

Abstract
Background
Auxiliary partial liver transplantation (APLT) in humans is a therapeutic modality used especially to treat liver failure in children or congenital metabolic disease. Animal models of APLT have helped to explore therapeutic options. Though many groups have suggested improvements, standardizing the surgical procedure has been challenging. Additionally, the question of whether graft livers are reconstituted by recipient-derived cells after transplantation has been controversial. The aim of this study was to improve experimental APLT in rats and to assess cell recruitment in the liver grafts.
Methods
To inhibit recipient liver regeneration and to promote graft regeneration, we treated recipients with retrorsine and added arterial anastomosis. Using green fluorescence protein transgenic rats as recipients, we examined liver resident cell recruitment within graft livers by immunofluorescence co-staining.
Results
In the improved APLT model, we achieved well-regenerated grafts that could maintain regeneration for at least 4 weeks. Regarding the cell recruitment, there was no evidence of recipient-derived hepatocyte, cholangiocyte or hepatic stellate cell recruitment into the graft. Macrophages/monocytes, however, were consistently recruited into the graft and increased over time, which might be related to inflammatory responses. Very few endothelial cells showed co-localization of markers.
Conclusions
We have successfully established an improved rat APLT model with arterial anastomosis as a standard technique. Using this model, we have characterized cell recruitment into the regenerating grafts.
Introduction
Since the first report of auxiliary partial liver transplantation (APLT) in 19651, many groups have adopted this procedure. The results of APLT in humans during the initial 2 decades were discouraging2. In the 1990s, however, successful auxiliary partial orthotopic liver transplantation (APOLT) cases were documented in hepatic failure3,4 and metabolic liver disease5. Subsequently, several case series were reported6–10. Many of these achieved favorable results leading to recommendation of APLT, especially in childhood hepatic failure, due to its beneficial long-term effect on life expectancy and the possibility of immunosuppressive drug withdrawal. Some reports, however, suggested that APOLT was associated with higher mortality and morbidity rates compared with orthotopic liver transplantation (OLT)8,11. While APOLT has exhibited more rejection than OLT12,13, the exact mechanism underlying this phenomenon has yet to be elucidated. It has been postulated that resident macrophages (Kupffer) cells in native livers might play a role in promoting rejection in APLT13. Vascular and bile duct complications caused by technical problems or incorrect clinical indications have also been implicated in these relatively unfavorable results of APOLT10. As a result, APOLT has been performed by only a limited number of institutions6,7,14,15.
Auxiliary partial liver transplantation (APLT) with portal vein (PV) and arterial anastomosis in rats was introduced by Lee and Edgington in 196816. Hess et al17 improved the model by adding bile duct reconstruction of the graft and ligation of the native bile duct resulting in long-term graft survival. Although this APLT model was first reported more than 4nine decades ago, due to technical difficulty and relatively poor outcomes, it has not been used commonly as an experimental model. Several groups have tried to improve the model. Thus Yoo et al18 reported that PV reconstruction without hepatic arterial inflow could sustain the graft after APLT. Muller et al19 explored the PV arterialization technique and Marni et al20 proposed a cuff method to simplify the procedure and decrease operation time, achieving an improved survival rate. Overall, however, both technical difficulties and immunological challenges12,13 have precluded general application of the model.
To overcome these obstacles, recently we developed a simple technique of APLT in the rat that recapitulates many physiological effects of human procedures (see Matsubara et al, 2015)21 by incorporating standardized techniques for PV, bile duct reconstruction and the use of retrorsine (RS) (a pyrrolizidine alkaloid that blocks the hepatocyte cell cycle)22 before transplantation to allow regenerative advantage to the donor liver. Thus, providing for the first time, a new tool to study compensatory regeneration in transplanted liver grafts21.
With regard to cell repopulation in liver transplants, there are several reports that bone marrow (BM)-derived cells play an important role in renewal and repair during liver regeneration23–27. These studies suggest that hematopoietic stem cells are recruited to the graft and transdifferentiate into hepatic parenchymal or endothelial cells. However, these interpretations have been questioned following reports of spontaneous fusion of BM cells with other cell types28,29. Thus the role of BM cell recruitment or transdifferentiation in hepatic cell renewal remains controversial.
Establishment of an improved rat APLT model as a standard procedure would aid investigation of clinically relevant APLT and cell liver regeneration/immune cell recruitment. Here, based on our previously developed APLT model, we have improved the graft function and regeneration by incorporating arterial anastomosis and modifying bile duct reconstruction, and we characterized cell recruitment into the graft during regeneration.
Materials and Methods
Rat strains and care
Outbred Sprague-Dawley (SD) male rats were obtained from Charles River Laboratories and used for auxiliary liver transplantation as both donors and recipients. Nagase analbuminemic rats (NAR) of SD background were also used as recipients, Inbred Fisher (F)344 male rats (Charles River Laboratories) were used as donors and homozygous F344 rats expressing green fluorescence protein (GFP) under the ubiquitin C promoter (line 307; F344-Tg(EGFP) F455/Rrrc) obtained from the Rat Resource and Research Center (University of Missouri, Columbia, MO) used as recipients. For APLT, 200 – 350g rats were used as donors or recipients. The animals were maintained in accordance with the guidelines established by the Committee on Laboratory Resources, National Institutes of Health and experiments were conducted under an Institutional Animal Care and Use Committee - approved protocol.
Experimental groups
Five groups were constituted (total, n=28): groups 1 and 2, SD recipients transplanted from SD donors (each group, n=8); groups 3 and 4, NAR recipients transplanted from SD donors (each group; n = 3). To evaluate cell recruitment into the liver graft, F344 GFP transgenic (tg) rats were used as recipients and F344 rats as donors (group 5; n = 6). Group 1 and 2 rats were euthanized at 2 and 4 weeks and group 3 and 4 rats at 4 weeks after APLT. Group 5 rats were euthanized at 1, 2 and 4 weeks postoperatively. Retrorsine (RS) (Sigma Chemical Co., St. Louis, MO) was administered to graft recipients in groups 1, 3 and 5 to inhibit the mitosis of host native hepatocytes, as previously described21, 22. Briefly, 2 intra-peritoneal injections of 30 mg/kg RS were administered to rats weighing 90 – 140 g, 2 weeks apart. Four weeks after the second injection, APLT with 1/3 hepatectomy was performed in each rat.
Donor surgery
Donor surgery was performed as described21. We improved the previous model by adding arterial anastomosis. The celiac artery (CeA) were skeletonized, the side branches (left gastric artery, splenic artery and gastroduodenal artery) were ligated and divided. An arterial stent made from 22 or 24-gauge catheter (BD) was inserted into the celiac artery from the proximal side and secured in place with 7-0 silk.
Recipient surgery (see Supplementary Video online)
After laparotomy, right nephrectomy and IVC anastomosis (Figure 1A) was performed as described21. For arterial anastomosis (Figure 1B), the operating table was rotated 45° anti-clockwise. The graft CeA was anastomosed to the recipient right renal artery (RA) using a 22 or 24 G stent. Just after completion of arterial anastomosis, the IVC clamp and arterial clamp were removed and the graft re-perfused with arterial flow. At the same time, the proximal side of the PV was clamped to avoid backflow bleeding. The graft SMV was then anastomosed to the recipient PV/SMV confluence by end-to-side anastomosis (Figure 1C). After completing the PV anastomosis, the vascular clamp on the PV was released and PV flow provided to the graft. Following reperfusion, the PV between the pyloric vein and the PV anastomosis was ligated completely using 6-0 silk to increase inflow to the graft. After vascular anastomoses, a left lateral lobectomy (1/3 hepatectomy) of the native liver was performed.
