est celle de catalyseur de cascades d'apoptose. Cette activité met en cause une apparition dans le noyau de la GAPDH, dont une proportion importante nécessite la synthèse de nouvelle protéine de GAPDH et a des caractéristiques qui indiquent la présence d'un isozyme nouveau. L'élévation de la GAPDH nucléaire observée dans des échantillons postmortem prélevés chez des patients atteints de la maladie de Parkinson, la maladie d'Alzheimer, la chorée d'Huntington et le glaucome, notamment, met en évidence la pertinence de ces constatations dans le cas de problèmes neurodégénératifs humains. On a maintenant identifié un certain nombre de composés à petites molécules qui ont une activité anti-apoptotique parce qu'ils peuvent réagir avec la GAPDH et l'empêcher de s'accumuler dans le noyau. Ces composés, dont l'un fait actuellement l'objet d'essais cliniques de phase II, qui achèvent, comme traitement de fond contre la maladie de Parkinson, offrent des possibilités pour le traitement des maladies neurodégénératives humaines.
Introduction
Glyceraldehyde-3-phosphate dehydrogenase (GAPDH [EC 1.2.1.12]) is widely regarded as a ubiquitously occurring, nonregulated enzyme that functions exclusively within the glycolytic pathway. Despite an increasing number of publications showing that GAPDH mRNA levels vary in response to a variety of stimuli, GAPDH is still widely used as a “housekeeping” gene for standardizing Northern blots. Several recent reviews, however, have highlighted the multifunctional nature of GAPDH.1,2,3 One of the most intriguing of the nonglycolytic functions of GAPDH is as an initiation signal for cellular apoptosis or programmed cell death (PCD), which may be significant for a number of human disorders. The role of GAPDH in some PCD cascades has recently been reviewed,4 and a detailed discussion of this is beyond the scope of the current article. Rather, this commentary will focus on recent advances in determining the mechanism by which GAPDH initiates PCD and the clinical relevance of GAPDH-mediated PCD, with particular emphasis on the potential of GAPDH as a target for novel, small-molecule, anti-apoptotic, disease-modifying therapies.
Relevance of GAPDH to human neurodegenerative disorders
Although still questioned by some, accumulating evidence suggests that PCD pathways play at least some role in both chronic and acute human neurodegenerative disorders.5,6,7,8,9,10 As previously discussed,4,11 there is now good evidence showing that GAPDH can play a role in the initiation of some PCD cascades in a variety of cell culture models. Whether GAPDH is also involved in the initiation of PCD cascades in humans, and more specifically during neurodegeneration, has rarely been examined. Recent studies suggest, however, that GAPDH does indeed play a role.
Polyglutamine-repeat disorders
By far the best documented evidence for this role comes from studies of the polyglutamine-repeat disorders. These diverse diseases occur because of the presence of long polyglutamine repeats in the disease proteins. In each instance, the disease is known to be the result of the presence of the expanded polyglutamine tract. Shortly after the causal relation between these polyglutamine tracts and disease severity and progression was demonstrated, it was reported that the polyglutamine regions selectively bound to a number of proteins. One of these proteins is GAPDH.12 The resulting protein aggregates are found within the nucleus, and it has been suggested that the presence of these proteins within the nucleus is sufficient for cell death cascades to be initiated.13,14 This finding is remarkably similar to those of previous cell culture studies where the appearance of GAPDH in the nucleus appears to trigger cell death cascades. Recently, nuclear GAPDH has been shown to be present in vulnerable neurons in postmortem samples from patients with Machado–Joseph disease15,16 and dentatorubral-pallidoluysian atrophy (DRLPA),16 which are both polyglutamine expansion diseases, and not in control samples. Intriguingly, the presence of nuclear GAPDH was demonstrated using an antibody raised against the nuclear GAPDH isolated from cells undergoing apoptosis, which appears to show selectivity toward nuclear GAPDH. Further, white-matter lesions associated with variant forms of DRLPA have recently been reported to be associated with increased GAPDH immunoreactivity.17 A high molecular weight GAPDH immunoreactive protein has also been observed in nuclear fractions from fibroblasts of patients with Huntington's disease. Similar GAPDH species were not observed in fibroblasts from age-matched controls.18
Parkinson's disease
Nuclear GAPDH immunostaining has now been shown in melanized substantia nigra neurons from postmortem samples from patients with Parkinson's disease.19 Again, these are the neurons primarily affected in Parkinson's disease. Similar observations have also been made using the nuclear-selective GAPDH antibody.15 Further, recent reports suggest that a fraction of less than 10 kd isolated from the cerebrospinal fluid (CSF) of patients with Parkinson's disease causes apoptosis when added to cells in culture. Equivalent fractions from the CSF samples of individuals who did not have Parkinson's disease showed no such toxicity. Remarkably, the toxicity of the CSF from individuals with Parkinson's disease appears to be prevented by treatment of cells with either a GAPDH antisense oligonucleotide,20 suggesting that synthesis of new GAPDH protein is required in order to initiate the cell death cascade, or with the anti-apoptotic GAPDH ligand, R-deprenyl.21
Alzheimer's disease
Several years ago, it was shown that antibodies raised against Alzheimer's disease plaques reacted with nuclear GAPDH in cells undergoing apoptosis in culture.22 More recently, the antibody raised against nuclear GAPDH from dying cells was shown to react with amyloid plaques from postmortem samples obtained from patients with Alzheimer's disease.15,16 In this regard, it is of interest to note that GAPDH was previously shown to bind to the C-terminal region of the amyloid precursor protein (APP).23 C-terminal fragments of APP have received considerable attention recently as a possible neurotoxin in Alzheimer's disease. The presence of a high molecular weight GAPDH immunoreactive species has also been reported in Alzheimer's disease, but not in fibroblasts from healthy controls.24
Glaucoma
Postmortem retinal samples from patients with glaucoma have also been reported to show an accumulation of nuclear GAPDH in the susceptible retinal ganglion cells.25 Glaucoma may be regarded as a form of retinal ischemia and, in this regard, it is interesting to note that in an animal model of focal ischemia, the appearance of nuclear GAPDH has been documented to occur in both the ischemic core and the penumbral regions.26
Relevance of GAPDH
From the preceding, it is clear that there is increasing evidence that nuclear accumulation of GAPDH initiating cell death mechanisms in cell culture systems has relevance to a number of human neurodegenerative disorders.
