

Abstract
Small aerial robots are limited to short mission times because aerodynamic and energy conversion efficiency diminish with scale. One way to extend mission times is to perch, as biological flyers do. Beyond perching, small robot flyers benefit from manoeuvring on surfaces for a diverse set of tasks, including exploration, inspection and collection of samples. These opportunities have prompted an interest in bimodal aerial and surface locomotion on both engineered and natural surfaces. To accomplish such novel robot behaviours, recent efforts have included advancing our understanding of the aerodynamics of surface approach and take-off, the contact dynamics of perching and attachment and making surface locomotion more efficient and robust. While current aerial robots show promise, flying animals, including insects, bats and birds, far surpass them in versatility, reliability and robustness. The maximal size of both perching animals and robots is limited by scaling laws for both adhesion and claw-based surface attachment. Biomechanists can use the current variety of specialized robots as inspiration for probing unknown aspects of bimodal animal locomotion. Similarly, the pitch-up landing manoeuvres and surface attachment techniques of animals can offer an evolutionary design guide for developing robots that perch on more diverse and complex surfaces.
Keywords: bioinspired, biomimetics, bimodal, flight, surface, locomotion
1. Introduction
In contrast to flying robots, animals can land on and take off from a diverse range of complex natural and engineered surfaces. Further, generalist birds, bats and many flying insects can fly for much longer than small aerial robots [1,2]. Regardless, flying animals and robots need to land frequently if they cannot acquire energy mid-flight. This requirement is especially important for small flying robots as compared to larger aircraft, because energy efficiencies associated with aerodynamics and on-board energy conversion greatly reduce with size. This helps explain why current aerial robots are typically limited to mission times of about half an hour [2].
The smaller the flyer, the lower its aerodynamic efficiency, which decreases with the Reynolds number (Re). For small animals and robots, 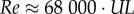 , in which U is the flight velocity, L is the chord length and 68 000 is calculated by taking the inverse of the kinematic viscosity of air at sea level [3]. The velocity of a flyer in equilibrium diminishes with size, because velocity is proportional to the square-root of weight divided by wing surface area,
, in which U is the flight velocity, L is the chord length and 68 000 is calculated by taking the inverse of the kinematic viscosity of air at sea level [3]. The velocity of a flyer in equilibrium diminishes with size, because velocity is proportional to the square-root of weight divided by wing surface area, 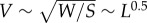 , the square-root of length [4]. Consequently, as flyers get smaller and are thus slower, they have to operate at lower Reynolds numbers. Low Reynolds numbers not only reduce the maximal lift coefficient, CL, they also increase the minimal drag coefficient, CD, because of complex flow separation phenomena associated with low Reynolds number effects [5,6]. These effects reduce the aerodynamic efficiency of small wings, in particular the glide ratio CL/CD, which determines flight distance, and the power factor
, the square-root of length [4]. Consequently, as flyers get smaller and are thus slower, they have to operate at lower Reynolds numbers. Low Reynolds numbers not only reduce the maximal lift coefficient, CL, they also increase the minimal drag coefficient, CD, because of complex flow separation phenomena associated with low Reynolds number effects [5,6]. These effects reduce the aerodynamic efficiency of small wings, in particular the glide ratio CL/CD, which determines flight distance, and the power factor 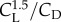 , which determines flight time [5,7]. For example, the lift to drag ratio of a well performing aerofoil of a wing operating at Re ∼ 106 (e.g. Liebeck L 1003) is about ten-times higher than that of a pigeon at Re ∼ 50 000 and 30 times that of a dragonfly wing at Re ∼ 5000 [8,9]. These scale effects reduce the aerodynamic efficiency of animals and aerial robots similarly.
, which determines flight time [5,7]. For example, the lift to drag ratio of a well performing aerofoil of a wing operating at Re ∼ 106 (e.g. Liebeck L 1003) is about ten-times higher than that of a pigeon at Re ∼ 50 000 and 30 times that of a dragonfly wing at Re ∼ 5000 [8,9]. These scale effects reduce the aerodynamic efficiency of animals and aerial robots similarly.
Energy storage and conversion also suffer at small scales. Small robots typically have to rely on electric motors powered by batteries, because scaling effects greatly reduce the efficiency of small gas engines and turbines [10]. The poor performance of fuel-based energy conversion stems from unstable combustion in small volumes, higher friction and heat loss, and sealing and manufacturing issues [10]. However, batteries have a much lower specific energy density (up to approx. 0.150 kWh kg−1 [11]) than large aircraft fuels (approx. 12 kWh kg−1 [12]) and the fat of animals (approx. 11 kWh kg−1 [13]). Even with energy conversion efficiencies of 90% for most electric motors [14], 20% for large engines [14] and 5–20% for animals [15,16], both large fuel-powered vehicles and animals have a much larger flight range and duration than electrically powered robots.
Current small flying robots are useful for inspection, imaging, surveillance and communication, provided these missions can be split-up in short cycles to accommodate the limited flight time. Furthermore, they need to fly sufficiently high to avoid collision with myriad obstacles in the near-ground environment, unless they can mitigate collisions [17]. The utility of aerial robots will be much improved, and their mission extended, if they can perch and locomote on these obstacles to collect physical samples and sense at lower energetic cost [18]. For similar reasons, animals locomote on the regular patterned surfaces of urban environments and the complex surfaces of the natural environment to exploit these habitats. In contrast, robots are missing both the autonomy and bimodal terrestrial and aerial locomotory apparatus needed to succeed in these conditions [18]. Whereas autonomy is an ultimate requirement, the first step is the development of a new class of bimodal robots to embody such navigational capabilities.
All aerial animals locomote by combining terrestrial, arboreal or aquatic locomotion with flight, because of the necessity to return to the Earth to feed, rest and reproduce. The diversity in effective biological solutions offers ‘out of the box’ design inspiration for roboticists [19–21]. The fundamental physical mechanism that facilitates these bioinspired solutions is evolution. Over millions of years, through the process of natural selection, animal ‘designs’ have been refined to increase their fitness, their ability to survive and reproduce in the face of an uncertain and complex environment [22]. A universal solution is animal locomotion, which facilitates resource retrieval, security and habitat location [23–25]. However, no animal morphology can be presumed to be optimal for locomotion in an engineering sense, because animals have additional constraints for survival and species preservation, including eating, digestion and mating [26]. Understanding the biomechanical basis of animal locomotion principles, sometimes referred to as ‘bio-understanding’ [27], is thus essential for harnessing future aerial robots with the extraordinary abilities of flying animals. Biomechanical trends in locomotion, body plan and surface attachment solutions emerge when function is compared across the diversity of organismal solutions. An integrated overview of the current state of bimodal aerial robotics and animal mechanics might thus provide a particularly valuable resource for new design inspiration [28–31].
Here, we review the present state of aerial bimodal robotics and our comparative biomechanical understanding of aerial animals that land and take off from solid surfaces. The key locomotion phases are: landing, surface attachment, terrestrial locomotion, surface detachment and take-off (figure 2). Each has specific, as well as interrelated, constraints. A given animal or robot may employ different solutions for each of them, tailored to the properties of the contact surfaces with which it interacts. The diverse biological solutions are organized based on how the flight dynamics, the surface grip mechanism and the physical surface properties scale. To understand how the surface contact requirements drive bimodal solutions, we first survey and characterize the diverse engineered and biological surfaces available for bimodal locomotion. We then review bimodal animal locomotion and existing aerial robotics realizations of landing and attachment, surface locomotion and take-off. Next these biological and engineering solutions are contrasted across scale to identify differences driven by scaling laws. Finally, we summarize the current research approaches in comparative biomechanics and bimodal robotics to determine opportunities for future crossover—to show how bio-understanding could transform the ability of robots to succeed at the interface of flight and surface locomotion.
Figure 2.

Biological solutions for landing, locomotion and take-off on highly irregular surfaces are diverse and depend on scale and flight adaptation. Powered flyers, shown in blue and teal colours, are able to use their wings to control their landing location with high precision while gliders, shown in purple and green colours, must land reliably at relatively higher velocities. Most animals pitch up to land on vertical surfaces, although bats are unusual in that they frequently invert to land upside-down. Larger animals predominantly use claws for attachment on vertical surfaces. Smaller animals typically use a combination of spines and adhesive pads. Frogs are unusual in that they typically land on leaves and stick to these smooth surfaces with wet adhesion. The flexibility of leaves dampens their impact. Flying snakes are also highly specialized and perch by wrapping their body around a landing branch. These animals all benefit from many joints that enable their bodies to adapt, comply and absorb energy upon landing. For take-off, most animals push-off from the surface with their legs or bodies. By contrast, bats usually hang upside-down and simply initiate their flight by dropping. (Animal drawings: Margarethe Roderick.)
2. Diversity of natural and engineered surfaces
The physical properties and topography of surfaces range from the hard, smooth, regularly patterned surfaces of glass or concrete to the soft, highly textured, irregular surfaces of trees (figure 1). These surface characteristics have a direct effect on suitable attachment strategies. As such, it is important to distinguish between macroscopic and microscopic surface architecture with respect to the length scale of the attachment mechanism. We describe the macroscopic shape through surface orientation or surface angle and the microscopic shape through rugosity of the surface features. Rugosity is related to fractal dimension [38,39] for irregular surfaces and is defined as Ar/Ag, where Ar is the real area of contact considering microscopic undulations, and Ag is the apparent or projected geometric surface area at the length scale of the region of attachment. For example, relatively smooth, low rugosity surfaces are conducive to using suction for attachment while textured, high rugosity surfaces are suited to claws. A related geometric parameter, La/Rs, depends on the magnitude of the local surface radius of curvature, Rs, compared to the characteristic length, La, of the attachment mechanism or appendage. For birds or insects grasping a small branch, La/Rs > 1, which allows an enveloping grasp. When landing on a smooth, flat wall, La/Rs → 0, which requires the use of some adhesive strategy such as dry or wet adhesion or suction. This difference, in turn, has implications for the landing strategy. An enveloping grasp on a small branch is tolerant of misalignment; moreover, the branch is likely to be compliant so that a broad range of incoming directions and velocities are possible. Conversely, when attaching to a hard, flat wall with adhesives, it is important to conform intimately to the wall surface to prevent local stress concentrations and adhesive failure. At the same time, flat walls often provide significant aerodynamic ‘ground effects’, which the biological or robotic flyer needs to adapt to, or exploit, to reduce velocity and align itself to the surface [40,41].
Figure 1.

An overview of the range of engineered and natural surfaces that flying animals and robots can encounter for landing, locomotion and take-off. The target surfaces include cables, building walls and windows, the metallic surfaces of cars, the bark of tree trunks and branches, animal skin and ground vegetation. Engineered surfaces typically have regular surface angles, high surface hardness and low roughness. Natural surfaces typically have irregular surface angles, relatively low surface hardness and high roughness. Accordingly, each surface is labelled with the surface name, range of typical surface orientations, approximate surface hardness using the Brinell hardness scale, and a classification of the approximate surface rugosity. The surface orientation determines the range of angles at which an animal or robot can approach and perch on a landing target. The Brinell hardness, HB, determines the ability of a claw to grip onto the surface by deforming it. Surface rugosity is a measure of the surface roughness that determines the ability of a claw to grip onto the surface without indentation. It is quantified by the ratio of the three-dimensional surface area to the two-dimensional projected surface area with respect to the characteristic (average) normal direction of the three-dimensional surface. Brinell hardness values from: [32–37]. (Left background photograph: courtesy of Monica Bond.)
