

Abstract
Background
Xian-Fang-Huo-Ming-Yin (XFHM), a traditional herbal formula, has been used to treat sores and carbuncles for hundreds of years in Asia. Nowadays, its clinical effects in treatment of rheumatoid arthritis (RA) have been validated. In this study, we want to study its possible molecular mechanisms of regulating the differentiation of lymphocytes and production of pro-inflammatory cytokines in collagen-induced arthritis (CIA) mice for RA treatment.
Methods
A high performance liquid chromatography-electrospray ionization/mass spectrometer (HPLC-ESI/MSn) system was used to analyze the constituents of XFHM granules. An arthritics mouse model was induced by collagen and leflunomide (LEF) was used as a positive control medicine. Pathological changes at the metatarsophalangeal joint were studied through Safranin O and immunohistochemical staining. The differentiation of T, B and NK cells was examined by flow cytometry and pro-inflammatory cytokines were assayed using an Inflammation Antibody Array assay. The expression of key molecules of the nuclear factor κB (NF-κB) and Janus kinase/signal transducers and activators of transcription (JAK/STAT) signaling pathways in spleen were studied by western-blot analysis.
Results
In our study. 21 different dominant chemical constituents were identified in XFHM. Treatment with XFHM suppressed the pathological changes in arthrosis of CIA. Additionally, XFHM down-regulated the proliferation and differentiation of CD3+ T cells and CD3−CD19+ B cells significantly. However, XFHM had no significant effect on CD3−NK1.1+ NK cells. Further study showed that the production of pro-inflammatory cytokines had been suppressed by inhibiting the activation of NF-κB and JAK/STAT signaling.
Conclusions
XFHM can regulate and maintain the immunologic balance of lymphocytic immunity and inhibit the production of pro-inflammatory cytokines, thus suppressing the pathological changes of RA. Therefore, XFHM may be used as an application of traditional medicine against RA in modern complementary and alternative therapeutics.
Electronic supplementary material
The online version of this article (doi:10.1186/s12906-016-1526-x) contains supplementary material, which is available to authorized users.
Keywords: Xian-fang-huo-ming-Yin (XFHM), Lymphocyte, Pro-inflammatory cytokine, Nuclear factor κB (NF-κB), Janus-activated kinase-signal transducer and activator of transcription (JAK/STAT), Collagen-induced arthritis (CIA)
Background
Rheumatoid arthritis (RA) is a chronic inflammatory autoimmune disease with articular and systemic effects [1]. The pathogenesis of RA is not fully understood. Various inflammatory cells, including innate immune cells (mast cells, macrophages, dendritic cells and natural killer (NK) cells) and adaptive immune cells (T and B cells), are activated in RA joints, contributing to the pathogenesis of RA. Endothelial cells and fibroblast-like synoviocytes also participate in the disease progress of RA [2]. These cells constitute the synovial lining layer and participate in an inflammatory cascade, eventually inducing the destruction of cartilage and bone [3].
It is well known that RA is a T-cell dependent autoimmune disease. CD3+ T cells, cooperating with the inflammatory cytokines excreted by T cells, induce synovial inflammation, further leading to the destruction of articular cartilage [4]. B cells also contribute to RA pathogenesis by antigen presentation and inflammatory cytokines production [5]. Cell-cell interaction of T and B cells promote the excretion of cytokines and chemokines, reinforce the interaction of the feedback loop among T cells, macrophages and B cells, and maintain the progress of synovial inflammation and destruction of cartilage and bone [6]. NK cells have been reported to play an important role in the tissue pathology of RA [7]. NK cells have a disease-promoting function on RA tissue. A subset of NK cells expanding in inflamed synovium has been demonstrated, after which the production of interferon (IFN) γ is induced to a significant degree [8, 9]. NK cells cooperating with other cell types by cytokines and chemokines may be a potential risk for RA pathogenesis.
Pro-inflammatory cytokines play key roles in the pathophysiology of RA. It is well established that tumor necrosis factor (TNF) α and interleukin (IL)-6 play the dominant roles [10, 11]. IL-1, IL-17 and IFNγ also contribute to the process of RA [10, 12]. These pro-inflammatory cytokines mediate cell migration and cause synovial inflammation, which eventually result in cartilage and bone destruction. Nuclear factor κB (NF-κB) plays a central role in the differentiation, activation and survival of mammalian cells, contributing to RA in multiple ways. It can induce the production of pro-inflammatory cytokines and other mediators of inflammation, thereby accelerating the progress of RA pathology [13]. In addition, the Janus kinase/signal transducers and activators of transcription (JAK/STAT) signaling cascade activated by IL-6 participates in the pathogenesis of RA [14].
Xian-Fang-Huo-Ming-Yin (XFHM), is a formula described in the ancient Chinese herbal treatise Jiaozhu Furen Liangfang, which was compiled by Xue Ji during the Ming dynasty of ancient China. This typical traditional Chinese medical (TCM) formula has been used for hundreds of years. It is used to treat many diseases such as sores and carbuncles. We used an optimized formula of XFHM (other used name Lijie capsule) as a TCM treatment for RA and found a good therapeutic effect [15]. Additionally, previous studies from our laboratory suggested that the optimized formula of XFHM has inhibitory effects on apoptosis of lymphocytes in rats with adjuvant arthritis [16]. In previous studies, we discovered that chlorogenic acid and Luteolin, the main components of Caulis Lonicerae Japonicae (the monarch drug in XFHM), can inhibit the inflammatory proliferation of rat synovial cells induced by IL-1β [17] and IL-6 [18]. In addition, several studies have indicated that both the crude herbs and the active ingredients of these herbs have beneficial effects on RA. These effective properties include anti-inflammation [19, 20], anti-oxidation [21], anti-proliferation [22], promoting bone metabolism[23] and stimulating osteoblasts proliferation [24]. Therefore, we suggest that XFHM has inhibitory effects on the inflammatory proliferation of synoviocytes and the subsequent destruction of cartilage and bone.
In this study, full ingredient granules of XFHM were used as the treatment drug. The quantity control of the full composition granules of XFHM was assayed using the infrared fingerprint spectrum (IRFP) technique [25]. High performance liquid chromatography-electrospray ionization/mass spectrometer (HPLC-ESI/MSn) analysis was used to characterize the phytochemicals of XFHM. Leflunomide (LEF), a disease-modifying anti-rheumatic drug (DMARD), was used as a positive control medicine. Collagen-induced arthritis (CIA) in DBA1/J mice induced by immunization with bovine CII in freund’s complete adjuvant (CFA) was used as an animal model. This investigation was undertaken to determine the regulatory effects of XFHM on the proliferation and differentiation of T, B, and NK cells, and the production of pro-inflammatory cytokines in CIA mice.
Methods
Herb materials and preparation of XFHM
The modified formula of XFHM was composed of 12 medicinal herbs. Full composition granules of the 12 herbs were provided by Beijing Tcmages Pharmaceutical Co. LTD (Beijing, China). Quality control of the XFHM granules was executed through infrared spectrum fingerprint (IFRP). The IRFP graph is shown in Additional file 1: Figure S1. The constitution ratio of 12 herbs was Atractylodes lancea (2), Ligusticum chuanxiong Hort (2), Paeonia veitchii Lynch (2), Tail of Radix Angelicae sinensis (2), Angelica dahurica (3), Radix Saposhnikoviae (3), Boswellia carteri Birdw (1), Commiphora myrrha (1), Astragalus membranaceus (8), Caulis Lonicerae Japonicae (6), Gentiana macrophylla Pall (3) and Rehmannia glutinosa Libosch (3).
