

Abstract
As findings on the neuropathological and behavioral components of Alzheimer’s disease (AD) continue to accrue, converging evidence suggests that macroscale brain functional disruptions may mediate their association. Recent developments on theoretical neuroscience indicate that instantaneous patterns of brain connectivity and metastability may be a key mechanism in neural communication underlying cognitive performance. However, the potential significance of these patterns across the AD spectrum remains virtually unexplored. We assessed the clinical sensitivity of static and dynamic functional brain disruptions across the AD spectrum using resting-state fMRI in a sample consisting of AD patients (n = 80) and subjects with either mild (n = 44) or subjective (n = 26) cognitive impairment (MCI, SCI). Spatial maps constituting the nodes in the functional brain network and their associated time-series were estimated using spatial group independent component analysis and dual regression, and whole-brain oscillatory activity was analyzed both globally (metastability) and locally (static and dynamic connectivity). Instantaneous phase metrics showed functional coupling alterations in AD compared to MCI and SCI, both static (putamen, dorsal and default-mode) and dynamic (temporal, frontal-superior and default-mode), along with decreased global metastability. The results suggest that brains of AD patients display altered oscillatory patterns, in agreement with theoretical premises on cognitive dynamics.
Dementia is a highly prevalent syndrome with a large economic and societal impact, characterized by the progressive impairment of cognition, behavioral changes and reduced quality of life1. Alzheimer’s disease (AD), the most common form of dementia, has complex and heterogeneous etiopathogenic underpinnings, including both genetic and environmental factors2. While some clinical, cognitive and neuropathological components of AD are widely acknowledged1,3,4, a unified frame for the intricate architecture of AD is still needed in order to make sense of the pathways from molecular changes to clinical phenomenology, which is critical for early identification of at-risk individuals.
Converging evidence suggests that large-scale brain oscillatory activity disruptions, frequently assessed using resting-state functional magnetic resonance imaging (fMRI), may reflect key mechanisms bridging the gap between neuropathology and cognitive alterations in AD5. Virtually all the resting-state fMRI literature on oscillatory alterations in AD has focused on measures of coupling between different brain regions averaged across a scanning session –the so-called (static) functional connectivity. For example, AD patients have been shown to exhibit stronger hippocampal connectivity within the default mode network and weaker hippocampal-cingulate co-activation6. Furthermore, suggesting that the clinical manifestation may be mirrored in preclinical stages in individuals with high genetic risk, young and middle-aged carriers of the AD-associated APOE-epsilon4 allele have shown higher hippocampal synchronization within the default mode network7,8. However, results across studies are largely mixed, suggesting both increases and decreases in coupling associated with aging and AD9.
Importantly, recent conceptual, theoretical and methodological advances may shed light on these mixed findings; among them, the observation that time-varying coupling and de-coupling of brain regions embeds information that may be important in clinical settings10,11. This novel and biologically feasible framework has been conceptualized in the communication through coherence theory, which postulates that cognition is linked to neural communication emerging from coherent oscillatory activity, and that cognitive functions require a flexible set of signaling processes occurring on top of the anatomical backbone of the brain12,13,14. Namely, in order to fully benefit from the fixed anatomical connections, neural ensembles should traverse distinct dynamic states for adequate cognitive and other mental functions to arise15,16. Consequently, brain disorders typically implicating cognitive and emotional functions are expected to be characterized by a reduced dynamic repertoire17: a smaller set of brain functional configurations. Of note, the underlying neurobiological mechanisms can take place at different timescales, including the very slow frequencies measured non-invasively via fMRI15, and can be measured by analyzing variations in synchrony among regions over time18,19.
In this study, we aim to test this conjecture by assessing the potential clinical sensitivity of brain oscillatory characteristics across the AD spectrum in a sample of patients with AD, individuals with mild cognitive impairment (MCI), and those with only subjective memory complaints (also known as subjective cognitive impairment, SCI). Time-series from functional “seeds” defined with independent component analysis (ICA) were estimated using dual regression7, and potential alterations in brain oscillatory activity were analyzed using two methods: 1) the standard, averaged coupling between pairs of resting-state fMRI signals20, and 2) phase-based metrics of coupling and de-coupling across time: metastability (whole-brain dynamics) and pairwise synchrony changes (dynamics of paired brain regions)11. Phase-based metrics of coupling have conventionally been used in electroencephalography studies, in measures such as the phase-locking value21,22,23, and have only recently been adapted for fMRI signal analysis of time-varying connectivity10,11,24. To the best of our knowledge, no previous study has evaluated this type of phase-based dynamic alterations in fMRI activity in an aging and neurodegeneration context. Based on established knowledge about the anatomical distribution of the pathophysiology and resting-state fMRI alterations in AD, and also considering theoretical concepts on the relevance of a wide set of time-varying connectivity, we hypothesized that patients with AD exhibit altered sFC and dFC, both at the whole-brain level and at specific brain nodes implicating temporal, parietal and subcortical structures, reflecting a graded pattern of differences corresponding with disease severity.
Results
Demographic and clinical data
The demographic data summarized in Table 1 and Fig. 1 shows statistically significant differences in the distribution of age, years of education and MMSE scores across the different diagnostic categories. Briefly, patients with AD (71.5 ± 7.9 years) were older than the MCI (63.5 ± 11.1) and SCI (63.6 ± 9.6) groups. Also, patients with AD had lower MMSE scores (22.8 ± 4.8) than MCI (28.2 ± 2.1) and SCI (29.4 ± 0.6), and shorter education. There were no differences in sex distribution between groups. The correlation coefficients between age, MMSE and education indicate small/medium effect sizes25, which justifies their inclusion as covariates without strong collinearity issues.
Table 1. Clinical and demographic information.
| Age |
MMSE score |
Years of education |
Sample size (n) |
|||||
|---|---|---|---|---|---|---|---|---|
| Mean (SD) | Range | Mean (SD) | Range | Mean (SD) | Range | male/female | ||
| AD | 71.5 (7.9) | 53–91 | 22.8 (4.8) | 5–30 | 12.4 (3.1) | 7–19 | 35/45 | |
| MCI | 63.5 (11.1) | 42–85 | 28.2 (2.1) | 20–30 | 14.3 (2.8) | 9–20 | 26/18 | |
| SCI | 63.6 (9.6) | 49–84 | 29.4 (0.6) | 28–30 | 15 (3.3) | 9–22 | 12/14 | |
| Between-group differences (test statistics) | ||||||||
| t | p | t | p | t | p | Χ2 | p | |
| ADvsMCI | 4.6 | 9.5e-6* | −6.6 | 1.1e-9* | −3.3 | 0.001* | 2.1 | 0.148 |
| ADvsSCI | 4.2 | 6.7e-5* | −7 | 2.2e-10* | −3.6 | 4.5e-4* | 1.7e-4 | 0.989 |
| MCIvsSCI | −0.04 | 0.968 | −2.9 | 0.005* | −0.9 | 0.351 | 0.6 | 0.423 |
Notes: Between-group differences were assessed with either two-tailed t-tests (continuous variables) or chi-square (Χ2, contingency tables degree of freedom). Abbreviations: SD, standard deviation; *statistically-significant p-value.
Figure 1. Correlation matrix chart of demographic features.