Much research on perching aerial robots has focused on operating in towns and cities because there are many aerial robot applications in urban areas, such as inspection [18]. Human-made structures are typically designed for efficient cost-effective large-scale manufacturing and as a result, they are generally regular and uniformly patterned. Many of these surfaces are vertically oriented and hard, and a subset, including glass and metal surfaces, have a very low rugosity. Attachment of robots and animals to these surfaces requires close surface proximity using an adhesive pad or a suction seal (figure 3c,g,k,l,p) [42–50]. Other relatively hard and vertically oriented surfaces include brick, concrete and stucco with many asperities formed by holes and bumps, which facilitate the use of spines (figure 3b,f,j,o) [18,41,51–54]. In addition, urban areas feature many poles, pipes, posts and power lines. These cylindrical shapes have predictable surface properties that can be approached and grasped from a wide range of angles (figure 3a,e,i,n) [55–59].
Figure 3.

Existing robots that can land on, locomote around and take off from simple structured surfaces. The robots are organized by their aerial approach manoeuvre (vertical) and their surface attachment technique (horizontal). The aerial manoeuvres include pitch-up, direct ascent or descent, direct horizontal approach and inversion. Each manoeuvre has a specific required situational awareness, control and force sustained by the attachment mechanism. The different platforms are also organized by their ability to land from hovering: rotary robots in red, flapping robots in yellow, fixed wing robots in orange (other platforms in grey). A diverse set of approaches have been demonstrated to work for different structured surfaces, which include: cylinder grasping and hanging with La/Ls > 1, rough surface clinging using claws or spines with La/Ls ∼ 1, and smooth surface sticking using adhesives or suction with La/Ls < 1. Other specialized attachment techniques for specific surfaces include claws for soft surfaces, electrostatic adhesion, glue, as well as aerodynamic suction. The corresponding specific locomotion and take-off solutions depend on the aerial approach and attachment mechanism selected. These constraints limit the bimodal locomotion ability of these robots in specific ways, unlike the more generalist and flexible solutions found in nature. Models based on: [18,41,43,44,46–55,57–59,139–147].
Outside of cities, much of the Earth's land surface is covered by the irregular structures of the natural world. Some inanimate objects found in natural habitats, such as rocks, have physical properties that are similar to the rough outer surfaces of buildings but with more variable angles. However, many landing sites of interest also include living organisms, such as trees, ground vegetation and animals. These surfaces are typically softer than the surfaces of the engineered world and span all angles (figure 1). Exploiting the principles of claws, van der Waals forces and wet adhesion, animals have evolved to generate the required attachment forces (figure 2) [60–68]. This enables animals to negotiate and exploit complex surfaces with a combination of effective aerial approaches, contact strategies, surface locomotion techniques and take-off manoeuvres of which the dynamics are not well understood [1,69–85]. By contrast, aerial robots are just starting to implement some of these successful perching and locomotion strategies.
3. Air–surface transitions in flying animals
Animals have evolved a wide range of mechanisms and techniques for manoeuvring towards, landing on, moving on and taking off from irregular surfaces (figure 2). These differences are associated with animal size and aerodynamic control authority [1,86]. Unlike in robotics, where we often find separate mechanisms for flight and locomotion, the bimodal systems of organisms are more integrated. For example, the wings of powered fliers (birds, bats and insects) tuck by the side of the body so as not to inhibit surface locomotion. In some animals, these systems are even coupled. For example, some bats, squirrels and frogs use the same appendages for gliding as for surface attachment and locomotion [71,73–75,87]. To create flight trajectories, aerial animals rely on fusion of multiple sensory modalities with the visual system. Animals typically orient their body to be close to parallel with the surface just before touchdown to slow down and facilitate attachment. The attachment strategy of aerial animals involves a selection or combination of claws and adhesive pads, and their jointed appendages allow for adapting to surface features for locomotion. Many animals jump to take-off while others drop from their perches. Whereas the morphological adaptations of these systems in small animals with an exoskeleton, such as insects and spiders (Arthropoda), differ from larger animals with an internal skeleton, such as birds, bats, mammals, reptiles and amphibians (Chordata) [1,88], the flight control systems of all animals are well adapted for bimodal locomotion (figure 2).
Powered flyers, birds, bats and many insects, manoeuvre in the air by generating lift and thrust with their flapping wings (figure 2, blue and teal coloured animals) [23]. Without functional flapping wings, gliders must steer towards landing targets with their limbs or extended skin surfaces (figure 2, green and purple coloured animals) [89]. As a result, these animals typically approach a landing target at relatively higher speeds than powered flyers do. Gliders that use their limbs and body to direct their descent include many arthropods, such as some spiders and wingless hexapods [90–94]. Gliding vertebrates, including flying squirrels, colugos, snakes, lizards and frogs, use their extended aerodynamic surfaces to navigate in the air [71,74–79]. Flying squirrels and colugos employ their patagium, the skin connecting the feet to the hands, for manoeuvring [71,74,75]. Snakes, as well as lizards of the genus Draco, extend their ribs to achieve a higher glide ratio [76,79]. Geckos use their body and extra skin flaps for aerial control, and frogs rely heavily on their feet for aerial manoeuvres [77,89,95,96].
All of these animals depend on their eyes for situational awareness and navigation [69,70,82–84,97], which is enhanced by echolocation in many bats and some cave dwelling birds [98,99]. The majority of the flying animals rely thus on vision to land on a surface, and their visual feedback allows them to slow before perching. The widely used theory to describe this slowing in diurnal animals is called Tau Theory [97]. Tau is defined as 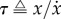 , where x and
, where x and 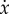 are the distance to landing and its derivative with respect to time; τ thus approximates the time to contact. Several birds, including pigeons and hummingbirds, have been shown to keep the time derivative of τ approximately constant during the approach [69,70]. Similarly, insects, including flies and bees, have been shown to use image expansion and optic flow on the retina to trigger deceleration, though with less precision [82–84].
are the distance to landing and its derivative with respect to time; τ thus approximates the time to contact. Several birds, including pigeons and hummingbirds, have been shown to keep the time derivative of τ approximately constant during the approach [69,70]. Similarly, insects, including flies and bees, have been shown to use image expansion and optic flow on the retina to trigger deceleration, though with less precision [82–84].
Upon landing, aerial animals must absorb the impact on the surface, which may vary in orientation and compliance (figure 2, contact). Some finches and doves, for example, have been shown to use their wingbeats to absorb 3–10 times the energy absorbed by the legs while landing [100]. Many of these animals pitch upwards to reduce their speed before landing on vertical surfaces, such as tree trunks [1,71,74,78–81,85]. This technique is found in a wide range of animals from insects to mammals, though it is more quantified in larger animals. Colugos, for example, are able to reduce their velocity by approximately to 60% (to 4 m s−1) using pitch-up manoeuvres [71,78]. Birds, flying squirrels and gliding lizards of the genus Draco have also been found to pitch up for rapid deceleration prior to landing [1,74,79–81]. By contrast, bees perform a pitch-up slow hover manoeuvre just before touchdown, though some small insects, such as flies, fly directly into the surface without substantial deceleration before contact [1,85].
Flying animals can also land (and sometimes must land) on the approximately cylindrical surfaces of branches, the angled tops of leaves, or inclined ceilings of caves. Branches and vegetation are popular landing sites for many gliders because they require less complex landing manoeuvres as the need to reduce speed is inversely proportional to surface compliance. For example, it is not known whether gliding snakes can land on vertical surfaces, but they have been observed landing on branches [76]. Some gliding rainforest frogs land on large leaves, which absorb some of the falling energy [87]. On the other hand, upside-down landings require special manoeuvres. To perch on branches or in caves, bats perform a head-under-heels manoeuvre, using their wings to change their moments of inertia to improve their flight control [73,101]. Similarly, some insects, including flies, ascend vertically when landing on overhung surfaces, and rotate the lower part of their body up upon impact [102].
For surface attachment, aerial arthropods and vertebrates generally use a selection or combination of adhesive pads or directional claws (or spines; figure 2, surface attachment) [86]. The underlying physics of these techniques include dry adhesion, wet adhesion or suction, interlocking, friction and grasping, [68,86]. The differences between and the effectiveness of all attachment strategies depend in large part on the rugosity. Adhesion relies on close contact with the surface for stable attachment, therefore, it is suitable for low rugosity surfaces [86]. Wet adhesion uses capillary forces for attachment, while dry adhesion relies on van der Waals attractive forces from hairs as a result of the high surface contact area [103–105]. Van der Waals adhesion has a higher theoretical interfacial strength [105]; however, wet adhesion can fill small irregularities with fluid [105], which is beneficial, because dry adhesion is less effective when a large number of small irregularities reduce the number of contact points. Many animals with adhesive pads use microscopic hairs that can adapt to irregularities [105]. At a larger scale, claws and spines allow for interlocking or grasping depending on relative claw and feature size. Interlocking occurs when the attachment mechanism engages mechanically with the surface, allowing for high shear forces [68,106]. This phenomenon can occur on high rugosity surfaces or on soft surfaces where the claws can produce local deformation. Grasping or wrapping allows for friction controllability via internal forces, which do not affect the external force balance [107]. This kind of attachment distributes stress over the grasped area rather than at concentrated points and can occur on high rugosity surfaces with curvatures La/Rs much greater than 1. Lastly, friction can occur whenever surfaces slide relative to each other.
The specific attachment techniques of animals differ depending on the lineage. Many aerial vertebrates, with a few exceptions, rely on hands and/or feet with claws for surface attachment. Furthermore, many perching birds and bats have specialized mechanisms in the feet to remain perched with low energy cost [60,61]. Smaller animals, including insects, geckos and frogs, typically use a combination of claws, spines or similar mechanisms along with adhesive pads. For example, claws have also been shown to be critical for navigating rough surfaces in beetles and geckos [62–64]. Similarly, frogs may use the specialized peg-like projections on their toes for interlocking on rough surfaces [68]. Adhesive pads are common among insects, geckos, skinks and tree frogs [65,66]. These pads can be categorized into smooth and hairy (fibrillar) pads as well as wet and dry adhesives [67]. Frogs, for example, use toe pads with mucous glands for wet adhesion [68]. The capillarity of the wet adhesion also introduces suction on smooth surfaces [68]. For many insects, both smooth and hairy pads excrete fluids for wet adhesion [108–111]. Other insects, spiders and geckos use hairy dry adhesive pads, which do not produce secretions [23,109,112–115]. The small hairs on foot pads are designed to self-clean, control detachment and increase adhesion [105]. Exceptions to these attachment trends include some bats and snakes. Certain bats, relatively large powered flyers, use wet adhesion to roost under leaves [61,116]. Snakes are unusual in that they use their flexible bodies to wrap around structures upon landing [76].
For movement on surfaces, aerial animals typically use their jointed appendages (legs, feet, arms, hands), with the exception of snakes (figure 2, surface locomotion) [23]. Surface navigation can be quasi-static or dynamic, and animals frequently adopt different gaits depending on the speed and irregularity of surfaces [23]. For example, flying squirrels can walk, bound and leap while moving on trees [117]. As another example, many birds navigate trees by hopping while foraging [118–122]. Many insects use an alternating tripod gait while climbing, with similar force patterns to those found in geckos [123]. While surface detachment has not been studied in great detail, many of these animals unload their directional attachment or peel their feet and hands from the surface to disengage [86]. For claws, animals open their hands to release the force holding the claws in contact with the surface. Some adhesive pads are also directional, which allows them to be detached by applying force in the non-preferred direction [124]. Other adhesive pads are typically controllable by concentrating stress for detachment [86].