HPLC-ESI/MSn analysis
HPLC-ESI/MSn analysis was performed on a Shimadzu 20LC (Kyoto, Japan) coupled to a diode array detector and TripleTOF 4600+ CDS mass spectrometer (AB Sciex, MA, USA). The chromatographic separations were carried out on an Agilent Poroshell C18 (2.1 mm × 100 mm, 2.7 μm). The mobile phase consisted of a combination of A (0.5‰ formic acid and 2 mM acetic acid) and B (0.5‰ formic acid and 2 mM acetic acid in acetonitrile methyl alcohol (1:1)) with a linear gradient, 0–10 min (5–20%, B), 10–22 min (20–95%, B). The flow rate was 0.4 mL/min, the sample injection volume was 5 μl and the column and sample temperature were BOTH 40 °C. The diode array detector (DAD) was set at 200, 220, 250 and 280 nm for the real-time monitoring of the peak intensity. Mass spectra were simultaneously acquired using electrospray ionization in the positive and negative ionization (POS and NEG) modes at fragmentation voltages (40 Psi) over the range of m/z 50–1250. The data was acquired with IDA (information dependent acquisition) method and analyzed by Peak View Software™ 2.2 (SCIEX, Foster City, CA, USA).
CIA induction in DAB1/J mice
DBA1/J male mice (7 to 8 weeks old) purchased from HFK Bioscience Co. Ltd. (Beijing, China) were immunized intradermally at the base of the tail with 150 μg of bovine type II collagen (CII) (Sigma, St. Louis, MO, USA) emulsified with an equal volume of complete Freund’s adjuvant (CFA) (Sigma, St. Louis, MO, USA). The DBA1/J mice were boosted 21 days after immunization by intradermal injection with 150 μg of CII emulsified with incomplete Freund’s adjuvant (IFA). Animal care and use were in accordance with institutional guidelines, and all animal experiments were approved by the Institutional Animal Care and Use Committee of the National Institute of state Scientific and Technological Commission.
Drug treatment
Mice were randomly divided into 4 groups as follows (n = 6 per group): normal group, fed with control diet and orally administrated sterile saline; model group, fed same as the normal group; LEF (Batch No. 130126, Cinkate Corporation, Beijing, China) group, LEF, fed with control diet and orally daily administrated 2 mg/kg LEF daily for 28 days; and XFHM group, fed with control diet and orally administrated 5.3 g/kg XFHM daily for 28 days. Drug treatment started from the first day after booster immunization. Mice were treated with 0.1 mL drug solution by gavage every day. Mice were sacrificed on day 29 after treatment. Mice were anesthetized by isoflurane anesthesia (2–3% isoflurane with oxygen supply). The sample size calculation was based on the basic principles of hypothetical randomized controlled trial [26]. Peripheral blood (PB) was obtained by removing the eyeballs prior to sacrifice during anesthesia, and the left legs and hind paws and spleens were removed after sacrifice.
Arthritic severity scores of CIA
Arthritic severity scores of CIA were monitored every 7 days after booster immunization. The arthritic severity scores for paws were used to reflect the severity of arthritis. Scores were classified as 0 (normal joints), 1 (swelling in 1 digit or joint inflammation), 2 (swelling in 2 or 3 digits or slight paw swelling), 3 (swelling in more than 4 digits and moderate swelling in the entire paw), 4 (severe swelling and deformation of the paw). Each paw was graded, and the four scores were added together so that the maximum possible score was 16 per mouse [27, 28].
Safranin O and immunohistochemical staining
The left hind legs and paws of the mice were removed, fixed with 4% paraformaldehyde in PBS, decalcified for 10 days with EDTA, embedded in paraffin, and sectioned at 5μm thickness. For Safranin O staining, sections were placed in hematoxylin for 2 min and then washed in water for 5 min. the sections were then placed in 0.1% Safranin O solution for a further 2 min and washed again in water for 30 s. Finally, the sections were passed through a series of industrial methylated spirit concentrations (70 to 100%) for 2 min at each concentration. The sections were then clarified in xylene for 2 min. the sections were then viewed, and images were taken using a light microscope.
Immunohistochemical staining for IL-1β and IL-17 was performed. After the sections were de-paraffinized, rehydrated and washed, the sections were then antigen-retrieved with pepsin and incubated with 0.3% hydrogen peroxidase for 20 min to block endogenous peroxidase activity, followed by processing with serum for 30 min to block non-specific ligations. The sections were then treated with rabbit anti-IL-1β and IL-17 (1:100, Santa Cruz Biotechnology, CA, USA) primary antibodies overnight at 4 °C, washed and incubated with reagents from an immunohistochemical kit (Zhongshan Biotechnology Ltd, Beijing, China) in compliance with the manufacturer’s instructions and visualized with 3,3-diaminobenzidine tetrahydrochloride (DAB). Finally, the sections were counterstained with hematoxylin. The sections were then viewed and images were taken using a light microscope. The IOD (integrated optical density) analysis was performed by Image-Pro Plus 6.0 (Media Cybernetics, MD, USA). Three samples in each group were reviewed five sections. The histological changes were assessed by two experienced pathologists who were blind to the treatment.
Flow cytometry (FACS) analysis
PB (0.2 mL) and half of the spleen were harvested after the mice anesthetized. Single-cell suspensions from CIA mouse spleens and PB were isolated. For surface marker staining, fluorescence conjugated anti-mouse-CD3e-PE, anti-mouse-CD19-PE-Cyanine7 and anti-mouse- NK1.1-FITC antibodies (CST, Boston, MA, USA) were used. FACS was performed using a FACS Calibur cytometer and analyzed using the CellQuest software (Beckman Coulter, Fullerton, CA, USA).
Inflammation antibody array assay
PB (0.6 mL) of the mice was harvested. Sera were separated through centrifugation at 1000 g/25 min. The levels of inflammatory antibodies in sera were detected by RayBiotech® Mouse Inflammation Antibody Array 1 (AAM-INF-G1, Ray Biotech Inc. Norcross, GA, USA), according to the manufacturer’s instructions. Briefly, 50 μL of serum was used in inflammation cytokines assays. The standard array matrix consisted of an 11 × 8 dot grid on a 20 mm × 30 mm nitrocellulose membrane with 40 unique capture antibodies. The array kit included a biotinylated-antibody solution and chemiluminescent substrate. The cytokine array membrane was incubated with 50 μL of serum for two hours, and the membrane was then washed three times with washing buffer 1 for 5 min each, followed by washing buffer II for 5 min each. Cytokines were detected using the cytokine antibody for one hour, followed by HRP-labeled streptavidin incubation for one hour. Fluorescence signals were scanned by a Microarray Scanner (GenePix 4000B, Axon, SFO, CO, USA) at 532 nm. Data acquisition and analysis were executed using the data analysis software AAM-INF-G1.
Western blot analysis
The other half of the spleens of CIA mouse was homogenized in 1 ml of a lysis buffer (Sigma, CA, USA). The extracts were cleared by spinning at 10,000 g at 4 °C for 15 min, and then diluted with the lysis buffer to achieve approximately 2 mg/ml protein concentration. Protein samples were separated on 10% sodium dodecyl sulfate (SDS)-polyacrylamide gel electrophoresis (PAGE) and transferred onto nitrocellulose membranes (Amersham Pharmacia Biotech, Uppsala, Sweden). The membranes were incubated with primary antibodies, including anti- NF-κB kinase inhibitor (IKK)α/β, anti-NF-κB p50, anti- glycoprotein (gp)-130, anti-JAK1 and anti-STAT3 rabbit anti-mouse monoclonal antibodies (CST, Boston, MA, USA), and were then incubated with horseradish peroxidase-conjugated secondary antibody. All immunoreactive proteins were visualized with SuperSignals west Pico Chemiluminescent Substrate (Thermo Scientific, Rockford, IL, USA). Densitometry plots showing the expression of proteins were normalized to glyceraldehyde 3 - phosphate dehydrogenase (GAPDH) and expressed as fold relative to the levels in mice of the normal group.
Statistical analysis
All data are presented as the means ± standard deviation (S.D.). The statistical analyses were performed using SPSS13.0 (SPSS Inc., Chicago, IL, USA). One-way analysis of variance (ANOVA) followed by the Tukey-Kramer test for multiple comparisons were used to compare the groups. P < 0.05 was considered to indicate statistical significance.