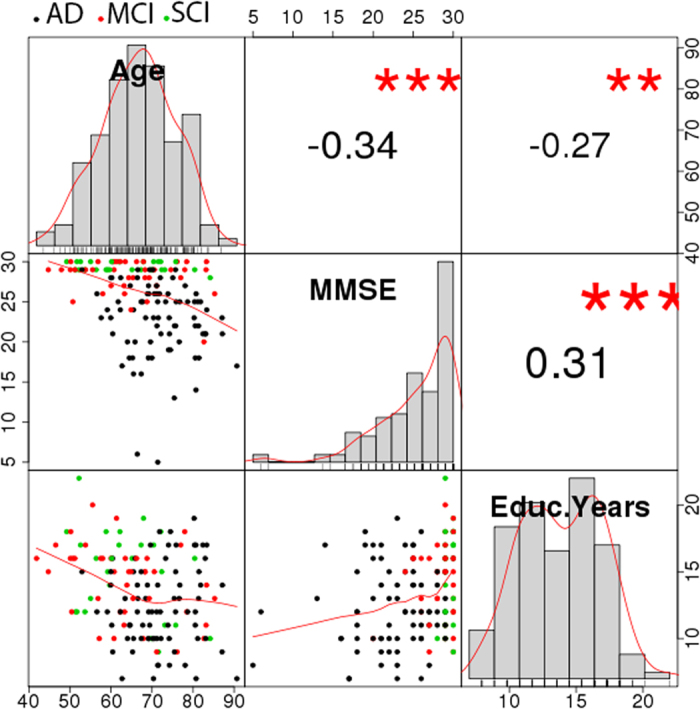
Entries in the upper triangle correspond to Pearson’s correlation coefficients, and the number of “*” represents the significance level (**p < 10−2, ***p < 10−3).
Statistical analysis of average coupling (sFC)
Figure 2 (top) displays regression coefficients and unadjusted p-values obtained from logistic regression analysis of diagnosis pairs (ADvsMCI, ADvsSCI and MCIvsSCI), using each of the 325 partial correlation coefficients as independent variable, and controlling for gender, age, headcoil and motion. Network-based statistics (NBS) analysis using data from all participants showed a 39-edge subnetwork with diagnose-dependent disruptions (p = 0.037) (Fig. 3, uppermost heatmap). As shown by the sum of F-statistics in Fig. 3 (bottom), the brain nodes with the largest contributions to the statistical significance of that subnetwork were IC13 (putamen), IC14 (dorsal), IC11 (default-mode) and IC6 (default-mode).
Figure 2. Logistic regression results for sFC and dFC at the 325 IC pairs.

The results represent raw p-values were obtained from logistic regression models adjusting by gender, age, in-scanner motion and headcoil. The horizontal red line indicates the p-value threshold (0.05) in the corresponding logarithmic scale. Colorbars display regression coefficients for the different pairs.
Figure 3. IC pairs in the statistically significant sub-networks (sFC and dFC) from NBS.
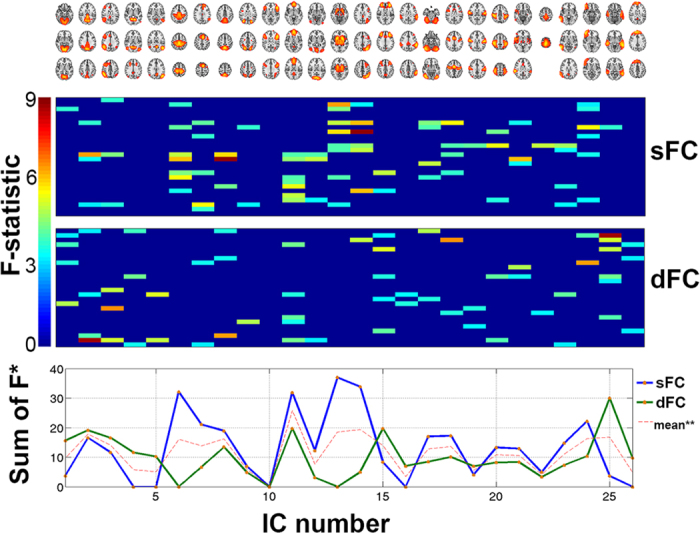
Heatmap matrices: the entries represent F-statistics of the 28-edge network with NBS significance. Non-significant entries were set to 0. Notes: *sum of F-statistics from the statistically significant results, illustrating the relative importance of each IC; **average of “sum of F” over dFC and sFC.
Figure 4 (top) shows the partial correlation coefficients (z-transformed) of the 39 edges constituting the mentioned network. Notice that the plot contains unadjusted data, which are not directly comparable to the F-statistics derived from NBS multivariate tests; visual analysis of the boxplot notches (namely, non-overlapping notches as evidence of median differences26) indicates that the largest between-group differences are in the following IC pairs: 36 (IC2-13: 0 < AD < MCI = SCI), 116 (IC6-7: 0 < MCI = SCI < AD), 137 (IC7-9: AD < MCI = SCI < 0), 160 (IC8-14: AD < MCI = SCI = 0), 166 (IC8-20: AD > MCI = SCI > 0), 207 (IC11-13: 0 < AD < MCI = SCI), 214 (IC11-20: AD = SCI < 0 < MCI), 216 (IC11-22: 0 < MCI < AD = SCI), 217 (IC11-23: 0 < MCI < AD = SCI), 222 (IC12-14: AD < MCI < SCI < 0), 239 (IC13-18: 0 = AD < MCI = SCI), 254 (IC14-21: 0 < AD = SCI < MCI), 261 (IC15-17: 0 < MCI < AD = SCI), 265 (IC15-21: 0 < AD = SCI < MCI), 281 (IC17-18: 0 < MCI < AD = SCI) and 307 (IC20-23: 0 < MCI < AD = SCI). Notice that, from the previous list, IC pairs 137, 222, 261, 281 and 307 show, on average, large z-transformed correlations (absolute value > 1), and all other IC pair coefficients were reduced via regularized partial correlation, as explained in Methods. These results indicate the presence of both increases and decreases in functional coupling which could discriminate between diagnostic categories.
Figure 4. Raw sFC and dFC values at the IC pairs with significance from the NBS tests.

The trends shown here corresponds to the direct measures of pairwise coupling (see Methods), whereas the test results discussed in the text show that the association between diagnostic status and dFC persists even after adjustments for gender, age, and headcoil. Notes: *coefficient of variation (standard deviation divided by mean) of the normalized phase differences (see Methods); **z-transformed value of the regularized regression coefficients.
Statistical analysis of time-varying connectivity (dFC)
The 325 dFC measures were largely uncorrelated with each other, and showed only weak correlations with their sFC counterparts (Figure S1), further justifying their inclusion as alternative phenotypes. Coefficients and p-values of the different IC pairs, derived from logistic regression tests over pairs of diagnoses, are shown in Fig. 2 (bottom). While most of the test statistics at p < 0.05 indicate decreased dFC with increased disease severity (negative-valued coefficients depicted as dark blue dots), none of these results survives a stringent Bonferroni correction. However, after multiple-testing adjustments using NBS on the three-diagnose data, there was a 30-edge dFC sub-network with alterations related to disease severity (p = 0.018) (Fig. 3, lowermost heatmap). The brain regions with the largest contributions to the statistical significance were related to the temporal/auditory (IC 25), frontal (IC 15) and default-mode (IC2 and IC11) areas.