Figure 1. Images and schematic drawings of each anastomosis during APLT in the rat and postoperative appearance of the graft and native liver.
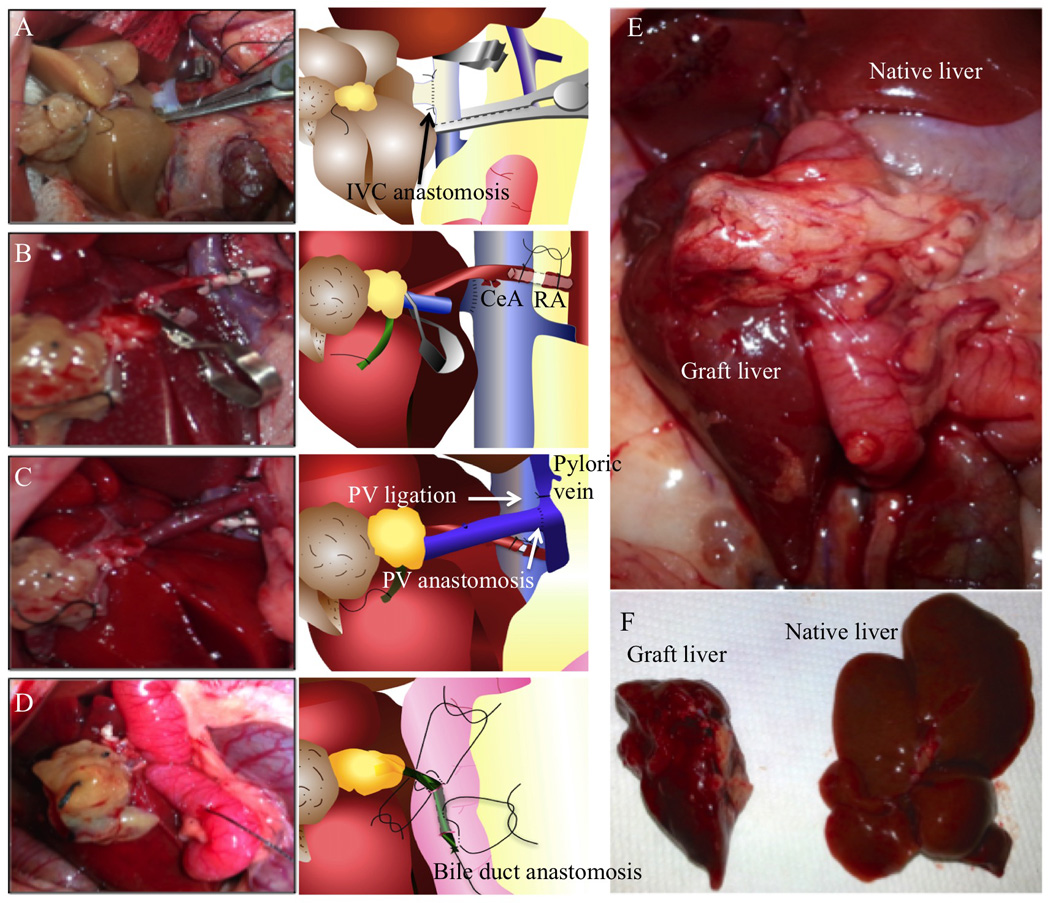
(A) IVC anastomosis. After clamping the proximal and distal ends of the IHIVC, the anterior wall of the IHIVC was cut to make a 7 mm vertical incision, as close to the ligated right renal vein as possible, for the IVC anastomosis. A stay suture was placed with 10-0 nylon at the distal end before beginning the continuous suture. Prior to completion of the anastomosis, saline was injected slowly using a L-shaped injector into the IHIVC anastomosis to remove any air trapped inside. (B) Arterial anastomosis. The thread tied to the right RA was held and pulled to the left side of the recipient. The proximal side of the right RA was clamped and the anterior wall of the artery cut with scissors and the blood inside the artery washed out. Then, the arterial stent tied on the graft celiac artery was inserted into the recipient’s right RA. The arterial stent was ligated in place with 7-0 silk to complete the arterial anastomosis. (C) PV anastomosis. The inside of the graft PV was washed and 2 stay sutures placed from outside to inside on each edge of the graft SMV using 10-0 nylon. The superior mesenteric artery (SMA) was clamped during PV anastomosis to minimize intestinal congestion. The splenic vein, the PV and SMV were clamped, to provide space for the anastomosis. The PV was cut about 4 mm on the opposite side of the splenic vein for anastomosis. An end-to-side anastomosis was performed between the graft SMV and the recipient’s PV. (D) Bile duct reconstruction. A small stitch was placed on the duodenal wall and on the right side wall of the graft bile duct close to the hilum using 10-0 nylon. Then, another small stitch was placed from the duodenum to the left side of the bile duct, 2 mm proximal to the previous suture. A 24-gauge needle was inserted into the duodenum 1 cm distal from the sutures and exited from the duodenal lumen between the sutures to introduce the bile duct into the duodenum. The anterior wall of the bile duct was then cut to create an opening for bile drainage. The bile duct was pulled with the bile duct opening inside the duodenum to avoid bile leakage and the 10-0 sutures tied to connect the duodenum and the bile duct. One Lembert suture was placed on the duodenum to close the insertion hole from the 24-gauge needle. (E) Appearance of the graft and native livers in situ 4 weeks after surgery. The liver graft has regenerated well and is similar macroscopically to the native liver. (F) Appearance of the liver graft (left) and native liver (right) following excision 4 weeks after surgery.
The bile duct was reconstructed using choledocho-duodenostomy (Figure 1D). Two stitches were placed between the bile duct and the duodenal wall and the graft bile duct inserted into the recipient duodenum.
Postoperative management
Following surgery, 1.0 mg/kg tacrolimus was injected intramuscularly each day until the end of the experiment30,31.
Assessment and definition of “graft weight ratio”
To evaluate graft regeneration, the following formulae were applied:
pre GRWR = preoperative graft weight/recipient body weight (at APLT surgery);
post GRWR = postoperative graft weight/recipient body weight (at euthanasia);
“graft weight ratio” = post GRWR/pre GRWR
Serum albumin levels
Serum albumin levels were determined in serum samples obtained from NAR (group 3 and 4) 1, 3, 7, 14, 21 and 28 days after APLT by enzyme-linked immunosorbent assay (ELISA) (Bethyl Lab, Montgomery, TX).
Immunohistochemistry
Epithelial cell adhesion molecule (EpCAM) expression was assessed in 5 µm sections, prepared from 4% paraformaldehyde-fixed, paraffin-embedded tissue. After Ag retrieval in citrate buffer, the samples were incubated with rabbit anti-EpCAM (Abcam ab71916) 1:500. For immunoperoxidase detection, the Vectastatin ABC kit Elite PK-6101 rabbit IgG and ImmPACT DAB peroxidase substrate kit SK-4105 were used.
Immunofluorescence staining
For albumin and albumin/Ki67 co-staining, paraffin sections were incubated with primary Abs: sheep anti-albumin (Bethyl A110-134A) 1:100 and mouse anti-Ki67 (BD 550906) 1:50 at 4°C overnight. Alexa Fluor 488 anti-mouse IgG (Invitrogen) 1:250 and Jackson Cy3 anti-sheep 1:100 were used as secondary Abs. To assess proliferation, positive and negative Ki-67 hepatocytes were counted in 5 fields per section in each group at ×400 magnification and positive hepatocytes expressed as a percentage.