Results
Identification of chemical constituents in XFHM by HPLC-ESI/MSn
Representative liquid chromatography-mass spectrometry chromatograms are shown in Fig. 1. In the HPLC-ESI/MSn experiment, both negative (Fig. 1a) and positive (Fig. 1b) modes were tried. Twenty-one constituents were identified by comparing the retention time with IDA method. The identified compounds are shown in Table 1.
Fig. 1.
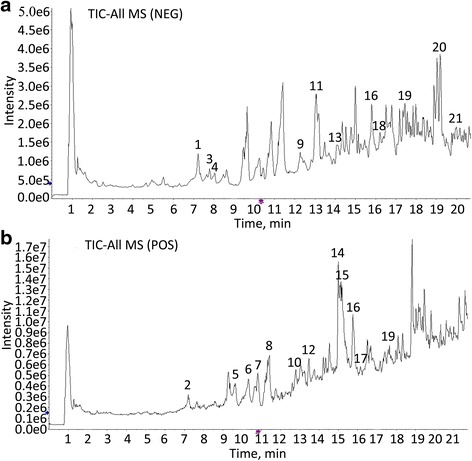
HPLC-ESI/MSn total ion chromatograms of XFHM processed by different methods. a The negative base peak MS spectrum, b the positive base peak MS spectrum
Table 1.
Chemical components identified from XFHM by HPLC-ESI/MSn
| No. | TR (min) | [M + H]/[M-H] (m/z) | Fragments (m/z) | Formula | Identification |
|---|---|---|---|---|---|
| 1 | 7.03 | /167.03 | /152.0,132.0 | C8H8O4 | Vanillic acid |
| 2 | 7.21 | 375.13 | 321.1,169.1 | C16H24O10 | Loganic acid |
| 3 | 7.73 | /179.03 | /135.0 | C9H8O4 | Caffeic acid |
| 4 | 8.03 | /353.09 | /173.0,135.0 | C16H18O9 | Chlorogenic acid |
| 5 | 9.63 | 357.12 | 195.1,121.1 | C16H20O9 | Gentiopicroside |
| 6 | 10.16 | 481.17 | 179.1,151.1 | C23H28O11 | Paeoniflorin |
| 7 | 10.81 | 391.16 | 179.1,149.0 | C17H26O10 | Loganin |
| 8 | 11.35 | 179.07 | 77.0,105.1 | C10H10O3 | Methyl 4-hydroxycinnamate |
| 9 | 12.28 | /563.14 | /383.1,473.1 | C26H28O14 | Apiin |
| 10 | 12.79 | 469.17 | 307.1,261.1 | C22H28O11 | Prim-O-glucosylcimifugin |
| 11 | 13.27 | /631.17 | /491.1,271.0 | C30H32O15 | Galloylpaeoniflorin |
| 12 | 13.44 | 307.12 | 259.1,235.1 | C16H18O6 | Cimifugin |
| 13 | 14.03 | /623.20 | /461.2,161.0 | C29H36O15 | Acteoside |
| 14 | 15.01 | 431.13 | 267.07 252.04 | C22H22O9 | Ononin |
| 15 | 15.39 | 463.16 | 301.1,167.1 | C23H26O10 | Lactiflorin |
| 16 | 15.81 | 285.06/283.06 | 220.1, 225.1 /268.0,211.0 |
C16H12O5 | Wogonin |
| 17 | 16.14 | 461.11 | 270.1,285.1 | C22H20O11 | Wogonoside |
| 18 | 16.2 | /271.06 | /151.0,119.1 | C15H12O5 | Naringenin |
| 19 | 17.55 | 269.07/267.07 | 197.1, 226.1 /252.0,223.0 |
C16H12O4 | Formononetin |
| 20 | 19.13 | /269.08 | /254.1,210.1 | C16H14O4 | Isoimperatorin |
| 21 | 20.00 | /829.46 | /621.40,651.41 | C41H68O14 | Astragaloside IV |
Effect of XFHM on histopathological changes in metatarsophalangeal joints of CIA mice
Mice treated with XFHM exhibited significant reductions in severity of CIA (Fig. 2a). On day 21 after booster immunization, the mean arthritic severity score for the group treated with XFHM was 6.17 ± 0.75, as opposed to 10.50 ± 1.05 for the model group. The XFHM treatment reduced arthritic severity scores at 0–28 days after booster injection in the CIA mice.
Fig. 2.
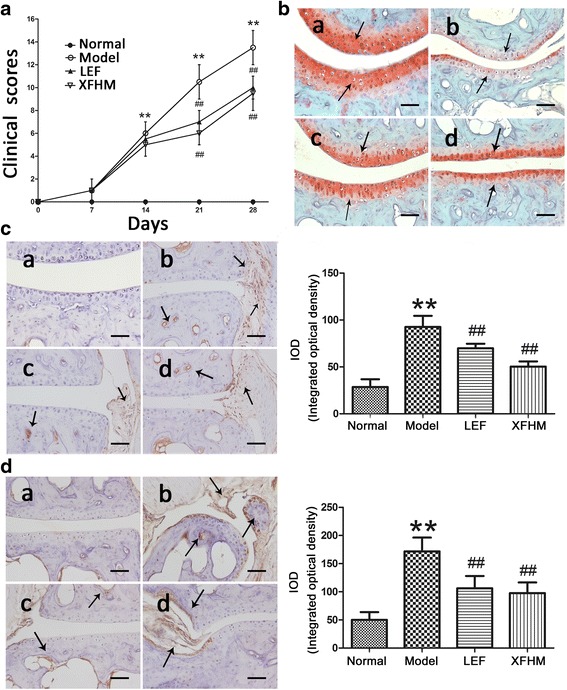
a Arthritic severity scores of CIA were monitored every 7 days after booster immunization. ** P < 0.01 indicates model group vs. normal group; # P < 0.05 and ## P < 0.01 indicate treatment groups vs. model group. b Safranin O staining of the articular cartilage. The arrows indicate Safranin O staining. c Immunohistochemical staining for IL-1β. The arrows indicate the location of IL-1β. d Immunohistochemistry staining for IL-17. The arrows indicate the location of IL-17. The results of integrated optical density (IOD) analysis are presented in the bar charts. Data are presented as the means ± S.D. (n = 3). The specimens were observed and photographed under a light microscope (Leica, DM RXA2) at magnification of 200 ×; a normal group, b model group, c group treated with LEF, d group treated with XFHM. The scale bar corresponds to 60 μm throughout
A decrease in Safranin O staining of the growth plates in CIA mice was evident prior to any morphological changes in the chondrocytes [29]. There were also differences in growth plates Safranin O staining among the groups (Fig. 2b). The intensity of Safranin O staining was significantly lower in the CIA mice relative to normal mice. After treatment with LEF and XFHM, the intensity of staining increased. Immunohistochemical detection and localization showed that IL-1β and IL-17 were expressed in the synovium, pannus and chondrocytes. As showed in Fig. 2c and d, significant differences were observed between normal and CIA mice. The abnormal expression of IL-1β and IL-17 decreased significantly with the LEF and XFHM treatments.
Intervention in the differentiation of T, B and NK cells in CIA mice
XFHM and LEF both could decrease the high levels of CD3+ T cells in PB of CIA mice (Fig. 3a and b) (P = 0.025 or 0.001). In addition, XFHM decreased the level of CD3+ T cells in spleen (Fig. 3c and d) (P = 0.018). The percentages of CD3−CD19+ B cells in PB (Fig. 3) and CD3−NK1.1+ NK cells in PB and spleen (Fig. 4) of CIA mice were increased significantly (P ≈ 0.001, 0.001 or 0.001). The abnormal rising of CD3−CD19+ B cells in PB was inhibited significantly after treatment with XFHM and LEF (Fig. 4) (P ≈ 0.001 or 0.001). Moreover, LEF inhibited the proliferation of CD3−NK1.1+ NK cells in PB and spleen (P = 0.001 or 0.029). However, XFHM had no significant regulatory effect on NK cells (Fig. 5).