The dFC values (coefficients of variation) at the 28 altered edges, grouped by diagnosis, are shown in Fig. 4 (bottom). As mentioned for the case of sFC, Fig. 4 displays unadjusted data, which are not directly comparable to the F-statistics derived from NBS multivariate tests; nevertheless, the different distributions of dFC values depending on diagnosis can be appreciated. Visual analysis of boxplot notches26 suggests that the strongest between-group dissimilarities observed in the raw data indicates localized decreases in dFC with increasing disease severity. More specifically, the following IC pairs showed that pattern: 1 (IC1-2: SCI < AD < MCI), 16 (IC1-17: SCI < MCI < AD), 48 (IC2-25: SCI = MCI < AD), 60 (IC3-14: SCI < MCI < AD), 64 (IC3-18: SCI < MCI < AD), 71 (IC3-25: SCI = MCI < AD), 79 (IC4-11: AD < MCI = SCI), 94 (IC4-26: AD < MCI < SCI), 136 (IC7-8: SCI < MCI = AD), 170 (IC8-24: SCI = MCI < AD), 184 (IC9-21: SCI = MCI < AD), 214 (IC11-20: SCI = AD < MCI), 233 (IC12-25: SCI = MCI < AD), 267 (IC15-23: MCI < SCI = AD), 271 (IC16-17: MCI < SCI = AD), 309 (IC20-25: AD < MCI = SCI) and 322 (IC23-26: AD = MCI < SCI). These results thus suggest that different patterns (both increases and decreases) of localized dFC may be related to diagnose.
The analysis of surrogate data showed that, in the present setting, the probabilities of detecting weak dFC are small. As shown in Fig. 5, in the whole dataset, the detection probabilities are above chance level only in the presence of strong dFC. For the specific case of the connections with (clinical) group differences, the strongest evidence for temporal dynamics at pairs 300 (IC19-22, p = 0.038), 60 (IC3-14, p = 0.062), 215 (IC11-21, p = 0.078) and 1 (IC1-2, p = 0.092) (Figs 5 and 6), suggesting that the current approach might have moderate power to detect weak dFC at such edges. As inferred from the data (Fig. 6), none of those results would be statistically significant when applying multiple testing adjustments; however, such corrections may be overly conservative here since the assessed edges come from another analysis, which already implemented a network-based multiple testing protocol. Since the evidence of true temporal dynamics observed here is only moderate, it should be noted that the group differences (AD/MCI/SCI) might have been influenced by non-dynamic properties of the time series.
Figure 5. Surrogate data test results at the whole-brain level and at edges with group differences.

Left: test of the sampled dFC values (sum of 325 edge weights) against a range of population means. Right: analysis of the dFC empirical values against their surrogate null distributions in the SCI group. The plot shows only the four edges with both statistically significant (clinical) group differences and p < 0.1 in the surrogate tests. Numbers next to the dashed line legends correspond to IC pairs. Additional details on the test procedures are described in Results.
Figure 6. Results of surrogate data tests for dFC in the SCI group.

Uppermost heatmap: T statistic maps from one-group T-tests (null hypothesis: dFC population mean = 0 at every edge). Only the original data was used for that plot. The upper diagonal shows only connections with statistically significant differences depending on clinical diagnose. Lowermost heatmap: using 10000 surrogate datasets, T* values were estimated, and 1 minus the proportion of times with T (original) > T* (surrogate) was considered the p-value for the presence of true dynamics. Note that the colormap corresponds to 1-p (lower p-values have warmer colors). In both heatmaps, the upper diagonal shows only connections with statistically significant differences depending on clinical diagnose. Bottom: sum of (empirical) T-statistic for each IC.
Time-varying synchronization at the whole-brain level: metastability
After adjusting for potential confounders, whole-brain metastability levels were higher in the healthier subgroup (SCI), and progressively decreased through MCI and AD (multinomial logit with SCI as reference group: βMCI = −25, SEMCI = 9.1, pMCI = 0.006; βAD = −29.1, SEAD = 8.9, pAD = 0.001; pseudo-R2 = 0.154, log-likelihood = −126.8, Fig. 7). Of note, neither the raw motion estimates obtained with MCFLIRT nor the variance kept after filtering were related to that metastability metric, and there was no evident movement*diagnose interaction associated with metastability (Figures S2 and S3). There was no association between metastability and tSNR, neither before nor after running FIX (linear regression metastability~tSNR: before FIX: β = −1.4 * 10−4, SE = 1.1 * 10−4, p = 0.217; after FIX: β = −1.7 * 10−4, SE = 1.1 * 10−4, p = 0.223; Figure S2). Similarly, there was no significant interaction with diagnosis (linear regression metastability~tSNR*diagnosis, with AD as reference group: before FIX: βMCI = 6 * 10−5, SEMCI = 2.6 * 10−4, pMCI = 0.817, βSCI = −2.2 * 10−4, SESCI = 3.2 * 10−4, pSCI = 0.495; after fix: βMCI = 1.6 * 10−4, SEMCI = 3.1 * 10−4, pMCI = 0.603, βSCI = 9.6 * 10−5, SESCI = 4.7 * 10−4, pSCI = 0.838; Figure S3).
Figure 7. Association between metastability and diagnostic status.
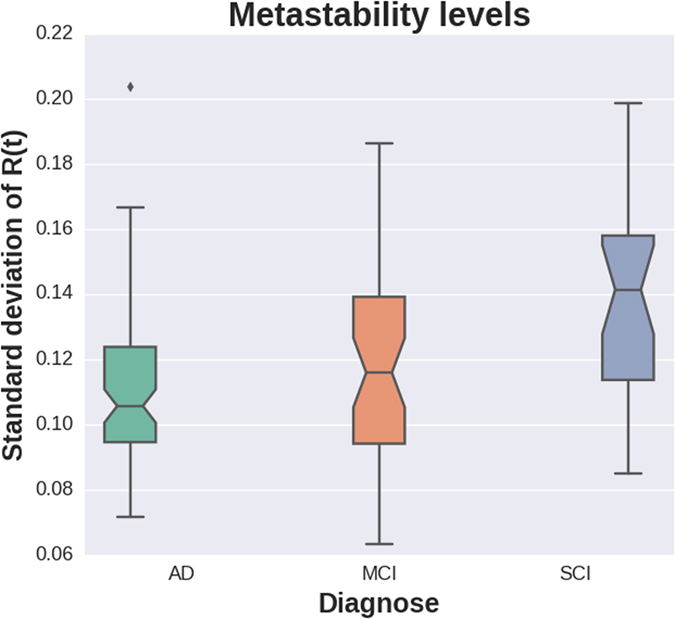
Note: The trend shown here corresponds to the direct measures of metastability, whereas the test results discussed in the text show that the association between diagnostic status and metastability persists even after adjustments for gender, age and headcoil.
Discussion
The present study evaluated the potential association between fMRI based neural oscillatory coupling disruptions and clinical severity across a phenotypical continuum related to cognitive status (AD, MCI and SCI). Two different types of between-region oscillatory coupling were analyzed: the average across the scanning session (static, sFC), defined as the regularized partial temporal correlation between pairs of nodes, and between-region oscillatory coupling was measured as the instantaneous transitions between synchronization and de-synchronization (dynamic, dFC), defined as the coefficient of variation of the phase differences over time. Distinct group differences in sFC and dFC were observed. Additionally, a statistically significant pattern of whole-brain metastability alterations was observed, with AD showing lower values than MCI, and MCI displaying lower values than SCI.
The findings on static coupling data (sFC) indicate between-group differences in coupling patterns mainly implicating the putamen, dorsal and default-mode areas, indicative of either hyper- or hypo-synchronization at specific regions depending on diagnosis. This is in line with previous data on the anatomical location of neurodegenerative and AD-related changes, and also resembles the accumulated literature findings of both increases and decreases in functional connectivity associated with aging and AD27.
To our knowledge, this is the first report on metastability and related phase coupling alterations (dFC) measured with fMRI in AD-spectrum phenotypes. The results agree with recent theories on brain computation, which posit metastability as the optimal state of neural activity at rest: a dynamical regime characterized by rapid, flexible, engaging and disengaging neural ensembles16,28,29,30. Within that framework, higher metastability at rest is thought to underlie optimal cognitive-behavioral function28,31; the findings here suggest that metastability gradually decreases from the healthier (SCI) to partly affected (MCI) and more severely altered cognitive states (AD). It is also worth noting that the dynamic differences measured globally (whole-brain metastability) were accompanied by localized disruptions in dFC, implicating a wide range of brain network nodes and their connections, suggesting a key role for temporal, frontal and default-mode areas in AD dFC.