GAPDH mechanisms: protein translocation or new protein synthesis?
The mechanism(s) by which the nuclear appearance of GAPDH initiates cell death remains unknown. GAPDH has been shown to have a number of nonglycolytic roles in the nucleus that could be of importance, including activation of transcription and nuclear DNA repair.1,2,3,4 This latter function is seen as uracil DNA glycosylase activity.27 Previous studies, however, have demonstrated that changes in GAPDH-associated uracil DNA glycosylase activity are not involved in the initiation of cell death.28 At present, activation of transcription by nuclear GAPDH is perhaps the most likely explanation for the induction of PCD cascades.
Examination of the literature regarding GAPDH and apoptosis reveals frequent references to translocation of GAPDH to the nucleus being required for the initiation of cell death.29,30,31 There are, however, few direct demonstrations of pre-existing GAPDH protein relocation to the nucleus directly initiating PCD. Rather, most studies show the appearance of GAPDH in the nucleus. Indeed, there appears to be very good evidence that rather than a translocation of existing GAPDH per se, PCD is initiated following the synthesis of new GAPDH protein.
Shashidharan et al,32 using GAPDH tagged with green fluorescent protein (GFP), have shown that there is a time-dependent appearance of fluorescent signal in the nucleus, following exposure of transfected cells to H2O2. Although this is consistent with a translocation of pre-existing GAPDH, as previously discussed,11 such a translocation was detected in only a small proportion of cells (about 10%), and H2O2 treatment routinely causes a much higher level of apoptosis. Ishitani et al16 have also recently reported a time-dependent appearance of GFP-tagged GAPDH in about 13% of cells during the 48 hours after transfection. These cells did not appear to have been stimulated to undergo apoptosis, other than that resulting from an overexpression of GAPDH, which the authors reported was sufficient to induce cell death. Whether the nuclear-accumulated GAPDH correlated with apoptosis was not reported. A previous study did report that translocation to the nucleus of a fraction of overexpressed GFP-tagged GAPDH resulted in an increase in the basal level of apoptosis.30
In contrast to the translocation evidence, there is much evidence that synthesis of new GAPDH protein is required for initiation of PCD cascades. By far the most convincing evidence is the demonstration that pretreatment with GAPDH antisense oligonucleotides prevents not only the increases seen in GAPDH mRNA33,34,35 and nuclear GAPDH protein,28,33,34 but also cell death.29,33,34,36,37 This effect was similar to that observed with both the protein synthesis inhibitor cycloheximide and the inhibitor of transcription actinomycin D.38 The effect of GAPDH antisense oligonucleotides is dose dependent, with an observed maximum protective effect in excess of 80%.11 Although there is still controversy as to the precise mechanism by which antisense treatments work, there is no suggestion in the literature that antisense treatment can prevent the translocation of existing protein from one subcellular compartment to another. Thus, whereas there may be a small amount of GAPDH that initially translocates from the cytoplasm to the nucleus, the vast majority of nuclear GAPDH appears to be derived from new protein synthesis. Such a conclusion is in agreement with the apparent almost-complete prevention of the appearance of nuclear GAPDH reported following either antisense treatment28,33,34 or administration of protein synthesis inhibitors,38 and with the recent description of increased Golgi apparatus GAPDH immunoreactivity before the appearance of GAPDH in the nucleus, observed in cells that subsequently undergo apoptosis.31
At least a proportion of the induced nuclear GAPDH appears to be a novel isozyme. Six nuclear GAPDH species have been identified by Saunders et al28 using 2-dimensional gel electrophoresis. Induction of cell death is associated with the appearance of distinct acidic isoforms of GAPDH, an effect that is prevented by antisense pretreatment.28 This prevention of acidic GAPDH appearance by antisense pretreatment suggests that these isoforms represent newly synthesized, distinct protein, rather than post-translationally modified existing protein. Our own preliminary studies appear to confirm this appearance of acidic GAPDH isoforms (Fig. 1; Dr. Paula Ashe, ALviva Biopharmaceuticals Inc., Saskatoon, Sask.: personal communication, 2003).
