

Abstract
Face recognition is an important component of successful social interactions in humans. A large literature in social psychology has focused on the phenomenon termed “the other race” (ORE) effect, the tendency to be more proficient with face recognition within one’s own ethnic group, as compared to other ethnic groups. Several potential hypotheses have been proposed for this effect including perceptual expertise, social grouping, and holistic face processing. Recent work on mnemonic discrimination (i.e. the ability to resolve mnemonic interference among similar experiences) may provide a mechanistic account for the ORE. In the current study, we examined how discrimination and generalization in the presence of mnemonic interference may contribute to the ORE. We developed a database of computerized faces divided evenly among ethnic origins (Black, Caucasian, East Asian, South Asian), as well as morphed face stimuli that varied in the amount of similarity to the original stimuli (30%, 40%, 50%, and 60% morphs). Participants first examined the original unmorphed stimuli during study, then during test were asked to judge the prior occurrence of repetitions (targets), morphed stimuli (lures), and new stimuli (foils). We examined participants’ ability to correctly reject similar morphed lures and found that it increased linearly as a function of face dissimilarity. We additionally found that Caucasian participants’ mnemonic discrimination/generalization functions were sharply tuned for Caucasian faces but considerably less tuned for East Asian and Black faces. These results suggest that expertise plays an important role in resolving mnemonic interference, which may offer a mechanistic account for the ORE.
Keywords: memory, interference, pattern separation, perceptual expertise
Introduction
Memory for faces is critical to many facets of our daily lives. It allows us to communicate effectively in social settings, to understand and recognize emotional states, and to create associations (e.g. faces and names). We use faces as perhaps the most prominent and effective means of identifying one another and facial features to identify each individual as coming from a different culture and ethnic background (Blais et al. 2008). A well documented phenomenon in psychology known as the “other race” effect (ORE) is a particular example in which the role of cultural influences in face recognition are important. This effect has been observed as far back as 100 years ago, and can be characterized as the tendency of members of one race to have difficulty remembering faces from other races relative to faces from one’s own race (Feingold 1914; Bothwell, Brigham, & Malpass, 1989). The ORE has prominently been used as a criticism of eyewitness testimony, which can rely on perceivers’ attempting to recognize faces of individuals outside of their own race group (Behrman & Davey, 2001; Sporer 2001).
The ORE has been studied extensively in the social psychology literature, and several mechanisms have been proposed for its mechanisms. Prominent among these mechanisms are holistic face processing, social and motivational grouping, and perceptual expertise. Given the complex nature of face processing, these mechanisms are not mutually exclusive and the ORE may arise from a combination of multiple mechanisms. Nevertheless, each of these proposed mechanisms provides valuable insight into the potential contributing processes of the ORE.
The holistic processing hypothesis suggests that the human visual system processes faces as an accumulation of interactions between multiple parts over the entire region of the face, as opposed to a decomposition of separable facial features (Michel et al. 2007; Taubert et al. 2011). According to this hypothesis, perceivers view same race faces as a unified object, while other race faces are processed as sets of features (Michel et al. 2006; Maurer et al., 2002; Tanaka et al. 2004; Young et al., 2012). Experiments using two paradigms, the composite face (Goffaux & Rossion, 2006; Hugenberg & Corneille, 2009; Michel et al. 2007) and the whole/part paradigms (Tanaka & Farah, 1993; Tanaka et al. 2004), have demonstrated evidence for such differential processing for other-race and same-race faces. Studies using inverted faces have also shown that face inversion has a significantly larger effect on recognition accuracy than object inversion (Yin 1969). When applied to same race and other race faces, inverted same race faces dropped to the same recognition levels as other race faces (Rhodes et al. 1989), suggesting that inversion removes some of the perceptual/mnemonic advantages of same race faces.
The social and motivational grouping hypothesis posits that perceivers categorize individuals based on inherent grouping characteristics. Thus, if a person shares a similar grouping characteristic (i.e. race, age, sex), the perceiver will process the face on an individual level. If the person does not share the grouping characteristic (out-group), then the perceiver will process this face on a categorical level (Sporer 2001; Hugenberg & Sacco, 2008; Hugenberg et al., 2010). Studies in social categorization theory have shown that merely labeling or categorizing faces as belonging to an in-group can cause significant differences in recognition rates between in-group and out-group faces, even when race and perceptual expertise are held constant and counterbalanced (Hugenberg et al. 2007).
The amount of experience and interaction that individuals have with other-race and same-race faces has also been theorized to modify the ORE (Walker & Tanaka 2003; Meissner & Brigham, 2001). This notion, known as Perceptual Expertise Theory, is based upon the idea that humans interact with faces so often that they become experts at identifying faces, much like a car enthusiast is an expert at identifying the makes of models of various automobiles (Tanaka & Gauthier, 1997; Tanaka, 2001). The Perceptual Expertise Theory has been supported by studies in which the ORE was observed in adolescents (Kelly et al. 2007; Walker & Hewstone, 2006), adults (O’toole, Valentin, & Abdi, 1994), and adolescents who had been adopted at a young age and developed face expertise among the race that they grew up with (Sangrigoli et al. 2005).