For GFP co-staining together with cell surface markers, sections were fixed in 2% paraformaldehyde, followed by 30% sucrose for 24 hr each, then snap frozen. Four µm sections were post-fixed in 4% paraformaldehyde, followed by incubation with primary Abs at 4°C overnight. The primary Abs and dilutions were: rabbit anti-GFP (Novus NB600-308), 1:1000; goat anti-GFP (Abcam ab5450), 1:1000; sheep anti-albumin (Bethyl A110-134A), 1:100; rabbit anti-EpCAM (Abcam ab71916), 1:100; rabbit anti-von Willebrand factor (vWF) (Abcam ab6994), 1:200; mouse anti-alpha smooth muscle actin (αSMA) (Abcam ab7817), 1:100; and mouse anti-CD68 (Serotec MCA341GA), 1:100. The secondary Abs were: Alexa Fluor 488 anti-rabbit/goat IgG (Invitrogen), 1:250; Alexa Fluor 594 anti-sheep IgG (Invitrogen), 1:250 and Jackson Cy3 anti-rabbit/mouse, 1:100. Sections were mounted using Vectashield mounting medium with 4,6-diamino-2-phenylindole (Vector) counterstaining of cell nuclei.
Statistics
All statistical analyses were performed using SPSS® version 23.0 (IBM, Armonk, New York,) and the Mann-Whitney U test was used for analysis of continuous variables. P ≤ 0.05 was considered statistically significant. Data are expressed as means ± SEM.
Results
Technologic refinements and surgical outcomes
After establishing the surgical model, 54 APLTs were performed. The recipient mortality rate was 25.9 % (n=14) and the morbidity rate was 30.0 % (n=12). The causes of mortality were graft congestion (42.7 %, n=6), bleeding (21.4 %, n=3), major bile leakage (14.3 %, n=2) and other causes (14.3 %, n=2). Causes of morbidity were bile obstruction (83.3 %, n=10) and localized bile leakage (16.7 %, n=2). Bile leakage and obstruction were the main complications of this procedure. The use of a stent for bile duct reconstruction produced a bile stone on the duodenal side of the stent in long-term survival cases; therefore the reconstruction method was modified (see bile duct reconstruction in the Materials and Methods). Arterial anastomosis was added to increase blood supply to the bile duct wall and decreased the morbidity rate of the choledocho-duodenostomy. Successful cases (Figure 1E and F, n=28) were defined as having a “graft weight ratio” > 1.0 and histologically less bile duct proliferation compared with complicated cases. In our experience, PV anastomosis completed within 30 min minimizes intestinal congestion or PV thrombosis and reduces mortality.
Histological assessment of graft livers after APLT
To assess the condition of the grafts, bile duct (EpCAM) and hepatocyte (albumin) marker expression was examined at 14 and 28 postoperative days. Graft livers showed high bile duct proliferation in complicated cases (n = 10). In successful cases (n = 28), although some bile duct proliferation was evident, the bile duct proportion in the parenchymal space was < 20 % (Figure 2A). There was no observable difference in bile duct proliferation between 2 and 4 weeks after surgery.
Figure 2. Histological appearance of liver graft and assessment of graft function.
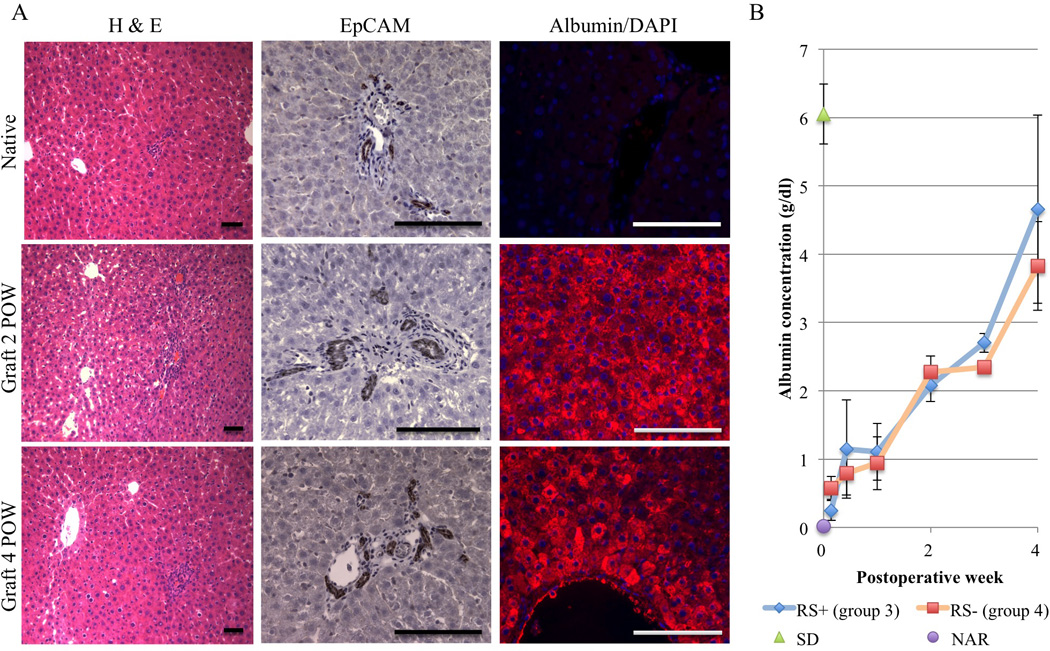
(A) H & E staining and immunohistochemical analyses of EpCAM and albumin expression in NAR native liver as a control and in the liver graft at 2 and 4 postoperative weeks (POW; representatives of groups 1 and 3). Scale bar = 50 µm. (B) Time course of serum albumin levels determined by ELISA in NAR recipient rats with normal SD grafts. Data show post-transplant RS+ NAR (group 3; n = 3) and RS− NAR (group 4; n = 3) albumin levels over time. As controls normal SD (n = 3) and NAR (n = 3) albumin levels are also shown at 0 POW. Values are shown as means ± SEM.
Serum albumin levels in analbuminemic rats with APLT graft
In groups 3 (n= 3) and 4 (n= 3), NARs were used as recipients to allow assessment of graft function by serum albumin measurement. Recipient sera were collected 1, 3, 7, 14, 21 and 28 days after APLT and ELISA performed. Serum albumin levels increased continuously for 4 weeks and reached 2/3 of the normal control level (serum albumin level of SD rats). There is no detected difference between the groups 3 (RS+) and 4 (RS−) in this study (Figure 2B).
Regenerative capacity of the APLT graft
Graft regenerative capacity was assessed in RS (groups 1; n= 8 and 3; n= 3) versus non-RS (groups 2; n= 8 and 4; n= 3) groups. Ki67 and albumin co-staining was performed on graft and native livers with RS treatment 2 weeks after APLT (Figure 3A). Ki67 analyses showed that the proportion of Ki67+ cells in the native liver was constantly low 2 and 4 weeks after APLT in both the RS and non-RS group (Figure 3B, left panel). Compared to native livers, Ki67+ cells in the grafts increased 2 weeks after APLT to almost the same level in both groups 1 (RS+; SD recipient) and 2 (RS−; SD recipient), indicating that the grafts in both groups were regenerating at 2 weeks. Four weeks after APLT, Ki67+ cells in the grafts of group 3 (RS+; NAR recipient) were significantly higher than those in group 4 (RS−; NAR recipient) (p = 0.009), suggesting that RS treatment was necessary for prolonged regeneration in the graft (Figure 3B, right panel). One reason that native livers regenerated less in the non-RS group could be decreased portal flow to the native liver because of PV ligation (Figure 1C).