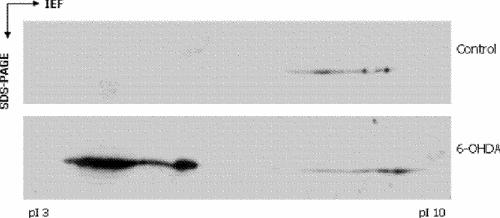
Fig. 1: Appearance of acidic isoforms of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) after treatment with 6-hydroxydopamine (6-OHDA). Human neuroblastoma (SH-SY5Y) cells were induced to undergo programmed cell death (PCD) by treatment with 6-OHDA. Cells were subcellular fractionated and nuclear extracts subjected to 2-dimensional gel electrophoresis. Control cultures were treated identically with the exception of vehicle treatment in place of 6-OHDA. The cultures treated with 6-OHDA show a pronounced appearance of acidic GAPDH bands in the nuclear fractions. Similar bands are absent from the control culture nuclear fractions.
We have extended these initial observations to show that the appearance of GAPDH in the nucleus following 6-hydroxydopamine treatment is associated with different nuclear compartments. In excess of 50% of this new GAPDH is resistant to standard extraction procedures, in comparison with less than 5% of nuclear GAPDH under control conditions (Fig. 2; Dr. Paula Ashe: personal communication, 2003). We are currently in the process of replicating these findings; they are however, similar to those of Sawa et al,29 who reported that induced nuclear GAPDH was not extractable.
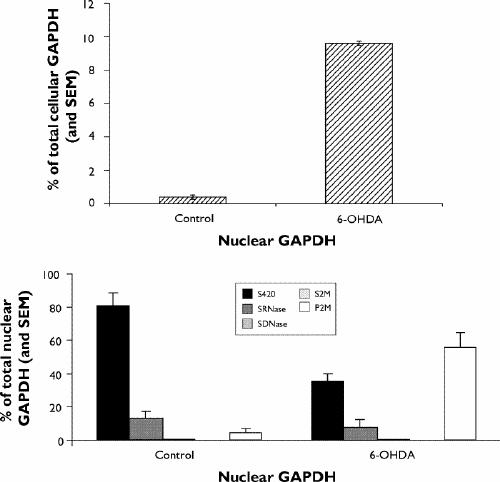
Fig. 2: Changes in nuclear GAPDH after treatment with 6-OHDA. SH-SY5Y cells were induced to undergo PCD by the addition of 6-OHDA. Cells were subcellular fractionated and GAPDH in the nuclear fractions examined. Upper panel: 6-OHDA treatment was associated with a pronounced increase in nuclear GAPDH levels. Lower panel: under control conditions, most (> 80%) of the low levels of nuclear GAPDH present were readily extracted as a soluble fraction after centrifugation. Less than 5% of nuclear GAPDH was inextractable by RNase/DNase/2M salt treatment under control conditions. After treatment with 6-OHDA, only 35% of the increased nuclear GAPDH was extractable as a soluble fraction by centrifugation. In excess of 55% of nuclear GAPDH was resistant to extraction by RNase/DNase/2M salt after 6-OHDA treatment. SEM = standard error of the mean, S420 = the soluble fraction from a 420-mmol/L salt extraction, SRNase = the soluble fraction following RNase treatment, SDNase = the soluble fraction following DNase treatment, S2M = the soluble fraction following 2-mol/L salt extraction, P2M = the insoluble fraction following 2-mol/L salt extraction.
Finally, Ishitani et al38 have reported the preparation of an antibody to nuclear GAPDH from cells undergoing PCD. This antibody appears to react selectively with the induced nuclear GAPDH, having little cross-reactivity to cytosolic GAPDH.16 Paradoxically, these authors have been unable to detect changes in the primary sequence of GAPDH during PCD. As such, the basis of the antigenic selectivity remains to be elucidated. It is worth noting that although there is a large increase in nuclear GAPDH, this still represents only a small proportion (< 10%) of total cellular GAPDH (Fig. 2). Thus, attempts to detect novel GAPDH sequences should be directed toward GAPDH isolated from distinct subcellular compartments, under both control and PCD conditions. Which, if any, of the apparent novel isoforms described here this nuclear GAPDH antibody recognizes is not known.
With respect to the possible involvement of GAPDH isoforms, it is worth noting that, although they receive little attention, isoforms of GAPDH have previously been described.39,40,41 Indeed, multiple copies of the GAPDH gene are present, with in excess of 20 believed to be present in the human genome.42 Although these multiple copies have largely been discounted as being pseudogenes, in light of recent data this should be revisited, bearing in mind that a lack of glycolytic activity need not preclude a functional, nonglycolytic role for any expressed GAPDH protein.