The relevance of the regions and brain networks found using sFC and dFC (including default-mode, frontal, putamen, dorsal and temporal areas) has been highlighted in former research on brain changes throughout the AD continuum, such as functional alterations detected with resting-state fMRI32,33,34,35, volumetric atrophy observed with MRI36,37, and amyloid neuropathological disturbances measured with positron emission tomography38. Jointly, these results expand on a system-wide view of AD dementia5,39, by endorsing that the anatomically-constrained AD neuropathology may be linked to some localized macro-scale brain function alterations ultimately leading to the clinical phenotypes.
Two behavioral fMRI reports on humans deserve discussion in light of the present dFC results. In perhaps the most similar study to this one, Hellyer et al.28 investigated cognition and metastable dynamics in a sample of 89 participants (either healthy or with traumatic brain injury); they found that reduced metastability (as measured with resting-state fMRI) was associated with structural connectivity damage, reduced cognitive flexibility and disrupted information processing. Added to the previous findings, this may suggest that altered resting-state dynamics are a landmark of neurocognitive alterations across distinct clinical phenotypes. Furthermore, in a study using sliding-window analysis, Sourty et al.40 reported that some brain function alterations in Lewy body dementia can be detected using dFC but not static functional connectivity. Importantly, measuring dFC might have been troublesome in that report due to the low statistical detection power of single-session sliding-window correlations18. Novel methodologies, such as the phase-synchrony approaches adopted in this study10,24, might help overcome those issues. Here, they allow expanding on the previous clinical findings in dementia with Lewy bodies40 by suggesting that dFC alterations can be found in another form of dementia (AD) when measuring phase differences.
Finally, it is worth highlighting that the observed metastability decreases in AD might be related to a shrinkage of the dynamic repertoire, which constitutes a candidate physiological mechanism to connect classical neuropathological and clinical observations, and can be interpreted in that context. For instance, neuropathological findings have shown that aging, AD and amyloid alterations are associated with key myelin aberrations41,42,43,44. In parallel, research combining theoretical neuroscience and resting-state fMRI measurements experiments indicates that the dynamic patterns of the brain are constrained in cases of degradation of myelination, due to a restricted capacity of the neural ensembles to change its spatial configuration (transmission speed restrictions)17. Namely, micro-scale alterations of axonal conduction velocities –commonly due to myelin pathology– are thought to underlie macro-scale dFC reductions observed with fMRI. Additionally, at the psycho(patho)logical level, the efficient dynamical processing (over time) of neural information by segregating and integrating information is thought to sustain flexible cognitive states16,31,45,46, making fMRI-based metastability and dFC decreases feasible candidate indices of neurocognitive pathology. The metastability-related alterations observed in patients with AD fit well into biological and clinical frameworks, to suggest that large-scale brain oscillatory disruptions may link micro-scale neural alterations with disrupted cognitive architectures reported from the clinical phenomenology, which may eventually inform new therapy development using pathoconnectomics47.
Some limitations should be noted. First, the moderate sample size could have restricted the power to detect some effects. However, the cases where statistically significant outcomes were detected may indicate the presence of relatively large population effect sizes. Secondly, potential clinical heterogeneities across participants might affect the consistency of the findings: different subtypes of SCI and MCI may be associated with considerable differences in risk for dementia48,49, and probably also with distinct brain characteristics (i.e., Nobili et al.50). Robust neuroimaging findings from new studies may help address this issue; also, data on genetic and neuropathological markers might allow better diagnostic classifications. Additional studies might take advantage of those markers, which may be better suited for study within larger samples that allow improved phenotypic (diagnostic) stratification. Furthermore, some clinical data indicates that SCI and MCI could be relatively close transitional states preceding AD51,52,53, which may be reflected as similar brain dynamics in the two diagnoses. Although there were robust statistical associations, the lack of a healthy control group that had not been referred from the memory clinic may have limited the power to detect some MCI- and AD-specific effects. Even though the category of SCI is still controversial and may lack clinical relevance, some reports indicate that it could be associated with minor brain changes54. Hence, future studies may benefit from the inclusion of healthy controls without cognitive complaints to improve specificity, and prospective longitudinal designs are needed to assess the predictive value of the sFC, dFC and metastability indices in a clinical dementia context. Besides, even though stringent image pre-processing methods were applied, the influence of artifacts cannot be completely ruled out. Moreover, since the relationship between neural responses and blood-oxygen-level dependent fMRI signals has not been fully elicited55,56, and also considering the issues associated with causation analysis in fMRI57, complementary experiments are needed to explain the ultimate pathological mechanisms behind the observed patterns. Finally, as discussed by Glerean et al.24, the phase synchrony methods used to estimate dFC have some limitations: for instance, the current results correspond to the chosen narrowband (0.04–0.07 Hz), but other frequency domains could contain complementary information; also, the amplitude envelope is not used, even though it might complement the analyzed phase metrics. It is also worth mentioning that the ongoing dFC approach offered moderate evidence of temporal dynamics within the dataset considered here. This observation emphasizes the importance of conducting surrogate data tests –or equivalent– in order to verify the presence of dynamics in fMRI signals. That being said, having detected a different set of relevant alterations in AD when using sFC and the current dFC metrics suggests that they might inform on complementary pathophysiological mechanisms.
Overall, the results reported here constitute empirical evidence on relatively unexplored neural activity disruptions through different cognitive impairment phenotypes related to neurodegeneration, which are in line with previous clinical and neuropathological observations of an AD continuum52,54,58,59,60. Notwithstanding the limitations of this work, the observed metastability, sFC and dFC patterns are in agreement with the literature31 and deserve further attention across various cognitive and clinical phenotypes.
Methods
Ethics statement
The Regional Committee for Medical Research Ethics in South-Eastern Norway approved the study. Patients were only enrolled if determined to have capacity for consent by the evaluating physician. All participants gave written informed consent. All procedures were conducted in accordance with the Helsinki Declaration.
Participants
Cross-sectional patient data were obtained from the “Norwegian registry for persons being evaluated for cognitive symptoms in specialized care (NorCog)”. NorCog is a national patient registry comprising consecutively enrolled patients referred to one of 27 participating memory outpatient clinics because of suspected cognitive impairment or dementia.
All patients in the present study were recruited from one of the NorCog centers, the memory clinic at Oslo University Hospital, between 2010 and 2014. Patients were assessed in accordance with a standardized examination protocol61. Patients at the Oslo University Hospital memory clinic are usually referred to MRI of the brain as part of the diagnostic workup. Between 2010 and 2014 a limited capacity research protocol MRI was available. Selection of referral to this alternative was only based on accepted waiting time. The patients were diagnosed by two doctors in consensus (KE/AB or MLB/KP), based on all available information from the extensive clinical examination. Only patients with AD (n = 80, 71 ± 8 years, 35 males) according to the ICD-10 criteria62, patients with MCI (n = 44, 64 ± 11 years, 26 males) according to Winblad criteria63, and patients with a subjective cognitive complain that did not fulfill the AD or MCI criteria (SCI, n = 26, 64 ± 11 years, 26 males)64, were included in this study. The results of the Mini-Mental State Examination (MMSE, Norwegian version) from the clinical assessment were used as a measure of cognitive impairment65,66. Additional descriptive information is summarized in Table 1 and Fig. 1.
