

Abstract
Advanced age is one of the risk factors for vascular diseases that are mainly caused by impaired nitric oxide (NO) production. It has been demonstrated that endothelial arginase constrains the activity of endothelial nitric oxide synthase (eNOS) and limits NO generation. Hence, arginase inhibition is suggested to be vasoprotective in aging. In this study, we examined the effects of intravenous injection of Piceatannol, an arginase inhibitor, on aged mice. Our results show that Piceatannol administration reduced the blood pressure in aged mice by inhibiting arginase activity, which was associated with NO production and reactive oxygen species generation. In addition, Piceatannol administration recovered Ca2+/calmodulin-dependent protein kinase II phosphorylation, eNOS phosphorylation and eNOS dimer stability in the aged mice. The improved NO signaling was shown to be effective in attenuating the phenylephrine-dependent contractile response and in enhancing the acetylcholine-dependent vasorelaxation response in aortic rings from the aged mice. These data suggest Piceatannol as a potential treatment for vascular disease.
Keywords: Age-associated diseases, Arginase inhibition, Endothelial dysfunction, Endothelial nitric oxide synthase, Intravenous injection, Piceatannol
INTRODUCTION
Advanced age increases vulnerability to age-associated diseases including hypertension, atherosclerosis, and diabetes, which are collectively the greatest health problem worldwide. The relationship between the risk of cardiovascular diseases and aging is characterized by the shift from a vasodilatory, anti-coagulative, anti-proliferative and anti-inflammatory state to a vasoconstrictor, pro-coagulative, pro-proliferative and pro-inflammatory state. In the latter state, vessel reactivity to vasoconstrictors is enhanced, while endothelium-dependent vasorelaxation progressively declines with age. In contrast, endothelium-independent relaxation in response to sodium nitroprusside [a donor of nitric oxide (NO)] is unaffected by aging. The potential underlying mechanism is that aging is attributable, in part, to the progression of oxidative stress and as a result, the reduced nitric oxide (NO) level is a marker of impaired endothelial function [1,2,3].
NO is an important messenger in blood vessel dilation and immune response. Contributing to the formation of reactive nitrogen species and post-translational protein modification, NO is involved in blood vessel dilation and immune response. Furthermore, it is indicated that NO mediates mitochondrial biogenesis [4], which could play significant roles in mitochondrial function, metabolism, and age-related diseases. In the vascular endothelium, NO is synthesized from L-arginine by endothelial nitric oxide synthase (eNOS) and contributes to vasculature hemostasis by regulating vasoreactivity, platelet activation, leukocyte adhesion, smooth muscle-cell proliferation, and migration [5].
It is established that endothelial arginase, the enzyme that converts L-arginine into L-ornithine and urea, limits eNOS activity by substrate depletion [2]. Therefore, as a symptom in age-associated cardiovascular diseases, increased arginase expression/activity constrains NO bioavailability and impairs endothelial functions. It is suggested that endothelial arginase could be a novel target for the design of therapies to treat vascular diseases and other age-related diseases [6].
Piceatannol and its glucopyranoside derivative are active phenolic compounds found in the roots of rhubarbs, which have been found to possess diverse biological activities [7,8,9,10,11]. Recently, we showed that Piceatannol-3’-O-β-D-glucopyranoside was a potent inhibitor of arginase isoforms and improved endothelial function in an atherogenic mouse model via the activation of eNOS [12,13]. In particular, we showed that arginase inhibition by Piceatannol-3’-O-β-D-glucopyranoside increased the intracellular L-arginine concentration and enhanced stability of eNOS dimer, which was associated with increased NO release, and thus, Piceatannol-3’-O-β-D-glucopyranoside improved the endothelium-dependent relaxation of aortic vessels [12,13].
Our previous data indicated that arginase inhibition by a Piceatannol derivative restored endothelial dysfunction. Therefore, we tested the in vivo effect of intravenously administered Piceatannol, the main active compound, on endothelial function in aged mice.
METHODS
Animals and Piceatannol administration
Male wild-type mice (WT, C57BL/6) (Dae Han Biolink Co., South Korea) were used in this study. Mice from 15 to 16 weeks of age were considered young mice (Young), and those more than 65 weeks of age were considered aged mice (Old). Aged mice were administered with Piceatannol (Old+Pc) by intravenous injection (30 mg/kg body weight/day, dissolved 1:9 v/v in ethanol/phosphate-buffered saline, with the injection volume from 100 to 150 µl) for 4 consecutive days before experiments were carried out. The study was approved in accordance with the Guide for the Care and Use of Laboratory Animals (Institutional Review Board, Kangwon National University).