Fig. 3.
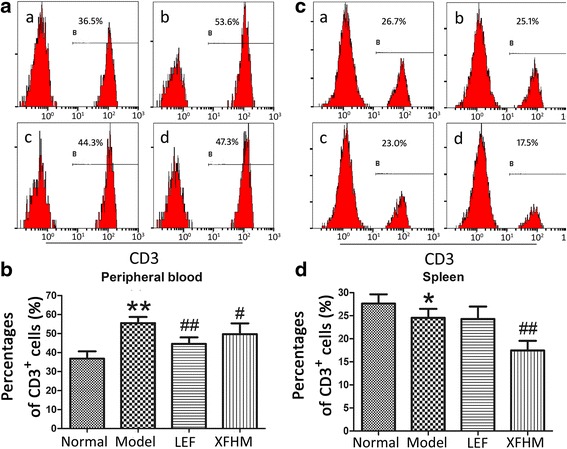
Percentages of CD3+ T cells in peripheral blood cells (PB) and splenocytes. a Histogram of flow cytometry (FCAS): a normal group, b model group, c LEF group and d XFHM group. b The results are presented in the bar charts. Data are presented as the means ± S.D. (n = 6). ** P < 0.01 indicates model group vs. normal group; # P < 0.05 and ## P < 0.01 indicate treatment groups vs. model group
Fig. 4.
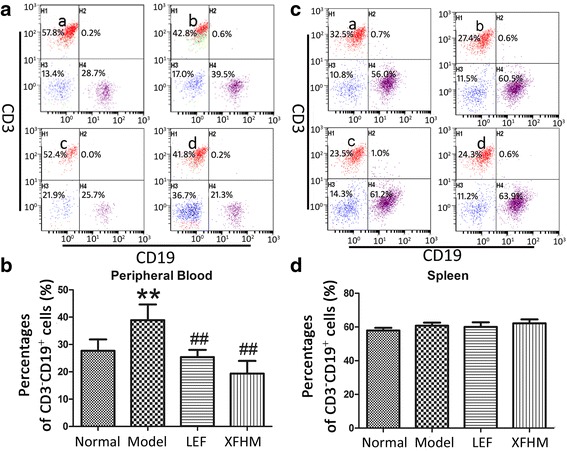
Percentages of CD3−CD19+ B cells in PB and splenocytes. a Histogram of FCAS: a normal group, b model group, c LEF group and d XFHM group. b The results are presented in the bar charts. Data are presented as the means ± S.D. (n = 6). ** P < 0.01 indicates model group vs. normal group; ## P < 0.01 indicate treatment groups vs. model group
Fig. 5.
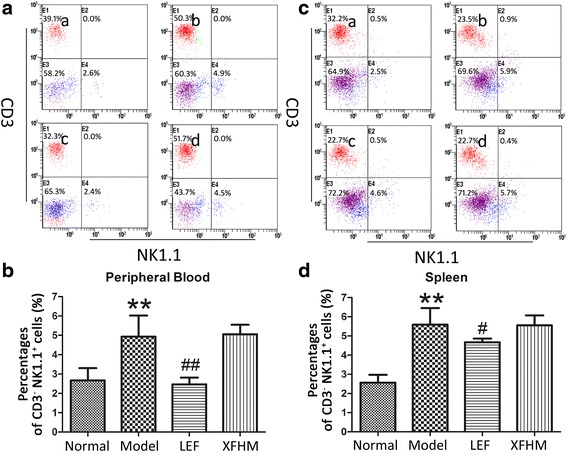
Percentages of CD3−NK1.1+ NK cells in PBMCs and splenocytes. a Histogram of FCAS: a normal group, b model group, c LEF group and d XFHM group. b The results are presented in the bar charts. Data are presented as the means ± S.D. (n = 6). ** P < 0.01 indicates model group vs. normal group; # P < 0.05 and ## P < 0.01 indicate treatment groups vs. model group
Effect of XFHM on the production of pro-inflammatory and anti-inflammatory cytokines
Next, we detected 40 types of pro-inflammatory cytokines in serum by Inflammation Antibody Array analysis. The results for 9 cytokines associated with RA are shown in Fig. 6. The levels of predominant pro-inflammatory cytokines contributing to the pathogenesis of RA, including IL-1β, TNFα, IFNγ, IL-6 and IL-17, were increased significantly in CIA mice. The production of those cytokines was restrained to different extent after treatment with LEF or XFHM. Amazingly, significant inhibitory effects appeared on the productions of IL-1β, TNFα, IFNγ and IL-6. However, there were no positive regulatory effects on IL-4, IL-10 or IL-13 from XFHM treatment.
Fig. 6.
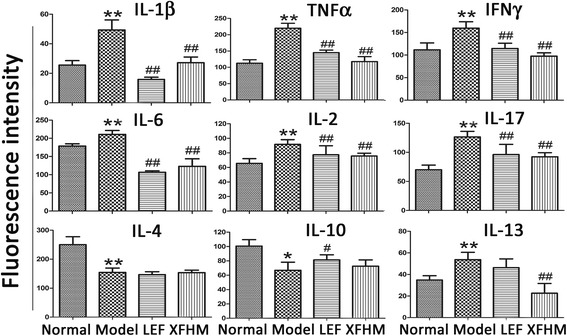
The levels of pro-inflammatory and anti-inflammatory cytokines in serum detected by inflammation antibody array assay. Data are presented as the means ± S.D. (n = 6). * P < 0.05 and ** P < 0.01 indicate model group vs. normal group; # P < 0.05 and ## P < 0.01 indicate treatment groups vs. model group
Inhibition of the activation of the NF-κB and JAK/STAT signaling pathways by XFHM treatment
We also performed western-blotting to test the production of key protein molecules of the NF-κB and JAK/STAT signaling pathways in spleen. The production of IKKα/β, NF-κB p50 (NF-κB signaling) and gp130, JAK1, and STAT3 (JAK/STAT signaling) proteins in CIA mice were increased notably, as compared with mice in the normal group (P ≈ 0.001, 0.008, 0.001, 0.001 or 0.001). As shown in Figs. 7 and 8, the abnormally higher levels of those molecules decreased significantly after LEF or XFHM treatment. These data suggest that XFHM exerts inhibitory effects on the production of pro-inflammatory cytokines by suppressing the activation of the NF-κB and JAK/STAT signaling pathways.
Fig. 7.
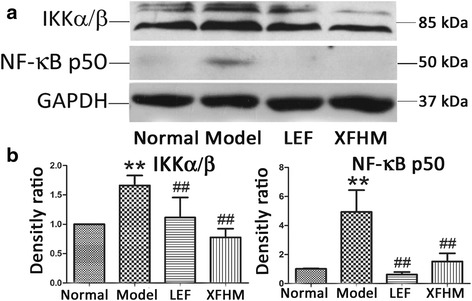
Effect of XFHM on inhibiting activation of the NF-κB signaling pathway. a and b IKKα/β and NF-κB p50 were detected in whole spleen tissue lysates by western-blot analysis. The quantified results are presented in a bar chart. GAPDH was used as an internal control. Data are presented as means ± S.D. (n = 6). ** P < 0.01 indicates model group vs. normal group; # P < 0.01 and ## P < 0.01 indicate treatment groups vs. model group
Fig. 8.