From the evidence presented here, our research group has developed a working model to explain the role of GAPDH in the initiation of PCD (Fig. 3). A central tenet of this model is that the synthesis of new GAPDH protein is an absolute requirement. The initiation events for this new GAPDH protein may vary and include, although are not limited to, 1 or more of the following:
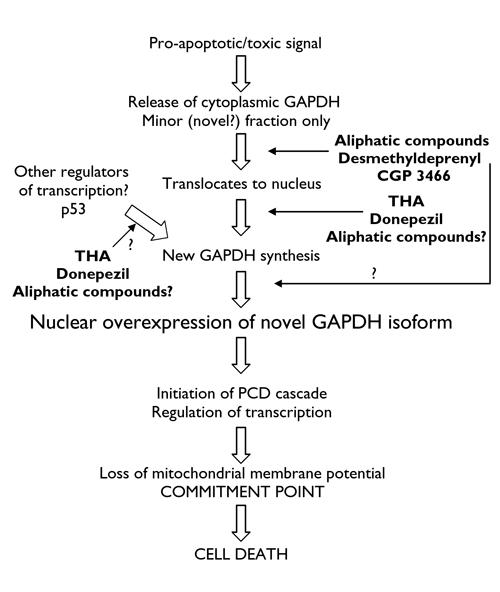
Fig. 3: A schematic representation of the GAPDH- mediated PCD cascade. A minor fraction of GAPDH is initially released from cytoplasmic binding sites. This minor fraction translocates to the nucleus and initiates synthesis of a novel nuclear-targeted GAPDH isoform. Transcription of this novel, nuclear GAPDH could also conceivably be induced by other factors. The novel, nuclear GAPDH then initiates PCD cascades through regulation of transcription, leading to loss of mitochondrial membrane potential and cell death. Small-molecule anti-apoptotic compounds have been identified that selectively bind to a fraction of GAPDH, preventing the increase in nuclear GAPDH and downstream events. Tetrahydroaminoacridine (THA) and donepezil have been reported to act at the level of new GAPDH transcription by binding to promoter regions. Aliphatic compounds, which like THA and donepezil have been shown to prevent increases in GAPDH mRNA, could also act at the level of transcription, although this has yet to be studied. Binding of small molecules to the newly synthesized GAPDH, thereby preventing its expression in the nucleus, is also possible in addition to binding to GAPDH released from cytoplasmic binding sites. The question marks above arrows indicate possible sites of action that have yet to be studied.
translocation of small amounts of existing GAPDH to the nucleus
altered p53 signalling, which has been reported to regulate GAPDH transcription35
hypoxia-induced transcription factors (the GAPDH gene contains hypoxia response elements43,44)
GAPDH, when present in the nucleus, has been shown to act as a transcriptional activator.45 In our model, there is an initial translocation of small amounts of GAPDH to the nucleus. This occurs following release of a small proportion of GAPDH from its cytoplasmic binding sites, which, it has been suggested, may occur in response to, for example, reactive oxygen species.46Translocation to the nucleus does not directly initiate PCD and occurs on a small scale, possibly below the detection limits for GFP-tagged GAPDH in most cells. The mechanism by which GAPDH enters the nucleus is unclear and may or may not require 1 or more chaperone proteins. The translocated GAPDH in the nucleus then acts as a transcription factor for the synthesis of new GAPDH protein, which is nuclear targeted and has characteristics of a novel isozyme (e.g., acidic pI values). The novel isozyme then acts as a transcription factor for pro-apoptotic signals. This model explains all the observations made so far and most notably explains the previous description of GAPDH antisense oligonucleotides preventing “translocation” of GAPDH to the nucleus, while still allowing for the translocation observed using GFP-tagged GAPDH. Further, whereas this model involves an initial, small-scale translocation of existing GAPDH protein, this is not an absolute requirement. Expression of the novel isozyme could equally be stimulated by other induced transcription factors or activators of transcription, although this requires verification. Interestingly, PCD-specific promoter regions have recently been described in the GAPDH gene,16 although their binding factors have yet to be identified.
GAPDH and anti-apoptotic compounds
From the preceding, it is readily apparent that GAPDH may serve as a target for small-molecule anti-apoptotic compounds with the potential to slow or prevent the progression of neurodegenerative disorders. Indeed, a number of well-documented anti-apoptotic agents have been shown to interact with GAPDH (Table 1).47,48,49 Although for 17β-estradiol this interaction has yet to be correlated with anti-apoptotic activity, it has been suggested that R-desmethyldeprenyl (the active metabolite of R-deprenyl) and CGP 3466 may be anti-apoptotic because of their binding to GAPDH.48,49
Table 1

We have developed a series of aliphatic compounds, initially using the propargylamine moiety found in deprenyl. The first example of these compounds, R-2HMP (R-N-(2-heptyl)-N-methylpropargylamine hydrochloride), has previously been reported to possess anti-apoptotic activity resulting from an interaction with the same binding site as R-deprenyl.50 In agreement with this, we have now obtained evidence that these compounds bind selectively to GAPDH. Intriguingly, using affinity columns and purified human recombinant GAPDH, these aliphatic compounds appear to bind only to a subset of GAPDH protein. As such, if unbound GAPDH (in excess of 90% of that initially loaded) from an initial run through an immobilized drug-affinity column is collected and added to a second naive drug-affinity column, very little further GAPDH binding is observed. This occurs despite the use of recombinant and, in theory, single-species GAPDH. It is of interest that although a single molecular weight GAPDH protein is identified with denaturing polyacrylamide gel electrophoresis, multiple bands are observed during isoelectric focusing. We are currently confirming these observations, and examining in more detail the nature of this apparent GAPDH subspecies. The possibility that the fraction of GAPDH that is selectively bound by our aliphatic compounds corresponds to a fraction of GAPDH that is more susceptible to release from cytoplasmic binding sites is intriguing and requires investigation. Likewise, the ability to distinguish a subset of an apparently homogeneous recombinant protein requires further study and explanation.