Blood pressure and heart rate measurements
Blood pressure and heart rate were measured daily by the non-invasive tail-cuff method (CODA-HT8, Kent Scientific Corporation, CT, USA) following the manufacturer’s instructions. For all groups, 15~20 measurements were taken for each mouse, and the averages of systolic blood pressure (SBP), diastolic blood pressure (DBP), mean blood pressure (MBP), and heart rate were then calculated.
Arginase activity assay
Tissue lysate was prepared using lysis buffer (50 mM Tris-HCl pH7.4, Triton X-100 0.1%, 0.1 mM EDTA and protease inhibitors) for homogenization at 4℃ followed by centrifugation for 20 min at 14,000 g and 4℃. The supernatant was used to assay for arginase activity as previously described [14].
Estimation of NO or reactive oxygen species (ROS) generation in isolated mouse aorta using DAF-FM or DHE
Fluorescence from the aortic endothelium was measured at different time intervals under microscopy as previously described [13].
Western blot analysis
Aortic vessel lysates were subjected to SDS-PAGE followed by western blot. eNOS dimer blot was performed as previously described [15]. Band intensities were analyzed using NIH ImageJ.
Aortic vascular tension assay
Mice were anesthetized using isoflurane, and the thoracic aorta was rapidly isolated and cut into 1.5 mm rings. The aortic ring were placed in ice-cold oxygenated Krebs-Ringer bicarbonate buffer (NaCl 118.3 mM, KCl 4.7 mM, MgSO4 1.2 mM, CaCl2 1.6 mM, NaHCO3 25 mM, glucose 11.1 mM; pH 7.4) and suspended between two wire stirrups (150 µm) in a myograph (Multi Myograph System, DMT-620) containing 10 ml Krebs-Ringer (95% O2-5% CO2, pH 7.4, 37℃). One stirrup was connected to a three-dimensional micromanipulator, and the other to a force transducer. The aortic rings were passively stretched at 10 min intervals in increments of 100 mg to reach the optimal tone (600 mg). After the aortic rings had been stretched to their optimal resting tone, the contractile response to 60mM KCl was determined. The response to a maximal dose of KCl was used to normalize the responses to agonist across vessel rings. Dose responses to the vasoconstrictor phenylephrine (PE, 10−9~10−5 M) were assessed, and responses to the vasodilators acetylcholine (Ach, 10−9~10−5 M) and sodium nitroprusside (SNP, 10−10~10−6 M) were assessed after pre-constriction with PE (10−5 M). To further confirm the NO-dependent vasorelaxation activity, aortic rings were treated with 1H-[1,2,4]oxadiazolo[4,3-a]quinoxalin-1-one (ODQ, 10−5 M), a soluble guanylyl cyclase inhibitor.
Statistics
Each graph represents cumulative data from three independent experiments performed in duplicate. Statistical significance was determined by one-way analysis of variance (ANOVA) (mean±SD) or two-way ANOVA (mean±SD) followed by an appropriate post-hoc test (GraphPad Prism 5 software, La Jolla, CA, USA). For all experiments, p<0.05 was considered statistically significant.
RESULTS
Piceatannol administration lowered blood pressure in mice with age-associated hypertension
As shown in Table 1, SBP, DBP, and MBP in the aged mice were higher than those in the young mice (Table 1, * vs. Young, p<0.01). After 4 consecutive days of Piceatannol administration, the SBP, DBP and MBP of the aged mice were all decreased to levels similar to those in the young mice (Table 1, † vs Old, p<0.01). Although there was no statistically significant difference in heart rate among the groups, it seemed that the administration of Piceatannol minimally increased the heart rate in the aged mice (Table 1, Old vs. Old+Pc, 536.8±48.4 vs. 580.2±45.1 bpm, ns), suggesting that arginase inhibition might enhance sympathetic outflow, as shown in a previous study [16]. Next, we sought the mechanism by which Piceatannol administration rescued blood pressure in the aged mice.
Table 1. Piceatannol administration reduces blood pressure in mice with age-associated hypertension.

Blood pressure and heart rate from each group (Young, Old, Old+Pc) were measured daily during 4 days of Piceatannol administration. *p<0.01 vs. Young, †p<0.01 vs. Old. Data are presented as the mean±SD of n=6 animals from each group.