When taking off, many gliding and powered flight animals use their appendages to significantly push-off from the substrate [71,72] (figure 2, take-off). This behaviour offsets the more energy demanding cost of flight and helps keep extended body parts from hitting surfaces. It helps gliders, in particular, because it enables them to reach flight speed more quickly. For example, colugos take off with a force equivalent to four-times body weight [71]. Finches and doves can produce forces in their legs that contribute at their peak to 77% and 85% of the whole-body accelerations, respectively [72]. Some animals capable of hovering, such as hummingbirds and swifts, rely more heavily on their wings rather than their legs for take-off or else drop from their perch [125]. Because bats roost upside-down, these animals are unusual in that they do not necessarily need to use additional leg forces to take off [101].
4. Air–surface transitions in aerial robots
While many robots can perform individual modes of locomotion reliably, transitions between modes remain a challenge. In the past few years, several robots have been developed that can perform aerial approaches, landings, surface locomotion and take-offs [18,41–59]. The simplest case of landing and locomotion is interacting with flat, horizontal ground. In these cases, robots have used wheels [126,127], cylindrical or spherical exoskeletons [17,128–130], or leg-like mechanisms [131–138] for locomotion. However, for the highly irregular or steep surfaces abundant on the Earth, new techniques have had to be developed. In landing on surfaces from a range of angles, current robots pitch up, approach directly, or invert themselves before landing. Under such conditions, aerial robots must exert substantial forces to remain attached or navigate along the surface. Surface attachment solutions include grasping, claws, adhesive pads and suction [18,41–59]. Some prototypes have begun to navigate surfaces with these techniques [41,44,139–141]. Take-off typically depends on the landing approach. So far, much research has focused on specific transitions or capabilities because for many applications of human interest only a subset of the touchdown to take-off process is required. Accordingly, there are two key foci: (i) aerial approach and take-off and (ii) surface contact and locomotion.
4.1. Landing and take-off in aerial robots
The landing and take-off manoeuvres of aerial robots on steep and inclined surfaces are highly interdependent. Landings of present aerial robots typically involve one of three manoeuvres: pitch-up, direct approach or inversion (figure 3) [18,41–59]. Each manoeuvre has specific trade-offs in required situational awareness, aerodynamic control and force on the attachment mechanism to enable landing and perching. Take-off has not been as widely studied and is generally designed based on the landing strategy.
Pitch-up manoeuvres require precise situational awareness and control but are typically associated with lower forces on the attachment mechanism (figure 3a–d). In similar ways as for animals, robot pitch-up manoeuvres reduce speed and can expose the landing gear to the surface for landing. However, this high angle of attack reduces control because translating wings will stall [57,148]. Thus, the timing of the pitch-up manoeuvre is critical [51,57]: too soon and the robot will fall, too late and it will crash into the surface. Pitch-up manoeuvres remain a challenge in part because current analytical aerodynamic techniques cannot predict the force fluctuations during these manoeuvres reliably [149,150]. Furthermore, standard solid wings offer less manoeuvrability than morphing wings, like those of animals [151–155]. As a result, many demonstrations of pitch-up manoeuvres with physical prototypes, both with fixed wings and quadrotors, have relied on motion capture systems for absolute situational awareness. These systems have been used for experiments to empirically generate the required control for trajectory planning (figure 3a) [57,156] and can give continuous position feedback (figure 3c) [142,143,157–162]. The few robots that have not required a motion capture system use sensors to detect a wall and initiate a pitch-up manoeuvre (figure 3b,c) [18,51,52,163]. To compensate for the lack of situational awareness using on-board sensing, some of the work in this field has been focused on how to recover from a failure [144,164]. Despite the strict control requirement, pitch-up manoeuvres offer design advantages for surface attachment. In a pitched up configuration, the robot can align its structure more closely to the surface, which minimizes the destabilizing pitch-back moment from the centre of mass with respect to the surface contact points (figure 3b,c) [18,43,51,52,142–144,159].
Many robots mitigate the challenges of dynamic pitch-up manoeuvres by using direct approaches, including horizontal, vertical or inverted approaches towards engineered surfaces. This reduces the required situational awareness and controllability of the robot, but results in a large pitch-back moment that needs to be addressed. The robots that perform these manoeuvres can typically hover or have a specialized suspension system to mitigate impact. Rotor-based vehicles are particularly well suited for vertical approaches, as they can vertically descend to land on cylinders (figure 3e) [55,58,165–167] or ascend to ceilings (figure 3f–h) [50,53,145,146]. In this configuration, robots must sustain a small pitch-back moment. By contrast, horizontal approach manoeuvres, for both fixed wings and rotor-based robots require the contact mechanism to hold while the robot is cantilevered, unless the robot aligns its structure near to the surface (figure 3i–m) [44,46–48,54,59,147]. Some quadrotors align themselves with the surface by pivoting nose-down (figure 3o–q) [41,49,53,140,141] or dropping below (figure 3n) [58] into an inverted configuration. The extra thrust from the rotors in the nose-down configuration can facilitate engagement of the attachment mechanism. However, this method does not work as effectively on non-vertical inclines and more complex surfaces.
In contrast to landing, take-off is rarely discussed in detail in the robotics literature. The take-off strategies of present aerial robots are mostly derived from the landing and contact strategy used. There are three main design considerations when taking off. Firstly, if the attachment is not directional, the robot must expend additional effort to disengage. Typically, disengagement involves reversing the attachment mechanism. Secondly, if the robot cannot hover, it needs to reach some speed by jumping or diving before flight can commence. Thirdly, robots must avoid hitting the surface too hard during take-off. Despite these constraints, present detachment mechanisms and rotor propulsion systems have proved to be quite successful for take-off from engineered surfaces. As a result, robots that pitch up to perch also pitch back to fly away (figure 3b) [18,51,52], while robots that make direct approaches also use direct take-offs (figure 3e–h) [47,54,139], and most aerial robots that perch in the inverted orientation pitch back into their stable flight configuration (figure 3o,p) [41].
4.2. Surface contact and locomotion in aerial robots
While landing and take-off depend primarily on the aerial platform, surface contact and locomotion mechanisms in bimodal aerial robots are generally highly specialized for the target surface type. These techniques include grasping for cylinders, claws and spines for rough surfaces, adhesive pads and suction of smooth surfaces, and additional miscellaneous strategies including electrostatic adhesion, aerodynamic suction and glue. Unlike in animals, there are few robots that have mechanisms to absorb the energy of impact other than with their structure [54,168]. None of these robots comes close to the bimodal locomotion capability of flying animals, which succeed on a more diverse suite of engineered and natural surfaces.
Enclosed grasping, or wrapping, has been a popular method used for attaching to relatively small cylindrical objects, such as cylinders, pipes and beams with La/Rs > 1 (figure 3e) [55,56]. These surfaces can be gripped from any direction, and allow for slight misalignment on contact. Many enclosed grasping mechanisms use jointed or compliant fingers [55,56,169–175], though others involve simply hooking and hanging (figure 3a,n) [58]. By contrast, one unusual modular snake-like robot wraps its entire body around cylinders upon contact (figure 3i) [59].
The most widely used attachment techniques for attaching to surfaces with macroscopic features (La/Rs ∼ 1), including stucco, concrete and wood, are claws and spines (figure 3b,f,j,o) [18,41,51–54]. Claws and spines can either use existing geometry or create local features by deforming the surface. Examples include claws that can catch on asperities and protrusions [18,41,51–53]. Using arrays of these microspines to share the load can improve the reliability of perching [18,41,51–53]. Spines have also been shown to enable soft surface penetration for passive, reliable attachment [54]. In both cases, these mechanisms can sustain high shear loads in combination with modest out of plane loads (normal to the surface).
Dry fibrillar adhesives and suction cups are frequently used for attachment on low rugosity surfaces, such as glass or polished metal (figure 3c,g,k,l,p) [43–50,176]. Fibrillar adhesives, inspired by the setae of gecko toes, are well suited to both porous and non-porous materials because of their reliance on van der Waals forces [176]. The molecular interaction arising from van der Waals forces supports large shear forces in addition to some normal adhesion [176]. These attributes make these adhesives appropriate for vertical and steeply inclined surfaces (figure 3c,k,l) [42–47]. Suction cups forming seals have been demonstrated to work for smooth, non-porous surfaces (figure 3g,p) [48–50] and require reliable vacuum pumps for continued attachment.
Additional techniques employed by aerial robots for maintaining proximity to a surface include electrostatic adhesion, aerodynamic suction, magnetism and glue. Electrostatic adhesion works well for a range of surfaces, but requires continuous power for attachment (figure 3h) [145]. Quadrotors that are aligned with the surface can also use their thrust to remain in contact, which also requires continuous power (figure 3q) [140]. This aerodynamic contact mechanism can also be used to regain contact upon slipping [41]. In addition, magnets offer reliable attachment, but only on magnetic surfaces [146,160,161]. Glue, such as rat trap glue, has been demonstrated as a reliable adhesive on a wide range of surfaces, but does not offer reliable detachment (figure 3m) [147].
While there is a wide range of robotic solutions for ascending specific surfaces, only a few have been demonstrated with aerial robots. For example, some climbing robots have rolled up walls using fibrillar adhesives [177,178] and pressure-sensitive adhesives [179] while others have used a stepping motion with spines [180]. Similarly, linkage-based and rolling locomotion have been the primary methods of movement for aerial robots on regular vertical and overhung surfaces. A central requirement for surface locomotion on steep surfaces is reliable attachment and detachment with each increment of motion. Repeated stepping robots with directional attachment mechanisms are specialized for climbing on flat surface types, ranging from high rugosity to smooth surfaces (figure 3d,l,o) [41,44,139]; rolling robots can use differential rotor thrust to navigate on wheels over a range of surface rugosities (figure 3q) [140,141].
5. Principles for bimodal robot design inspired by animal trends
5.1. Scaling implications for flying animals and perching robots
In comparing flying animals and robots, scaling trends emerge with implications for improving aerial design, structure and surface attachment strategies as a function of size. Specifically, aerodynamic control is closely associated with mass and wingspan combinations; larger structures must withstand larger resting stresses [23]. Accordingly, larger animals tend to rely more heavily on claws in contrast to insects, which rely on the combination of claws and an adhesive pad [86]. Figure 4 plots wingspan as a function of mass for different aerial systems (distinguished by colour) and attachment mechanism (distinguished by shape). A trend line emphasizes the utility of scaling to explain the cubic variation of mass with wingspan, as would be expected if all dimensions were increased proportionately. Whereas animals usually deviate from isometry within groups, trend lines that include multiple groups tend to follow isometric trends more closely [23]. Rotorcraft and gliding animals are typically heavier for their wingspan than fixed wing robots and active flying animals. Among these flyers, active flying animals and rotor-based robots can generally control their landing more precisely across a wide range of surfaces and orientations.
Figure 4.

Surface attachment solutions in animals and robots as a function of wingspan versus mass. Animal groups (open symbols) and aerial robots (filled symbols) are distinguished by colour, while attachment mechanism is distinguished by symbol shape. The trend line shows the utility of scaling to explain the cubic variation in mass with wingspan, which has profound implications for the structural and aerodynamic constraints on animals and robots. The plot also shows how the preferred attachment solution depends on scale accordingly. A great divide in attachment strategy exists between insects and vertebrates, which corresponds with the differences in skeletal structure. The reason adhesive pads did not evolve in larger animals is unclear at present, though this divide may stem from the relatively greater utility of claws for larger fliers. The robot data suggest that the customization of robots for specific applications, attachment strategies and surfaces has allowed them to be more variable in size and mass. The definition of wingspan is as follows: for flapping and fixed wing flyers it is the tip to tip distance, for rotorcraft it is the distance between the outer rotors, for lizards, snakes and amphibians it is the width between the outermost positions of their appendages. Data were listed or estimated from [4,74,77,79,95,181–185] and given by Jim McGuire for the Draco lizards. (Animal drawings: Margarethe Roderick.)