Image acquisition
T2*-weighted MRI was obtained on a 3 T General Electric Signa HDxt with different head coils (8-channel, HDNV and HNS), using an echo planar sequence with 203 volumes and the following parameters: repetition time (TR) = 2638 ms; echo time (TE) = 30 ms; flip angle = 90°; acquisition matrix = 64 × 64; in-plane resolution = 4 × 4 mm; 45 axial slices; slice thickness = 3 mm. Each dataset comprised 202 volumes, summing up a total of 202 × 2.638 s = 532.876 s (8 minutes, 53 seconds). For technical reasons, three different head coils were used: standard GE 8-channel (57 AD, 34 MCI and 19 SCI), HDNV (1 AD and 1 SCI) and HNS (21 AD, 10 MCI and 6 SCI). Fisher’s exact test for contingency tables did not reveal significant differences in the distribution of participants across headcoils (two-tailed p = 0.708). T1-weighted data was also collected, and used for co-registration purposes in the current study (FSPGR sequence; TR = 7800 ms; TE = 2.956 ms; TI = 450 ms, flip angle = 12°; in-plane resolution = 1 × 1 mm; number of sagittal slices = 166; slice thickness = 1.2 mm; acquisition time = 7 min 8 s).
Image pre-processing
fMRI data pre-processing was conducted using diverse tools of the FMRI Expert Analysis Tool from the FMRIB Software Library (FSL)67,68,69. Pre-processing steps included brain extraction, motion correction, spatial smoothing (Gaussian kernel, full-width at half-maximum = 6 mm) and high-pass filtering (100 s). The distribution of estimated mean relative motion (volume-to-volume displacement) obtained with FSL’s MCFLIRT is shown in Figure S4. The estimated mean relative in-scanner head motion measured with FSL’s MCFLIRT differed among diagnostic groups, due to a few AD participants with high movement rates (Kruskal-Wallis rank sum test X2 = 6.15, 2 degrees of freedom, p = 0.046; Figure S4). Mean relative motion was later used as a covariate in the statistical analyses. FMRIB’s ICA-based Xnoiseifier (FIX)70,71 was used for data denoising, with a conservative threshold of 60. Briefly, ICA is applied on single-subject data, and the resulting components are submitted to classification as “noise” or “signal” by comparison with a standard catalogue of independent components. Components classified as “noise” are then regressed out. Based on the temporal signal to noise ratio (tSNR)72 computed on the denoised data, three participants were discarded (initially, n = 153). Before FIX, there were between-group differences (Kruskal-Wallis rank sum test: X2 = 7.6, 2 d.f., p = 0.022; Table S1), due to lower tSNR in the SCI subset. Of note, those between-group differences in tSNR were removed after FIX (Kruskal-Wallis rank sum test: X2 = 2.7, 2 d.f., p = 0.259; Table S1).
To obtain precise brain masks for fMRI co-registration, automated whole-brain segmentation was performed on the T1-weighted anatomical references in FreeSurfer73,74; the surface reconstructions were visually inspected and manually edited when necessary. The fMRI datasets were then registered to the individual structural scan using FLIRT and boundary-based registration68,75. The structural scan was warped to the Montreal Neurological Institute MNI152 template (2 mm) using FNIRT75,76,77, before applying the same warping to the fMRI data.
Next, in order to avoid bias in the group-level ICA, three balanced subsets of participants were formed (26 AD, 26 MCI and 26 SCI; no significant differences in gender and age), as proposed by Kaufmann et al.78; an automatic estimation of model order generated 34 ICs. Individual component spatial maps and corresponding time-series were obtained for each subject in the full sample (n = 150) by means of dual regression7. Following standard procedures79, eight components were removed after visual screening, giving a total of 26 ICs’ time-series included in the final analyses. Additional information about those 26 ICs is shown in Fig. 8.
Figure 8. Parcellation-free spatial maps obtained using ICA.

(A) After applying an automatic model order selection and manually removing potential artifacts, 26 spatial components remained. (B) Activation levels derived from fMRI were set into a matrix of [150 subjects]×[202 time points]×[26 components]. Abbreviations: *, sampling time point, corresponding to the TR (here, 2638 ms); a.u., arbitrary units (of activation intensity). (C) Anatomical reference for IC pairs: each pair of ICs was assigned an arbitrary number, which is used as reference in the manuscript.
Average coupling of fMRI signals: static functional connectivity (sFC)
The 26 time-series mentioned above were then submitted to further processing using FSLNets20, running under Matlab (The Mathworks Inc.). In short, the pairwise coupling of these signals (a unique session-wide average per pair) was obtained by L1-norm regularization of the estimated inverse covariance matrix78,80, in order to force to zero the small and potentially noisy values. A total of 325 unique regularized partial correlation measures was obtained and standardized using Fisher’s z transformation; each pairwise correlation is typically considered a functional connection. In the present context, such statistical relationship is referred to as sFC (static functional connectivity), in contrast to the time-varying dynamic patterns described below.
Phase-based metrics of time-varying patterns: dynamic functional connectivity (dFC)
Two different metrics were computed to measure dFC. For whole-brain, metastability was measured by calculating the standard deviation of the Kuramoto order parameter (an index of oscillatory coupling of all regions at every instant; see Fig. 9). A wide range (large standard deviation) of Kuramoto order parameter values would thus characterize brains that traverse different dynamic stages of coupling over time: a broad set of dynamic states, measured as high metastability. Additionally, for pairs of brain regions, the level of fluctuation between synchronization and de-coupling across the scanning session was estimated as the normalized differences in their wave phases.
Figure 9. Computation of phase-based coupling parameters.

The pictures exemplify computation of pairwise phase-based coupling and metastability, for data from 4 ICs (one participant). (A) Four time series collected using FSL’s ICA pipelines. (B) The time series are band-pass filtered (0.04–0.07 Hz), to later compute the Hilbert transform. (C) Individual phases obtained from the imaginary component of the Hilbert transform. (D) Kuramoto order parameter estimated across time using the phases. (E) Pairwise phase-coupling matrices, C(t), estimated for the 4 phases, across all time points; delta phi (Δϕ) is normalized as described in Methods.
The specific procedure was conducted as follows. The 26 time-series obtained from ICA and dual regression were further processed in Matlab (The Mathworks Inc.) to estimate metrics of time-varying connectivity across the scanning session, based upon relevant literature10,11,24. First, each series was narrow-band filtered within 0.04–0.07 Hz, which is required to later obtain meaningful phases24. Then, the Hilbert transform was applied by computing, for each time-series x(t),
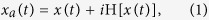 |
where H[·] stands for Hilbert transform and 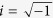 . x(t) can also be expressed as an amplitude-modulated signal a(t) with carrier frequency φ(t), so that
. x(t) can also be expressed as an amplitude-modulated signal a(t) with carrier frequency φ(t), so that
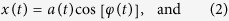 |
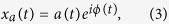 |
where a(t) is the instantaneous envelope and ϕ(t) the instantaneous phase. As suggested by Ponce-Alvarez, et al.11, the first and last ten time points were removed to minimize border effects from the Hilbert transform. The instantaneous phase values are computed for each of the 26 ICs, and then the Kuramoto order parameter (a proxy for the instantaneous whole-brain synchronization) is estimated as:
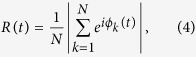 |
where k = {1, …, N} stands for IC number (here, N = 26), and ϕk(t) is the instantaneous phase of the k-th oscillator (time-series) at time t. The metastability was then calculated as the standard deviation of R(t)15. Additionally, pairwise estimates of dynamic phase (de)coupling are estimated as follows: for each time instance t, the pairwise difference in phase between the time-series i and time series j is computed as:
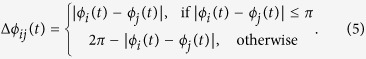 |
For each t, an instantaneous coupling matrix C(t) is obtained by normalizing those phase differences as 1 − Δϕij/π. The coefficient of variation (standard deviation divided by mean) of each element Cij(t) across all time points is the final pairwise metric of phase coupling/de-coupling. From here, there are 325 unique values in the matrix representing relationships between couples of time-series - which can be considered measures of dFC. The whole procedure is schematized in Fig. 9.