Figure 3. Assessment of the regenerative capacity of the transplanted liver in RS-treated (group 1 and 3) versus non RS-treated recipients (group 2 and 4).
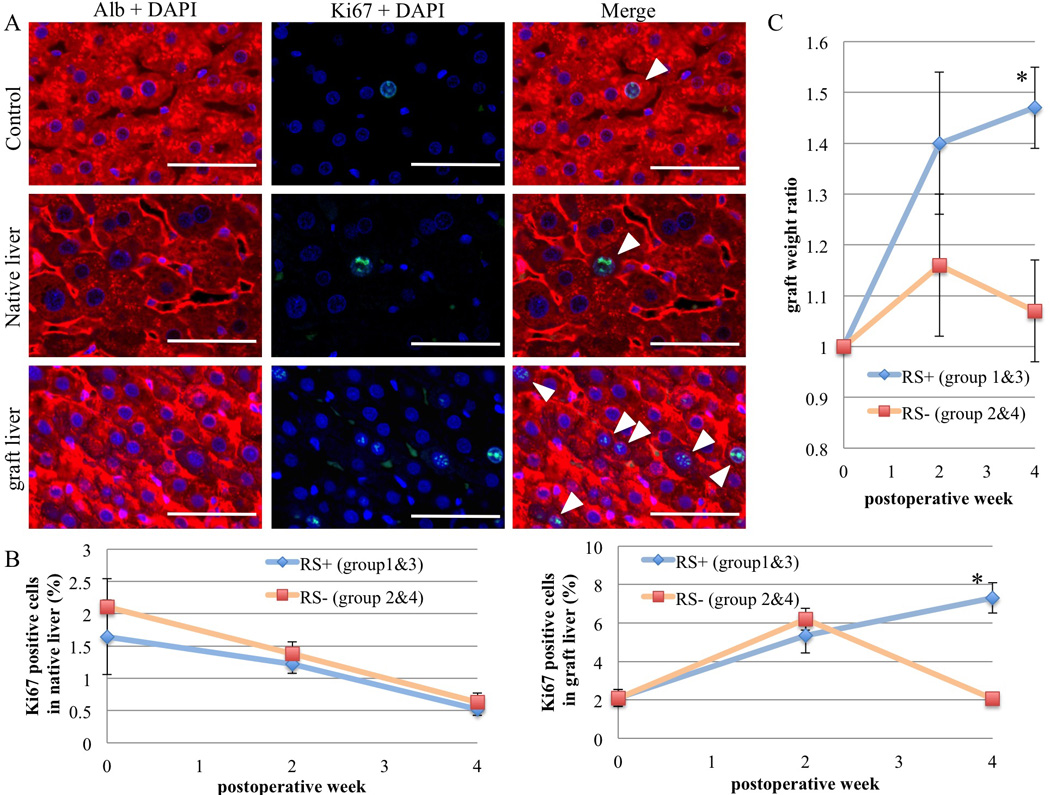
(A) Ki67/Albumin co-immunofluorescence staining in the graft and native liver with RS treatment 2 weeks after APLT. White arrowheads indicate Ki67+ cells. Scale bars = 50 µm. (B) To assess proliferation, Ki67 positive and negative hepatocytes were counted in 5 high power fields of each group at ×400 magnification and positive hepatocytes expressed as a percentage of total hepatocytes. (C) “Graft weight ratio” at 2 and 4 weeks after APLT comparing the RS and non RS-treated groups. Values are means ± SEM. *, P ≤ 0.05.
“Graft weight ratios” in the RS treatment groups were significantly higher than those in non-RS groups, especially at postoperative week 4 (p = 0.05), while non-RS group ratios decreased at 4 weeks after increasing at 2 weeks (Figure 3C). These results also indicate that RS treatment could prolong graft regeneration for at least 4 weeks. The total liver weight to body weight ratios were consistently in the range of 3.5 – 4.0 % for all groups.
Migration of recipient-derived GFP+ cells into the graft liver
To assess cell recruitment to the APLT, GFP− liver grafts were transplanted into RS-treated GFP tg rats. To evaluate cell recruitment from the host to graft livers, markers for hepatocytes (Alb), cholangiocytes (EpCAM), endothelial cells (vWF), hepatic stellate cells (αSMA) and Kupffer cells/monocytes (CD68) were co-stained with GFP immunofluorescence. Since all recipient native liver cells were GFP+, they co-localized with these different markers (Figure 4). In contrast, in the liver grafts, some GFP+ cells were found, mainly in perivascular areas. We detected co-expression of GFP and CD68 at 1, 2 and 4 postoperative weeks (POW) (Figure 4 and 5); first with a perivascular distribution, then after 4 weeks, with a more homogenous distribution in the parenchyma. These co-expressing cells increased over time (Figure 6). At 4 weeks postsurgery, very few graft endothelial cells co-expressed GFP and vWF (Figures 4 and 5).
Figure 4. Detection of host-derived cells in liver grafts.
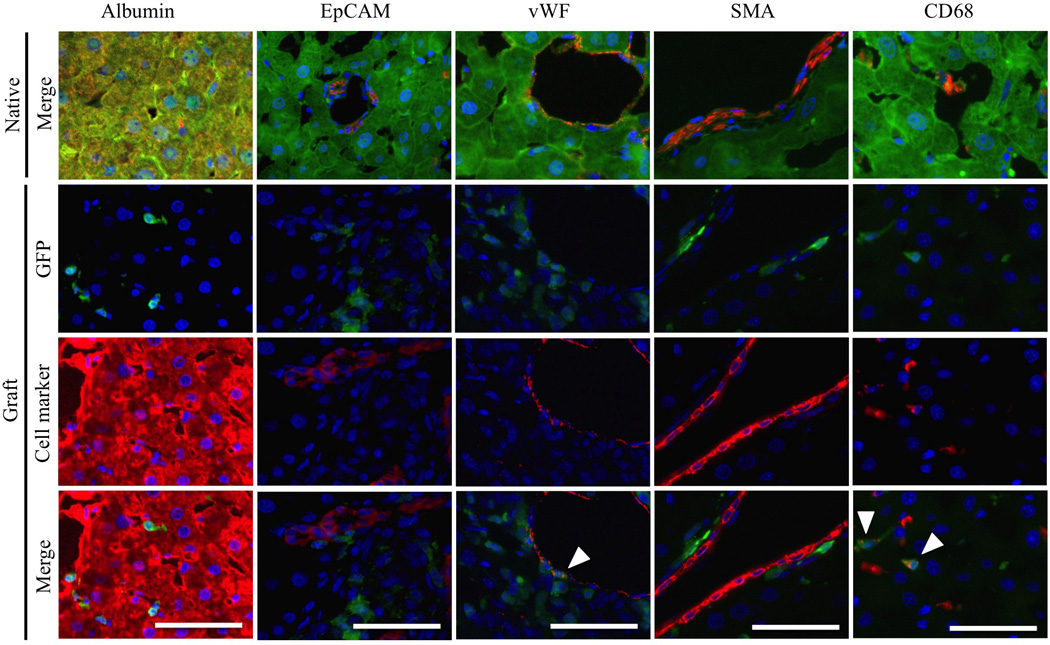
Staining was performed on native livers and liver grafts for hepatocytes (Alb), cholangiocytes (EpCAM), sinusoidal and vessel endothelial cells (vWF), hepatic stellate cells (αSMA) and Kupffer cells/monocytes (CD68), together with GFP immunofluorescence, 4 weeks after APLT. White arrowheads indicate co-localization. Scale bars = 50µm.