We sought to examine the effect of perceptual expertise on mnemonic discrimination of same- and other-race faces using a novel interference paradigm. We propose that interference among similar experiences may mechanistically contribute to this phenomenon. This “mnemonic” interference has been studied frequently in recent literature in the context of hippocampal learning (Yassa and Stark, 2011; Reagh et al. 2013; Roberts et al. 2014). The hippocampus is well known for playing a critical role in the encoding and storage of episodic memories (Squire, 1992). One mechanism by which the hippocampus is thought to encode unique episodic memories is by reducing interference among similar inputs (i.e. pattern separation). This computation is thought to underlie our ability to discriminate among similar experiences. The hippocampus is also implicated in pattern completion, the process of re-instantiating a pre-existing representation when presented with a partial or degraded input. This computation is thought to underlie our ability to generalize or perform recognition in the face of noise. These computational processes have been long hypothesized (Marr, 1971; McClelland et al. 1995; O’Reilly & Norman, 2002; Shapiro & Olton, 1994; Treves & Rolls, 1994) and examined in the hippocampus empirically over the past ten years (cf. review by Yassa & Stark, 2011).
We suggest that it is the delicate balance among these computational processes in our memory system that leads to finely tuned discrimination/generalization abilities in same-race faces relative to other-race faces. We hypothesize that perceptual expertise when viewing same-race faces would allow for more efficient resolution of interference, manifesting in sharper representations of same-race faces and should be able to discriminate and generalize more effectively. In other words, we predict that for stimuli that have more precise representations, mnemonic discrimination functions should be more finely tuned (i.e. “sigmoidal” or “S-shaped”) with respect to stimulus similarity. This would have the consequence that there would be a sharp transition at a distinct level of similarity where observers would switch from generalization to discrimination. For other-race faces (i.e. no expertise), we propose that mnemonic representations are less precise leading to a discrimination/generalization function that is less tuned (i.e. more linear).
It is important to note that the sigmoidal function implies that effective generalization is just as important as effective discrimination. Thus, having a neural system that only discriminates would not be optimal. Take for example, a situation in which the same person changes his or her appearance slightly. It is important to generalize across such a minor perturbation, but be able to discriminate effectively when change surpasses a specific threshold that is crossed when the person has changed identity and not just appearance. Thus, having efficient generalization as well as discrimination is an important hallmark of a sharpened or tuned representation. Our predictions were also informed by work in categorical perception of face identities. Behavioral studies using classification identity tasks have shown that large changes in discrimination occur in the 40–60% morph range for participants familiar with task faces, while little to no change occurs in low percentage morphs (Angeli et al. 2008; Jacques & Rossion, 2006; Campanella et al. 2000; Rotshtein et al. 2005).
We specifically used computer generated faces to parametrically manipulate interference. While synthetic faces may be less ecologically valid than naturalistic faces, they allow us to quantitatively manipulate various facial features and incrementally change the similarity of their appearance to make recognition and discrimination parametrically challenging. This parametric manipulation of interference is necessary to test our proposal that discrimination/generalization functions are differentially tuned based on expertise.
In the current study, we used face morphs with varying levels of similarity that were distributed equally among three ethnic groups (Black, Caucasian, and East Asian) and tested a sample of Caucasian participants’ ability to mnemonically discriminate Caucasian vs. Black and East Asian faces. We find evidence for a tuned (i.e. sigmoidal) discrimination/generalization curve in Caucasian faces and an “un-tuned” (i.e. linear) curve for Black and East Asian faces, consistent with our hypothesis. These results suggest that expertise in resolving mnemonic interference may be a core mechanism for the ORE.
Materials and Methods
Participants
Sixty young adults (mean age = 19.86 yr SD = 1.39) participated in the subsequent memory experiment. All participants were recruited from Johns Hopkins University and received course credit for their participation for the experiment. All participants were cognitively intact and did not suffer from major neurological, psychiatric, or health conditions. Before testing, all participants provided written informed consent and at the conclusion of the experiment, participants were debriefed. Data from three subjects were excluded due to chance performance or technical problems. The remaining 57 subjects were divided into four groups according to ethnicity: Black (n=9), Caucasian (n=29), East Asian (n=12) and South Asian (n=7).
Interference task
The stimuli were images of novel faces that were created using commercially available software (FaceGen Modeller 3.1). An initial 104 faces were generated evenly across gender and 4 ethnic origins that the FaceGen program provided; Black, Caucasian, East Asian, and South Asian. Each of the initial generated faces were then morphed by a specific percentage that we designated a priori using a similarity function within FaceGen. The function modified characteristics of the face by a specific percentage that we chose and generated a set of 9 morphed faces per percentage, from which we chose our stimuli. These “morphs” created similar, but not identical faces from the original image. We created morphs of 30, 40, 50 and 60%, with the 30% morph being highly similar to the original face, and the 60% being more dissimilar.
During the experiment, participants saw a total of 104 face stimuli during the encoding phase, and 200 stimuli during the retrieval phase. The retrieval phase consisted of a mixture of faces that participants saw during study (targets), the similar looking morphed faces (lures), and completely novel face stimuli that had never been seen before (foils). Participants were shown 52 target faces, 96 lure faces evenly split between similarity, and 52 foil faces. Lures were 4 bins of 24 stimuli. Within those 24 stimuli, 6 stimuli were 30% morphs, 6 stimuli were 40% morphs, 6 stimuli were 50% morphs, and 6 stimuli were 60% morphs (Fig 1).”
Fig 1. Mnemonic discrimination of similar face stimuli task schematic.