Piceatannol inhibited the increased arginase activity in the aortic vessels of aged mice
Consistent with a previous report [17], the aged mice showed an increased aortic arginase activity (Fig. 1, * vs. Young, 161.5±0.1 vs. 100.0±10.1%, p<0.001). The administration of Piceatannol reduced the aortic arginase activity in the aged mice (Fig. 1, † vs. Old, 124.5±0.5 vs. 161.5±0.1%, p<0.001).
Fig. 1. Piceatannol administration inhibits aortic arginase activity in aged mice.
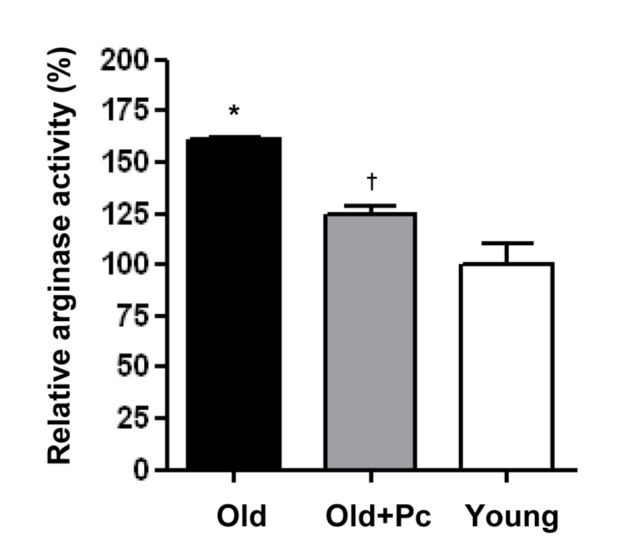
Arginase activity of isolated mouse aorta from each group (Young, Old, Old+Pc) was measured and represented as the percentage of that in the Young group (mean±SD). n=6 animals from each group. *p<0.01 vs. Young, †p<0.01 vs. Old.
Piceatannol administration recovered endothelial dysfunction in aged mice
Our previous studies indicated that arginase inhibition was associated with enhanced endothelial NO production. Hence, we used 4-Amino-5-Methylamino-2',7'-Difluorofluorescein Diacetate (DAF-FM, 5 µM), an NO-sensitive fluorescent dye, to test whether Piceatannol-dependent arginase inhibition resulted in increased NO production. As expected, endothelial NO production was attenuated in the aged mice, as indicated by reduced slope of DAF fluorescence intensity over time intervals (Fig. 2A, * vs. Young, 0.54±0.06 vs. 1.03±0.13 intensity/sec, p<0.001). This was improved by Piceatannol administration (Fig. 2A, † vs. Old, 0.86±0.07 vs. 0.54±0.06 intensity/sec, p<0.001). On the other hand, 30 min incubation of aortic vessels with NG-nitro-L-arginine methyl ester (L-NAME, 100 µM), an inhibitor of NOS, acutely reduced the DAF fluorescence slope.
Fig. 2. Arginase inhibition by Piceatannol restores NO and ROS balance in aged mice.

(A) The NO level in the mouse aortic endothelium was measured based on DAF-FM fluorescent dye. NG-nitro-L-arginine methyl ester (L-NAME, 100 µM), an inhibitor of NOS, was used for control. (B) ROS generation was estimated in isolated mouse aorta using DHE fluorescent dye. MnTBAP (Manganese (III) Tetrakis (4-Benzoic Acid) Porphyrin, 10 µM), a superoxide scavenger, was used for control. Data are presented as the mean±SD from n=6 animals from each group. *p<0.01 vs. Young, †p<0.01 vs. Old.
To determine whether increased NO production induced by arginase inhibition contributes to ROS reduction, we measured superoxide-radical generation using the fluorescent dye, Dihydroethidium (DHE, 5 µM). Compared with that in the young mice, the time-dependent slope of DHE fluorescence intensity in the aged mice was significantly increased (Fig. 2B, 0.34±0.05 vs. 0.26±0.03 intensity/sec, p<0.01). Piceatannol administration reduced the DHE fluorescence slope in the aged mice (Fig. 2B, 0.24±0.02 vs. 0.34±0.05 intensity/sec, p<0.001). Interestingly, L-NAME reduced the DHE fluorescence slope in the aged mice (Fig. 2B, 0.28±0.02 vs. 0.34±0.05 intensity/sec, p<0.001) but not in the other groups, indicating that NOS was a source of ROS generation in the aged mice. Manganese (III) Tetrakis (4-Benzoic Acid) Porphyrin (MnTBAP, 10 µM), a superoxide scavenger, nearly quenched the DHE signal.