Figure 5. Co-localization of GFP and CD68 or vWF immunofluorescence staining in liver grafts 4 weeks after transplantation.
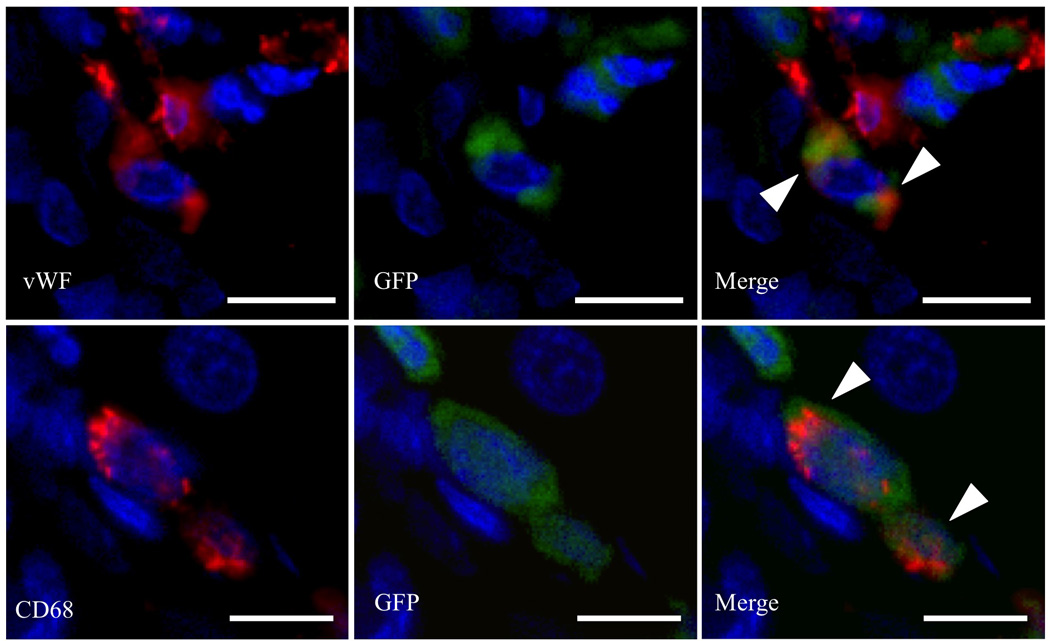
White arrowheads indicate co-localization. Scale bars = 10µm.
Figure 6. Host-derived CD68+/GFP+ cells increase over time after APLT.
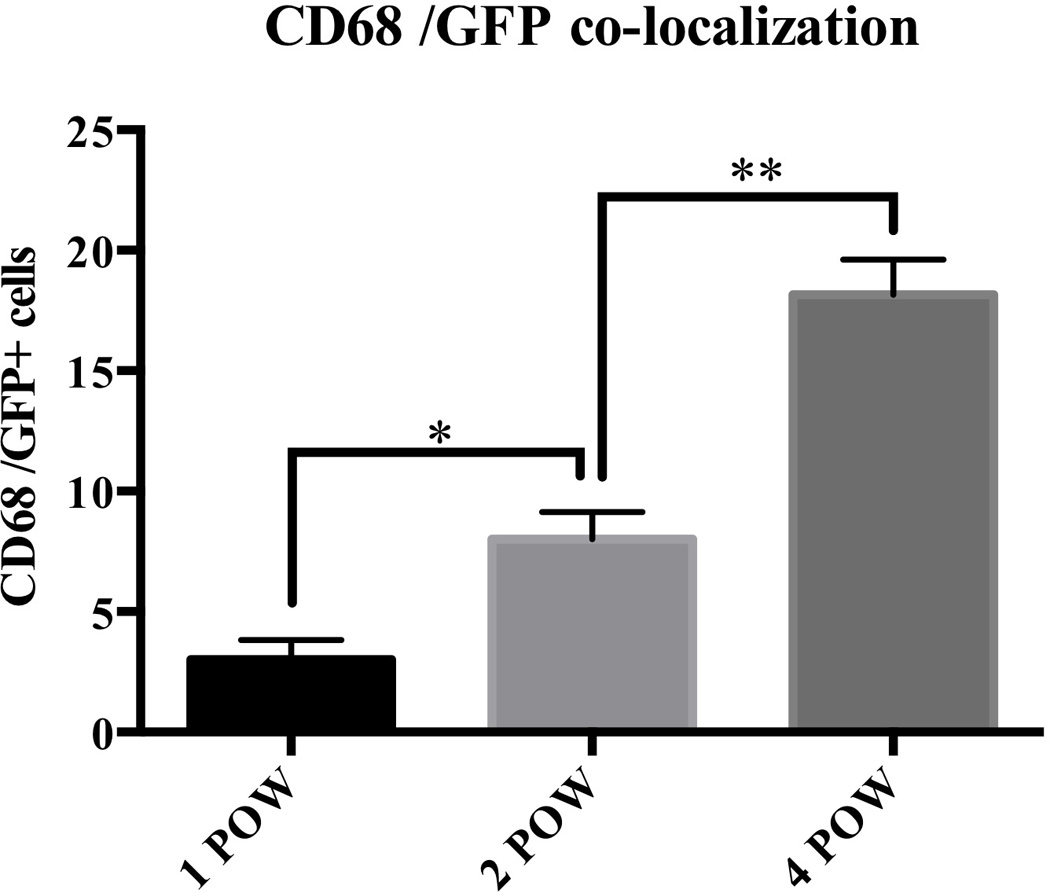
To assess co-localization of CD68+/GFP+ cells in the liver graft, double-positive cells were counted in 5 fields per representative of each group at ×200 magnification. These co-expressing cells increased over time (1POW; 3.0 ± 0.82, 2POW; 8.0 ± 1.1, 4POW; 18 ± 1.4 cells). POW = 4. Scale bars = 50µm. Values are shown as means ± SEM. *, P = 0.0238, **, P = 0.0043.
Discussion
Several reports have addressed technical issues concerning heterotopic APLT in the rat. Thus, Yoo et al18 suggested that PV reconstruction without arterial anastomosis could be sufficient to sustain partial liver transplants long-term. However, graft survival in their study was low (<50% by 30 postoperative days) and they also ligated the native bile duct; consequently the native liver shrank, similarly to the procedure of Hess et al with a survival rate of only 44.4%17. Others have reported portal arterialization19 producing a survival rate at 14 days of 77.7%; however, in these studies, hepatocytes did not survive long-term with arterial blood flow alone32,33. Marni et al20 also achieved a high survival rate of 80% at 2 weeks and 73.3% at 8 weeks by using a cuff technique for PV and IVC anastomosis. However, in their study arterial anastomosis was not performed, and the morbidity rate and histological assessment of the graft were not reported. The survival rate in the present study was 74% at 4 weeks.
The procedure in the present study was adopted from a model previously developed by Matsubara et al21 and incorporates similar characteristics to clinical APLT and maintained healthy conditions in both the graft and native livers for at least 4 POW with a morbidity rate of 30%. When the native livers were treated with RS, the grafts increased their regeneration rates and compensated for native liver disability. Indeed, grafts in the RS-treated group maintained regenerative function for at least 4 weeks, which was longer than in the non-RS treated group.