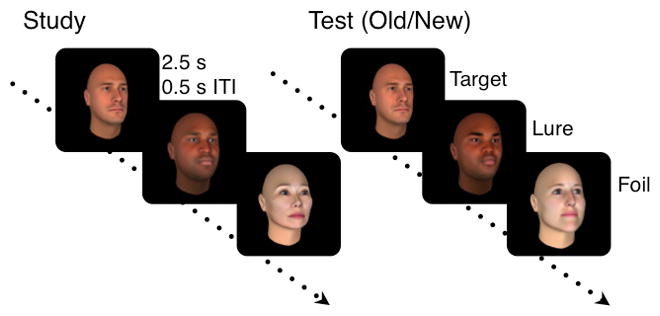
All faces were presented for 2.5s, with a 0.5s inter-stimulus interval. Study phase consisted of 104 trials. Judgments were incidental (“How pleasant is this face on a scale from 1–9?”), and subjects were unaware of a subsequent memory test. (B) Test phase consisted of 200 trials (52 repeated Targets, 96 similar Lures divided equally among 30%, 40%, 50%, and 60% morphs, and 52 novel Foils). Judgments were overtly mnemonic (“Old/New?”).
All experiments were programmed in Psychtoolbox 3.0 (Brainard, 1997; Pelli, 1997) running under MATLAB (Natick, MA) release R2010a. Each trial consisted of 2 displays: an image display and a fixation display. During both encoding and retrieval phases, images were presented on the center of the screen with a black background for 3000 ms. The fixation display consisted of a white fixation cross on the center of the screen with a black background for 1000 ms.
Participants underwent an incidental encoding phase where they were shown the initial, non-morphed face stimuli in randomized order, and were asked to rate the pleasantness of each face using a 1–9 scale (1 being most positive, 9 being most negative). Participants were told to spread their responses across the scale. This was only intended to be an incidental judgment such that we can ascertain that participants paid attention to our stimuli and had no bearing on the subsequent mnemonic judgment. Participants were given a surprise subsequent memory test immediately after encoding. Participants were told they would once again see faces appear one at a time on the screen and were asked to indicate whether the faces on the screen had been seen during the previous phase by pressing button responses on the keyboard corresponding to “old” or “new”. Participants were explicitly told that in order for a face image to be called “old,” it had to be the same exact image they saw before. However, we did not alert participants to the possibility that there would be similar looking faces presented in the task.
Our two key outcomes measures of interest were target recognition and lure discrimination index (LDI). Target recognition was measured by a discriminability index (d′), which was calculated as z(Hits) - z(False Alarms). Hits and false alarms refer to correct recognition of old items and false recognition of new items, respectively. d′ is calculated as the difference of z-transformed values. In order to measure how well participants discriminated similar items (lures), we examined performance using a bias-corrected LDI operationalized as p(New|Lure) - p(New|Target). This corrected for the general tendency to reject (i.e. call an item ‘New’) and is similar to other metrics we used in prior work (Yassa, Mattfeld, Stark, & Stark, 2011; Yassa et al., 2011).
Results
Raw performance data are shown in Table 1. First, we examined the role of interference in mnemonic discrimination of face stimuli by running a repeated measures ANOVA across the entire sample (with four interference levels: 30, 40, 50, and 60% morphs). We observed a significant main effect of interference [F(3,168) = 91.86, P < .0001]. A post hoc trend analysis revealed a significant linear [F(1,56) = 202.19, P <.0001] trend. Significant quadratic [F(1,56) = 13.99, P<.0001] and cubic [F(1,56) = 12.63, P<.001] trends were also identified (Fig 2).
Table 1.
| GROUP | AFRICAN | EUROPEAN | EAST ASIAN | SOUTH ASIAN | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
| ||||||||||||||||
| N | 9 | M:F | 5:4 | N | 29 | M:F | 12:17 | N | 12 | M:F | 7:5 | N | 7 | M:F | 3:4 | |
| TARGETS | ||||||||||||||||
|
| ||||||||||||||||
| Hits | Misses | Hits | Misses | Hits | Misses | Hits | Misses | |||||||||
| Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | |
| Black | 0.68 | 0.06 | 0.32 | 0.06 | 0.66 | 0.04 | 0.34 | 0.04 | 0.69 | 0.04 | 0.31 | 0.04 | 0.61 | 0.05 | 0.39 | 0.05 |