Piceatannol induced an increase in the dimer stability and phosphorylation at Ser1177 of eNOS
To understand the mechanisms by which Piceatannol-dependent arginase inhibition improved eNOS-derived NO production, we performed western blot analysis of eNOS Ser1177 phosphorylation, a critical post-translational modification that enhanced eNOS activity. The aged mice showed a reduced eNOS Ser1177 phosphorylation level (Fig. 3A, * vs. Young, 0.39±0.11 vs. 1.00±0.17 AU, p<0.05). The administration of Piceatannol had no effect on arginase II expression (Fig. 3A, Old+Pc vs. Old, 1.07±0.25 vs. 1.14±0.29 AU, ns), but raised the level of eNOS Ser1177 phosphorylation (Fig. 3A, † vs. Old, 0.94±0.15 vs. 0.39±0.11 AU, p<0.05). As Ca2+/calmodulin-dependent protein kinase II (CaMKII) is known as a regulator of eNOS phosphorylation, we also checked CaMKII Thr286 phosphorylation, which is the activated state of CaMKII. We found that there was a decrease in CaMKII Thr286 phosphorylation in the aged mice (Fig. 3A, * vs. Young, 0.32±0.18 vs. 1.00±0.14 AU, p<0.05), which was recovered by Piceatannol administration (Fig. 3A, † vs. Old, 0.98±0.02 vs. 0.32±0.18 AU, p<0.05). These data suggested a relationship between arginase activity and CaMKII-dependent eNOS phosphorylation.
Fig. 3. Arginase inhibition by Piceatannol administration alters eNOS phosphorylation and dimer stability.

(A) Western blot and densitometric analysis of eNOS pSer1177 and arginase II in mouse aorta. (B) eNOS dimer blot and densitometric analysis. Data are presented as the mean±SD from n=6 animals from each group. *p<0.05 vs. Young, †p<0.05 vs. Old, ns, non-significant.
In addition, eNOS dimer stability was reduced in the aged mice (Fig. 3B, * vs. Young, 65.7±7.4 vs. 100.0±3.8%, p<0.01), but augmented in the Piceatannol-treated mice (Fig. 3B, † vs. Old, 147.2±10.1 vs. 65.7±7.4%, p<0.01).
Piceatannol enhanced NO signaling in the vasculature of aged mice
As Piceatannol improved NO production in the aortic endothelium through enhanced eNOS dimer stability and Ser1177 phosphorylation and eNOS dimer stability, we tested the effect of Piceatannol administration on the endothelium-dependent vasorelaxation. Mouse aortic vessels were pre-constricted with PE, and dose-response curves to Ach and SNP were constructed. An attenuated maximum response (Emax) to Ach in the aged mice was observed (Fig. 4A, * vs. Young, 55.0±2.3 vs. 79.3±2.8%, p<0.05). The reduced Ach maximum response was markedly improved by Piceatannol administration (Fig. 4A, † vs. Old, 73.6±3.4 vs. 55.0±2.3%, p<0.05). However, SNP, the NO donor, induced similar dose-response curves in aortic vessels from all the groups (Fig. 4B). The cumulative PE-dependent vasoconstriction response was increased in the aortic vessels of the aged mice (Fig. 4C, * vs. Young, 131.7±4.7 vs. 69.4±2.1%, p<0.01), which was reversed in the Piceatannol-administered mice (Fig. 4C, † vs. Old, 63.5±4.3 vs. 131.7±4.7%, p<0.01). To further confirm that the changes in vessel reactivity were dependent on NO signaling, the responses to ODQ, an inhibitor of soluble guanylyl cyclase, were measured. After pre-constriction with PE, incubation with ODQ further constricted aortic vessels in all the groups, but the effects were not different among the groups (Fig. 4D).
Fig. 4. Effects of arginase inhibition by Piceatannol administration on vessel reactivity.

(A) Cumulative dose-response curve to acetylcholine (Ach) after pre-constriction by phenylephrine (PE, 10−3 M). (B) Cumulative dose-response curve to sodium nitroprusside (SNP) after pre-constriction by phenylephrine (10−3 M). (C) Cumulative dose-response curve to phenylephrine normalized to the maximum response to KCl (60 mM). (D) Constriction response to a soluble guanylyl cyclase inhibitor, 1H-[1,2,4]oxadiazolo[4,3-a]quinoxalin-1-one (ODQ, 10−5 M). Data are presented as the mean±SD from n=6 animals from each group. *p<0.05 vs. Young, †p<0.05 vs. Old, ns, non-significant.