This approach was originally introduced by Glerean et al.24, who provided evidence that phase synchronization from fMRI signals can convey reliable information on time-varying connectivity; it was later adapted by Demirtas et al.10; in both cases, surrogate data was generated to test for the presence of dynamic patterns using previously described randomization procedures81,82. Considering the set of narrowband fMRI time-series as an ensemble of chaotic oscillators83,84,85, these techniques are based on the observation that phase synchronization can be measured from their Hilbert-transformed signals86.
To assess whether, in the current dataset, the coefficient of variation of Cij(t) can be considered a measure of temporal dynamics, statistical tests were conducted using surrogate data. Briefly, as recommended by Prichard and Theiler82, multivariate time series of the phase-randomized Fourier transform were obtained using the original dataset; a total of 10000 surrogates was generated, mimicking both the autocorrelations of each variable (IC signal) and the cross correlations between all variables. This simultaneous phase randomization method was implemented using publicly available Matlab code (https://se.mathworks.com/matlabcentral/fileexchange/32621-phase-randomization/content/phaseran.m) with the unfiltered time series; then, after filtering each of the 10000 datasets separately, the coefficient of variation matrix of Cij(t) was computed. The evidence of temporal dynamics was then evaluated at both the whole-brain level (by averaging all individual dFC values of each subject) and for single connections retrieved from the inter-group results. First, the sum of all unique dFC values was estimated for every participant as a general measure of dFC, and a series of one-sample T-tests was performed to evaluate whether the sampled dFC values (in the 150 participants) were drawn from a population with mean dFC ranging from 0 (weak dFC) to the maximum observed value in the surrogates (strong dFC). This set of T statistics was compared with the surrogate T* metrics, and a p-value was obtained for each of the assessed population means as 1 minus the fraction of times that T > T*. Additionally, similar tests were performed for single connections, with a special focus on the edges with statistically significant differences at the group level. Briefly, for each dFC metric (connection) in the original data, a one-sample T-test was conducted to evaluate whether in the 26-participant SCI group (the healthiest participants in this study, where the dFC signal should not be confounded by brain alterations), the sampled dFC values had been drawn from a population with a mean value of 0. From here, 325 T-statistics were retrieved (one for each unique edge), and the same procedure was repeated for each of the 10000 surrogate datasets, obtaining 10000 × 325 T* values. The difference of 1 minus the proportion of times each original T was larger than its corresponding 10000 T* statistics was considered the test p-value.
Statistical analyses
The putative association between metastability and AD diagnose was tested by means of a multinomial logit model, using diagnose as independent variable in
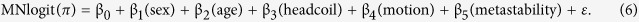 |
Given the distribution of headcoils in this study (Table 1), β3’s regressor was as a dummy variable with a value of “1” for 8-channel data and “0” for both HDNV and HNS. In-scanner motion was adjusted for using regressor β4. This analysis was implemented using Python’s statsmodels module87,88.
Additionally, between-diagnose differences in phase-based dynamic connectivity were tested by means of the network-based statistic (NBS) approach89. Briefly, NBS allows the statistical examination of potential differences in edge weights across different groups or conditions. It controls the family-wise error rate when statistical tests are conducted at single edges comprising a whole graph, on the basis of conventional cluster-based thresholding of statistical parametric maps. In the present scenario, the input matrices (graphs) represent the dFC measures between pairs of ICs; thus, potential dynamic connectivity disruptions are assessed at the edge level using NBS. The design matrices used for NBS here included adjustments for potential confounders (gender, age, headcoil and in-scanner motion); they were submitted to ANCOVA with the standard NBS parameters (5000 permutations; threshold: p = 0.05). Also, when appropriate, logistic regression tests were conducted individually on each of the 325 unique IC pairs. Some data were visualized using R’s PerformanceAnalytics package90,91.
Additional Information
How to cite this article: Córdova-Palomera, A. et al. Disrupted global metastability and static and dynamic brain connectivity across individuals in the Alzheimer’s disease continuum. Sci. Rep. 7, 40268; doi: 10.1038/srep40268 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Material
Acknowledgments
Supported by the Research Council of Norway (204966/F20, 223273, 213837); the South-Eastern Norway Regional Health Authority (2012-047, 2013-123, 2015-073, 2016-083); European Community’s 7th Framework Programme (602450, IMAGEMEND), and Kristian Gerhard Jebsen Foundation. We acknowledge the contribution of patient data from the Norwegian registry for persons being evaluated for cognitive symptoms in specialized care (NorCog) by the Norwegian National Advisory Unit on Ageing and Health.
Footnotes
Author Contributions A.C.-P., T.K. and L.T.W. conceived and designed the experiments, and analyzed the data; A.C.-P., T.K., D.A., N.T.D., T.M., K.P., M.J.L., M.L.B., A.E., A.B., O.A.A., K.E., G.S. and L.T.W. performed the experiments, wrote the paper and gave approval of this version; K.P., M.J.L., M.L.B., A.E., A.B., O.A.A., K.E. and G.S. collected the sample and contributed materials/analysis tools.