| European | 0.65 | 0.05 | 0.35 | 0.05 | 0.61 | 0.04 | 0.39 | 0.04 | 0.53 | 0.04 | 0.47 | 0.04 | 0.60 | 0.04 | 0.40 | 0.04 |
| East Asian | 0.67 | 0.06 | 0.33 | 0.06 | 0.57 | 0.04 | 0.43 | 0.04 | 0.63 | 0.05 | 0.37 | 0.05 | 0.52 | 0.06 | 0.48 | 0.06 |
| South Asian | 0.71 | 0.04 | 0.29 | 0.04 | 0.67 | 0.03 | 0.33 | 0.03 | 0.61 | 0.06 | 0.39 | 0.06 | 0.56 | 0.07 | 0.44 | 0.07 |
|
| ||||||||||||||||
| LURES 30% | ||||||||||||||||
|
| ||||||||||||||||
| CRs | FAs | CRs | FAs | CRs | FAs | CRs | FAs | |||||||||
| Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | |
| Black | 0.34 | 0.05 | 0.66 | 0.05 | 0.37 | 0.03 | 0.63 | 0.03 | 0.42 | 0.08 | 0.58 | 0.08 | 0.43 | 0.06 | 0.57 | 0.06 |
| European | 0.35 | 0.05 | 0.65 | 0.05 | 0.46 | 0.03 | 0.54 | 0.03 | 0.43 | 0.04 | 0.57 | 0.04 | 0.53 | 0.09 | 0.47 | 0.09 |
| East Asian | 0.39 | 0.10 | 0.61 | 0.10 | 0.48 | 0.05 | 0.52 | 0.05 | 0.55 | 0.04 | 0.45 | 0.04 | 0.39 | 0.07 | 0.61 | 0.07 |
| South Asian | 0.39 | 0.09 | 0.61 | 0.09 | 0.56 | 0.03 | 0.44 | 0.03 | 0.65 | 0.06 | 0.35 | 0.06 | 0.47 | 0.04 | 0.53 | 0.04 |
|
| ||||||||||||||||
| LURES 40% | ||||||||||||||||
|
| ||||||||||||||||
| CRs | FAs | CRs | FAs | CRs | FAs | CRs | FAs | |||||||||
| Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | |
| Black | 0.55 | 0.06 | 0.45 | 0.06 | 0.58 | 0.04 | 0.42 | 0.04 | 0.56 | 0.08 | 0.44 | 0.08 | 0.49 | 0.07 | 0.51 | 0.07 |
| European | 0.54 | 0.05 | 0.46 | 0.05 | 0.49 | 0.03 | 0.51 | 0.03 | 0.61 | 0.06 | 0.39 | 0.06 | 0.54 | 0.08 | 0.46 | 0.08 |
| East Asian | 0.64 | 0.08 | 0.36 | 0.08 | 0.68 | 0.03 | 0.32 | 0.03 | 0.72 | 0.05 | 0.28 | 0.05 | 0.62 | 0.04 | 0.38 | 0.04 |
| South Asian | 0.46 | 0.06 | 0.54 | 0.06 | 0.54 | 0.04 | 0.46 | 0.04 | 0.66 | 0.07 | 0.34 | 0.07 | 0.50 | 0.05 | 0.50 | 0.05 |
|
| ||||||||||||||||
| LURES 50% | ||||||||||||||||
|
| ||||||||||||||||
| CRs | FAs | CRs | FAs | CRs | FAs | CRs | FAs | |||||||||
| Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | |
| Black | 0.73 | 0.06 | 0.27 | 0.06 | 0.67 | 0.03 | 0.33 | 0.03 | 0.73 | 0.05 | 0.27 | 0.05 | 0.67 | 0.12 | 0.33 | 0.12 |
| European | 0.84 | 0.05 | 0.16 | 0.05 | 0.79 | 0.03 | 0.21 | 0.03 | 0.75 | 0.04 | 0.25 | 0.04 | 0.61 | 0.10 | 0.39 | 0.10 |
| East Asian | 0.79 | 0.06 | 0.21 | 0.06 | 0.76 | 0.03 | 0.24 | 0.03 | 0.83 | 0.03 | 0.17 | 0.03 | 0.71 | 0.05 | 0.29 | 0.05 |
| South Asian | 0.69 | 0.05 | 0.31 | 0.05 | 0.70 | 0.04 | 0.30 | 0.04 | 0.72 | 0.07 | 0.28 | 0.07 | 0.71 | 0.07 | 0.29 | 0.07 |
|
| ||||||||||||||||
| LURES 60% | ||||||||||||||||
|
| ||||||||||||||||
| CRs | FAs | CRs | FAs | CRs | FAs | CRs | FAs | |||||||||
| Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | |
| Black | 0.74 | 0.06 | 0.26 | 0.06 | 0.77 | 0.04 | 0.23 | 0.04 | 0.69 | 0.06 | 0.31 | 0.06 | 0.58 | 0.07 | 0.42 | 0.07 |
| European | 0.73 | 0.05 | 0.27 | 0.05 | 0.81 | 0.03 | 0.19 | 0.03 | 0.82 | 0.06 | 0.18 | 0.06 | 0.64 | 0.13 | 0.36 | 0.13 |
| East Asian | 0.68 | 0.08 | 0.32 | 0.08 | 0.80 | 0.04 | 0.20 | 0.04 | 0.69 | 0.05 | 0.31 | 0.05 | 0.66 | 0.09 | 0.34 | 0.09 |
| South Asian | 0.71 | 0.06 | 0.29 | g | 0.68 | 0.03 | 0.32 | 0.03 | 0.76 | 0.05 | 0.24 | 0.05 | 0.68 | 0.06 | 0.32 | 0.06 |
|
| ||||||||||||||||
| FOILS | ||||||||||||||||
|
| ||||||||||||||||
| CRs | FAs | CRs | FAs | CRs | FAs | CRs | FAs | |||||||||
| Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | |
| Black | 0.56 | 0.04 | 0.44 | 0.04 | 0.61 | 0.03 | 0.39 | 0.03 | 0.58 | 0.06 | 0.42 | 0.06 | 0.52 | 0.06 | 0.48 | 0.06 |
| European | 0.50 | 0.06 | 0.50 | 0.06 | 0.56 | 0.03 | 0.44 | 0.03 | 0.66 | 0.06 | 0.34 | 0.06 | 0.55 | 0.05 | 0.45 | 0.05 |
| East Asian | 0.44 | 0.05 | 0.56 | 0.05 | 0.51 | 0.04 | 0.49 | 0.04 | 0.55 | 0.06 | 0.45 | 0.06 | 0.47 | 0.02 | 0.53 | 0.02 |
| South Asian | 0.48 | 0.05 | 0.52 | 0.05 | 0.51 | 0.04 | 0.49 | 0.04 | 0.65 | 0.04 | 0.35 | 0.04 | 0.59 | 0.06 | 0.41 | 0.06 |
Fig 2. Results from the Test phase across all participants.