DISCUSSION
There is emerging evidence showing that enhanced arginase expression/activity is associated with vascular diseases. In the vasculature, arginase exists in two isoforms, namely arginase I and arginase II, but only arginase II expression is detectable in the mouse aorta [17]. We found that in aged mice, although arginase activity was increased, there was no change in the arginase II protein expression level. Hence, further study needs to be carried out to find the mechanisms associated with arginase II up-regulation.
The decline in eNOS-derived NO during aging could be due to several mechanisms: 1) L-arginine (substrate of eNOS) depletion, 2) BH4 (cofactor of eNOS) depletion, 3) eNOS uncoupling, and 4) altered eNOS phosphorylation. We have shown that increased arginase activity in aged mice was associated with a loss of eNOS dimerization and reduced eNOS Ser1177 phosphorylation. We also found that the administration of Piceatannol, as an arginase inhibitor, improved eNOS dimer stability and eNOS Ser1177 phosphorylation, which could be partially explained by the activation of CaMKII. The recovered eNOS/NO signaling was shown to be effective in vasoreactivity.
Despite the fact that eNOS normally produces NO, it also produces superoxide in the uncoupled state due to the absence of either BH4 or L-arginine. Oxidative stress in aging may damage and deplete BH4, contributing to eNOS uncoupling [18]. In addition, limited L-arginine bioavailability also promotes eNOS uncoupling [19]. Neither BH4 nor L-arginine level in endothelial cells was measured in our present study. However, we demonstrated that in aged vessels, there is an up-regulation of arginase activity, which might be a modulator of the intracellular L-arginine concentration [20]. In particular, it is suggested that endothelial cells contain two pools of L-arginine, pool I and pool II (divided into IIA and IIB) [21,22]. Extracellular L-arginine, as a basic amino acid like L-lysine, can be transported through the plasma membrane by cationic transporters to build up pool I. eNOS is accessible to the L-arginine in pool II, which is distinct from pool I and poorly interchangeable with the extracellular space. As pool IIB is used by mitochondrial arginase II, increased arginase activity may deplete the local concentration of L-arginine available to eNOS. It has also previously been demonstrated that the silencing of arginase II, but not that of arginase I, recovered the lost L-arginine bioavailability in aged mouse vessels [17]. Our present study is in accordance with that concept.
Uncoupled eNOS, along with the mitochondrial electron transport chain and activation of xanthine oxidase and NADPH oxidase, is a major sources of enhanced vascular ROS production in pathophysiological states [23]. Our data support the hypothesis that in age-related endothelial dysfunction, eNOS uncoupling is associated with increased ROS production, and Piceatannol-dependent arginase inhibition restores eNOS dimer state and recovers the nitroso-redox balance in the cardiovascular system. Hence, endothelial arginase is a valuable target in age-related vascular diseases.
As NO has multiple vasoprotective characteristics, NO signaling-based therapeutics are under investigation such as dietary L-arginine [24,25,26,27,28], drug-eluting stents [29], inhalational NO gas [30,31], and NOS gene therapy [32,33]. Unfortunately, oral L-arginine therapy produced controversial results [25,27,28]. The inhalation of NO gas can cause systemic toxicity [34,35], and pharmacological NO donors may contribute to vascular injury prior to the initiation of therapy because of surgical manipulation. In addition, gene therapy has a limited use because of inadequate delivery systems and mutational concerns. In contrast, pharmaceutical arginase inhibitors benefit vascular function in atherogenesis, diabetes, and aging [2,6,14,36,37]. Our data support the hypothesis that arginase inhibition is curative in vivo, and hence, the identification of a highly potent arginase inhibitor would contribute significantly to the treatment of vascular diseases.
In summary, Piceatannol administration by intravenous injection can improve vascular function in aged mice. Piceatannol inhibited arginase activity and reciprocally increased NO generation by increasing eNOS Ser1177 phosphorylation and eNOS dimer stability. Our results suggests that Piceatannol, as an arginase inhibitor, could be used for the development of therapies for vascular disease.
ACKNOWLEDGEMENTS
This study was supported by the Basic Science Research Program of the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (2015R1D1A3A01017911 and 2016M3A9B6903185 to S.R.). This study was supported by 2016 Research Grant from Kangwon National University (No. 520150402).