How binding of anti-apoptotic compounds to GAPDH can prevent apoptosis is unclear at present. Carlile et al48 have proposed that drug binding causes a change in the multimeric nature of GAPDH in solution. It has been proposed that this change may prevent GAPDH from entering the nucleus and, thus, by preventing initial translocation, apoptosis is prevented. We have not observed a similar change in the multimeric state of recombinant human GAPDH following aliphatic compound binding, despite having previously reported good evidence for these aliphatic compounds, deprenyl/desmethyldeprenyl and CGP 3466 being active because of an interaction with the same binding site. This may relate to species differences in the purified GAPDH used. We used a recombinant human GAPDH, whereas Carlile et al48 used GAPDH from Leishmania mexicana. Drugs designed to inhibit Leishmania but not human GAPDH have been described51 and, as such, species differences in the response of GAPDH to binding ligands are not without precedent. Further, we have observed no change in the multimeric nature of cytosolic GAPDH in cells induced to undergo apoptosis, in either the presence or absence of anti-apoptotic GAPDH ligands.
It is clear, however, that the compounds described here do prevent the increased nuclear accumulation of GAPDH. Because these compounds have been shown to bind to purified preparations of cytosolic GAPDH48,49 and at least the aliphatic compounds prevent the increase in GAPDH mRNA,52 it appears most likely that these compounds bind to cytosolic GAPDH and prevent its initial translocation to the nucleus following release from cytoplasmic binding sites. The possibility that at least some of these compounds show selective binding to released GAPDH is intriguing and merits further investigation. Whether such binding involves a change in the multimeric state of GAPDH in a cellular environment has yet to be conclusively demonstrated.
Finally, 3 compounds (tetrahydroaminoacridine [THA], donepezil and ONO-1603) have been described that, while preventing the increase in GAPDH mRNA and therefore nuclear GAPDH accumulation that occurs during PCD, thereby reducing apoptosis, do not bind to GAPDH. Donepezil and THA, both of which are marketed as symptomatic therapies for Alzheimer's disease, appear to be active by interacting with promoter regions distinct for the nuclear GAPDH.16 In contrast, ONO-1603, which is currently undergoing clinical trials as an antidementia compound, showed no such activity,16 and its mechanism of action requires further study. These studies raise the possibility that THA and donepezil may possess, in addition to symptomatic benefits, disease-modifying effects in patients with Alzheimer's disease. There do not appear to be, however, clinical reports suggesting such a disease-modifying action of either donepezil or THA.
Conclusion
In summary, various cell culture models have conclusively demonstrated that initiation of apoptosis by a variety of insults involves an increase in the nuclear expression of GAPDH. This increased nuclear expression is dependent on the synthesis of new GAPDH protein. Whereas some GAPDH translocation from the cytosol may occur, this appears to be on a small scale and results in the activation of GAPDH transcription. This new GAPDH protein appears to have novel characteristics and is nuclear targeted. This nuclear-targeted GAPDH then regulates transcription to initiate PCD cascades. There is now accumulating evidence that these observations may have relevance to human neurodegenerative conditions. As such, recent reports have shown increased nuclear GAPDH associated with susceptible neurons in postmortem samples from patients with a variety of neurodegenerative conditions. Small-molecule compounds with demonstrated anti-apoptotic activity that selectively interact with GAPDH have been identified. These compounds have also been shown to prevent the increase in nuclear GAPDH associated with the cell culture models in which they show anti-apoptotic activity. It is proposed that this effect is the result of binding to GAPDH, preventing the subsequent increase in GAPDH synthesis and nuclear accumulation. One of these compounds, CGP 3466, is currently undergoing Phase II clinical trials as a disease-modifying agent for Parkinson's disease, while others are in development.
Additional note
Following acceptance of this manuscript, Kusner et al53 reported nuclear accumulation of GAPDH in human retinal Muller cells undergoing apoptosis because of high glucose concentrations, effects that were prevented by treatment with R-deprenyl. In addition, Takasaki et al54 reported that GAPDH autoantigens were involved in the autoimmune response in patients with systemic lupus erythematosus, whereas Mazzola and Sirover55 provided evidence of structural changes in GAPDH protein associated with age-related neurodegenerative conditions. Further, Brown et al56 have reported that the nuclear accumulation of GAPDH is the result of a novel CRM-1-mediated nuclear export signal. Finally, it has been reported by Kanai et al57 that several histone deacetylase inhibitors that show anti-apoptotic activity also prevent the associated nuclear accumulation of GAPDH.
Acknowledgments
The author gratefully acknowledges Dr. Paula Ashe for helpful discussions and for making preliminary data available for publication, and he acknowledges Drs. Alan A. Boulton and Bruce A. Davis for their critical appraisal of the manuscript.
Footnotes
Competing interests: Dr. Berry was employed by ALviva Biopharmaceuticals at the time of writing and owns stock options in the company.