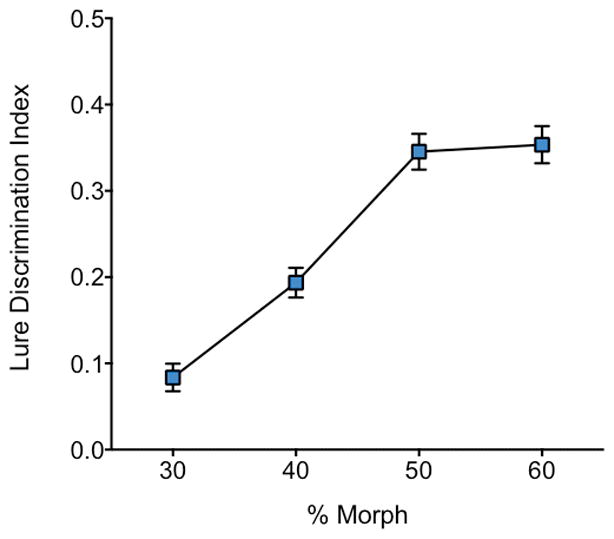
Lure correct rejection Tbias-corrected Lure Discrimination IndexT improved as face morphs got increasingly dissimilar.
We then examined the possibility that performance would be modulated by the ethnic origin of the face stimuli by conducting a three-way repeated measures ANOVA with interference level and stimulus ethnicity as within subject factors, and participant ethnicity as a between subject factor. We observed a significant main effect of interference as before [(F(3,477) = 72.1, P<.0001] as well as a significant interference by stimulus ethnicity interaction [F(9,477) = 2.715, P<.05]. The three-way interaction was not significant (P=.28).
After receiving feedback from participants, we identified a potential pitfall in one of our stimulus categories. During testing, many of our participants of Middle Eastern and Indian descent felt that they did not identify with any of the categories and that while the South Asian category was close, did not fully reflect their background. Due to the ambiguity of this particular stimulus set, we chose to limit our remaining analysis to the more robust stimulus sets (Black, Caucasian, and East Asian).
In order to query the ORE in sufficient detail, we decided to limit our analyses to the race group with the highest sample size, as the number of subjects in other race groups was not comparable. Thus, for the remaining analyses, we only used Caucasian individuals (n=29).
We subsequently conducted a repeated measures ANOVA with interference (four levels) and face stimulus ethnicity (three levels) only in Caucasian subjects and observed a main effect of interference level [F(3,84) = 4.54, P<.05] and ethnicity [F(2,56) = 4.0, P<.05] as well as a significant interference by ethnicity interaction [F(6,168) = 24.77, P<.0001] (Fig 3a). The effect size of this interaction was moderate (ηp2 = .47).
Fig 3.
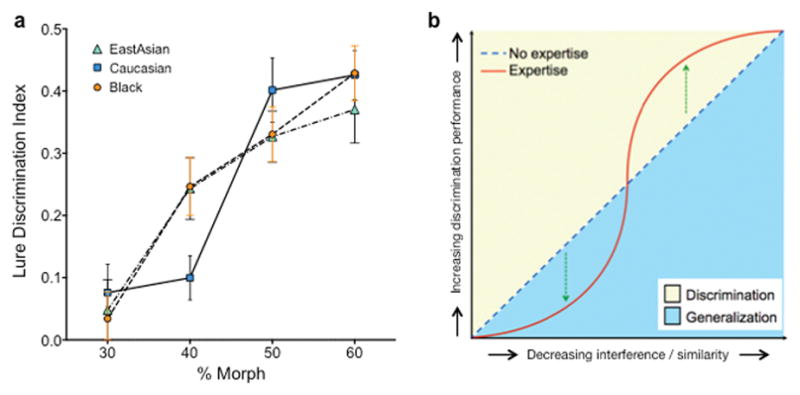
(a) Results for Caucasian participants categorized by race of face stimuli. Black and East Asian faces were better fit with linear functions, while Caucasian faces were significantly better fit with a sigmoid than linear. (b) A conceptual model of the effects of expertise on discrimination performance. For faces with which perceivers have expertise, discrimination and generalization performance should be more tuned, thus yielding a sigmoidal pattern. For faces with which perceives do not have expertise, discrimination performance should increase linearly with decreasing interference.
In order to examine the tuning curves for each stimulus ethnicity we attempted to fit both a linear curve and a sigmoidal curve with variable slope to each of the stimulus categories using ordinary least squares (OLS). The procedure used nonlinear regression which aimed to find the values of the parameters that make the curve come as close as possible to the data via minimizing the sum of the squares of the vertical distances of the points from the curve.
For Black faces, only the linear fit was possible, which yielded an R2 of .26. For East Asian faces, only the linear fit was possible, which yielded an R2 of .17. For the Caucasian faces, the linear fit had an R2 of .287 whereas the sigmoidal had an R2 of .336. We compared the two fits using the Extra sum-of-squares F-test. The null hypothesis was that the linear fit was correct. The improvement of the sigmoidal model was quantified as the difference in sum-of-squares. The difference in fit was significant (F(2,112) = 4.09, P=.019), suggesting that performance on Caucasian faces were better fit with a sigmoidal S-shaped function, as we predicted (Fig 3b).
Discussion
This study sought to examine how perceptual expertise may influence mnemonic discrimination of same- and other-race faces. Results during a face discrimination task suggest that when subjects are shown face lures from within their own race, their ability to discriminate these faces from previously viewed faces increases sharply at a threshold between 40 and 50% dissimilarity. In contrast, when viewing other-race faces, the same subjects’ ability to perform this discrimination process increases linearly as a function of dissimilarity. These distinct “tuning curves” suggest that judgments are not simply a function of interference but also vary as a function of expertise.