Scaling principles also put constraints on body structure [186]. Assuming isometric scaling, ‘tiny’ exoskeletons can withstand much larger loadings than ‘big’ animal skeletons or robot structures can when measured as multiples of body weight [23]. Thus, larger animals must land more carefully. To compensate, larger animals bend their joints less, which reduces the stress from the applied moments [23]. The requirement to adapt as a function of scale generally holds when comparing ‘tiny’ with ‘big’ scales. Certain flies can fly straight into a surface to secure adhesion [102], while birds must land with high precision to avoid injury (figure 4) [1]. This trend has also been found in robots with respect to their landing manoeuvres [187]. The smallest perching aerial robots require very little sensing and can rely on passive [188] or mechanically triggered mechanisms to safely interact with surfaces [187]. Larger robots typically use a complex control system (either a human pilot or a motion capture system) to avoid crashing.
Size and mass relationships have a large effect on surface attachment strategies and may explain why larger animals depend primarily on claws while small animals frequently use spines and adhesive pads. The overall division between insects and vertebrates is apparent in figure 4. Both dry and wet adhesion techniques scale with area, while mass scales with volume [86]. Therefore, larger animals must have disproportionally large pads or high efficiency per pad to attach. Indeed, larger species do typically have larger pads and/or higher adhesive pad efficiency, the largest being geckos [189]. For robots, similar scaling applies. However, by employing systems to distribute adhesive stress evenly, rotorcraft that perch with dry adhesives can exceed 0.5 kg [142]. As animals get larger, they rely increasingly on claws. The effectiveness of the claw depends on the claw shape (tip radius and structure diameter) and the strength of the claw material [86,106,190]. Smaller tip radii enable attachment to more surface features. Once interlocked, the claw is constrained only by its mechanical strength, determined by the shape and material of the claw [86]. For high rugosity surfaces, such as stone, stucco or concrete, the number of ‘usable’ asperities per unit area scales with the inverse of the tip radius [190]. Furthermore, stress varies with the square of the tip radius [190]. Therefore, for a given surface, isometrically larger animals will find fewer usable asperities, and their contact points will be more prone to failure, either from claw or surface fracture [86]. This constraint limits climbing for animals when  . Still, while adhesive pads of a certain size have an adhesion limit, big animals can still climb with large relatively blunt claws on sufficiently soft or irregular surfaces. For robots, similar trends again apply.
. Still, while adhesive pads of a certain size have an adhesion limit, big animals can still climb with large relatively blunt claws on sufficiently soft or irregular surfaces. For robots, similar trends again apply.
5.2. Future directions in the field of air–surface transitions and locomotion
Current research goals in bioinspired robotics and biomechanics have shaped our knowledge in these fields and offer paths for future crossover (figure 5a). Knowing that animals can perch almost without failure on a variety of surfaces, biomechanists in the past few decades have been searching for the principles that determine force production, energy expenditure and efficiency. On the other hand, roboticists can typically measure these quantities in robots. Therefore, roboticists have focused on realizing reliability, versatility and robustness within the constraints of current engineering materials, manufacturing, sensing and algorithms. Thus, these fields are poised to offer complementary insight. In the coming years, we expect to see more biomechanists use cutting-edge robots for controlled studies of specific phenomena, especially for understanding flow fields and surface attachment physics (figure 5b). We further expect roboticists to look to animals for design guidance in uncontrolled environments, in particular, for pitch-up manoeuvres and using claws and adhesive pads for surface attachment.
Figure 5.

Future directions for bimodal locomotion research. (a) Current comparative biomechanics focus and bioinspired robot design paradigms. In comparative biomechanics, researchers focus on relatively simple behaviours to measure the forces animals generate and how much energy this costs to determine the overall efficiency of the locomotion apparatus, for which they use engineering mechanics and instruments (indicated by the white arrow labelled ‘mechanics’). In bioinspired robotics, researchers can readily measure forces, energy, and efficiency and aim for improving reliability, robustness and versatility inspired by biological principles (indicated by the white arrow labelled ‘principles’). (b) Overview of particularly promising opportunities for advancing bioinspired robotics (blue) and bio-understanding based on comparative biomechanics (green). The importance of scale is underscored by how tiny versus big animals converged on different attachment solutions (figure 4), and must thus be considered explicitly in comparative biomechanical research and robotic implementation of bimodal locomotion principles. We note that robots can serve as informative platforms for dissecting the physical constraints that shaped these solutions and can be used to test evolutionary hypotheses.
Because of the ability in engineering to control variables, perform rapid repeatable experiments and distill physical phenomena, many opportunities have arisen for further studying the biomechanics of landing and surface locomotion (figure 5b). Large robots will continue to enable measuring and modelling flow fields around wings [148], claw engagement [106], contact forces during dynamic movements [123,191] and dynamic take-offs. Similarly, small robots can improve our understanding of the robustness, spine and adhesive attachment mechanisms, dynamic movements and flapping wing take-offs of tiny animals [192]. Despite these opportunities, studies in neuroscience and muscle physiology are needed to elucidate how sensorimotor control enables the musculoskeletal system of animals to outperform robot sensing, control and actuation.
While robots are specialized to be well suited for specific tasks, animal evolution offers inspiration to help engineers design robots for the uncertainty of the natural world. Continuous rotation actuators, electrical power storage, simple mechanisms, high-performance materials [193], and simple high-speed sensor suites have enabled aerial robots to perch on surface types on which many animals cannot land. Robotic solutions, although far less versatile, sophisticated, or robust than those of animals, can take advantage of more limited objectives and stronger materials so that very simple solutions work surprisingly well. As a consequence, bird-sized quadrotors can function with landing and perching strategies more like those seen in insects than in birds or mammals. On the other hand, animals have independently evolved certain common attributes as a result of adapting to uncertain conditions in nature. This evolutionary convergence in locomotive systems suggests that these strategies are well suited for reliable and versatile locomotion in these environments and can serve as a valuable design guide for engineers (figure 5b). In particular, for larger robots, there are opportunities in the study of closed loop pitch-up manoeuvres, claw mechanics, surface locomotion with airborne phases using jointed limbs, and jumping to move away from a surface to avoid wing–substrate collision during take-off. For small robots, we expect designers to begin to take advantage of the high specific strength with direct landing approaches as well as the spines and adhesive pads, jointed limbs for stepping and direct take-offs of insects.
6. Outlook
The fields of bimodal biomechanics and robotics are beginning to take off and could greatly benefit from a concerted effort. Animals use a wide range of landing and attachment techniques, though there are many commonalities among them distinguishable by size and body structure. Robotics can be an effective tool for dissecting the underlying physical constraints that have driven the evolution of bimodal animal locomotion, as well as mechanistically underpin complex behaviour. Similarly, modern perching aerial robots use a variety of aerial manoeuvres in combination with specific attachment methods that are optimized for specific engineered landing surfaces. Evolutionarily convergent commonalities among animals can offer key principles for designing versatile, reliable and robust aerial robots that can venture reliably into the outside world. These ventures can be enhanced by concurrent advances in engineering control systems, high-performance lightweight materials, mechanism design, and computer-based modelling and rapid prototyping [28]. This collaboration may yield unprecedented capabilities of a new generation of robots that not only embody the bimodal capacities of animals, but also harness the fruits of current efforts in machine learning and artificial intelligence.
Acknowledgements
We thank the members of the Lentink Lab and the Biomimetics and Dextrous Manipulation Lab, in particular, Hao Jiang, Hannah Stuart and Alice Wu, for their constructive feedback. We also thank two anonymous reviewers for their thoughtful input. We are grateful to Margarethe Roderick for the animal drawings, and Jim McGuire for the Draco lizard wingspan and mass data. We thank Morgan Pope for helpful discussions.
Authors' contributions
W.R.T.R. came up with the main ideas, drafted and revised the manuscript. D.L. and M.R.C. conceived of the overall focus of the work, contributed central ideas and revised drafts of the manuscript. All authors gave final approval for publication.
Competing interests
We have no competing interests.
Funding
This research was supported by a Stanford Graduate Engineering Fellowship and NSF Graduate Research Fellowship (DGE-114747) to W.R.T.R. A Micro Autonomous Systems and Technology at the Army Research Laboratory—Collaborative Technology Alliance Center grant MCE-16-17-4.4 and NSF Hybrid IIS_1161679 to M.R.C., and grant MCE-16-17-4.3 and NSF CAREER Award 1552419 to D.L. This material is based upon work supported by the National Science Foundation Graduate Research Fellowship Program.
Disclaimer
Any opinions, findings and conclusions or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the National Science Foundation.
References
- 1.Alexander DE, Vogel S. 2004. Nature‘s flyers: birds, insects, and the biomechanics of flight Baltimore, MD: JHU Press. [Google Scholar]
- 2.Crutsinger GM, Short J, Sollenberger R. 2016. The future of UAVs in ecology: an insider perspective from the Silicon Valley drone industry. J. Unmanned Veh. Syst. 4, 161–168. ( 10.1139/juvs-2016-0008) [DOI] [Google Scholar]
- 3.Bertin JJ, Cummings RM. 2014. Aerodynamics for engineers, 6th edn New York City, NY: Pearson. [Google Scholar]
- 4.Tennekes H. 1997. The simple science of flight from insects to jumbo jets. Cambridge, MA: MIT Press. [Google Scholar]
- 5.Leishman GJ. 2007. Principles of helicopter aerodynamics. Cambridge, UK: Cambridge University Press. [Google Scholar]
- 6.Jacobs EN, Sherman A. 1937. Airfoil section characteristics as affected by variations of the Reynolds number. NACA Rep. 586.