Correspondence to: Dr. Mark D. Berry, Department of Chemistry, J.R. Brodie Science Centre, Brandon University, Brandon MB R7A 6A9; fax 204 728-7346; berrym@brandonu.ca
Submitted June 3, 2003; Revised Oct. 15, 2003; Accepted Oct. 20, 2003
References
- 1.Sirover MA. Emerging new functions of the glycolytic protein, glyceraldehyde-3-phosphate dehydrogenase, in mammalian cells. Life Sci 1996;58:2271-7. [DOI] [PubMed]
- 2.Sirover MA. Role of the glycolytic protein, glyceraldehyde-3-phosphate dehydrogenase in normal cell function and in cell pathology. J Cell Biochem 1997;66:133-40. [PubMed]
- 3.Sirover MA. New insights into an old protein: the functional diversity of mammalian glyceraldehyde-3-phosphate dehydrogenase. Biochim Biophys Acta 1999;1432:159-84. [DOI] [PubMed]
- 4.Berry MD, Boulton AA. Glyceraldehyde-3-phosphate dehydrogenase and apoptosis. J Neurosci Res 2000;60:150-4. [DOI] [PubMed]
- 5.Lev N, Melamed E, Offen D. Apoptosis and Parkinson's disease. Prog Neuropsychopharmacol Biol Psychiatry 2003;27:245-50. [DOI] [PubMed]
- 6.Raina A, Hochman A, Ickes H II, Zhu X, Ogawa O, Cash AD, et al. Apoptotic promoters and inhibitors in Alzheimer's disease: Who wins out? Prog Neuropsychopharmacol Biol Psychiatry 2003; 27:251-4. [DOI] [PubMed]
- 7.Hickey MA, Chesselet MF. Apoptosis in Huntington's disease. Prog Neuropsychopharmacol Biol Psychiatry 2003;27:255-65. [DOI] [PubMed]
- 8.Love S. Apoptosis and brain ischaemia. Prog Neuropsychopharmacol Biol Psychiatry 2003;27:267-82. [DOI] [PubMed]
- 9.Barber AJ. A new view of diabetic retinopathy: a neurodegenerative disease of the eye. Prog Neuropsychopharmacol Biol Psychiatry 2003;27:283-90. [DOI] [PubMed]
- 10.Waldmeier PC. Prospects for antiapoptotic drug therapy of neurodegenerative diseases. Prog Neuropsychopharmacol Biol Psychiatry 2003;27:303-21. [DOI] [PubMed]
- 11.Berry MD, Ashe PC. Glyceraldehyde-3-phosphate dehydrogenase as a target for antiapoptotic drugs. In: LeBlanc AC, editor. Neuromethods: apoptosis techniques and protocols. Vol. 37. 2nd ed. Totowa (NJ): Humana Press; 2002. p. 149-61.
- 12.Burke JR, Enghild JJ, Martin ME, Jou YS, Myers RM, Roses AD, et al. Huntingtin and DRLPA proteins selectively interact with the enzyme GAPDH. Nat Med 1996;2:347-50. [DOI] [PubMed]
- 13.Klement IA, Skinner PJ, Kaytor MD, Yi H, Hersch SM, Clark HB, et al. Ataxin-1 nuclear localization and aggregation: role in polyglutamine-induced disease in SCA1 transgenic mice. Cell 1998;95:41-53. [DOI] [PubMed]
- 14.Saudou F, Finkbeiner S, Devys D, Greenberg ME. Huntingtin acts in the nucleus to induce apoptosis but death does not correlate with the formation of intranuclear inclusions. Cell 1998; 95: 55-66. [DOI] [PubMed]
- 15.Ishitani R, Fushiki S, Tatton NA, Tsuchiya K, Kuwae T, Yamada M, et al. Pro-apoptotic GAPDH protein: implications in the pathogenesis of neurodegenerative disorders [poster]. Soc Neurosci Abstr Vol. 27, Program No. 572.14, 2001. Available: http://sfn.scholarone.com/itin2001/index.html (accessed 2004 Aug. 18).
- 16.Ishitani R, Tajima H, Takata H, Tsuchiya K, Kuwae T, Yamada M, et al. Proapoptotic protein glyceraldehyde-3-phosphate dehydrogenase: a possible site of action of antiapoptotic drugs. Prog Neuropsychopharmacol Biol Psychiatry 2003;27:291-301. [DOI] [PubMed]
- 17.Shiozawa M, Fukutani Y, Arai N, Cairns NJ, Mizutani T, Isaki K, et al. Glyceraldehyde 3-phosphate dehydrogenase and endothelin-1 immunoreactivity is associated with cerebral white matter damage in dentatorubral-pallidoluysian atrophy. Neuropathology 2003;23:36-43. [DOI] [PubMed]
- 18.Mazzola JL, Sirover MA. Alteration of nuclear glyceraldehyde-3-phosphate dehydrogenase structure in Huntington's disease fibroblasts. Mol Brain Res 2002;100:95-101. [DOI] [PubMed]
- 19.Tatton NA. Increased caspase 3 and Bax immunoreactivity accompany nuclear GAPDH translocation and neuronal apoptosis in Parkinson's disease. Exp Neurol 2000;166:29-43. [DOI] [PubMed]
- 20.Fukuhara Y, Takeshima T, Kashiyawa Y, Mishima K, Ishitani R, Nakashima K. GAPDH antisense oligonucleotide protects dopaminergic neuronal death occurring with exposure of the cerebrospinal fluid from PD patients [poster]. Soc Neurosci Abstr Vol. 27, Program No. 198.17, 2001. Available: http://sfn.scholarone.com/itin20021/index.html (accessed 2004 Aug. 18).
- 21.Fukuhara Y, Takeshima T, Kashiwaya Y, Mishima K, Ishitani R, Nakashima K. (-)-Deprenyl and GAPDH antisense oligonucleotides rescue mesencephalic dopaminergic neurons from neuronal death induced by cerebrospinal fluid from PD patients (poster). Program no. 361.14. 2002 Abstract Viewer/Itinerary Planner. Washington: Society for Neuroscience; 2002. Available: http://sfn.scholarone.com/itin2002/index.html (accessed 2004 Aug. 18).