In the current study, the impairment of native liver regeneration by RS treatment continued for 4 weeks. If native livers are healthy, both native and graft livers orchestrate a “hepatostat” balance, that ensures liver weight is controlled for the performance of its homeostatic functions34. In the present model, 1/3 of the donor liver was transplanted and 1/3 hepatectomy was performed on the recipient. As a result, total liver volume was maintained without massive hepatocyte loss. If the hepatostat was driven mainly by the hepatic mass, the liver would not regenerate, but even in the non-RS treatment group, the graft liver regenerated within 2 POW to the same level as in the RS-treated group (Figure 3).
Liver regeneration after hepatectomy usually completes 1 week after surgery35,36, however, the grafts in our model maintained their regenerative capacity for at least 2 weeks. This may be because the graft received adequate PV blood flow essential for liver regeneration37,38 compared with the native liver, that had reduced portal venous flow (only pyloric venous flow remaining), and likely retained the stimulus for regeneration. Recent studies39,40 suggest that portal pressure plays an important role in modulating liver regeneration. In the present study, portal flow was completely re-directed to the graft, thus, maintaining an adequate portal pressure for its regeneration. Future studies could be directed to elucidate the role of portal pressure on regulating graft regeneration in this model by modulating the extent of portal ligation.
On the other hand, although graft regeneration in the non-RS treated group did not continue for 4 weeks, serum albumin levels increased continuously during this period. Even if there were differences in graft size, albumin production was not significantly different between the non-RS and RS groups. This may be due to the fact that the liver grafts already had a large liver mass (nearly 40% of a whole liver) able to produce albumin and also since albumin has a long half-life (approximately 20 days) and could accumulate over time. However, additional APLTs will have to be performed in greater numbers on analbuminemic rats in the future to study significant differences in the capacity to produce serum albumin. Moreover, previously, other APLT models, with only portal arterialized flow, have been used for bioengineered liver grafts implantation41,42. The advantage of the present model is that it ensures both arterial and PV flow, mimicking physiologically the same conditions as in the native liver, thus, representing an adequate APLT model to test the function and regeneration of bioengineered livers.
Many investigators have attempted to elucidate the mechanistic basis of liver regeneration. In regard to cell recruitment into the liver, Wang et al43,44 suggested that sinusoidal endothelial cells that transdifferentiated from BM cells could repopulate chronically-injured livers, whereas Okabayashi et al45 reported that host BM stem cells could differentiate into hepatocytes under specific conditions. Hepatocytes are assumed to have the capacity to transdifferentiate into cholangiocytes46,47, but whether and how BM cells can constitute hepatic cells remains controversial48. Here, we investigated cell origin/cell migration into the regenerating liver graft by performing immunofluorescence co-staining. Markers specific for hepatocytes, cholangiocytes, endothelial cells, stellate cells and Kupffer cells/monocytes were used to determine the origin of cells within the APLT. There was no evidence of hepatocyte, cholangiocyte or hepatic stellate cell migration into the grafts. CD68+ cells (Kupffer cells/monocytes) however, were consistently recruited into the grafts and increased over time from a perivascular to a parenchymal sinusoid location.
Recent studies have revealed that Kupffer cells are derived from embryonic progenitors in the yolk sac and maintained by self-renewal in situ49,50. Thus, the recruitment of CD68+ cells into the graft might be related to inflammatory responses of circulating blood monocytes. Very few cells exhibited co-expression of vWF and GFP only in the cytoplasm, which could be the result of endothelial cell phagocytosis. Our finding supports previously published evidence29 that only inflammatory cells are detected in human liver grafts. However, the limitation of this study is that we have investigated only liver resident cells and have not evaluated the significance of liver stem cells, nor the role of other cells that were recruited into the graft. Also, this study has focused on early regeneration and not long-term regeneration. Further investigation should be conducted to elucidate the mechanistic basis of liver regeneration.
An important aspect affecting the outcome after APLT in the clinics is graft atrophy due to early and late acute graft rejection. Different contributor factors have been associated to especially late acute rejection, for instance, lowering immunosuppression51, native liver affecting allograft tolerance12,13,19, competition of metabolic function and/or portal circulation between the graft and the native liver52. However, the etiologies of this issue have not been clearly elucidated. Although in these particular experimental settings, we did not investigate the immunological rejection of the liver graft. The described APLT model could be used for future experiments to study graft rejection and the role of immune cells (eg T cells, NK cells, granulocytes, dendritic cells).
In summary, we have successfully established a rat APLT model with arterial anastomosis. Using this model, we have evaluated cell recruitment into the graft. Of the graft cells post-APLT, only macrophages/monocytes were recruited into the grafts and these findings support a self-renewal ability of the liver. This improved model may prove valuable in addressing questions related to liver regeneration, recruitment, immunology and transplantation of bioengineered livers.
Supplementary Material
{kind=link}
Acknowledgments
We thank the Center for Biologic Imaging at the University of Pittsburgh for microscope technical support and Dawn E. McCormick PhD., University of Pittsburgh, for editing this article and giving helpful advice. We also thank Dr. Ira J. Fox for providing the starting colony of Nagase analbuminemeic rats.
Funding
Funding from the US National Institutes of Health (DK099257 to A.S.-G. and DK090325 to M.O.) and the Competitive Medical Research Fund Program of the UPMC Health System and the University of Pittsburgh Physicians (UPP) Academic Foundation to A.S.-G. Also funding from Japan Agency for Medical Research and Development (AMED), Research Center Network for Realization of Regenerative Medicine Projects for Technological Development to H.Y. The Amigos del Instituto Nacional de Ciencias Medicas y Nutricion Foundation funded the stipend of A.P-G.
Abbreviations
- αSMA
alpha smooth muscle actin
- APLT
auxiliary partial liver transplantation
- BM
bone marrow
- CeA
celiac artery
- ELISA
enzyme-linked immunosorbent assay
- EpCAM
epithelial cell adhesion molecule
- GFP
green fluorescence protein
- GRWR
graft-recipient weight ratio
- IHIVC
infra-hepatic inferior vena cava
- IVC
inferior vena cava
- NAR
Nagase analbuminemic rats
- OLT
orthotopic liver transplantation
- PBS
phosphate-buffered saline
- POW
postoperative weeks
- PV
portal vein
- RA
renal artery
- RS
retrorsine
- SD
Sprague-Dawley
- SHIVC
supra-hepatic inferior vena cava
- SMA
superior mesenteric artery
- SMV
superior mesenteric vein
- tg
transgenic
- vWF
von Willebrand factor
Footnotes
Author contributions
- Substantial contributions to the conception or design of the work; or the acquisition, analysis, or interpretation of data for the work;
- Drafting the work or revising it critically for important intellectual content;
- Final approval of the version to be published;
- Agreement to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved;
As specific contributions, Y.O., K.M., H.Y. and A.S-G., designed the study; Y.O., S.Y., and K.M. performed the APLT surgical model; Y.O, A. P-G. and J. G-L performed immunohistochemistry and immunofluorescence staining; Y.O., K.H., A.P-G., J. G-L, A.C.I., K.M., H.Y., M.O., and A.S-G. analyzed and interpreted data; M.I.Y and M.O., breed and prepared F344 rats expressing green fluorescence protein; Y.O., A.P-G., S.Y., K.M., M.I.Y, D.A.G., A.W.T., H.Y., M.O., A.S-G. prepared the manuscript; A.S-G. obtained funding for these studies. All authors revised the manuscript.