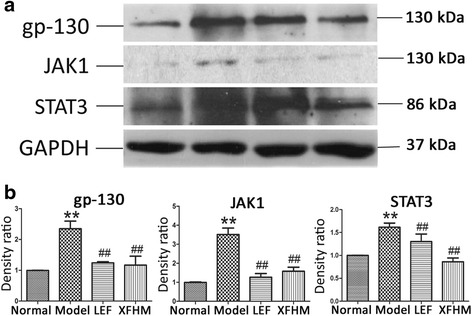
Effect of XFHM on inhibiting activation of the JAK/STAT signaling pathway. a and b gp130, JAK1 and STAT3 were detected in whole spleen tissue lysates by western-blot analysis. The quantified results are indicated by a bar chart. GAPDH was used as an internal control. GAPDH was used as an internal control. Data are presented as the means ± S.D. (n = 6). ** P < 0.01 indicates model group vs. normal group; # P < 0.01 and ## P < 0.01 indicate treatment groups vs. model group
Discussion
In TCM, combinatory therapeutic strategies according to patient symptoms are often applied to various types of diseases [30]. The combinations of herbs form multi-herbal formulae for diseases treatment. Treatment by such formulae has been verified scientifically for treatment on several diseases as an effective complementary or alternative therapy [31]. In this study, we focused on the therapeutic efficacy of a traditional herbal formula XFHM on RA, XFHM, and the mechanisms by which it regulates both the differentiation of T, B and NK cells and the production of pro-inflammatory cytokines. Suppressing effects on NF-κB and JAK/STAT signaling pathways were also detected.
As described previously, the formula of XFHM is consists of 12 medicinal herbs. It has been suggested that these herbs or the active ingredients in these herbs offer beneficial effects on RA. Ligusticum chuanxiong Hort has been demonstrated to have anti-inflammatory function [19]. Radix Astragali could reduce cell accumulation, relieve the swelling and arthritic index of joints, and down-regulate the serum concentrations of TNFα and IL-1β in adjuvant-induced arthritis [22]. Lonicera japonica Thunb was proved to have anti-inflammatory, anti-oxidative and anti-carcinogenic effects [21, 32]. The extract of Radix Gentianae Macrophyllae possesses significant antinociceptive and anti-inflammatory activities [20]. Tail Radix Angelicae Sinensis is characterized by hematopoietic, antioxidant, and immunoregulatory activities [23]. Rehmannia glutinosa Libosch and Manis tetradactyla were found to play effective roles in bone metabolism. Rehmannia glutinosa Libosch has been proved to stimulate the proliferation and activity of osteoblasts and to inhibit the generation and resorptive activity of osteoclasts [24].
The HPLC-ESI/MSn analysis was used to identify the predominant chemical constituents in XFHM. Negative and positive base peak mass spectrums were measured to obtain information about their chemical constituents. The 21 chemical constituents shown in Table 1 were identified from full composition granules of XFHM. Several lines of evidence indicate that Veratric acid [33], Loganic acid [34], Paeoniflorin [35], Loganin [36] and apiin [37] have anti-inflammatory and immune-regulatory activities. Gentiopicroside [38], Astragaloside IV [39], Acteoside [40] and Prime-O-glucosylcimifugin [41] exert anti-inflammatory effects by inhibiting the activation of NF-κB and other signaling pathways. Isoimperatorin can inhibit TNFα-induced expression of VCAM-1 by up-regulating the production of PPAR-γ and translating signals to ERK1/2, PI3K, and PKC [42]. Chlorogenic acid [17] and Caffeic acid [43] have inhibitory effects on the inflammatory proliferation of synoviocytes. Wogonin can down-regulate the production of MMP-3, acting directly on articular chondrocytes [44]. Wogonoside inhibits LPS-induced angiogenesis both in vitro and in vivo [45]. Naringenin can manipulate the immunostimulatory properties of DCs and thus represents a potential therapeutic for the treatment of RA [46]. In addition, Formononetin can increase allergic responses by enhancing IL-4 production in T cells [47]. Taken together, these findings suggest that functions of the constituents of XFHM synergistically contributed to the therapeutic effects for RA treatment.
In the pathogenesis of RA, inflammatory cells including monocytes and leukocytes infiltrate into the joint tissues, along with the proliferation of synovial lining cells, then leading to pannus formation which covers the surface of articular cartilage and bone. Pannus produces various pro-inflammatory cytokines and chemokines, leading to the destruction of cartilage and bone [48]. The present results of Safranin O and immunohistochemical staining showed that XFHM could relieve the articular cartilage injury characterized by proteoglycan loss, reduced the level of pro-inflammatory cytokines IL-1β and IL-17, inhibit the inflammatory infiltration in the synovium, and thus repair the destruction of cartilage and bone.
Antigen-presenting cells present arthritis-associated antigens to T cells, thereby initiating RA pathogenesis. Activated T cells infiltrate into the synovial membrane, combine with autocrine pro-inflammatory cytokines, and then induce inflammation in the synovium. B cells, also known as antigen-presenting cells in the pathogenesis of RA, can secrete auto-antibodies, thus further stimulating the production of pro-inflammatory cytokines. The activation of T and B cells increases the production of cytokines and chemokines, contributing to a feedback loop for the interaction of T cells, macrophages and B cells [5]. NK cells have been reported to have protective and pathogenic roles in RA [49]. Interactions of NK cells with other immune cells boost the release of pro-inflammatory cytokines, contributing to inflammation of the synovium in RA. Our results indicated that XFHM can modulate the differentiation of T and B cells, inhibiting the inflammatory progress in the synovium of RA.
Within the microenvironment of inflamed articular cartilage in RA, IL-1β, TNFα, IFNγ, IL-6 and IL-17 are the dominant pro-inflammatory cytokines. These cytokines have pleiotropic effects [48]. Myeloid origin cells, including macrophages and dendritic cells, produce most of the TNFα, IL-1β and IL-6. IFNγ and IL-17 produced by T helper 1 (Th1) and Th17cells respectively, are known as the defining cytokines for these cells. These pro-inflammatory cytokines interact synergistically, participating in the proliferation and differentiation of pathogenic cells, inflammatory cell migration, pannus formation, and the process of destruction of cartilage and bone [50, 51]. IL-4, IL-10 and IL-13, as the classic anti-inflammatory cytokines, can decrease the production of inflammatory cytokines, and thus inhibit synovial inflammation and bone damage [52, 53]. In the present study, we found that XFHM decreased the production of pro-inflammatory cytokines that promote autoimmune pathology and that it displayed immunosuppressive activities. Strangely, it had no positive regulatory effect on the production of anti-inflammatory cytokines. In TCM, combinations of plant species and minerals, called formulae, are often prescribed based on clinical experience. In formulae, multiple components could interact with multiple targets and exert synergistic therapeutic efficacy. The regulatory mechanism of XFHM on the production of pro-inflammatory and anti-inflammatory cytokines needs to be researched deeply.
NF-κB, a transcription factor, is involved in inflammation, cell survival, proliferation and differentiation. Increased NF-κB activity contributes to the chronic inflammatory characteristic of RA. NF-κB protein normally exists as homo- or hetero-dimers. The homo- or hetero-dimers combine with an NF-κB inhibitor (IκB) to form a cytoplasmic complex that inhibits its entry into the nucleus. IKKα/β mediates the phosphorylation of IκB. IκB phosphorylation prompts its proteasome-mediated degradation through ubiquitination. NF-κB dimers are released and free to migrate into the nucleus. The released NF-κB dimers bind to promoter regions and induce the expression of target genes. Previous investigations have detected NF-κB activation in RA synovium, and the expression of p50 and p65 has been observed in synovial intimal lining cells by immunohistochemical analysis [54, 55]. Furthermore, the JAK/STAT signaling pathway, known as the IL-6 pathway, offers novel potential therapeutic opportunities that aim to prevent bone destruction in RA [56]. IL-6 activates the JAK/STAT signaling cascade through binding to gp130, which triggers the phosphorylation of JAK/STAT in primary rheumatoid synoviocytes. Phosphorylated STAT3 proteins are dimerized, consequently translocating into the nucleus, and then regulate target gene transcription [57]. In this study, we discovered that XFHM could regulate the secretion of pro-inflammatory cytokines by inhibiting the activation of the NF-κB and JAK/STAT signaling pathway induced by TNFα, IL-1β or IL-6.