Footnotes
Author contributions: M.C.N., performed the experiments and wrote manuscript. S.R. supervised and coordinated the study and wrote the manuscript.
CONFLICTS OF INTEREST: The authors declare no conflicts of interest.
References
- 1.Seals DR, Jablonski KL, Donato AJ. Aging and vascular endothelial function in humans. Clin Sci (Lond) 2011;120:357–375. doi: 10.1042/CS20100476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Berkowitz DE, White R, Li D, Minhas KM, Cernetich A, Kim S, Burke S, Shoukas AA, Nyhan D, Champion HC, Hare JM. Arginase reciprocally regulates nitric oxide synthase activity and contributes to endothelial dysfunction in aging blood vessels. Circulation. 2003;108:2000–2006. doi: 10.1161/01.CIR.0000092948.04444.C7. [DOI] [PubMed] [Google Scholar]
- 3.Katusic ZS. Mechanisms of endothelial dysfunction induced by aging: role of arginase I. Circ Res. 2007;101:640–641. doi: 10.1161/CIRCRESAHA.107.162701. [DOI] [PubMed] [Google Scholar]
- 4.Nisoli E, Falcone S, Tonello C, Cozzi V, Palomba L, Fiorani M, Pisconti A, Brunelli S, Cardile A, Francolini M, Cantoni O, Carruba MO, Moncada S, Clementi E. Mitochondrial biogenesis by NO yields functionally active mitochondria in mammals. Proc Natl Acad Sci U S A. 2004;101:16507–16512. doi: 10.1073/pnas.0405432101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Moncada S, Higgs A. The L-arginine-nitric oxide pathway. N Engl J Med. 1993;329:2002–2012. doi: 10.1056/NEJM199312303292706. [DOI] [PubMed] [Google Scholar]
- 6.Ryoo S, Gupta G, Benjo A, Lim HK, Camara A, Sikka G, Lim HK, Sohi J, Santhanam L, Soucy K, Tuday E, Baraban E, Ilies M, Gerstenblith G, Nyhan D, Shoukas A, Christianson DW, Alp NJ, Champion HC, Huso D, Berkowitz DE. Endothelial arginase II: a novel target for the treatment of atherosclerosis. Circ Res. 2008;102:923–932. doi: 10.1161/CIRCRESAHA.107.169573. [DOI] [PubMed] [Google Scholar]
- 7.Choi KH, Kim JE, Song NR, Son JE, Hwang MK, Byun S, Kim JH, Lee KW, Lee HJ. Phosphoinositide 3-kinase is a novel target of piceatannol for inhibiting PDGF-BB-induced proliferation and migration in human aortic smooth muscle cells. Cardiovasc Res. 2010;85:836–844. doi: 10.1093/cvr/cvp359. [DOI] [PubMed] [Google Scholar]
- 8.Choi SZ, Lee SO, Jang KU, Chung SH, Park SH, Kang HC, Yang EY, Cho HJ, Lee KR. Antidiabetic stilbene and anthraquinone derivatives from Rheum undulatum. Arch Pharm Res. 2005;28:1027–1030. doi: 10.1007/BF02977396. [DOI] [PubMed] [Google Scholar]
- 9.Matsuda H, Morikawa T, Toguchida I, Park JY, Harima S, Yoshikawa M. Antioxidant constituents from rhubarb: structural requirements of stilbenes for the activity and structures of two new anthraquinone glucosides. Bioorg Med Chem. 2001;9:41–50. doi: 10.1016/s0968-0896(00)00215-7. [DOI] [PubMed] [Google Scholar]
- 10.Moon MK, Kang DG, Lee JK, Kim JS, Lee HS. Vasodilatory and anti-inflammatory effects of the aqueous extract of rhubarb via a NO-cGMP pathway. Life Sci. 2006;78:1550–1557. doi: 10.1016/j.lfs.2005.07.028. [DOI] [PubMed] [Google Scholar]
- 11.Ngoc TM, Minh PT, Hung TM, Thuong PT, Lee I, Min BS, Bae K. Lipoxygenase inhibitory constituents from rhubarb. Arch Pharm Res. 2008;31:598–605. doi: 10.1007/s12272-001-1199-0. [DOI] [PubMed] [Google Scholar]
- 12.Woo A, Shin W, Cuong TD, Min B, Lee JH, Jeon BH, Ryoo S. Arginase inhibition by piceatannol-3'-O-β-D-glucopyranoside improves endothelial dysfunction via activation of endothelial nitric oxide synthase in ApoE-null mice fed a high-cholesterol diet. Int J Mol Med. 2013;31:803–810. doi: 10.3892/ijmm.2013.1261. [DOI] [PubMed] [Google Scholar]