References
- Reitz C. & Mayeux R. Alzheimer disease: epidemiology, diagnostic criteria, risk factors and biomarkers. Biochem Pharmacol 88, 640–651, doi: 10.1016/j.bcp.2013.12.024 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tanzi R. E. The genetics of Alzheimer disease. Cold Spring Harb Perspect Med 2, doi: 10.1101/cshperspect.a006296 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- McKhann G. et al. Clinical diagnosis of Alzheimer’s disease: report of the NINCDS-ADRDA Work Group under the auspices of Department of Health and Human Services Task Force on Alzheimer’s Disease. Neurology 34, 939–944 (1984). [DOI] [PubMed] [Google Scholar]
- Nelson P. T. et al. Correlation of Alzheimer disease neuropathologic changes with cognitive status: a review of the literature. J Neuropathol Exp Neurol 71, 362–381, doi: 10.1097/NEN.0b013e31825018f7 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones D. T. et al. Cascading network failure across the Alzheimer’s disease spectrum. Brain 139, 547–562, doi: 10.1093/brain/awv338 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greicius M. D., Srivastava G., Reiss A. L. & Menon V. Default-mode network activity distinguishes Alzheimer’s disease from healthy aging: evidence from functional MRI. Proc Natl Acad Sci USA 101, 4637–4642, doi: 10.1073/pnas.0308627101 (2004). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Filippini N. et al. Distinct patterns of brain activity in young carriers of the APOE-epsilon4 allele. Proc Natl Acad Sci USA 106, 7209–7214, doi: 10.1073/pnas.0811879106 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Westlye E. T., Lundervold A., Rootwelt H., Lundervold A. J. & Westlye L. T. Increased hippocampal default mode synchronization during rest in middle-aged and elderly APOE epsilon4 carriers: relationships with memory performance. J Neurosci 31, 7775–7783, doi: 10.1523/JNEUROSCI.1230-11.2011 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dennis E. L. & Thompson P. M. Functional brain connectivity using fMRI in aging and Alzheimer’s disease. Neuropsychol Rev 24, 49–62, doi: 10.1007/s11065-014-9249-6 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Demirtas M. et al. Dynamic functional connectivity reveals altered variability in functional connectivity among patients with major depressive disorder. Hum Brain Mapp, doi: 10.1002/hbm.23215 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ponce-Alvarez A. et al. Resting-state temporal synchronization networks emerge from connectivity topology and heterogeneity. PLoS Comput Biol 11, e1004100, doi: 10.1371/journal.pcbi.1004100 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fries P. A mechanism for cognitive dynamics: neuronal communication through neuronal coherence. Trends Cogn Sci 9, 474–480, doi: 10.1016/j.tics.2005.08.011 (2005). [DOI] [PubMed] [Google Scholar]
- Fries P. Neuronal gamma-band synchronization as a fundamental process in cortical computation. Annu Rev Neurosci 32, 209–224, doi: 10.1146/annurev.neuro.051508.135603 (2009). [DOI] [PubMed] [Google Scholar]
- Senden M., Deco G., de Reus M. A., Goebel R. & van den Heuvel M. P. Rich club organization supports a diverse set of functional network configurations. Neuroimage 96, 174–182, doi: 10.1016/j.neuroimage.2014.03.066 (2014). [DOI] [PubMed] [Google Scholar]
- Deco G. & Kringelbach M. Metastability and Coherence: Extending the Communication through Coherence Hypothesis Using a Whole-Brain Computational Perspective. Trends Neurosci 39, 432, doi: 10.1016/j.tins.2016.04.006 (2016). [DOI] [PubMed] [Google Scholar]
- Tognoli E. & Kelso J. A. The metastable brain. Neuron 81, 35–48, doi: 10.1016/j.neuron.2013.12.022 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghosh A., Rho Y., McIntosh A. R., Kotter R. & Jirsa V. K. Noise during rest enables the exploration of the brain’s dynamic repertoire. PLoS Comput Biol 4, e1000196, doi: 10.1371/journal.pcbi.1000196 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hindriks R. et al. Can sliding-window correlations reveal dynamic functional connectivity in resting-state fMRI? Neuroimage 127, 242–256, doi: 10.1016/j.neuroimage.2015.11.055 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hutchison R. M. et al. Dynamic functional connectivity: promise, issues, and interpretations. Neuroimage 80, 360–378, doi: 10.1016/j.neuroimage.2013.05.079 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith S. M. et al. Functional connectomics from resting-state fMRI. Trends Cogn Sci 17, 666–682, doi: 10.1016/j.tics.2013.09.016 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aydore S., Pantazis D. & Leahy R. M. A note on the phase locking value and its properties. Neuroimage 74, 231–244, doi: 10.1016/j.neuroimage.2013.02.008 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lachaux J. P., Rodriguez E., Martinerie J. & Varela F. J. Measuring phase synchrony in brain signals. Hum Brain Mapp 8, 194–208 (1999). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Varela F., Lachaux J. P., Rodriguez E. & Martinerie J. The brainweb: phase synchronization and large-scale integration. Nat Rev Neurosci 2, 229–239, doi: 10.1038/35067550 (2001). [DOI] [PubMed] [Google Scholar]
- Glerean E., Salmi J., Lahnakoski J. M., Jaaskelainen I. P. & Sams M. Functional magnetic resonance imaging phase synchronization as a measure of dynamic functional connectivity. Brain Connect 2, 91–101, doi: 10.1089/brain.2011.0068 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cohen J. A power primer. Psychol Bull 112, 155–159 (1992). [DOI] [PubMed] [Google Scholar]
- Chambers J. M. Graphical methods for data analysis. (Wadsworth International Group; Duxbury Press, 1983). [Google Scholar]
- Braskie M. N., Ringman J. M. & Thompson P. M. Neuroimaging measures as endophenotypes in Alzheimer’s disease. Int J Alzheimers Dis 2011, 490140, doi: 10.4061/2011/490140 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hellyer P. J., Scott G., Shanahan M., Sharp D. J. & Leech R. Cognitive Flexibility through Metastable Neural Dynamics Is Disrupted by Damage to the Structural Connectome. J Neurosci 35, 9050–9063, doi: 10.1523/JNEUROSCI.4648-14.2015 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kahana M. J. The cognitive correlates of human brain oscillations. J Neurosci 26, 1669–1672, doi: 10.1523/JNEUROSCI.3737-05c.2006 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shanahan M. Metastable chimera states in community-structured oscillator networks. Chaos 20, 013108, doi: 10.1063/1.3305451 (2010). [DOI] [PubMed] [Google Scholar]
- Kringelbach M. L., McIntosh A. R., Ritter P., Jirsa V. K. & Deco G. The Rediscovery of Slowness: Exploring the Timing of Cognition. Trends Cogn Sci 19, 616–628, doi: 10.1016/j.tics.2015.07.011 (2015). [DOI] [PubMed] [Google Scholar]
- Agosta F. et al. Resting state fMRI in Alzheimer’s disease: beyond the default mode network. Neurobiol Aging 33, 1564–1578, doi: 10.1016/j.neurobiolaging.2011.06.007 (2012). [DOI] [PubMed] [Google Scholar]
- Binnewijzend M. A. et al. Resting-state fMRI changes in Alzheimer’s disease and mild cognitive impairment. Neurobiol Aging 33, 2018–2028, doi: 10.1016/j.neurobiolaging.2011.07.003 (2012). [DOI] [PubMed] [Google Scholar]
- Gour N. et al. Functional connectivity changes differ in early and late-onset Alzheimer’s disease. Hum Brain Mapp 35, 2978–2994, doi: 10.1002/hbm.22379 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hafkemeijer A. et al. Increased functional connectivity and brain atrophy in elderly with subjective memory complaints. Brain Connect 3, 353–362, doi: 10.1089/brain.2013.0144 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Jong L. W. et al. Strongly reduced volumes of putamen and thalamus in Alzheimer’s disease: an MRI study. Brain 131, 3277–3285, doi: 10.1093/brain/awn278 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sluimer J. D. et al. Accelerating regional atrophy rates in the progression from normal aging to Alzheimer’s disease. Eur Radiol 19, 2826–2833, doi: 10.1007/s00330-009-1512-5 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klunk W.E. et al. Imaging brain amyloid in Alzheimer’s disease with Pittsburgh Compound-B. Ann Neurol 55, 306–319, doi: 10.1002/ana.20009 (2004). [DOI] [PubMed] [Google Scholar]