Disclosures
There is no authors’ conflict of interest to disclose as described by the ICMJE
References
- 1.Absolon KB, Hagihara PF, Griffen WO, Jr, Lillehei RC. Experimental and clinical heterotopic liver homotransplantation. Rev Int Hepatol. 1965;15(8):1481–1490. [PubMed] [Google Scholar]
- 2.Terpstra OT, Schalm SW, Weimar W, et al. Auxiliary partial liver transplantation for end-stage chronic liver disease. N Engl J Med. 1988;319(23):1507–1511. doi: 10.1056/NEJM198812083192303. [DOI] [PubMed] [Google Scholar]
- 3.Metselaar HJ, Hesselink EJ, de Rave S, et al. Recovery of failing liver after auxiliary heterotopic transplantation. Lancet. 1990;335(8698):1156–1157. doi: 10.1016/0140-6736(90)91158-7. [DOI] [PubMed] [Google Scholar]
- 4.Boudjema K, Jaeck D, Simeoni U, Bientz J, Chenard MP, Brunot P. Temporary auxiliary liver transplantation for subacute liver failure in a child. Lancet. 1993;342(8874):778–779. doi: 10.1016/0140-6736(93)91542-t. [DOI] [PubMed] [Google Scholar]
- 5.Whitington PF, Emond JC, Heffron T, Thistlethwaite JR. Orthotopic auxiliary liver transplantation for Crigler-Najjar syndrome type 1. Lancet. 1993;342(8874):779–780. doi: 10.1016/0140-6736(93)91543-u. [DOI] [PubMed] [Google Scholar]
- 6.Kato T, Selvaggi G, Levi D, et al. Routine use of auxiliary partial orthotopic liver transplantation for children with fulminant hepatic failure: Preliminary report. Transplant Proc. 2006;38(10):3607–3608. doi: 10.1016/j.transproceed.2006.10.038. [DOI] [PubMed] [Google Scholar]
- 7.Faraj W, Dar F, Bartlett A, et al. Auxiliary liver transplantation for acute liver failure in children. Ann Surg. 2010;251(2):351–356. doi: 10.1097/SLA.0b013e3181bdfef6. [DOI] [PubMed] [Google Scholar]
- 8.Kasahara M, Takada Y, Egawa H, et al. Auxiliary partial orthotopic living donor liver transplantation: Kyoto University experience. Am J Transplant. 2005;5(3):558–565. doi: 10.1111/j.1600-6143.2005.00717.x. [DOI] [PubMed] [Google Scholar]
- 9.Rela M, Muiesan P, Vilca-Melendez H, et al. Auxiliary partial orthotopic liver transplantation for Crigler-Najjar syndrome type I. Ann Surg. 1999;229(4):565–569. doi: 10.1097/00000658-199904000-00017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.van Hoek B, de Boer J, Boudjema K, Williams R, Corsmit O, Terpstra OT. Auxiliary versus orthotopic liver transplantation for acute liver failure. EURALT Study Group. European Auxiliary Liver Transplant Registry. J Hepatol. 1999;30(4):699–705. doi: 10.1016/s0168-8278(99)80202-5. [DOI] [PubMed] [Google Scholar]
- 11.Azoulay D, Samuel D, Ichai P, et al. Auxiliary partial orthotopic versus standard orthotopic whole liver transplantation for acute liver failure: a reappraisal from a single center by a case-control study. Ann Surg. 2001;234(6):723–731. doi: 10.1097/00000658-200112000-00003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kamada N, Muller GH, Katami M, Tamaki T, Baguerizo A, Shinomiya T. Sensitization of rats for rejection of heart allografts by heterotopic auxiliary liver grafting or administration of liver cell suspensions. Transplantation. 1985;40(1):106–108. [PubMed] [Google Scholar]
- 13.Gugenheim J, Houssin D, Tamisier D, et al. Spontaneous long-term survival of liver allografts in inbred rats: influence of the hepatectomy of the recipient's own liver. Transplantation. 1981;32(5):445–450. [PubMed] [Google Scholar]
- 14.Matsuura T, Soejima Y, Taguchi T. Auxiliary partial orthotopic living donor liver transplantation with a small-for-size graft for congenital absence of the portal vein. Liver Trasnpl. 2010;16(12):1437–1439. doi: 10.1002/lt.22179. [DOI] [PubMed] [Google Scholar]
- 15.Ohno Y, Mita A, Ikegami T, et al. Temporary auxiliary partial orthotopic liver transplantation using a small graft for familial amyloid polyneuropathy. Am J Transplant. 2012;12(8):2211–2219. doi: 10.1111/j.1600-6143.2012.04061.x. [DOI] [PubMed] [Google Scholar]
- 16.Lee S, Edgington TS. Heterotopic liver transplantation utilizing inbred rat strains. I. Characterization of allogeneic graft rejection and the effects of biliary obstruction and portal vein circulation on liver regeneration. Am J Pathol. 1968;52(3):649–669. [PMC free article] [PubMed] [Google Scholar]
- 17.Hess F, Jerusalem C, van der Heyde MN. Advantages of auxiliary liver homotransplantation in rats. Arch Surg. 1972;104(1):76–80. doi: 10.1001/archsurg.1972.04180010070018. [DOI] [PubMed] [Google Scholar]
- 18.Yoo CH, Wang Y, Ha J, et al. Long-term fate of heterotopic liver transplants in rats with portal vein inflow only. Microsurgery. 1999;19(2):78–82. doi: 10.1002/(sici)1098-2752(1999)19:2<78::aid-micr7>3.0.co;2-w. [DOI] [PubMed] [Google Scholar]
- 19.Muller G. A simple technique for heterotopic auxiliary liver transplantation in the rat. Transplantation. 1983;36(2):221–222. doi: 10.1097/00007890-198308000-00028. [DOI] [PubMed] [Google Scholar]
- 20.Marni A, Ferrero ME. Heterotopic liver grafting in the rat. A simplified method using cuff techniques. Transplantation. 1985;39(3):329–331. doi: 10.1097/00007890-198503000-00029. [DOI] [PubMed] [Google Scholar]
- 21.Matsubara K, Yokota S, Ono Y, et al. Surgical Model of Auxiliary Partial Liver Transplantation in the Rat. [published online ahead of print December 22, 2015] Protoc exch [Google Scholar]
- 22.Laconi E, Oren R, Mukhopadhyay DK, et al. Long-term, near-total liver replacement by transplantation of isolated hepatocytes in rats treated with retrorsine. Am J Pathol. 1998;153(1):319–329. doi: 10.1016/S0002-9440(10)65574-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Petersen BE, Bowen WC, Patrene KD, et al. Bone marrow as a potential source of hepatic oval cells. Science. 1999;284(5417):1168–1170. doi: 10.1126/science.284.5417.1168. [DOI] [PubMed] [Google Scholar]
- 24.Lagasse E, Connors H, Al-Dhalimy M, et al. Purified hematopoietic stem cells can differentiate into hepatocytes in vivo. Nat Med. 2000;6(11):1229–1234. doi: 10.1038/81326. [DOI] [PubMed] [Google Scholar]
- 25.Theise ND, Badve S, Saxena R, et al. Derivation of hepatocytes from bone marrow cells in mice after radiation-induced myeloablation. Hepatology. 2000;31(1):235–240. doi: 10.1002/hep.510310135. [DOI] [PubMed] [Google Scholar]
- 26.Theise ND, Nimmakayalu M, Gardner R, et al. Liver from bone marrow in humans. Hepatology. 2000;32(1):11–16. doi: 10.1053/jhep.2000.9124. [DOI] [PubMed] [Google Scholar]