- 22.Sunaga K, Takahashi H, Chuang DM, Ishitani R. Glyceraldehyde-3-phosphate dehydrogenase is over-expressed during apoptotic death of neuronal cultures and is recognized by a monoclonal antibody against amyloid plaques from Alzheimer's brain. Neurosci Lett 1995;200:133-6. [DOI] [PubMed]
- 23.Schulze H, Schuler A, Stuber D, Dobeli H, Langen H, Huber G. Rat brain glyceraldehyde-3-phosphate dehydrogenase interacts with the recombinant cytoplasmic domain of Alzheimer's β-amyloid precursor protein. J Neurochem 1993; 60: 1915-22. [DOI] [PubMed]
- 24.Mazzola JL, Sirover MA. Subcellular alteration of glyceraldehyde-3-phosphate dehydrogenase in Alzheimer's disease fibroblasts. J Neurosci Res 2003;71:279-85. [DOI] [PubMed]
- 25.Mittag TW, Tatton NA, Wax M, Edward D, Chalmers-Redman RME, Tatton WG. Increased retinal ganglion cell (RGC) GAPDH, Bax and activated caspase 3 in a rat model and in human glaucoma [poster]. Association for Research in Vision and Ophthalmology meeting; 2001 Apr 25–May 4; Fort Lauderdale, Fla.
- 26.Tanaka R, Mochizuki H, Suzuki A, Katsube N, Ishitani R, Mizuno Y, et al. Induction of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) expression in rat brain after focal ischemia/reperfusion. J Cereb Blood Flow Metab 2002;22:280-8. [DOI] [PubMed]
- 27.Meyer-Siegler K, Mauro DJ, Seal G, Wurzer J, DeRiel JK, Sirover MA. A human nuclear uracil DNA glycosylase is the 37kDa subunit of glyceraldehyde-3-phosphate dehydrogenase. Proc Natl Acad Sci U S A 1991;88:8460-4. [DOI] [PMC free article] [PubMed]
- 28.Saunders PA, Chen RW, Chuang DM. Nuclear translocation of glyceraldehyde-3-phosphate dehydrogenase isoforms during neuronal apoptosis. J Neurochem 1999;72:925-32. [DOI] [PubMed]
- 29.Sawa A, Khan AA, Hester LD, Snyder SH. Glyceraldehyde-3-phosphate dehydrogenase: nuclear translocation participates in neuronal and nonneuronal cell death. Proc Natl Acad Sci U S A 1997;94:11669-74. [DOI] [PMC free article] [PubMed]
- 30.Dastoor Z, Dreyer JL. Potential role of nuclear translocation of glyceraldehyde-3-phosphate dehydrogenase in apoptosis and oxidative stress. J Cell Sci 2001;114:1643-53. [DOI] [PubMed]
- 31.Maruyama W, Oya-Ito T, Shamoto-Nagai M, Osawa T, Naoi M. Glyceraldehyde-3-phosphate dehydrogenase is translocated into nuclei through Golgi apparatus during apoptosis induced by 6-hydroxydopamine in human dopaminergic SH-SY5Y cells. Neurosci Lett 2002;321:29-32. [DOI] [PubMed]
- 32.Shashidharan P, Chalmers-Redman RME, Carlile GW, Rodic V, Gurvich N, Yuen T, et al. Nuclear translocation of GAPDH-GFP fusion protein during apoptosis. NeuroReport 1999; 10: 1149-53. [DOI] [PubMed]
- 33.Ishitani R, Chuang DM. Glyceraldehyde-3-phosphate dehydrogenase antisense oligodeoxynucleotides protect against cytosine arabinonucleoside-induced apoptosis in cultured cerebellar neurons. Proc Natl Acad Sci U S A 1996;93:9937-41. [DOI] [PMC free article] [PubMed]
- 34.Ishitani R, Sunaga K, Tanaka M, Aishita H, Chuang DM. Overexpression of glyceraldehyde-3-phosphate dehydrogenase is involved in low K+-induced apoptosis but not necrosis of cultured cerebellar granule cells. Mol Pharmacol 1997;51:542-50. [DOI] [PubMed]
- 35.Chen RW, Saunders PA, Wei H, Li Z, Seth P, Chuang DM. Involvement of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and p53 in neuronal apoptosis: evidence that GAPDH is upregulated by p53. J Neurosci 1999;19:9654-62. [DOI] [PMC free article] [PubMed]
- 36.Saunders PA, Chalecka-Franaszek E, Chuang DM. Subcellular distribution of glyceraldehyde-3-phosphate dehydrogenase in cerebellar granule cells undergoing cytosine arabinoside- induced apoptosis. J Neurochem 1997;69:1820-8. [DOI] [PubMed]
- 37.Fukuhara Y, Takeshima T, Kashiwaya Y, Shimoda K, Ishitani R, Nakashima K. GAPDH knockdown rescues mesencephalic dopaminergic neurons from MPP+-induced apoptosis. Neuroreport 2001;12:2049-52. [DOI] [PubMed]
- 38.Ishitani R, Tanaka M, Sunaga K, Katsube N, Chuang DM. Nuclear localization of overexpressed glyceraldehyde-3-phosphate dehydrogenase in cultured cerebellar neurons undergoing apoptosis. Mol Pharmacol 1998;53:701-7. [DOI] [PubMed]