LEF is a classic DMARD for RA treatment. It inhibits the enzymatic activity of dihydroorotate dehydrogenase and has anti-proliferative effects. The activation of T cells depends on signal transduction of the dihydroorotate dehydrogenase pathway particularly. LEF also inhibits the activity of pro-inflammatory cytokines such as TNF and IL-1, relieves inflammation, delays the progress of joint and cartilage destruction, and thus improves the quality of life of RA patients [58]. In this study, we also obtained significant anti-inflammatory effects and inhibitory action on bone destruction through LEF treatment for CIA. However, a recent study indicated that LEF had myelosuppressive and hepatotoxic potential in a rat model of RA [59]. Adverse drug reactions to LEF, such as pruritus, loss of appetite, weak, dizziness, diarrhea and erythra, also occur frequently in clinics. Multi-herbal formulas based on traditional medicine have been scientifically verified as a complementary and alternative therapy for the treatment of various diseases. Formulae composed of a mixture of natural products have the presumed ability to target multiple sites. In our study, XFHM was verified as an effective medicine for RA treatment. It can also avoid the myelosuppressive, hepatotoxic potential and other adverse reactions that occur due to chemical drugs including LEF. In the present study, the results indicated that the effects of anti-inflammation and inhibition of bone and cartilage destruction of XFHM were equal to or exceeded those of LEF.
Conclusions
In summary, our results indicate that XFHM can suppress the inflammatory swelling in joints, inhibit inflammatory infiltration in synovium and relieve cartilage and bone destruction in CIA mice. The main mechanisms likely involve to the modulation of T, B and NK cell differentiation, a decrease in inflammatory hyperplasia of synovium, and pro-inflammatory cytokine secretion by suppressing the activation of NF-κB and the JAK/STAT signaling pathway. The use of XFHM should be an optional complementary and alternative therapy for RA treatment.
Acknowledgements
Not applicable.
Funding
This work was supported by the National Natural Science Foundation of China [81173228] and the fundamental Research Funds for Beijing University of Chinese medicine [2015-JYB -XJQ002]. These funding bodies have not participated in the design of the study and collection, analysis, and interpretation of data and in writing the manuscript.
Availability of data and material
The datasets used and/or analysed during the current study available from the corresponding author on reasonable request. All data generated or analysed during this study are included in this published article and its supplementary information files.
Authors’ contributions
JL, YW, XL, DZ and JZ carried out the experiments and analyzed the constituents of XFHM. These five authors contributed equally to this work. BN, LL, BD, AW, DZ, MC, LZ, MM and JZ participated in animal experiments and discussed results. JL, YC carried out the H&E staining and assessed the pathological changes in sections. LC participated in overall experiments, designed and coordinated the study and drafted the manuscript. All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Ethics approval and consent to participate
In this study, animal care and use were in accordance with institutional guidelines, and all animal experiments were approved by the Institutional Animal Care and Use Committee of the National Institute of state Scientific and Technological Commission.
Abbreviations
- CFA
Freund’s complete adjuvant
- CIA
Collagen-induced arthritis
- DAD
Diode array detector
- DMARD
Disease-modifying anti-rheumatic drug
- FACS
Flow cytometry
- GAPDH
Glyceraldehyde 3 - phosphate dehydrogenase
- HPLC-ESI/MSn
High performance liquid chromatography-electrospray ionization/mass spectrometer
- IFA
Incomplete Freund’s adjuvant
- IFN
Interferon
- IFRP
Infrared spectrum fingerprint
- IL
Interleukin
- JAK
Janus kinase
- LEF
Leflunomide
- NF-κB
Nuclear factor κB
- NK
Natural killer
- RA
Rheumatoid arthritis
- S.D.
Means ± standard deviation
- SDS-PAGE
Sodium dodecyl sulfate-polyacrylamide gel electrophoresis
- STAT
Signal transducers and activators of transcription
- TCM
Traditional Chinese medical
- TNF
Tumor necrosis factor
- XFHM
Xian-Fang-Huo-Ming-Yin
Additional file
The infrared spectrum fingerprint (IRFP) of YQHXJD compound. The x-axis indicated wavelength of absorption, and y-axis indicated absorption intensity. (PDF 128 kb)
Contributor Information
Jinyu Li, Email: leery_5566@163.com.
Yi Wei, Email: 251021414@qq.com.
Xue Li, Email: lxsr1210@outlook.com.
Dashuai Zhu, Email: chnzhuds1990@163.com.
Bo Nie, Email: nieboww_1977@163.com.
Jingwei Zhou, Email: 13910634708@163.com.
Lixia Lou, Email: lixialou@163.com.
Bin Dong, Email: dongbin6300@sina.com.
Aiming Wu, Email: wam688@163.com.
Yongzhe Che, Email: cheli@nankai.edu.cn.
Meng Chen, Email: cmazy@126.com.
Lingqun Zhu, Email: lingqunzhu@vip.sina.com.
Mingwei Mu, Email: never505@sohu.com.
Limin Chai, Email: liminchai@hotmail.com.
References
- 1.Choy E. Understanding the dynamics: pathways involved in the pathogenesis of rheumatoid arthritis. Rheumatology. 2012;51(Suppl 5):v3–11. doi: 10.1093/rheumatology/kes113. [DOI] [PubMed] [Google Scholar]
- 2.McInnes IB, Schett G. The pathogenesis of rheumatoid arthritis. N Engl J Med. 2011;365(23):2205–19. doi: 10.1056/NEJMra1004965. [DOI] [PubMed] [Google Scholar]
- 3.Bartok B, Firestein GS. Fibroblast-like synoviocytes: key effector cells in rheumatoid arthritis. Immunol Rev. 2010;233(1):233–55. doi: 10.1111/j.0105-2896.2009.00859.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Lundy SK, Sarkar S, Tesmer LA, Fox DA. Cells of the synovium in rheumatoid arthritis. T lymphocytes. Arthritis Res Ther. 2007;9(1):202. doi: 10.1186/ar2107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Smolen JS, Aletaha D, Koeller M, Weisman MH, Emery P. New therapies for treatment of rheumatoid arthritis. Lancet. 2007;370(9602):1861–74. doi: 10.1016/S0140-6736(07)60784-3. [DOI] [PubMed] [Google Scholar]
- 6.Smolen JS, Steiner G. Therapeutic strategies for rheumatoid arthritis. Nat Rev Drug Discov. 2003;2(6):473–88. doi: 10.1038/nrd1109. [DOI] [PubMed] [Google Scholar]
- 7.Shegarfi H, Naddafi F, Mirshafiey A. Natural killer cells and their role in rheumatoid arthritis: friend or foe? Sci World J. 2012;2012:491974. doi: 10.1100/2012/491974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Dalbeth N, Callan MF. A subset of natural killer cells is greatly expanded within inflamed joints. Arthritis Rheum. 2002;46(7):1763–72. doi: 10.1002/art.10410. [DOI] [PubMed] [Google Scholar]