- 13.Woo A, Min B, Ryoo S. Piceatannol-3'-O-beta-D-glucopyranoside as an active component of rhubarb activates endothelial nitric oxide synthase through inhibition of arginase activity. Exp Mol Med. 2010;42:524–532. doi: 10.3858/emm.2010.42.7.053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.White AR, Ryoo S, Li D, Champion HC, Steppan J, Wang D, Nyhan D, Shoukas AA, Hare JM, Berkowitz DE. Knockdown of arginase I restores NO signaling in the vasculature of old rats. Hypertension. 2006;47:245–251. doi: 10.1161/01.HYP.0000198543.34502.d7. [DOI] [PubMed] [Google Scholar]
- 15.Takimoto E, Champion HC, Li M, Ren S, Rodriguez ER, Tavazzi B, Lazzarino G, Paolocci N, Gabrielson KL, Wang Y, Kass DA. Oxidant stress from nitric oxide synthase-3 uncoupling stimulates cardiac pathologic remodeling from chronic pressure load. J Clin Invest. 2005;115:1221–1231. doi: 10.1172/JCI21968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Huynh NN, Andrews KL, Head GA, Khong SM, Mayorov DN, Murphy AJ, Lambert G, Kiriazis H, Xu Q, Du XJ, Chin-Dusting JP. Arginase II knockout mouse displays a hypertensive phenotype despite a decreased vasoconstrictory profile. Hypertension. 2009;54:294–301. doi: 10.1161/HYPERTENSIONAHA.108.121731. [DOI] [PubMed] [Google Scholar]
- 17.Shin W, Berkowitz DE, Ryoo SW. Increased arginase II activity contributes to endothelial dysfunction through endothelial nitric oxide synthase uncoupling in aged mice. Exp Mol Med. 2012;44:594–602. doi: 10.3858/emm.2012.44.10.068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Milstien S, Katusic Z. Oxidation of tetrahydrobiopterin by peroxynitrite: implications for vascular endothelial function. Biochem Biophys Res Commun. 1999;263:681–684. doi: 10.1006/bbrc.1999.1422. [DOI] [PubMed] [Google Scholar]
- 19.Böger RH, Bode-Böger SM, Mügge A, Kienke S, Brandes R, Dwenger A, Frölich JC. Supplementation of hypercholesterolaemic rabbits with L-arginine reduces the vascular release of superoxide anions and restores NO production. Atherosclerosis. 1995;117:273–284. doi: 10.1016/0021-9150(95)05582-h. [DOI] [PubMed] [Google Scholar]
- 20.Erdely A, Kepka-Lenhart D, Salmen-Muniz R, Chapman R, Hulderman T, Kashon M, Simeonova PP, Morris SM., Jr Arginase activities and global arginine bioavailability in wild-type and ApoE-deficient mice: responses to high fat and high cholesterol diets. PLoS One. 2010;5:e15253. doi: 10.1371/journal.pone.0015253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Simon A, Plies L, Habermeier A, Martiné U, Reining M, Closs EI. Role of neutral amino acid transport and protein breakdown for substrate supply of nitric oxide synthase in human endothelial cells. Circ Res. 2003;93:813–820. doi: 10.1161/01.RES.0000097761.19223.0D. [DOI] [PubMed] [Google Scholar]
- 22.Closs EI, Scheld JS, Sharafi M, Förstermann U. Substrate supply for nitric-oxide synthase in macrophages and endothelial cells: role of cationic amino acid transporters. Mol Pharmacol. 2000;57:68–74. [PubMed] [Google Scholar]
- 23.Cai H, Harrison DG. Endothelial dysfunction in cardiovascular diseases: the role of oxidant stress. Circ Res. 2000;87:840–844. doi: 10.1161/01.res.87.10.840. [DOI] [PubMed] [Google Scholar]