- 7.Drela M. 2014. Flight vehicle aerodynamics. Cambridge, MA: MIT Press. [Google Scholar]
- 8.Lissaman PBS. 1983. Low-Reynolds-number airfoils. Annu. Rev. Fluid Mech. 15, 223–239. ( 10.1146/annurev.fl.15.010183.001255) [DOI] [Google Scholar]
- 9.Shyy W, Lian Y, Tang J, Viieru D, Liu H. 2007. Aerodynamics of Low Reynolds number flyers. Cambridge, UK: Cambridge University Press. [Google Scholar]
- 10.Sher I, Levinzon-Sher D, Sher E. 2009. Miniaturization limitations of HCCI internal combustion engines. Appl. Therm. Eng. 29, 400–411. ( 10.1016/j.applthermaleng.2008.03.020) [DOI] [Google Scholar]
- 11.Scrosati B, Garche J. 2010. Lithium batteries: status, prospects and future. J. Power Sources 195, 2419–2430. ( 10.1016/j.jpowsour.2009.11.048) [DOI] [Google Scholar]
- 12.Hileman JI, Donohoo PE, Stratton RW. 2010. Energy content and alternative jet fuel viability. J. Propuls. Power 26, 1184–1196. ( 10.2514/1.46232) [DOI] [Google Scholar]
- 13.Livesey G, Elia M. 1988. Estimation of energy expenditure, and net fat oxidation calorimetry: evaluation of errors to the detailed composition of net carbohydrate and synthesis by indirect with special reference. Clin. Nutr. 47, 608–628. [DOI] [PubMed] [Google Scholar]
- 14.Larminie J, Lowry J. 2012. Electric vehicle technology explained, 2nd edn Chichester, UK: John Wiley & Sons, Ltd. [Google Scholar]
- 15.Kvist A, Lindström Å, Green M, Piersma T, Visser GH. 2001. Carrying large fuel loads during sustained bird flight is cheaper than expected. Nature 413, 730–732. ( 10.1038/35099556) [DOI] [PubMed] [Google Scholar]
- 16.Lehmann F-O, Dickinson MH. 1997. The changes in power requirements and muscle efficiency during elevated force production in the fruit fly Drosophila melanogaster. J. Exp. Biol. 200, 1133–1143. [DOI] [PubMed] [Google Scholar]
- 17.Briod A, Kornatowski P, Zufferey J-C, Floreano D. 2014. A collision-resilient flying robot. J. Field Robot. 31, 496–509. ( 10.1002/rob.21495) [DOI] [Google Scholar]
- 18.Desbiens AL, Cutkosky MR. 2010. Landing and perching on vertical surfaces with microspines for small unmanned air vehicles. J. Intell. Robot. Syst. 57, 313–327. ( 10.1007/s10846-009-9377-z) [DOI] [Google Scholar]
- 19.Lentink D, Biewener AA. 2010. Nature-inspired flight—beyond the leap. Bioinspir. Biomim. 5, 40201 ( 10.1088/1748-3182/5/4/040201) [DOI] [PubMed] [Google Scholar]
- 20.Lentink D. 2013. Biomimetics: flying like a fly. Nature 498, 306–307. ( 10.1038/nature12258) [DOI] [PubMed] [Google Scholar]
- 21.Lentink D. 2014. Bioinspired flight control. Bioinspir. Biomim. 9, 20301 ( 10.1088/1748-3182/9/2/020301) [DOI] [PubMed] [Google Scholar]
- 22.Stearns S, Hoekstra R. 2005. Evolution, 2nd edn Oxford, UK: Oxford University Press. [Google Scholar]
- 23.Alexander RM. 2006. Principles of animal locomotion. Princeton, NJ: Princeton University Press. [Google Scholar]
- 24.Vogel S. 1994. Life in moving fluids: the physical biology of flow. Princeton, NJ: Princeton University Press. [Google Scholar]
- 25.Alexander RM. 1999. Energy for animal life. Oxford, UK: Oxford University Press. [Google Scholar]
- 26.Alexander RM. 1996. Optima for animals. Princeton, NJ: Princeton University Press. [Google Scholar]
- 27.Cutkosky MR. 2015. Climbing with adhesion: from bioinspiration to biounderstanding. Interface Focus 5, 20150015 ( 10.1098/rsfs.2015.0015) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Sitti M, Menciassi A, Ijspeert AJ, Low KH, Kim S. 2013. Survey and introduction to the focused section on bio-inspired mechatronics. IEEE/ASME Trans. Mechatron. 18, 409–418. ( 10.1109/TMECH.2012.2233492) [DOI] [Google Scholar]
- 29.Low KH, Hu T, Mohammed S, Tangorra J, Kovac M. 2015. Perspectives on biologically inspired hybrid and multi-modal locomotion. Bioinspir. Biomim. 10, 20301 ( 10.1088/1748-3190/10/2/020301) [DOI] [PubMed] [Google Scholar]
- 30.Dudley R, Byrnes G, Yanoviak SP, Borrell B, Brown RM, McGuire JA. 2007. Gliding and the functional origins of flight: biomechanical novelty or necessity. Annu. Rev. Ecol. Evol. Syst. 38, 179–201. ( 10.1146/annurev.ecolsys.37.091305.110014) [DOI] [Google Scholar]
- 31.Lock RJ, Burgess SC, Vaidyanathan R. 2014. Multi-modal locomotion: from animal to application. Bioinspir. Biomim. 9, 11001 ( 10.1088/1748-3182/9/1/011001) [DOI] [PubMed] [Google Scholar]
- 32.Doyle J. 1980. The hardness of wood. PhD thesis, University of Canterbury, Christchurch, New Zealand.
- 33.Quinn GD, Green P, Xu K. 2003. Cracking and the indentation size effect for knoop hardness of glasses. J. Am. Ceram. Soc. 86, 441–448. ( 10.1111/j.1151-2916.2003.tb03319.x) [DOI] [Google Scholar]
- 34.Ainbinder SB, Laka MG. 1969. Hardness of polymers. Polym. Mech. 2, 211–217. ( 10.1007/BF00860285) [DOI] [Google Scholar]
- 35.ASTM. 2002. Standard hardness conversion tables for metals. E 140-02, 1-21. West Conshohocken, PA: ASTM International. [Google Scholar]
- 36.Hays C, Kendall EG. 1973. An analysis of Knoop microhardness. Metallography 6, 275–282. ( 10.1016/0026-0800(73)90053-0) [DOI] [Google Scholar]
- 37.Beke B. 2012. The process of fine grinding, vol. 1 Berlin, Germany: Springer Science & Business Media. [Google Scholar]
- 38.Ganti S, Bhushan B. 1995. Generalized fractal analysis and its applications to engineering surfaces. Wear 180, 17–34. ( 10.1016/0043-1648(94)06545-4) [DOI] [Google Scholar]
- 39.Brown SR. 1988. A note on the description of surface roughness using fractal dimension. Geophys. Res. Lett. 15, 286 ( 10.1029/GL015i003p00286) [DOI] [Google Scholar]
- 40.Rayner JMV. 1991. On the aerodynamics of animal flight in ground effect. Phil. Trans. R. Soc. Lond. B 334, 119–128. ( 10.1098/rstb.1991.0101) [DOI] [Google Scholar]
- 41.Pope MT, Cutkosky MR. 2016. Thrust-assisted perching and climbing for a bioinspired UAV. In Biomimetic and biohybrid systems (eds Lepora NF, Mura A, Krapp HG, PVerschure FMJ, Prescott TJ), pp. 288–296. Berlin, Germany: Springer. [Google Scholar]
- 42.Hawkes EW, Eason EV, Asbeck AT, Cutkosky MR. 2013. The Gecko's toe: scaling directional adhesives for climbing applications. IEEE/ASME Trans. Mechatron. 18, 518–526. ( 10.1109/TMECH.2012.2209672) [DOI] [Google Scholar]
- 43.Hawkes EW, Jiang H, Cutkosky MR. 2016. Three-dimensional dynamic surface grasping with dry adhesion. Int. J. Rob. Res. 35, 943–958. ( 10.1177/0278364915584645) [DOI] [Google Scholar]
- 44.Estrada MA, Hawkes EW, Christensen DL, Cutkosky MR. 2014. Perching and vertical climbing: design of a multimodal robot. In Proc. IEEE Int. Conf. on Robotics and Automation, 31 May–7 June, Hong Kong, China, pp. 4215–4221. Piscataway, NJ: IEEE.
- 45.Jiang H, Pope MT, Hawkes EW, Christensen DL, Estrada MA, Parlier A, Tran R, Cutkosky MR. 2014. Modeling the dynamics of perching with opposed-grip mechanisms. In IEEE Int. Conf. on Robotics and Automation (ICRA), 31 May–7 June, Hong Kong, China, pp. 3102–3108. Piscataway, NJ: IEEE.
- 46.Daler L, Klaptocz A, Briod A, Sitti M, Floreano D. 2013. A perching mechanism for flying robots using a fibre-based adhesive. In Proc. IEEE Int. Conf. on Robotics and Automation, 6–10 May, Karlsruhe, Germany, pp. 4433–4438. Piscataway, NJ: IEEE.
- 47.Kalantari A, Mahajan K, Donald I, Spenko M. 2015. Autonomous perching and take-off on vertical walls for a quadrotor micro air vehicle. In IEEE Int. Conf. on Robotics and Automation (ICRA), 26–30 May, Seattle, WA, pp. 4669–4674. Piscataway, NJ: IEEE. Piscataway, NJ: IEEE.
- 48.Liu Y, Sun G, Chen H. 2014. Impedance control of a bio-inspired flying and adhesion robot. In IEEE Int. Conf. on Robotics and Automation (ICRA), 31 May–7 June, Hong Kong, China, pp. 3564–3569. Piscataway, NJ: IEEE.
- 49.Tsukagoshi H, Watanabe M, Hamada T, Ashlih D, Iizuka R. 2015. Aerial manipulator with perching and door-opening capability. In IEEE Int. Conf. on Robotics and Automation (ICRA), 26–30 May, Seattle, WA, pp. 4663–4668. Piscataway, NJ: IEEE.
- 50.Du S, Chen H, Liu Y, Hu R. 2015. Unified switching between active flying and perching of a bioinspired robot using impedance control. J. Robot. 2015, 1–11. ( 10.1155/2015/763710) [DOI] [Google Scholar]
- 51.Desbiens AL, Asbeck AT, Cutkosky MR. 2011. Landing, perching and taking off from vertical surfaces. Int. J. Rob. Res. 30, 355–370. ( 10.1177/0278364910393286) [DOI] [Google Scholar]
- 52.Desbiens AL, Asbeck A, Cutkosky M. 2010. Hybrid aerial and scansorial robotics. In Proc. IEEE Int. Conf. on Robotics and Automation, 3–8 May, Anchorage, Alaska, pp. 72–77. Piscataway, NJ: IEEE.
- 53.Pope M. 2016. Microspines make it easy for drones to perch on walls and ceilings. In IEEE Spectrum. See http://spectrum.ieee.org/automaton/robotics/drones/microspines-make-it-easy-for-drones-to-perch-on-walls-and-ceilings. Piscataway, NJ: IEEE.
- 54.Kovač M, Germann J, Hürzeler C, Siegwart RY, Floreano D. 2009. A perching mechanism for micro aerial vehicles. J. Micro-Nano Mechatron. 5, 77–91. ( 10.1007/s12213-010-0026-1) [DOI] [Google Scholar]
- 55.Chi W, Low KH, Hoon KH, Tang J. 2014. An optimized perching mechanism for autonomous perching with a quadrotor. In Proc. IEEE Int. Conf. on Robotics and Automation, 31 May–7 June, Hong Kong, China, pp. 3109–3115. Piscataway, NJ: IEEE.
- 56.Coxworth B. 2016. Grasping drone brings new meaning to ‘armed aircraft’. New Atlas, 9 September 2016. See http://newatlas.com/prodrone-armed-drone/45351/.
- 57.Cory R, Tedrake R. 2008. Experiments in fixed-wing UAV perching. In AIAA Guidance, Navigation and Control Conf. and Exhibit, 18–21 August, Honolulu, HI, pp. 1–12. Reston, VA: AIAA.
- 58.Braithwaite A, Alhinai T, Haas-heger M, Mcfarlane E, Kovac M. 2015. Tensile web construction and perching with nano aerial vehicles. In Int. Symp. on Robotics Research, pp. 1–17.
- 59.Ackerman E. 2013. CMU snake robots can now strangle things on contact. In IEEE Spectrum. See http://spectrum.ieee.org/automaton/robotics/military-robots/cmu-snake-robots-can-now-strangle-things-on-contact. Piscataway, NJ: IEEE.
- 60.Galton PM, Shepherd JD. 2012. Experimental analysis of perching in the European starling (Sturnus vulgaris: Passeriformes; Passeres), and the automatic perching mechanism of birds. J. Exp. Zool. Part A Ecol. Genet. Physiol. 317 A, 205–215. ( 10.1002/jez.1714) [DOI] [PubMed] [Google Scholar]
- 61.Quinn TH, Baumel JJ. 1993. Chiropteran tendon locking mechanism. J. Morphol. 216, 197–208. ( 10.1002/jmor.1052160207) [DOI] [PubMed] [Google Scholar]
- 62.Stork NE. 1980. Experimental analysis of adhesion of Chrysolina polita (Chrysomelidae: Coleoptera) on a variety of surfaces. J. Exp. Biol. 88, 91–107. [Google Scholar]
- 63.Betz O. 2002. Performance and adaptive value of tarsal morphology in rove beetles of the genus Stenus (Coleoptera, Staphylinidae). J. Exp. Biol. 205, 1097–1113. [DOI] [PubMed] [Google Scholar]
- 64.Mahendra BC. 1941. Contributions to the bionomics, anatomy reproduction and development of the Indian house gecko, Hemidactylus faviviridis Rüppel. In Proc. of the Indian Academy of Sciences - Section B, pp. 288–306.