- 39.Edwards YH, Clark P, Harris H. Isozymes of glyceraldehyde-3-phosphate dehydrogenase in man and other mammals. Ann Hum Genet 1976;40:67-77. [DOI] [PubMed]
- 40.Soukri A, Valverde F, Hafid N, Elkebbaj MS, Serrano A. Characterization of muscle glyceraldehyde-3-phosphate dehydrogenase isoforms from euthermic and induced hibernating Jaculus orientalis. Biochim Biophys Acta 1995;1243:161-8. [DOI] [PubMed]
- 41.Mezquita J, Pau M, Mezquita C. Several novel transcripts of glyceraldehyde-3-phosphate dehydrogenase expressed in adult chicken testis. J Cell Biochem 1998;71:127-39. [DOI] [PubMed]
- 42.Piechaczyk M, Blanchard JM, Riaad-El Sabouty S, Dani C, Marty L, Jeanteur P. Unusual abundance of vertebrate 3-phosphate dehydrogenase pseudogenes. Nature 1984;312:469-71. [DOI] [PubMed]
- 43.Graven KK, Yu Q, Pan D, Roncarati JS, Farber HW. Identification of an oxygen responsive enhancer element in the glyceraldehyde-3-phosphate dehydrogenase gene. BiochimBiophysActa 1999;1447:208-18. [DOI] [PubMed]
- 44.Lu S, Gu X, Hoestje S, Epner DE. Identification of an additional hypoxia responsive element in the glyceraldehyde-3-phosphate dehydrogenase gene promoter. Biochim Biophys Acta 2002;1574:152-6. [DOI] [PubMed]
- 45.Morgenegg G, Winkler GC, Hubscher U, Heizmann CW, Mous J, Kuenzle CC. Glyceraldehyde-3-phosphate dehydrogenase is a nonhistone protein and a possible activator of transcription in neurons. J Neurochem 1986;47:54-62. [DOI] [PubMed]
- 46.Tatton WG. Apoptotic mechanisms in neurodegeneration: possible relevance to glaucoma. EurJOphthalmol 1999; 9 (Suppl 1): S22-9. [DOI] [PubMed]
- 47.Ramirez VD, Kipp JL, Joe I. Estradiol, in the CNS, targets several physiologically relevant membrane-associated proteins. BrainResBrainResRev 2001;37:141-52. [DOI] [PubMed]
- 48.Carlile GW, Chalmers-Redman RME, Tatton NA, Pong A, Borden KE, Tatton WG. Reduced apoptosis after nerve growth factor and serum withdrawal: conversion of tetrameric glyceraldehyde-3-phosphate dehydrogenase to a dimer. MolPharmacol 2000;57:2-12. [PubMed]
- 49.Kragten E, Lalande I, Zimmermann K, Roggo S, Schindler P, Muller D, et al. Glyceraldehyde-3-phosphate dehydrogenase, the putative target of the antiapoptotic compounds CGP 3466 and R-(-)-deprenyl. JBiolChem 1998;273:5821-8. [DOI] [PubMed]
- 50.Berry MD. R-2HMP: an orally active agent combining independent antiapoptotic and MAO-B-inhibitory activities. CNSDrugRev 1999;5:105-24.
- 51.Suresh S, Bressi JC, Kennedy KJ, Verlinde CL, Gelb MH, Hol WG. Conformational changes in Leishmania mexicana glyceraldehyde-3-phosphate dehydrogenase induced by designed inhibitors. JMolBiol 2001;309:423-35. [DOI] [PubMed]
- 52.Zhang D. Anti-apoptotic actions of R-2HMP in cerebellar granule cells: changes of mitochondrial membrane potential and sub-cellular GAPDH protein [dissertation]. Saskatoon (SK): University of Saskatchewan; 2001.
- 53.Kusner LL, Sarthy VP, Mohr S. Nuclear translocation of glyceraldehyde-3-phosphate dehydrogenase: a role in high glucose-induced apoptosis in retinal Muller cells. Invest Ophthalmol Vis Sci 2004;45(5):1553-61. [PubMed]
- 54.Takasaki Y, Kaneda K, Matsushita M, Yamada H, Nawata M, Matsudaira R, et al. Glyceraldehyde 3-phosphate dehydrogenase is a novel autoantigen leading autoimmune responses to proliferating cell nuclear antigen multiprotein complexes in lupus patients. Int Immunol 2004;16(9):1295-304. [DOI] [PubMed]
- 55.Mazzola JL, Sirover MA. Subcellular analysis of aberrant protein structure in age-related neurodegenerative disorders. J Neurosci Methods 2004;137(2):241-6. [DOI] [PubMed]
- 56.Brown VM, Krynetski EY, Krynetskaia NF, Grieger D, Mukatira ST, Murti KG, et al. A novel CRM1-mediated nuclear export signal governs nuclear accumulation of glyceraldehyde-3-phosphate dehydrogenase following genotoxic stress. J Biol Chem 2004;279(7):5984-92. [DOI] [PubMed]
- 57.Kanai H, Sawa A, Chen RW, Leeds P, Chuang DM. Valproic acid inhibits histone deacetylase activity and suppresses excitotoxicity-induced GAPDH nuclear accumulation and apoptotic death in neurons. Pharmacogenomics J 2004;Aug 3 (electonic publication ahead of print). [DOI] [PubMed]