- 24.Blum A, Hathaway L, Mincemoyer R, Schenke WH, Kirby M, Csako G, Waclawiw MA, Panza JA, Cannon RO., 3rd Oral L-arginine in patients with coronary artery disease on medical management. Circulation. 2000;101:2160–2164. doi: 10.1161/01.cir.101.18.2160. [DOI] [PubMed] [Google Scholar]
- 25.Dudek D, Legutko J, Heba G, Bartus S, Partyka L, Huk I, Dembinska-Kiec A, Kaluza GL, Dubiel JS. L-arginine supplementation does not inhibit neointimal formation after coronary stenting in human beings: an intravascular ultrasound study. Am Heart J. 2004;147:E12. doi: 10.1016/j.ahj.2003.10.025. [DOI] [PubMed] [Google Scholar]
- 26.Böger RH. L-Arginine therapy in cardiovascular pathologies: beneficial or dangerous? Curr Opin Clin Nutr Metab Care. 2008;11:55–61. doi: 10.1097/MCO.0b013e3282f2b0c3. [DOI] [PubMed] [Google Scholar]
- 27.Walker HA, McGing E, Fisher I, Böger RH, Bode-Böger SM, Jackson G, Ritter JM, Chowienczyk PJ. Endothelium-dependent vasodilation is independent of the plasma L-arginine/ADMA ratio in men with stable angina: lack of effect of oral L-arginine on endothelial function, oxidative stress and exercise performance. J Am Coll Cardiol. 2001;38:499–505. doi: 10.1016/s0735-1097(01)01380-8. [DOI] [PubMed] [Google Scholar]
- 28.Wilson AM, Harada R, Nair N, Balasubramanian N, Cooke JP. L-arginine supplementation in peripheral arterial disease: no benefit and possible harm. Circulation. 2007;116:188–195. doi: 10.1161/CIRCULATIONAHA.106.683656. [DOI] [PubMed] [Google Scholar]
- 29.Ansel GM, Lumsden AB. Evolving modalities for femoropopliteal interventions. J Endovasc Ther. 2009;16:II82–II97. doi: 10.1583/08-2654.1. [DOI] [PubMed] [Google Scholar]
- 30.Griffiths MJ, Evans TW. Inhaled nitric oxide therapy in adults. N Engl J Med. 2005;353:2683–2695. doi: 10.1056/NEJMra051884. [DOI] [PubMed] [Google Scholar]
- 31.Ichinose F, Roberts JD, Jr, Zapol WM. Inhaled nitric oxide: a selective pulmonary vasodilator: current uses and therapeutic potential. Circulation. 2004;109:3106–3111. doi: 10.1161/01.CIR.0000134595.80170.62. [DOI] [PubMed] [Google Scholar]
- 32.Barbato JE, Kibbe MR, Tzeng E. The emerging role of gene therapy in the treatment of cardiovascular diseases. Crit Rev Clin Lab Sci. 2003;40:499–545. [PubMed] [Google Scholar]
- 33.Kibbe MR, Tzeng E. Nitric oxide synthase gene therapy in vascular pathology. Semin Perinatol. 2000;24:51–54. doi: 10.1016/s0146-0005(00)80056-7. [DOI] [PubMed] [Google Scholar]
- 34.Matsumoto A, Momomura S, Hirata Y, Aoyagi T, Sugiura S, Omata M. Inhaled nitric oxide and exercise capacity in congestive heart failure. Lancet. 1997;349:999–1000. doi: 10.1016/s0140-6736(05)62897-8. [DOI] [PubMed] [Google Scholar]
- 35.Roger N, Barberà JA, Roca J, Rovira I, Gómez FP, Rodriguez-Roisin R. Nitric oxide inhalation during exercise in chronic obstructive pulmonary disease. Am J Respir Crit Care Med. 1997;156:800–806. doi: 10.1164/ajrccm.156.3.9611051. [DOI] [PubMed] [Google Scholar]
- 36.Romero MJ, Platt DH, Tawfik HE, Labazi M, El-Remessy AB, Bartoli M, Caldwell RB, Caldwell RW. Diabetes-induced coronary vascular dysfunction involves increased arginase activity. Circ Res. 2008;102:95–102. doi: 10.1161/CIRCRESAHA.107.155028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Steppan J, Ryoo S, Schuleri KH, Gregg C, Hasan RK, White AR, Bugaj LJ, Khan M, Santhanam L, Nyhan D, Shoukas AA, Hare JM, Berkowitz DE. Arginase modulates myocardial contractility by a nitric oxide synthase 1-dependent mechanism. Proc Natl Acad Sci U S A. 2006;103:4759–4764. doi: 10.1073/pnas.0506589103. [DOI] [PMC free article] [PubMed] [Google Scholar]