While our results suggest that mnemonic discrimination/generalization and the ability to reduce interference may mechanistically contribute to the ORE, it is a complex phenomenon that may also be driven by perceptual, motivational, social, and cultural factors (see Young et al. 2012 for a recent review). Based on our study, it is possible to see that some of these factors may have strong effects on mnemonic processes. We have only touched upon the effects of one proposed model on mnemonic discrimination. It is our hope that more studies will examine the effects of holistic processing and social categorization on mnemonic discrimination both behaviorally and neurally.
A potential concern is whether our results could be influenced by the amount of cultural exposure to other races during childhood and adolescent years. After the test session, all subjects completed self-report questionnaires that extensively assessed their experience with other races. Participants also indicated which racial category (based on the FaceGen categories) they most identified with. Despite the heterogeneity of modern societies and the prevalence of cross-cultural expertise, all subjects demonstrated more prevalent interactions and experiences within race than outside of race. Thus, we feel confident that our results are not confounded by inconsistent racial expertise.
These results raise a major question: what are the neural mechanisms that may underlie such mnemonic expertise with face stimuli? There are likely multiple mechanisms that span across sensory and perceptual processing, memory processing, as well as executive function. We propose that the hippocampus plays an important role in this ability, given its role in resolving mnemonic interference through pattern separation and pattern completion (potential neural substrates for discrimination and generalization behavior). However, the hippocampus by itself cannot give rise to such a complex phenomenon. Contributions of visual, attentional, and perceptual factors are necessary, as well as prior knowledge that accumulates over time building up such expertise. These effects are particularly important when considering proposed mechanisms such as holistic processing, social and motivational grouping, and perceptual expertise, all of which would have a potential influence on the quality and strength of information flow in and out of the hippocampus. For example, there is a large body of literature that has implicated the fusiform gyrus in facial recognition (Haxby et al. 1995; Sergent et al. 1992; Puce et al. 1995; Kanwisher et al. 1997; Golby et al. 2001; Rotshtein et al. 2005). More recent work has explored the contributions of a much more diverse set of regions involved in perceptual processing. Notably, an fMRI study by Bavel and colleagues (2008) showed that intergroup perception yielded greater activity in the amygdala, fusiform gyri, orbitofrontal cortex, and dorsal striatum. Future studies examining the neural basis of the ORE will need to bridge between perceptual and mnemonic processing networks.
One of the most important questions prompted by this work is whether a linear un-tuned discrimination/generalization function can be converted to a tuned s-shaped function with enough practice and exposure. Because our study was cross-sectional and not longitudinal we are unable to answer this question directly, but we believe that future studies should examine this possibility in detail. The idea of “expertise training” is not new by any means. This idea has been shown to be successful in the laboratory across multiple instances and racial groups (Elliot, Wills, & Goldstein, 1973). Training techniques have been used in the past to enhance face recognition expertise. For example, participants have been trained to individuate faces (Lebrecht et al. 2009). Other work has shown that perceivers can be trained to individuate rather than categorize faces (Hugenberg et al. 2007). Finally, other work has also shown that one can familiarize perceivers with other race faces through fostering holistic face processing practices (McKone et al. 2007). However, these studies did not specifically target mnemonic interference, and using an interference paradigm similar to ours may provide a more targeted alternative than simple exposure training. These questions will be very important to address in future work.
Acknowledgments
We thank Stephanie Leal, Zachariah Reagh, Jared Roberts, Craig Stark, Rebecca Stevenson, and Katherine Cameron for conceptual feedback and comments on earlier versions of this manuscript. This research was supported by NIH grants R01 MH102392, and P50 AG16573.
References
- Angeli A, Davidoff J, Valentine T. Face familiarity, distinctiveness, and categorical perception. The Quarterly Journal of Experimental Psychology. 2008;61(5):690–707. doi: 10.1080/17470210701399305. [DOI] [PubMed] [Google Scholar]
- Behrman BW, Davey SL. Eyewitness identification in actual criminal cases: an archival analysis. Law and human behavior. 2001;25(5):475. doi: 10.1023/a:1012840831846. [DOI] [PubMed] [Google Scholar]
- Bernstein MJ, Young SG, Hugenberg K. The cross-category effect mere social categorization is sufficient to elicit an own-group bias in face recognition. Psychological Science. 2007;18(8):706–712. doi: 10.1111/j.1467-9280.2007.01964.x. [DOI] [PubMed] [Google Scholar]