- 65.Creton C, Gorb SN. 2007. Sticky feet: from animals to materials. MRS Bull. 32, 466–472. ( 10.1557/mrs2007.79) [DOI] [Google Scholar]
- 66.Arzt E, Gorb S, Spolenak R. 2003. From micro to nano contacts in biological attachment devices. Proc. Natl Acad. Sci. USA 100, 10 603–10 609. ( 10.1073/pnas.1534701100) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Beutel RG, Gorb SN. 2001. Ultrastructure of attachment specializations of hexapods (Arthropoda): evolutionary patterns inferred from a revised ordinal phylogeny. J. Zool. Syst. Evol. Res. 39, 177–207. ( 10.1046/j.1439-0469.2001.00155.x) [DOI] [Google Scholar]
- 68.Emerson SB, Diehl D. 1980. Toe pad morphology and mechanisms of sticking in frogs. Biol. J. Linn. Soc. 13, 199–216. ( 10.1111/j.1095-8312.1980.tb00082.x) [DOI] [Google Scholar]
- 69.Lee DN, Davies MNO, Green PR, Van Der Weel FR. 1993. Visual control of velocity of approach by pigeons when landing. J. Exp. Biol. 180, 85–104. ( 10.4324/9780203936672) [DOI] [Google Scholar]
- 70.Lee DN, Reddish PE, Rand DT. 1991. Aerial docking by hummingbirds. Naturwissenschaften 78, 526–527. ( 10.1007/BF01131406) [DOI] [Google Scholar]
- 71.Byrnes G, Lim NT-L, Spence AJ. 2008. Take-off and landing kinetics of a free-ranging gliding mammal, the Malayan colugo (Galeopterus variegatus). Proc. R. Soc. B 275, 1007–1013. ( 10.1098/rspb.2007.1684) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Provini P, Tobalske BW, Crandell KE, Abourachid A. 2012. Transition from leg to wing forces during take-off in birds. J. Exp. Biol. 215, 4115–4124. ( 10.1242/jeb.074484) [DOI] [PubMed] [Google Scholar]
- 73.Riskin DK, Bahlman JW, Hubel TY, Ratcliffe JM, Kunz TH, Swartz SM. 2009. Bats go head-under-heels: the biomechanics of landing on a ceiling. J. Exp. Biol. 212, 945–953. ( 10.1242/jeb.026161) [DOI] [PubMed] [Google Scholar]
- 74.Paskins KE, Bowyer A, Megill WM, Scheibe JS. 2007. Take-off and landing forces and the evolution of controlled gliding in northern flying squirrels Glaucomys sabrinus. J. Exp. Biol. 210, 1413–1423. ( 10.1242/jeb.02747) [DOI] [PubMed] [Google Scholar]
- 75.Bahlman JW, Swartz SM, Riskin DK, Breuer KS. 2013. Glide performance and aerodynamics of non-equilibrium glides in northern flying squirrels (Glaucomys sabrinus). J. R. Soc. Interface 10, 20120794 ( 10.1098/rsif.2012.0794) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Socha JJ. 2011. Gliding flight in Chrysopelea: turning a snake into a wing. Integr. Comp. Biol. 51, 969–982. ( 10.1093/icb/icr092) [DOI] [PubMed] [Google Scholar]
- 77.McCay MG. 2001. Aerodynamic stability and maneuverability of the gliding frog Polypedates dennysi. J. Exp. Biol. 204, 2817–2826. [DOI] [PubMed] [Google Scholar]
- 78.Byrnes G, Spence AJ. 2011. Ecological and biomechanical insights into the evolution of gliding in mammals. Integr. Comp. Biol. 51, 991–1002. ( 10.1093/icb/icr069) [DOI] [PubMed] [Google Scholar]
- 79.McGuire JA, Dudley R. 2005. The cost of living large: comparative gliding performance in flying lizards (Agamidae: Draco). Am. Nat. 166, 93–106. ( 10.1086/430725) [DOI] [PubMed] [Google Scholar]
- 80.Ando M, Shiraishi S. 1993. Gliding flight in the Japanese giant flying squirrel Petaurista leucogenys. J. Mammal. Soc. Jpn 18, 19–32. [Google Scholar]
- 81.Stafford BJ, Thorington RW, Kawamichi T. 2002. Gliding behavior of Japanese giant flying squirrels (Petaurista leucogenys). J. Mammal. 83, 553–562. () [DOI] [Google Scholar]
- 82.Wagner H. 1982. Flow-field variables trigger landing in flies. Nature 297, 147–148. ( 10.1038/297147a0) [DOI] [Google Scholar]
- 83.Srinivasan MV, Zhang S-W, Chahl JS. 2001. Landing strategies in honeybees, and possible applications to autonomous airborne vehicles. Biol. Bull. 200, 216–221. ( 10.2307/1543319) [DOI] [PubMed] [Google Scholar]
- 84.Baird E, Boeddeker N, Ibbotson MR, Srinivasan MV. 2013. A universal strategy for visually guided landing. Proc. Natl Acad. Sci. USA 110, 18 686–18 691. ( 10.1073/pnas.1314311110) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Evangelista C, Kraft P, Dacke M, Reinhard J, Srinivasan MV. 2010. The moment before touchdown: landing manoeuvres of the honeybee Apis mellifera. J. Exp. Biol. 213, 262–270. ( 10.1242/jeb.037465) [DOI] [PubMed] [Google Scholar]
- 86.Labonte D, Federle W. 2015. Scaling and biomechanics of surface attachment in climbing animals. Phil. Trans. R. Soc. B 370, 20140027 ( 10.1098/rstb.2014.0027) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Roberts WE. 1994. Explosive breeding aggregations and parachuting in a neotropical frog, Agalychnis saltator (Hylidae). J. Herpetol. 28, 193 ( 10.2307/1564620) [DOI] [Google Scholar]
- 88.Dickinson MH. 2000. How animals move: an integrative view. Science 288, 100–106. ( 10.1126/science.288.5463.100) [DOI] [PubMed] [Google Scholar]
- 89.Jusufi A, Goldman DI, Revzen S, Full RJ. 2008. Active tails enhance arboreal acrobatics in geckos. Proc. Natl Acad. Sci. USA 105, 4215–4219. ( 10.1073/pnas.0711944105) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Yanoviak SP, Munk Y, Kaspari M, Dudley R. 2010. Aerial manoeuvrability in wingless gliding ants (Cephalotes atratus). Proc. R. Soc. B 277, 2199–2204. ( 10.1098/rspb.2010.0170) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Yanoviak SP, Munk Y, Dudley R. 2015. Arachnid aloft: directed aerial descent in neotropical canopy spiders. J. R. Soc. Interface 12, 20150534 ( 10.1098/rsif.2015.0534) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Munk Y, Yanoviak SP, Koehl MAR, Dudley R. 2015. The descent of ant: field-measured performance of gliding ants. J. Exp. Biol. 218, 1393–1401. ( 10.1242/jeb.106914) [DOI] [PubMed] [Google Scholar]
- 93.Yanoviak SP, Kaspari M, Dudley R. 2009. Gliding hexapods and the origins of insect aerial behaviour. Biol. Lett. 5, 510–512. ( 10.1098/rsbl.2009.0029) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Yanoviak SP, Dudley R, Kaspari M. 2005. Directed aerial descent in canopy ants. Nature 433, 624–626. ( 10.1038/nature03268) [DOI] [PubMed] [Google Scholar]
- 95.Young BA, Lee CE, Daley KM. 2002. On a flap and a foot: aerial locomotion in the ‘flying’ gecko, Ptychozoon kuhli. J. Herpetol. 36, 412–418. ( 10.1670/0022-1511(2002)036%5B0412:OAFAAF%5D2.0.CO;2) [DOI] [Google Scholar]
- 96.Mercellini DL, Keefer TE. 1976. Analysis of the gliding behavior of Ptychozoon lionatum (Reptilia: Gekkonidae). Herpetologica 32, 362–366. [Google Scholar]
- 97.Lee DN. 2009. General Tau Theory: evolution to date. Perception 38, 837–858. ( 10.1068/ldmk-lee) [DOI] [PubMed] [Google Scholar]
- 98.Jones G, Holderied MW. 2007. Bat echolocation calls: adaptation and convergent evolution. Proc. R. Soc. B 274, 905–912. ( 10.1098/rspb.2006.0200) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Fullard JH, Barclay RMR, Thomas DW. 1993. Echolocation in free-flying Atiu swiftlets (Aerodramus sawtelli). Biotropica 25, 334 ( 10.2307/2388791) [DOI] [Google Scholar]
- 100.Provini P, Tobalske BW, Crandell KE, Abourachid A. 2014. Transition from wing to leg forces during landing in birds. J. Exp. Biol. 217, 2659–2666. ( 10.1242/jeb.104588) [DOI] [PubMed] [Google Scholar]
- 101.Bergou AJ, Swartz SM, Vejdani H, Riskin DK, Reimnitz L, Taubin G, Breuer KS. 2015. Falling with style: bats perform complex aerial rotations by adjusting wing inertia. PLoS Biol. 13, 1–16. ( 10.1371/journal.pbio.1002297) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Hyzer WG. 1962. Flight behavior of a fly alighting on a ceiling. Science 137, 609–610. ( 10.1126/science.137.3530.609) [DOI] [PubMed] [Google Scholar]
- 103.Gao H, Wang X, Yao H, Gorb S, Arzt E. 2005. Mechanics of hierarchical adhesion structures of geckos. Mech. Mater. 37, 275–285. ( 10.1016/j.mechmat.2004.03.008) [DOI] [Google Scholar]
- 104.Autumn K, et al. 2002. Evidence for van der Waals adhesion in gecko setae. Proc. Natl Acad. Sci. USA 99, 12 252–12 256. ( 10.1073/pnas.192252799) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Federle W. 2006. Why are so many adhesive pads hairy? J. Exp. Biol. 209, 2611–2621. ( 10.1242/jeb.02323) [DOI] [PubMed] [Google Scholar]
- 106.Dai Z, Gorb SN, Schwarz U. 2002. Roughness-dependent friction force of the tarsal claw system in the beetle Pachnoda marginata (Coleoptera, Scarabaeidae). J. Exp. Biol. 205, 2479–2488. [DOI] [PubMed] [Google Scholar]
- 107.Prattichizzo D, Trinkle JC. 2008. Grasping. In Springer handbook of robotics (eds Siciliano B, Khatib O), pp. 955–988. Berlin, Germany: Springer Science & Business Media. [Google Scholar]
- 108.Frantsevich L, Gorb S. 2004. Structure and mechanics of the tarsal chain in the hornet, Vespa crabro (Hymenoptera: Vespidae): implications on the attachment mechanism. Arthropod. Struct. Dev. 33, 77–89. ( 10.1016/j.asd.2003.10.003) [DOI] [PubMed] [Google Scholar]
- 109.Bullock JMR, Drechsler P, Federle W. 2008. Comparison of smooth and hairy attachment pads in insects: friction, adhesion and mechanisms for direction-dependence. J. Exp. Biol. 211, 3333–3343. ( 10.1242/jeb.020941) [DOI] [PubMed] [Google Scholar]