- 27.Gao Z-h, McAlister VC, Williams GM. Repopulation of liver endothelium by bone-marrow-derived cells. Lancet. 2001;357(9260):932–933. doi: 10.1016/s0140-6736(00)04217-3. [DOI] [PubMed] [Google Scholar]
- 28.Terada N, Hamazaki T, Oka M, et al. Bone marrow cells adopt the phenotype of other cells by spontaneous cell fusion. Nature. 2002;416(6880):542–545. doi: 10.1038/nature730. [DOI] [PubMed] [Google Scholar]
- 29.Wu T, Cieply K, Nalesnik MA, et al. Minimal evidence of transdifferentiation from recipient bone marrow to parenchymal cells in regenerating and long-surviving human allografts. Am J Transplant. 2003;3(9):1173–1181. doi: 10.1046/j.1600-6143.2003.00137.x. [DOI] [PubMed] [Google Scholar]
- 30.Murase N, Kim DG, Todo S, Cramer DV, Fung J, Starzl TE. FK506 suppression of heart and liver allograft rejection. II: The induction of graft acceptance in rats. Transplantation. 1990;50(5):739–744. doi: 10.1097/00007890-199011000-00002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Hossain MA, Hamamoto I, Kobayashi S, Maeba T, Maeta H. Immunosuppression in auxiliary partial liver transplantation with FK506 in rats. Transplant Proc. 1997;29(8):3617–3618. doi: 10.1016/s0041-1345(97)01045-2. [DOI] [PubMed] [Google Scholar]
- 32.Schleimer K, Stippel DL, Kasper HU, et al. Competition between native liver and graft in auxiliary liver transplantation in a rat model. Transplantation proceedings. 2008;40(4):967–970. doi: 10.1016/j.transproceed.2008.03.030. [DOI] [PubMed] [Google Scholar]
- 33.Schleimer K, Stippel DL, Kasper HU, et al. Portal vein arterialization increases hepatocellular apoptosis and inhibits liver regeneration. J Surg Res. 2008;149(2):250–258. doi: 10.1016/j.jss.2008.01.021. [DOI] [PubMed] [Google Scholar]
- 34.Michalopoulos GK. Liver regeneration. Journal of cellular physiology. 2007;213(2):286–300. doi: 10.1002/jcp.21172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Gerlach C, Sakkab DY, Scholzen T, Dassler R, Alison MR, Gerdes J. Ki-67 expression during rat liver regeneration after partial hepatectomy. Hepatology. 1997;26(3):573–578. doi: 10.1002/hep.510260307. [DOI] [PubMed] [Google Scholar]
- 36.Zou Y, Bao Q, Kumar S, Hu M, Wang GY, Dai G. Four waves of hepatocyte proliferation linked with three waves of hepatic fat accumulation during partial hepatectomy-induced liver regeneration. PloS one. 2012;7(2):e30675. doi: 10.1371/journal.pone.0030675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Starzl TE, Porter KA, Kashiwagi N. Portal hepatotrophic factors, diabetes mellitus and acute liver atrophy, hypertrophy and regeneration. Surg Gynecol Obstet. 1975;141(6):843–858. [PMC free article] [PubMed] [Google Scholar]
- 38.Starzl TE, Francavilla A, Porter KA, Benichou J, Jones AF. The effect of splanchnic viscera removal upon canine liver regeneration. Surg Gynecol Obstet. 1978;147(2):193–207. [PMC free article] [PubMed] [Google Scholar]
- 39.Fahrner R, Patsenker E, de Gottardi A, et al. Elevated liver regeneration in response to pharmacological reduction of elevated portal venous pressure by terlipressin after partial hepatectomy. Transplantation. 2014;97(9):892–900. doi: 10.1097/TP.0000000000000045. [DOI] [PubMed] [Google Scholar]
- 40.Pourcher G, El-Kehdy H, Kanso F, et al. Volumetric Portal Embolization: A New Concept to Improve Liver Regeneration and Hepatocyte Engraftment. Transplantation. 2016;100(2):344–354. doi: 10.1097/TP.0000000000001024. [DOI] [PubMed] [Google Scholar]
- 41.Uygun BE, Soto-Gutierrez A, Yagi H, et al. Organ reengineering through development of a transplantable recellularized liver graft using decellularized liver matrix. Nat Med. 2010;16(7):814–820. doi: 10.1038/nm.2170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Hata T, Uemoto S, Fujimoto Y, et al. Transplantation of engineered chimeric liver with autologous hepatocytes and xenobiotic scaffold. Ann Surg. 2013;257(3):542–547. doi: 10.1097/SLA.0b013e31825c5349. [DOI] [PubMed] [Google Scholar]
- 43.Wang L, Wang X, Wang L, et al. Hepatic vascular endothelial growth factor regulates recruitment of rat liver sinusoidal endothelial cell progenitor cells. Gastroenterology. 2012;143(6):1555–1563. e1552. doi: 10.1053/j.gastro.2012.08.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Wang L, Wang X, Xie G, Wang L, Hill CK, DeLeve LD. Liver sinusoidal endothelial cell progenitor cells promote liver regeneration in rats. J CLin Invest. 2012;122(4):1567–1573. doi: 10.1172/JCI58789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Okabayashi T, Cameron AM, Hisada M, Montgomery RA, Williams GM, Sun Z. Mobilization of host stem cells enables long-term liver transplant acceptance in a strongly rejecting rat strain combination. Am J Transplant. 2011;11(10):2046–2056. doi: 10.1111/j.1600-6143.2011.03698.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Michalopoulos GK, Barua L, Bowen WC. Transdifferentiation of rat hepatocytes into biliary cells after bile duct ligation and toxic biliary injury. Hepatology. 2005;41(3):535–544. doi: 10.1002/hep.20600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Yovchev MI, Locker J, Oertel M. Biliary fibrosis drives liver repopulation and phenotype transition of transplanted hepatocytes. J Hepatol. 2016 doi: 10.1016/j.jhep.2016.01.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Michalopoulos GK. Liver regeneration: alternative epithelial pathways. Int J Biochem Cell Biol. 2011;43(2):173–179. doi: 10.1016/j.biocel.2009.09.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Schulz C, Gomez Perdiguero E, Chorro L, et al. A lineage of myeloid cells independent of Myb and hematopoietic stem cells. Science. 2012;336(6077):86–90. doi: 10.1126/science.1219179. [DOI] [PubMed] [Google Scholar]
- 50.Hashimoto D, Chow A, Noizat C, et al. Tissue-resident macrophages self-maintain locally throughout adult life with minimal contribution from circulating monocytes. Immunity. 2013;38(4):792–804. doi: 10.1016/j.immuni.2013.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Durand F, Belghiti J, Vilgrain V, Valla D. Vanishing graft syndrome after auxiliary liver transplantation. J Hepatol. 2000;33(3):513. doi: 10.1016/s0168-8278(00)80291-3. [DOI] [PubMed] [Google Scholar]
- 52.de Jonge J, Zondervan PE, JN IJ, Metselaar HJ, Tilanus HW. Importance of portal flow diversion in experimental auxiliary partial orthotopic liver transplantation. Transplantation. 2000;70(1):44–47. [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.