- Blais C, Jack RE, Scheepers C, Fiset D, Caldara R. Culture shapes how we look at faces. PLoS One. 2008;3(8):e3022. doi: 10.1371/journal.pone.0003022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bothwell RK, Brigham JC, Malpass RS. Cross-racial identification. Personality and Social Psychology Bulletin. 1989;15(1):19–25. [Google Scholar]
- Brainard DH. The psychophysics toolbox. Spatial vision. 1997;10:433–436. [PubMed] [Google Scholar]
- Elliott ES, Wills EJ, Goldstein AG. The effects of discrimination training on the recognition of white and oriental faces. Bulletin of the Psychonomic Society. 1973;2(2):71–73. [Google Scholar]
- Feingold GA. Influence of environment on identification of persons and things. J Am Inst Crim L & Criminology. 1914;5:39. [Google Scholar]
- Goffaux V, Rossion B. Faces are” spatial”--holistic face perception is supported by low spatial frequencies. Journal of Experimental Psychology: Human Perception and Performance. 2006;32(4):1023. doi: 10.1037/0096-1523.32.4.1023. [DOI] [PubMed] [Google Scholar]
- Golby AJ, Gabrieli JD, Chiao JY, Eberhardt JL. Differential responses in the fusiform region to same-race and other-race faces. Nature neuroscience. 2001;4(8):845–850. doi: 10.1038/90565. [DOI] [PubMed] [Google Scholar]
- Haxby JV, Horwitz B, Ungerleider LG, Maisog JM, Pietrini P, Grady CL. The functional organization of human extrastriate cortex: a PET-rCBF study of selective attention to faces and locations. Journal of Neuroscience. 1994;14(11):6336–6353. doi: 10.1523/JNEUROSCI.14-11-06336.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hugenberg K, Young SG, Bernstein MJ, Sacco DF. The categorization-individuation model: an integrative account of the other-race recognition deficit. Psychological review. 2010;117(4):1168. doi: 10.1037/a0020463. [DOI] [PubMed] [Google Scholar]
- Hugenberg K, Miller J, Claypool HM. Categorization and individuation in the cross-race recognition deficit: Toward a solution to an insidious problem. Journal of Experimental Social Psychology. 2007;43(2):334–340. [Google Scholar]
- Hugenberg K, Corneille O. Holistic Processing Is Tuned for In-Group Faces. Cognitive Science. 2009;33(6):1173–1181. doi: 10.1111/j.1551-6709.2009.01048.x. [DOI] [PubMed] [Google Scholar]
- Hugenberg K, Sacco DF. Social categorization and stereotyping: How social categorization biases person perception and face memory. Social and Personality Psychology Compass. 2008;2(2):1052–1072. [Google Scholar]
- Jacques C, Rossion B. The speed of individual face categorization. Psychological Science. 2006;17(6):485–492. doi: 10.1111/j.1467-9280.2006.01733.x. [DOI] [PubMed] [Google Scholar]
- Kanwisher N, McDermott J, Chun MM. The fusiform face area: a module in human extrastriate cortex specialized for face perception. The Journal of Neuroscience. 1997;17(11):4302–4311. doi: 10.1523/JNEUROSCI.17-11-04302.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kelly DJ, Quinn PC, Slater AM, Lee K, Ge L, Pascalis O. The other-race effect develops during infancy evidence of perceptual narrowing. Psychological Science. 2007;18(12):1084–1089. doi: 10.1111/j.1467-9280.2007.02029.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lebrecht S, Pierce LJ, Tarr MJ, Tanaka JW. Perceptual other-race training reduces implicit racial bias. PLoS One. 2009;4(1):e4215. doi: 10.1371/journal.pone.0004215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marr D. Simple memory: a theory for archicortex. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 1971:23–81. doi: 10.1098/rstb.1971.0078. [DOI] [PubMed] [Google Scholar]
- Maurer D, Le Grand R, Mondloch CJ. The many faces of configural processing. Trends in cognitive sciences. 2002;6(6):255–260. doi: 10.1016/s1364-6613(02)01903-4. [DOI] [PubMed] [Google Scholar]
- McClelland JL, McNaughton BL, O’Reilly RC. Why there are complementary learning systems in the hippocampus and neocortex: insights from the successes and failures of connectionist models of learning and memory. Psychological review. 1995;102(3):419. doi: 10.1037/0033-295X.102.3.419. [DOI] [PubMed] [Google Scholar]
- McKone E, Brewer JL, MacPherson S, Rhodes G, Hayward WG. Familiar other-race faces show normal holistic processing and are robust to perceptual stress. Perception. 2007;36(2):224. doi: 10.1068/p5499. [DOI] [PubMed] [Google Scholar]
- Meissner CA, Brigham JC. Thirty years of investigating the own-race bias in memory for faces: A meta-analytic review. Psychology, Public Policy, and Law. 2001;7(1):3. [Google Scholar]
- Michel C, Corneille O, Rossion B. Race categorization modulates holistic face encoding. Cognitive Science. 2007;31(5):911–924. doi: 10.1080/03640210701530805. [DOI] [PubMed] [Google Scholar]
- Michel C, Rossion B, Han J, Chung CS, Caldara R. Holistic processing is finely tuned for faces of one’s own race. Psychological Science. 2006;17(7):608–615. doi: 10.1111/j.1467-9280.2006.01752.x. [DOI] [PubMed] [Google Scholar]
- O’Reilly RC, Norman KA. Hippocampal and neocortical contributions to memory: Advances in the complementary learning systems framework. Trends in cognitive sciences. 2002;6(12):505–510. doi: 10.1016/s1364-6613(02)02005-3. [DOI] [PubMed] [Google Scholar]
- O’toole AJ, Deffenbacher KA, Valentin D, Abdi H. Structural aspects of face recognition and the other-race effect. Memory & Cognition. 1994;22(2):208–224. doi: 10.3758/bf03208892. [DOI] [PubMed] [Google Scholar]
- Pelli DG. The VideoToolbox software for visual psychophysics: Transforming numbers into movies. Spatial vision. 1997;10(4):437–442. [PubMed] [Google Scholar]