- 110.Federle W. 2002. An integrative study of insect adhesion: mechanics and wet adhesion of pretarsal pads in ants. Integr. Comp. Biol. 42, 1100–1106. ( 10.1093/icb/42.6.1100) [DOI] [PubMed] [Google Scholar]
- 111.Dirks J-H, Clemente CJ, Federle W. 2010. Insect tricks: two-phasic foot pad secretion prevents slipping. J. R. Soc. Interface 7, 587–593. ( 10.1098/rsif.2009.0308) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Betz O. 2003. Structure of the tarsi in some Stenus species (Coleoptera, Staphylinidae): external morphology, ultrastructure, and tarsal secretion. J. Morphol. 255, 24–43. ( 10.1002/jmor.10044) [DOI] [PubMed] [Google Scholar]
- 113.Eisner T, Aneshansley DJ. 2000. Defense by foot adhesion in a beetle (Hemisphaerota cyanea). Proc. Natl Acad. Sci. USA 97, 6568–6573. ( 10.1073/pnas.97.12.6568) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Gorb SN. 1998. The design of the fly adhesive pad: distal tenent setae are adapted to the delivery of an adhesive secretion. Proc. R. Soc. B 265, 747–752. ( 10.1098/rspb.1998.0356) [DOI] [Google Scholar]
- 115.Langer MG, Ruppersberg JP, Gorb S. 2004. Adhesion forces measured at the level of a terminal plate of the fly's seta. Proc. R. Soc. Lond. B 271, 2209–2215. ( 10.1098/rspb.2004.2850) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Riskin DK, Racey PA. 2010. How do sucker-footed bats hold on, and why do they roost head-up? Biol. J. Linn. Soc. 99, 233–240. ( 10.1111/j.1095-8312.2009.01362.x) [DOI] [Google Scholar]
- 117.Stafford BJ, Thorington RW, Kawamichi T. 2003. Positional behavior of Japanese giant flying squirrels (Petaurista leucogenys). J. Mammal. 84, 263–271. ( 10.1644/1545-1542(2003)084%3C0263:PBOJGF%3E2.0.CO;2) [DOI] [Google Scholar]
- 118.Fujita M, Kawakami K, Moriguchi S, Higuchi H. 2008. Locomotion of the Eurasian nuthatch on vertical and horizontal substrates. J. Zool. 274, 357–366. ( 10.1111/j.1469-7998.2007.00395.x) [DOI] [Google Scholar]
- 119.Fujita M, Kawakami K, Higuchi H. 2007. Hopping and climbing gait of Japanese pygmy woodpeckers (Picoides kizuki). Comp. Biochem. Physiol. - A Mol. Integr. Physiol. 148, 802–810. ( 10.1016/j.cbpa.2006.06.048) [DOI] [PubMed] [Google Scholar]
- 120.Norberg RÅ. 1983. Optimal locomotion modes of foraging birds in trees. Ibis (Lond. 1859). 125, 172–180. ( 10.1111/j.1474-919X.1983.tb03097.x) [DOI] [Google Scholar]
- 121.Norberg R. 1986. Treecreeper climbing; mechanics, energetics, and structural adaptations. Ornis Scand. 17, 191–209. ( 10.2307/3676828) [DOI] [Google Scholar]
- 122.Norberg RÅ. 2008. Why foraging birds in trees should climb and hop upwards rather than downwards. Ibis (Lond. 1859). 123, 281–288. ( 10.1111/j.1474-919X.1981.tb04030.x) [DOI] [Google Scholar]
- 123.Goldman DI, Chen TS, Dudek DM, Full RJ. 2006. Dynamics of rapid vertical climbing in cockroaches reveals a template. J. Exp. Biol. 209, 2990–3000. ( 10.1242/jeb.02322) [DOI] [PubMed] [Google Scholar]
- 124.Autumn K, Liang YA, Hsieh ST, Zesch W, Chan WP, Kenny TW, Fearing R, Full RJ. 2000. Adhesive force of a single gecko foot-hair. Nature 405, 681–685. ( 10.1038/35015073) [DOI] [PubMed] [Google Scholar]
- 125.Tobalske BW. 2004. Take-off mechanics in hummingbirds (Trochilidae). J. Exp. Biol. 207, 1345–1352. ( 10.1242/jeb.00889) [DOI] [PubMed] [Google Scholar]
- 126.Itasse M, Moschetta J-M. 2011. Equilibrium transition study for a hybrid MAV. In Int. Micro Air Vehicle Conference and Flight Competition 2011 (IMAV2011), 30 September, 't Harde, The Netherlands, pp. 76–83.
- 127.Kossett A, D'Sa R, Purvey J, Papanikolopoulos N. 2010. Design of an improved land/air miniature robot. In 2010 IEEE Int. Conf. on Robotics and Automation, 3–8 May, Anchorage, Alaska, pp. 632–637. Piscataway, NJ: IEEE.
- 128.Kalantari A, Spenko M. 2013. Design and experimental validation of HyTAQ, a Hybrid Terrestrial and Aerial Quadrotor. In 2013 IEEE Int. Conf. on Robotics and Automation, 6–10 May, Karlsruhe, Germany, pp. 4445–4450. Piscataway, NJ: IEEE.
- 129.Kawasaki K, Zhao M, Okada K, Inaba M. 2013. MUWA: Multi-field universal wheel for air-land vehicle with quad variable-pitch propellers. In IEEE Int. Conf. on Intelligent Robots and Systems, 3–7 November, Tokyo, Japan, pp. 1880–1885. Piscataway, NJ: IEEE.
- 130.Dudley CJ, Woods AC, Leang KK. 2015. A micro spherical rolling and flying robot. In 2015 IEEE/RSJ Int. Conf. on Intelligent Robots and Systems (IROS), 28 September–2 October, Hamburg, Germany, pp. 5863–5869. Piscataway, NJ: IEEE.
- 131.Daler L, Lecoeur J, Hahlen PB, Floreano D. 2013. A flying robot with adaptive morphology for multi-modal locomotion. In 2013 IEEE/RSJ Int. Conf. on Intelligent Robots and Systems, 3–7 November, Tokyo, Japan, pp. 1361–1366. Piscataway, NJ: IEEE.
- 132.Daler L, Mintchev S, Stefanini C, Floreano D. 2015. A bioinspired multi-modal flying and walking robot. Bioinspir. Biomim. 10, 16005 ( 10.1088/1748-3190/10/1/016005) [DOI] [PubMed] [Google Scholar]
- 133.Bachmann RJ, Boria FJ, Vaidyanathan R, Ifju PG, Quinn RD. 2009. A biologically inspired micro-vehicle capable of aerial and terrestrial locomotion. Mech. Mach. Theory 44, 513–526. ( 10.1016/j.mechmachtheory.2008.08.008) [DOI] [Google Scholar]
- 134.Peterson K, Fearing RS. 2011. Experimental dynamics of wing assisted running for a bipedal ornithopter. In IEEE Int. Conf. on Intelligent Robots and Systems, 25–30 September, San Francisco, CA, pp. 5080–5086. Piscataway, NJ: IEEE.
- 135.Mulgaonkar Y, et al. 2016. The flying monkey: a mesoscale robot that can run, fly, and grasp. In 2016 IEEE Int. Conf. on Robotics and Automation (ICRA), 16–21 May, Stockholm, Sweden, pp. 4672–4679. Piscataway, NJ: IEEE.
- 136.Polakowski MR. 2012. An improved lightweight micro scale vehicle capable of aerial and terrestrial locomotion. Masters thesis. Case Western Reserve University, Cleveland, OH, USA.
- 137.Peterson K, Birkmeyer P, Dudley R, Fearing RS. 2011. A wing-assisted running robot and implications for avian flight evolution. Bioinspir. Biomim. 6, 46008 ( 10.1088/1748-3182/6/4/046008) [DOI] [PubMed] [Google Scholar]
- 138.Pratt CJ, Leang KK. 2016. Dynamic underactuated flying-walking (DUCK) robot. In 2016 IEEE Int. Conf. on Robotics and Automation (ICRA), 16–21 May, Stockholm, Sweden, pp. 3267–3274. Piscataway, NJ: IEEE.
- 139.Dickson JD, Clark JE. 2013. Design of a multimodal climbing and gliding robotic platform. IEEE/ASME Trans. Mechatron. 18, 494–505. ( 10.1109/TMECH.2012.2223708) [DOI] [Google Scholar]
- 140.Myeong WC, Jung KY, Jung SW, Jung YH, Myung H. 2015. Development of a drone-type wall-sticking and climbing robot. In 2015 12th Int. Conf. on Ubiquitous Robots and Ambient Intelligence, URAI 2015, 28–30 October, Goyang City, Korea, pp. 386–389. Piscataway, NJ: IEEE.
- 141.Shin JU, Kim D, Kim JH, Myung H. 2013. Micro aerial vehicle type wall-climbing robot mechanism. In Proc. - IEEE Int. Workshop on Robot and Human Interactive Communication, 26–29 August, Gyeongju, Korea, pp. 722–725. Piscataway, NJ: IEEE.
- 142.Thomas J, Pope M, Loianno G, Hawkes EW, Estrada MA, Jiang H, Cutkosky MR, Kumar V. 2016. Aggressive flight with quadrotors for perching on inclined surfaces. J. Mech. Robot. 8, 51007 ( 10.1115/1.4032250) [DOI] [Google Scholar]
- 143.Mellinger D, Michael N, Kumar V. 2014. Trajectory generation and control for precise aggressive maneuvers with quadrotors. In Experimental Robotics, vol. 79, Springer Tracts in Advanced Robotics, pp. 361–373. Berlin, Germany: Springer.
- 144.Jiang H, Pope MT, Estrada MA, Edwards B, Cuson M, Hawkes EW, Cutkosky MR. 2015. Perching failure detection and recovery with onboard sensing. In 2015 IEEE/RSJ Int. Conf. on Intelligent Robots and Systems (IROS), 28 September–2 October, Gyeongju, Korea, pp. 1264–1270. Piscataway, NJ: IEEE.
- 145.Graule MA, Chirarattananon P, Fuller SB, Jafferis NT, Ma KY, Spenko M, Kornbluh R, Wood RJ. 2016. Perching and takeoff of a robotic insect on overhangs using switchable electrostatic adhesion. Sci. Mag. 352, 978–982. ( 10.1126/science.aaf1092) [DOI] [PubMed] [Google Scholar]
- 146.Yanagimura K, Ohno K, Okada Y, Takeuchi E, Tadokoro S. 2014. Hovering of MAV by using magnetic adhesion and winch mechanisms. In 2014 IEEE Int. Conf. on Robotics and Automation (ICRA), 31 May–7 June, Hong Kong, China, pp. 6250–6257. Piscataway, NJ: IEEE.
- 147.Anderson M, Perry C, Hua B. 2009. The sticky-pad plane and other innovative concepts for perching UAVs. In AIAA Aerospace Sciences Meeting, pp. 1–10. Reston, VA: AIAA.
- 148.Lentink D, Dickinson MH. 2009. Rotational accelerations stabilize leading edge vortices on revolving fly wings. J. Exp. Biol. 212, 2705–2719. ( 10.1242/jeb.022269) [DOI] [PubMed] [Google Scholar]
- 149.Seigler T, Lubbers J, Reich G. 2010. Perch landing manuevers for a rotating wing MAV. In 51st AIAA/ASME/ASCE/AHS/ASC Structures, Structural Dynamics, and Materials Conf., 12–15 April, Orlando, Florida, pp. 1–12. Reston, VA: AIAA.