- 9.Aramaki T, Ida H, Izumi Y, Fujikawa K, Huang M, Arima K, Tamai M, Kamachi M, Nakamura H, Kawakami A, et al. A significantly impaired natural killer cell activity due to a low activity on a per-cell basis in rheumatoid arthritis. Mod Rheumatol. 2009;19(3):245–52. doi: 10.3109/s10165-009-0160-6. [DOI] [PubMed] [Google Scholar]
- 10.McInnes IB, Schett G. Cytokines in the pathogenesis of rheumatoid arthritis. Nat Rev Immunol. 2007;7(6):429–42. doi: 10.1038/nri2094. [DOI] [PubMed] [Google Scholar]
- 11.Firestein GS. Evolving concepts of rheumatoid arthritis. Nature. 2003;423(6937):356–61. doi: 10.1038/nature01661. [DOI] [PubMed] [Google Scholar]
- 12.Bettelli E, Carrier Y, Gao W, Korn T, Strom TB, Oukka M, Weiner HL, Kuchroo VK. Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells. Nature. 2006;441(7090):235–8. doi: 10.1038/nature04753. [DOI] [PubMed] [Google Scholar]
- 13.Brown KD, Claudio E, Siebenlist U. The roles of the classical and alternative nuclear factor-kappaB pathways: potential implications for autoimmunity and rheumatoid arthritis. Arthritis Res Ther. 2008;10(4):212. doi: 10.1186/ar2457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Heinrich PC, Behrmann I, Muller-Newen G, Schaper F, Graeve L. Interleukin-6-type cytokine signalling through the gp130/Jak/STAT pathway. Biochem J. 1998;334(Pt 2):297–314. doi: 10.1042/bj3340297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Wei Shi NT, Jing X, Lijun Z, Limin C, Jinyu W. Clinical study on the treatment of rheumatoid arthritis in active phase with the Lijie capsule. Lishizhen Med Mater Med Res. 2012;12(21):3230–1. [Google Scholar]
- 16.Tang Nong CL, Zhang L, Lin G, Huang X. Influences of Lijie Capsules on expressions of Fas and FasL genes in lymphocytes in rats with adjuvant arthritis. J Beijing Univ Tradit Chin Med. 2006;29(7):4. [Google Scholar]
- 17.Lou L, Liu Y, Zhou J, Wei Y, Deng J, Dong B, Chai L. Chlorogenic acid and luteolin synergistically inhibit the proliferation of interleukin-1beta-induced fibroblast-like synoviocytes through regulating the activation of NF-kappaB and JAK/STAT-signaling pathways. Immunopharmacol Immunotoxicol. 2015;37(6):499–507. doi: 10.3109/08923973.2015.1095763. [DOI] [PubMed] [Google Scholar]
- 18.Lou L, Zhou J, Liu Y, Wei YI, Zhao J, Deng J, Dong B, Zhu L, Wu A, Yang Y, et al. Chlorogenic acid induces apoptosis to inhibit inflammatory proliferation of IL-6-induced fibroblast-like synoviocytes through modulating the activation of JAK/STAT and NF-kappaB signaling pathways. Exp Ther Med. 2016;11(5):2054–60. doi: 10.3892/etm.2016.3136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Huang J, Lu XQ, Zhang C, Lu J, Li GY, Lin RC, Wang JH. Anti-inflammatory ligustilides from Ligusticum chuanxiong Hort. Fitoterapia. 2013;91:21–7. doi: 10.1016/j.fitote.2013.08.013. [DOI] [PubMed] [Google Scholar]
- 20.Jia N, Li Y, Wu Y, Xi M, Hur G, Zhang X, Cui J, Sun W, Wen A. Comparison of the anti-inflammatory and analgesic effects of Gentiana macrophylla Pall. and Gentiana straminea Maxim., and identification of their active constituents. J Ethnopharmacol. 2012;144(3):638–45. doi: 10.1016/j.jep.2012.10.004. [DOI] [PubMed] [Google Scholar]
- 21.Chen WP, Wu LD. Chlorogenic acid suppresses interleukin-1beta-induced inflammatory mediators in human chondrocytes. Int J Clin Exp Pathol. 2014;7(12):8797–801. [PMC free article] [PubMed] [Google Scholar]
- 22.Jiang JB, Qiu JD, Yang LH, He JP, Smith GW, Li HQ. Therapeutic effects of astragalus polysaccharides on inflammation and synovial apoptosis in rats with adjuvant-induced arthritis. Int J Rheum Dis. 2010;13(4):396–405. doi: 10.1111/j.1756-185X.2010.01555.x. [DOI] [PubMed] [Google Scholar]
- 23.Chen XP, Li W, Xiao XF, Zhang LL, Liu CX. Phytochemical and pharmacological studies on Radix Angelica sinensis. Chin J Nat Med. 2013;11(6):577–87. doi: 10.3724/SP.J.1009.2013.00577. [DOI] [PubMed] [Google Scholar]
- 24.Oh KO, Kim SW, Kim JY, Ko SY, Kim HM, Baek JH, Ryoo HM, Kim JK. Effect of Rehmannia glutinosa Libosch extracts on bone metabolism. Clin Chim Acta. 2003;334(1–2):185–95. doi: 10.1016/S0009-8981(03)00238-9. [DOI] [PubMed] [Google Scholar]
- 25.Sun G, Li L, Li Y, Song A. Study on the digitized and quantified evaluating method for super information characteristics of herbal preparation by infrared spectrum fingerprints. Ann Transl Med. 2014;2(10):98. doi: 10.3978/j.issn.2305-5839.2014.09.13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Noordzij M, Dekker FW, Zoccali C, Jager KJ. Sample size calculations. Nephron Clin Pract. 2011;118(4):c319–23. doi: 10.1159/000322830. [DOI] [PubMed] [Google Scholar]
- 27.Kato G, Shimizu Y, Arai Y, Suzuki N, Sugamori Y, Maeda M, Takahashi M, Tamura Y, Wakabayashi N, Murali R, et al. The inhibitory effects of a RANKL-binding peptide on articular and periarticular bone loss in a murine model of collagen-induced arthritis: a bone histomorphometric study. Arthritis Res Ther. 2015;17:251. doi: 10.1186/s13075-015-0753-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Hsiao HB, Hsieh CC, Wu JB, Lin H, Lin WC. Kinsenoside inhibits the inflammatory mediator release in a type-II collagen induced arthritis mouse model by regulating the T cells responses. BMC Complement Altern Med. 2016;16:80. doi: 10.1186/s12906-016-1054-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Takahi K, Hashimoto J, Hayashida K, Shi K, Takano H, Tsuboi H, Matsui Y, Nakase T, Tomita T, Ochi T, et al. Early closure of growth plate causes poor growth of long bones in collagen-induced arthritis rats. J Musculoskelet Neuronal Interact. 2002;2(4):344–51. [PubMed] [Google Scholar]
- 30.Pan TL, Wang PW, Huang CH, Leu YL, Wu TH, Wu YR, You JS. Herbal formula, Scutellariae radix and Rhei rhizoma attenuate dimethylnitrosamine-induced liver fibrosis in a rat model. Sci Rep. 2015;5:11734. doi: 10.1038/srep11734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Xue T, Roy R. Studying traditional Chinese medicine. Science. 2003;300(5620):740–1. doi: 10.1126/science.300.5620.740. [DOI] [PubMed] [Google Scholar]
- 32.Lee HJ, Seo HS, Ryu J, Yoon YP, Park SH, Lee CJ. Luteolin inhibited the gene expression, production and secretion of MUC5AC mucin via regulation of nuclear factor kappa B signaling pathway in human airway epithelial cells. Pulm Pharmacol Ther. 2014;31:117–22. doi: 10.1016/j.pupt.2014.09.008. [DOI] [PubMed] [Google Scholar]
- 33.Choi WS, Shin PG, Lee JH, Kim GD. The regulatory effect of veratric acid on NO production in LPS-stimulated RAW264.7 macrophage cells. Cell Immunol. 2012;280(2):164–70. doi: 10.1016/j.cellimm.2012.12.007. [DOI] [PubMed] [Google Scholar]
- 34.Wei S, Chi H, Kodama H, Chen G. Anti-inflammatory effect of three iridoids in human neutrophils. Nat Prod Res. 2013;27(10):911–5. doi: 10.1080/14786419.2012.668687. [DOI] [PubMed] [Google Scholar]
- 35.Ni J, Yang D, Song L, Li C. Protective effects of paeoniflorin on alveolar bone resorption and soft-tissue breakdown in experimental periodontitis. J Periodontal Res. 2015;51(2):257–64. doi: 10.1111/jre.12305. [DOI] [PubMed] [Google Scholar]