- Puce A, Allison T, Gore JC, McCarthy G. Face-sensitive regions in human extrastriate cortex studied by functional MRI. Journal of neurophysiology. 1995;74:1192–1192. doi: 10.1152/jn.1995.74.3.1192. [DOI] [PubMed] [Google Scholar]
- Reagh ZM, Roberts JM, Ly M, DiProspero N, Murray E, Yassa MA. Spatial discrimination deficits as a function of mnemonic interference in aged adults with and without memory impairment. Hippocampus. 2014;24(3):303–314. doi: 10.1002/hipo.22224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rhodes G, Brake S, Atkinson AP. What’s lost in inverted faces? Cognition. 1993;47(1):25–57. doi: 10.1016/0010-0277(93)90061-y. [DOI] [PubMed] [Google Scholar]
- Roberts JM, Ly M, Murray E, Yassa MA. Temporal discrimination deficits as a function of lag interference in older adults. Hippocampus. 2014 doi: 10.1002/hipo.22303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rotshtein P, Henson RN, Treves A, Driver J, Dolan RJ. Morphing Marilyn into Maggie dissociates physical and identity face representations in the brain. Nature neuroscience. 2005;8(1):107–113. doi: 10.1038/nn1370. [DOI] [PubMed] [Google Scholar]
- Sangrigoli S, Pallier C, Argenti AM, Ventureyra VAG, De Schonen S. Reversibility of the other-race effect in face recognition during childhood. Psychological Science. 2005;16(6):440–444. doi: 10.1111/j.0956-7976.2005.01554.x. [DOI] [PubMed] [Google Scholar]
- Sergent J, Ohta S, MacDonald B. Functional neuroanatomy of face and object processing A positron emission tomography study. Brain. 1992;115(1):15–36. doi: 10.1093/brain/115.1.15. [DOI] [PubMed] [Google Scholar]
- Shapiro ML, Olton DS. Hippocampal function and interference. Memory systems. 1994;1994:141–146. [Google Scholar]
- Sporer SL. Recognizing faces of other ethnic groups: An integration of theories. Psychology, Public Policy, and Law. 2001;7(1):36. [Google Scholar]
- Squire LR. Memory and the hippocampus: a synthesis from findings with rats, monkeys, and humans. Psychological review. 1992;99(2):195. doi: 10.1037/0033-295x.99.2.195. [DOI] [PubMed] [Google Scholar]
- Tanaka JW. The entry point of face recognition: evidence for face expertise. Journal of Experimental Psychology: General. 2001;130(3):534. doi: 10.1037//0096-3445.130.3.534. [DOI] [PubMed] [Google Scholar]
- Tanaka JW, Farah MJ. Parts and wholes in face recognition. The Quarterly journal of experimental psychology. 1993;46(2):225–245. doi: 10.1080/14640749308401045. [DOI] [PubMed] [Google Scholar]
- Tanaka JW, Gauthier I. Expertise in object and face recognition. Psychology of learning and motivation. 1997;36:83–125. [Google Scholar]
- Tanaka JW, Kiefer M, Bukach CM. A holistic account of the own-race effect in face recognition: Evidence from a cross-cultural study. Cognition. 2004;93(1):B1–B9. doi: 10.1016/j.cognition.2003.09.011. [DOI] [PubMed] [Google Scholar]
- Taubert J, Apthorp D, Aagten-Murphy D, Alais D. Vision research. 11. Vol. 51. Chicago: 2011. The role of holistic processing in face perception: Evidence from the face inversion effect; pp. 1273–1278. [DOI] [PubMed] [Google Scholar]
- Treves A, Rolls ET. Computational analysis of the role of the hippocampus in memory. Hippocampus. 1994;4(3):374–391. doi: 10.1002/hipo.450040319. [DOI] [PubMed] [Google Scholar]
- Van Bavel JJ, Packer DJ, Cunningham WA. The neural substrates of in-group bias a functional magnetic resonance imaging investigation. Psychological Science. 2008;19(11):1131–1139. doi: 10.1111/j.1467-9280.2008.02214.x. [DOI] [PubMed] [Google Scholar]
- Walker PM, Hewstone M. A developmental investigation of other-race contact and the own-race face effect. British Journal of Developmental Psychology. 2006;24(3):451–463. [Google Scholar]
- Walker PM, Tanaka JW. An encoding advantage for own-race versus other-race faces. PERCEPTION-LONDON- 2003;32(9):1117–1126. doi: 10.1068/p5098. [DOI] [PubMed] [Google Scholar]
- Yassa MA, Lacy JW, Stark SM, Albert MS, Gallagher M, Stark CE. Pattern separation deficits associated with increased hippocampal CA3 and dentate gyrus activity in nondemented older adults. Hippocampus. 2011a;21(9):968–979. doi: 10.1002/hipo.20808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yassa MA, Stark CE. Pattern separation in the hippocampus. Trends in neurosciences. 2011b;34(10):515–525. doi: 10.1016/j.tins.2011.06.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yassa MA, Mattfeld AT, Stark SM, Stark CE. Age-related memory deficits linked to circuit-specific disruptions in the hippocampus. Proceedings of the National Academy of Sciences. 2011;108(21):8873–8878. doi: 10.1073/pnas.1101567108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yassa MA, Stark SM, Bakker A, Albert MS, Gallagher M, Stark CE. High-resolution structural and functional MRI of hippocampal CA3 and dentate gyrus in patients with amnestic Mild Cognitive Impairment. Neuroimage. 2010;51(3):1242–1252. doi: 10.1016/j.neuroimage.2010.03.040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yin RK. Looking at upside-down faces. Journal of experimental psychology. 1969;81(1):141. [Google Scholar]
- Young SG, Hugenberg K, Bernstein MJ, Sacco DF. Perception and Motivation in Face Recognition A Critical Review of Theories of the Cross-Race Effect. Personality and Social Psychology Review. 2012;16(2):116–142. doi: 10.1177/1088868311418987. [DOI] [PubMed] [Google Scholar]