- 150.Polet DT, Rival DE, Weymouth GD. 2015. Unsteady dynamics of rapid perching manoeuvres. J. Fluid Mech. 767, 323–341. ( 10.1017/jfm.2015.61) [DOI] [Google Scholar]
- 151.Wickenheiser AM, Garcia E. 2008. Optimization of perching maneuvers through vehicle morphing. J. Guid. Control. Dyn. 31, 815–823. ( 10.2514/1.33819) [DOI] [Google Scholar]
- 152.Wickenheiser AM, Garcia E. 2006. Longitudinal dynamics of a perching aircraft. J. Aircr. 43, 1386–1392. ( 10.2514/1.20197) [DOI] [Google Scholar]
- 153.Wright K, Lind R. 2007. Investigating sensor emplacement on vertical surfaces for a biologically-inspired morphing design from bats. In AIAA Atmospheric Flight Mechanics Conf. and Exhibit, 20–23 August, Hilton Head, SC, pp. 1–8. Reston, VA: AIAA. [Google Scholar]
- 154.Hurst A, Wickenheiser A, Garcia E. 2008. Localization and perching maneuver tracking for a morphing UAV. In 2008 IEEE/ION Position, Location and Navigation Symp., pp. 1238–1245. Piscataway, NJ: IEEE. [Google Scholar]
- 155.Hurst A, Garcia E. 2011. Controller design for a morphing, perching aircraft. In SPIE Proc. (ed. MN Ghasemi-Nejhad), pp. 79771 L.
- 156.Cory RE. 2010. Supermaneuverable perching. PhD thesis. MIT, Cambridge, MA, USA.
- 157.Ackerman E. 2012. Flapping robotic birdplane lands right on your hand. In IEEE Spectrum. See http://spectrum.ieee.org/automaton/robotics/robotics-hardware/robotic-bird-airplane-lands-right-on-your-hand.
- 158.Thomas J, Loianno G, Pope M, Hawkes EW, Estrada MA, Jiang H, Cutkosky MR, Kumar V. 2015. Planning and control of aggressive maneuvers for perching on inclined and vertical surfaces. In Volume 5C: 39th Mechanisms and Robotics Conf., 2–5 August, Boston, MA, pp. V05CT08A012. New York, NY: ASME.
- 159.Mellinger D, Shomin M, Kumar V. 2010. Control of quadrotors for robust perching and landing. In Proc. of the Int. Powered Lift Conf., 5–7 October, Philadelphia, PA, pp. 119–126.
- 160.Chirarattananon P, Ma KY, Wood RJ. 2014. Fly on the wall. In 5th IEEE RAS/EMBS Int. Conf. on Biomedical Robotics and Biomechatronics, 12–15 April, São Paulo, Brazil, pp. 1001–1008. Piscataway, NJ: IEEE.
- 161.Chirarattananon P, Ma KY, Wood RJ. 2016. Perching with a robotic insect using adaptive tracking control and iterative learning control. Int. J. Rob. Res. 35, 1185–1206. ( 10.1177/0278364916632896) [DOI] [Google Scholar]
- 162.Robertson DK, Reich GW. 2013. Design and perching experiments of bird-like remote controlled planes. In 54th AIAA/ASME/ASCE/AHS/ASC Structures, Structural Dynamics, and Materials Conf., 8–11 April, Boston, MA, pp. 1–17. Reston, VA: AIAA.
- 163.Glassman E, Desbiens AL, Tobenkin M, Cutkosky M, Tedrake R. 2012. Region of attraction estimation for a perching aircraft: a Lyapunov method exploiting barrier certificates. In 2012 IEEE Int. Conf. on Robotics and Automation, 14–18 May, Saint Paul, MN, pp. 2235–2242. Piscataway, NJ: IEEE.
- 164.Crandall KL, Minor MA. 2015. UAV fall detection from a dynamic perch using instantaneous centers of rotation and inertial sensing. In 2015 IEEE Int. Conf. on Robotics and Automation (ICRA), 26–30 May, Seattle, WA, pp. 4675–4679. Piscataway, NJ: IEEE.
- 165.Xie P, Ma O, Zhang Z. 2013. A bio-inspired approach for UAV landing and perching. In AIAA Guidance, Navigation, and Control (GNC) Conference, 19–22 August, Boston, MA, pp. 1–13. Reston, VA: AIAA.
- 166.Zhang Z, Xie P, Ma O. 2013. Bio-inspired trajectory generation for UAV Perching. In IEEE/ASME Int. Conf. on Advanced Intelligent Mechatronics (AIM), 9–12 July, Wollongon, Australia, pp. 997–1002. Piscataway, NJ: IEEE.
- 167.Zhang Z, Xie P, Ma O. 2014. Bio-inspired trajectory generation for UAV perching movement based on Tau Theory. Int. J. Adv. Robot. Syst. 11, 1–13. ( 10.5772/58898) [DOI] [Google Scholar]
- 168.Nagendran A, Crowther W, Richardson R. 2012. Biologically inspired legs for UAV perched landing. IEEE Aerosp. Electron. Syst. Mag. 27, 4–13. ( 10.1109/MAES.2012.6163608) [DOI] [Google Scholar]
- 169.Xie P, Ma O, Zhang L. 2015. A bio-inspired UAV leg-foot mechanism for landing, grasping and perching tasks. In AIAA Sci. Tech., pp. 1–15.
- 170.Doyle CE, Bird JJ, Isom TA, Kallman JC, Bareiss DF, Dunlop DJ, King RJ, Abbott JJ, Minor MA. 2013. An avian-inspired passive mechanism for quadrotor perching. IEEE/ASME Trans. Mechatron. 18, 506–517. ( 10.1109/TMECH.2012.2211081) [DOI] [Google Scholar]
- 171.Chi W, Low KH, Hoon KH, Tang J, Go TH. 2012. A bio-inspired adaptive perching mechanism for unmanned aerial vehicles. J. Robot. Mechatron. 24, 642–648. ( 10.20965/jrm.2012.p0642) [DOI] [Google Scholar]
- 172.Doyle CE, Bird JJ, Isom TA, Johnson CJ, Kallman JC, Simpson JA, King RJ, Abbott JJ, Minor MA. 2011. Avian-inspired passive perching mechanism for robotic rotorcraft. In IEEE Int. Conf. on Intelligent Robots and Systems, 25–30 September, San Francisco, CA, pp. 4975–4980. Piscataway, NJ: IEEE.
- 173.Yeol JW, Lin C. 2014. Development of multi-tentacle micro air vehicle. In Int. Conf. on Unmanned Aircraft Systems (ICUAS), 27–30 May, Orlando, FL, pp. 815–820.
- 174.Burroughs ML, Beauwen Freckleton K, Abbott JJ, Minor MA. 2015. A sarrus-based passive mechanism for rotorcraft perching. J. Mech. Robot. 8, 11010 ( 10.1115/1.4030672) [DOI] [Google Scholar]
- 175.Culler E, Thomas G, Lee C. 2012. A perching landing gear for a quadcopter. In 53rd AIAA/ASME/ASCE/AHS/ASC Structures, Structural Dynamics and Materials Conf., 23–26 April, Honolulu, HI, pp. 1–9. Reston, VA: AIAA.
- 176.Hawkes EW, et al. 2013. Dynamic surface grasping with directional adhesion. In IEEE Int. Conf. on Intelligent Robots and Systems, 3–7 November, Tokyo, Japan, pp. 5487–5493. Piscataway, NJ: IEEE.
- 177.Menon C, Murphy M, Sitti M. 2004. Gecko inspired surface climbing robots. In IEEE Int. Conf. on Robotics and Biomimetics, 22–26 August, Shenyang, China, pp. 431–436. Piscataway, NJ: IEEE.
- 178.Aksak B, Murphy MP, Sitti M. 2008. Gecko inspired micro-fibrillar adhesives for wall climbing robots on micro/nanoscale rough surfaces. In 2008 IEEE Int. Conf. on Robotics and Automation, 19–23 May, Pasadena, CA, pp. 3058–3063. Piscataway, NJ: IEEE.
- 179.Daltorio KA, Horchler AD, Gorb S, Ritzmann RE, Quinn RD. 2005. A small wall-walking robot with compliant, adhesive feet. In 2005 IEEE/RSJ Int. Conf. on Intelligent Robots and Systems, 2–6 August, Alberta, Canada, pp. 3648–3653. Piscataway, NJ: IEEE.
- 180.Spenko MJ, Haynes GC, Saunders JA, Cutkosky MR, Rizzi AA, Full RJ, Koditschek DE. 2008. Biologically inspired climbing with a hexapedal robot. J. F. Robot. 25, 223–242. ( 10.1002/rob.20238) [DOI] [Google Scholar]
- 181.The Cornell Lab of Ornithology. 2015. Bird guide: all about birds. See https//www.allaboutbirds.org/guide/search/.
- 182.Greenewalt CH. 1962. Dimensional relationships for flying animals. Washington, DC: The Smithsonian Institution. [Google Scholar]
- 183.Socha JJ, O'Dempsey T, LaBarbera M. 2005. A 3-D kinematic analysis of gliding in a flying snake, Chrysopelea paradisi. J. Exp. Biol. 208, 1817–1833. ( 10.1242/jeb.01579) [DOI] [PubMed] [Google Scholar]
- 184.Shine R, Keogh S, Doughty P, Giragossyan H. 1998. Costs of reproduction and the evolution of sexual dimorphism in a ‘flying lizard’ Draco melanopogon (Agimidae). J. Zool. 246, 203–213. [Google Scholar]
- 185.Garbutt N. 2008. Wallace's gliding frog (Rhacophorus nigropalmatus) in mid-glide. Danum Valley, Sabah, Borneo. Nat. Pict. Libr. See http//www.naturepl.com/search/preview/wallaces-gliding-frog-rhacophorus-nigropalmatus-in-mid-glide-danum-valley/0_01174907.html.
- 186.Templin RJ. 2000. The spectrum of animal flight: insects to pterosaurs. Prog. Aerosp. Sci. 36, 393–436. ( 10.1016/S0376-0421(00)00007-5) [DOI] [Google Scholar]
- 187.Kovac M. 2016. Learning from nature how to land aerial robots. Science 352, 895–896. ( 10.1126/science.aaf6605) [DOI] [PubMed] [Google Scholar]
- 188.Stowers AK, Lentink D. 2015. Folding in and out: passive morphing in flapping wings. Bioinspir. Biomim. 10, 25001 ( 10.1088/1748-3190/10/2/025001) [DOI] [PubMed] [Google Scholar]
- 189.Labonte D, Clemente CJ, Dittrich A, Kuo C-Y, Crosby AJ, Irschick DJ, Federle W. 2016. Extreme positive allometry of animal adhesive pads and the size limits of adhesion-based climbing. Proc. Natl Acad. Sci. USA 113, 201519459 ( 10.1073/pnas.1519459113) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Asbeck AT, Kim S, Cutkosky MR, Provancher WR, Lanzetta M. 2006. Scaling hard vertical surfaces with compliant microspine arrays. Int. J. Robot. Res. Indentation Size Eff. Knoop Hardness Glas. 25, 1165–1179. ( 10.1177/0278364906072511) [DOI] [Google Scholar]
- 191.McInroe B, Astley HC, Gong C, Kawano SM, Schiebel PE, Rieser JM, Choset H, Blob RW, Goldman DI. 2016. Tail use improves performance on soft substrates in models of early vertebrate land locomotors. Science 353, 154–158. ( 10.1126/science.aaf0984) [DOI] [PubMed] [Google Scholar]
- 192.Floreano D, Zufferey J-C, Srinivasan MV, Ellington C. (eds). 2010. Flying insects and robots. Berlin, Germany: Springer. [Google Scholar]
- 193.Cutkosky MR, Kim S. 2009. Design and fabrication of multi-material structures for bioinspired robots. Phil. Trans. R. Soc. A 367, 1799–1813. ( 10.1098/rsta.2009.0013) [DOI] [PubMed] [Google Scholar]