- 36.Yamabe N, Noh JS, Park CH, Kang KS, Shibahara N, Tanaka T, Yokozawa T. Evaluation of loganin, iridoid glycoside from Corni Fructus, on hepatic and renal glucolipotoxicity and inflammation in type 2 diabetic db/db mice. Eur J Pharmacol. 2010;648(1–3):179–87. doi: 10.1016/j.ejphar.2010.08.044. [DOI] [PubMed] [Google Scholar]
- 37.Mencherini T, Cau A, Bianco G, Della Loggia R, Aquino RP, Autore G. An extract of Apium graveolens var. dulce leaves: structure of the major constituent, apiin, and its anti-inflammatory properties. J Pharm Pharmacol. 2007;59(6):891–7. doi: 10.1211/jpp.59.6.0016. [DOI] [PubMed] [Google Scholar]
- 38.Zhao L, Ye J, Wu GT, Peng XJ, Xia PF, Ren Y. Gentiopicroside prevents interleukin-1 beta induced inflammation response in rat articular chondrocyte. J Ethnopharmacol. 2015;172:100–7. doi: 10.1016/j.jep.2015.06.031. [DOI] [PubMed] [Google Scholar]
- 39.Zhang WJ, Frei B. Astragaloside IV inhibits NF- kappa B activation and inflammatory gene expression in LPS-treated mice. Mediators Inflamm. 2015;2015:274314. doi: 10.1155/2015/274314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Jing W, Chunhua M, Shumin W. Effects of acteoside on lipopolysaccharide-induced inflammation in acute lung injury via regulation of NF-kappaB pathway in vivo and in vitro. Toxicol Appl Pharmacol. 2015;285(2):128–35. doi: 10.1016/j.taap.2015.04.004. [DOI] [PubMed] [Google Scholar]
- 41.Chen N, Wu Q, Chi G, Soromou LW, Hou J, Deng Y, Feng H. Prime-O-glucosylcimifugin attenuates lipopolysaccharide-induced acute lung injury in mice. Int Immunopharmacol. 2013;16(2):139–47. doi: 10.1016/j.intimp.2013.04.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Moon L, Ha YM, Jang HJ, Kim HS, Jun MS, Kim YM, Lee YS, Lee DH, Son KH, Kim HJ, et al. Isoimperatorin, cimiside E and 23-O-acetylshengmanol-3-xyloside from Cimicifugae rhizome inhibit TNF-alpha-induced VCAM-1 expression in human endothelial cells: involvement of PPAR-gamma upregulation and PI3K, ERK1/2, and PKC signal pathways. J Ethnopharmacol. 2011;133(2):336–44. doi: 10.1016/j.jep.2010.10.004. [DOI] [PubMed] [Google Scholar]
- 43.Mia MM, Bank RA. The pro-fibrotic properties of transforming growth factor on human fibroblasts are counteracted by caffeic acid by inhibiting myofibroblast formation and collagen synthesis. Cell Tissue Res. 2015;363(3):775–89. doi: 10.1007/s00441-015-2285-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Park JS, Lee HJ, Lee DY, Jo HS, Jeong JH, Kim DH, Nam DC, Lee CJ, Hwang SC. Chondroprotective Effects of Wogonin in Experimental Models of Osteoarthritis in vitro and in vivo. Biomol Ther. 2015;23(5):442–8. doi: 10.4062/biomolther.2015.045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Chen Y, Lu N, Ling Y, Gao Y, Wang L, Sun Y, Qi Q, Feng F, Liu W, Liu W, et al. Wogonoside inhibits lipopolysaccharide-induced angiogenesis in vitro and in vivo via toll-like receptor 4 signal transduction. Toxicology. 2009;259(1–2):10–7. doi: 10.1016/j.tox.2009.01.010. [DOI] [PubMed] [Google Scholar]
- 46.Li YR, Chen DY, Chu CL, Li S, Chen YK, Wu CL, Lin CC. Naringenin inhibits dendritic cell maturation and has therapeutic effects in a murine model of collagen-induced arthritis. J Nutr Biochem. 2015;26(12):1467–78. doi: 10.1016/j.jnutbio.2015.07.016. [DOI] [PubMed] [Google Scholar]
- 47.Park J, Kim SH, Cho D, Kim TS. Formononetin, a phyto-oestrogen, and its metabolites up-regulate interleukin-4 production in activated T cells via increased AP-1 DNA binding activity. Immunology. 2005;116(1):71–81. doi: 10.1111/j.1365-2567.2005.02199.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Karmakar S, Kay J, Gravallese EM. Bone damage in rheumatoid arthritis: mechanistic insights and approaches to prevention. Rheum Dis Clin North Am. 2010;36(2):385–404. doi: 10.1016/j.rdc.2010.03.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Conigliaro P, Scrivo R, Valesini G, Perricone R. Emerging role for NK cells in the pathogenesis of inflammatory arthropathies. Autoimmun Rev. 2011;10(10):577–81. doi: 10.1016/j.autrev.2011.04.017. [DOI] [PubMed] [Google Scholar]
- 50.Astry B, Harberts E, Moudgil KD. A cytokine-centric view of the pathogenesis and treatment of autoimmune arthritis. J Interferon Cytokine Res. 2011;31(12):927–40. doi: 10.1089/jir.2011.0094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Venkatesha SH, Dudics S, Acharya B, Moudgil KD. Cytokine-modulating strategies and newer cytokine targets for arthritis therapy. Int J Mol Sci. 2015;16(1):887–906. doi: 10.3390/ijms16010887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Woods JM, Tokuhira M, Berry JC, Katschke KJ, Jr, Kurata H, Damergis JA, Jr, Arai K, Koch AE. Interleukin-4 adenoviral gene therapy reduces production of inflammatory cytokines and prostaglandin E2 by rheumatoid arthritis synovium ex vivo. J Investig Med. 1999;47(6):285–92. [PubMed] [Google Scholar]
- 53.Vermeij EA, Broeren MG, Bennink MB, Arntz OJ, Gjertsson I, LEMvL P, van den Berg WB, Koenders MI, van de Loo FA. Disease-regulated local IL-10 gene therapy diminishes synovitis and cartilage proteoglycan depletion in experimental arthritis. Ann Rheum Dis. 2015;74(11):2084–91. doi: 10.1136/annrheumdis-2014-205223. [DOI] [PubMed] [Google Scholar]
- 54.Firestein GS. NF-kappaB: Holy Grail for rheumatoid arthritis? Arthritis Rheum. 2004;50(8):2381–6. doi: 10.1002/art.20468. [DOI] [PubMed] [Google Scholar]
- 55.Simmonds RE, Foxwell BM. Signalling, inflammation and arthritis: NF-kappaB and its relevance to arthritis and inflammation. Rheumatology. 2008;47(5):584–90. doi: 10.1093/rheumatology/kem298. [DOI] [PubMed] [Google Scholar]
- 56.Alghasham A, Rasheed Z. Therapeutic targets for rheumatoid arthritis: Progress and promises. Autoimmunity. 2014;47(2):77–94. doi: 10.3109/08916934.2013.873413. [DOI] [PubMed] [Google Scholar]
- 57.O’Brien CA, Gubrij I, Lin SC, Saylors RL, Manolagas SC. STAT3 activation in stromal/osteoblastic cells is required for induction of the receptor activator of NF-kappaB ligand and stimulation of osteoclastogenesis by gp130-utilizing cytokines or interleukin-1 but not 1,25-dihydroxyvitamin D3 or parathyroid hormone. J Biol Chem. 1999;274(27):19301–8. doi: 10.1074/jbc.274.27.19301. [DOI] [PubMed] [Google Scholar]
- 58.Wiacek R, Kolossa K, Jankowski T, Jeka S, Karmowski A, Karmowski M, Gworys B. The efficacy and safety of leflunomide in patients with active rheumatoid arthritis. Adv Clin Exp Med. 2012;21(3):337–42. [PubMed] [Google Scholar]
- 59.Bilasy SE, Essawy SS, Mandour MF, Ali EA, Zaitone SA. Myelosuppressive and hepatotoxic potential of leflunomide and methotrexate combination in a rat model of rheumatoid arthritis. Pharmacol Rep. 2015;67(1):102–14. doi: 10.1016/j.pharep.2014.08.009. [DOI] [PubMed] [Google Scholar]