- Pievani M., de Haan W., Wu T., Seeley W. W. & Frisoni G. B. Functional network disruption in the degenerative dementias. Lancet Neurol 10, 829–843, doi: 10.1016/S1474-4422(11)70158-2 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sourty M. et al. Identifying Dynamic Functional Connectivity Changes in Dementia with Lewy Bodies Based on Product Hidden Markov Models. Front Computat Neurosc 10, 60 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bartzokis G. Age-related myelin breakdown: a developmental model of cognitive decline and Alzheimer’s disease. Neurobiol Aging 25, 5–18; author reply 49-62 (2004). [DOI] [PubMed] [Google Scholar]
- Behrendt G. et al. Dynamic changes in myelin aberrations and oligodendrocyte generation in chronic amyloidosis in mice and men. Glia 61, 273–286, doi: 10.1002/glia.22432 (2013). [DOI] [PubMed] [Google Scholar]
- Marner L., Nyengaard J. R., Tang Y. & Pakkenberg B. Marked loss of myelinated nerve fibers in the human brain with age. J Comp Neurol 462, 144–152, doi: 10.1002/cne.10714 (2003). [DOI] [PubMed] [Google Scholar]
- Zhan X. et al. Myelin basic protein associates with AbetaPP, Abeta1-42, and amyloid plaques in cortex of Alzheimer’s disease brain. J Alzheimers Dis 44, 1213–1229, doi: 10.3233/JAD-142013 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deco G., Tononi G., Boly M. & Kringelbach M. L. Rethinking segregation and integration: contributions of whole-brain modelling. Nat Rev Neurosci 16, 430–439, doi: 10.1038/nrn3963 (2015). [DOI] [PubMed] [Google Scholar]
- Kello C. T. Critical branching neural networks. Psychol Rev 120, 230–254, doi: 10.1037/a0030970 (2013). [DOI] [PubMed] [Google Scholar]
- Rubinov M. & Bullmore E. Fledgling pathoconnectomics of psychiatric disorders. Trends Cogn Sci 17, 641–647, doi: 10.1016/j.tics.2013.10.007 (2013). [DOI] [PubMed] [Google Scholar]
- Fischer P. et al. Conversion from subtypes of mild cognitive impairment to Alzheimer dementia. Neurology 68, 288–291, doi: 10.1212/01.wnl.0000252358.03285.9d (2007). [DOI] [PubMed] [Google Scholar]
- Gallassi R. et al. Are subjective cognitive complaints a risk factor for dementia? Neurol Sci 31, 327–336, doi: 10.1007/s10072-010-0224-6 (2010). [DOI] [PubMed] [Google Scholar]
- Nobili F. et al. Brain SPECT in subtypes of mild cognitive impairment. Findings from the DESCRIPA multicenter study. J Neurol 255, 1344–1353, doi: 10.1007/s00415-008-0897-4 (2008). [DOI] [PubMed] [Google Scholar]
- Fonseca J. A. et al. Factors that predict cognitive decline in patients with subjective cognitive impairment. Int Psychogeriatr 27, 1671–1677, doi: 10.1017/S1041610215000356 (2015). [DOI] [PubMed] [Google Scholar]
- Reisberg B. & Gauthier S. Current evidence for subjective cognitive impairment (SCI) as the pre-mild cognitive impairment (MCI) stage of subsequently manifest Alzheimer’s disease. Int Psychogeriatr 20, 1–16, doi: 10.1017/S1041610207006412 (2008). [DOI] [PubMed] [Google Scholar]
- Reisberg B., Shulman M. B., Torossian C., Leng L. & Zhu W. Outcome over seven years of healthy adults with and without subjective cognitive impairment. Alzheimers Dement 6, 11–24, doi: 10.1016/j.jalz.2009.10.002 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stewart R. Subjective cognitive impairment. Curr Opin Psychiatry 25, 445–450, doi: 10.1097/YCO.0b013e3283586fd8 (2012). [DOI] [PubMed] [Google Scholar]
- Maier A. et al. Divergence of fMRI and neural signals in V1 during perceptual suppression in the awake monkey. Nat Neurosci 11, 1193–1200, doi: 10.1038/nn.2173 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zumer J. M., Brookes M. J., Stevenson C. M., Francis S. T. & Morris P. G. Relating BOLD fMRI and neural oscillations through convolution and optimal linear weighting. Neuroimage 49, 1479–1489, doi: 10.1016/j.neuroimage.2009.09.020 (2010). [DOI] [PubMed] [Google Scholar]
- Ramsey J. D. et al. Six problems for causal inference from fMRI. Neuroimage 49, 1545–1558, doi: 10.1016/j.neuroimage.2009.08.065 (2010). [DOI] [PubMed] [Google Scholar]
- Mecocci P. Oxidative stress in mild cognitive impairment and Alzheimer disease: a continuum. J Alzheimers Dis 6, 159–163 (2004). [DOI] [PubMed] [Google Scholar]
- Mesulam M., Shaw P., Mash D. & Weintraub S. Cholinergic nucleus basalis tauopathy emerges early in the aging-MCI-AD continuum. Ann Neurol 55, 815–828, doi: 10.1002/ana.20100 (2004). [DOI] [PubMed] [Google Scholar]
- Panza F. et al. Late-life depression, mild cognitive impairment, and dementia: possible continuum? Am J Geriatr Psychiatry 18, 98–116, doi: 10.1097/JGP.0b013e3181b0fa13 (2010). [DOI] [PubMed] [Google Scholar]
- Brækhus A., Ulstein I., Wyller T. B. & Engedal K. The Memory Clinic – outpatient assessment when dementia is suspected. Tidsskr Nor Legeforen 131, 2254–2257 (2011). [DOI] [PubMed] [Google Scholar]
- World Health Organization. The ICD-10 classification of mental and behavioural disorders: diagnostic criteria for research. (World Health Organization, 1993). [Google Scholar]
- Winblad B. et al. Mild cognitive impairment-beyond controversies, towards a consensus: report of the International Working Group on Mild Cognitive Impairment. J Intern Med 256, 240–246, doi: 10.1111/j.1365-2796.2004.01380.x (2004). [DOI] [PubMed] [Google Scholar]
- Garcia-Ptacek S. et al. Subjective cognitive impairment subjects in our clinical practice. Dement Geriatr Cogn Dis Extra 4, 419–430, doi: 10.1159/000366270 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Engedal K., Haugen P., Gilje K. & Laake P. Efficacy of short mental tests in the detection of mental impairment in old age. Compr Gerontol A 2, 87–93 (1988). [PubMed] [Google Scholar]
- Folstein M. F., Folstein S. E. & McHugh P. R. “Mini-mental state”. A practical method for grading the cognitive state of patients for the clinician. J Psychiatr Res 12, 189–198 (1975). [DOI] [PubMed] [Google Scholar]
- Beckmann C. F. & Smith S. M. Probabilistic independent component analysis for functional magnetic resonance imaging. IEEE Trans Med Imaging 23, 137–152, doi: 10.1109/TMI.2003.822821 (2004). [DOI] [PubMed] [Google Scholar]
- Jenkinson M., Bannister P., Brady M. & Smith S. Improved optimization for the robust and accurate linear registration and motion correction of brain images. Neuroimage 17, 825–841 (2002). [DOI] [PubMed] [Google Scholar]
- Smith S. M. et al. Advances in functional and structural MR image analysis and implementation as FSL. Neuroimage 23 Suppl 1, S208–219, doi: 10.1016/j.neuroimage.2004.07.051 (2004). [DOI] [PubMed] [Google Scholar]
- Griffanti L. et al. ICA-based artefact removal and accelerated fMRI acquisition for improved resting state network imaging. Neuroimage 95, 232–247, doi: 10.1016/j.neuroimage.2014.03.034 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salimi-Khorshidi G. et al. Automatic denoising of functional MRI data: combining independent component analysis and hierarchical fusion of classifiers. Neuroimage 90, 449–468, doi: 10.1016/j.neuroimage.2013.11.046 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roalf D. R. et al. The impact of quality assurance assessment on diffusion tensor imaging outcomes in a large-scale population-based cohort. Neuroimage 125, 903–919, doi: 10.1016/j.neuroimage.2015.10.068 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dale A. M., Fischl B. & Sereno M. I. Cortical surface-based analysis. I. Segmentation and surface reconstruction. Neuroimage 9, 179–194, doi: 10.1006/nimg.1998.0395 (1999). [DOI] [PubMed] [Google Scholar]
- Fischl B. et al. Whole brain segmentation: automated labeling of neuroanatomical structures in the human brain. Neuron 33, 341–355 (2002). [DOI] [PubMed] [Google Scholar]
- Greve D. N. & Fischl B. Accurate and robust brain image alignment using boundary-based registration. Neuroimage 48, 63–72, doi: 10.1016/j.neuroimage.2009.06.060 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jenkinson M., Beckmann C. F., Behrens T. E., Woolrich M. W. & Smith S. M. Fsl. Neuroimage 62, 782–790, doi: 10.1016/j.neuroimage.2011.09.015 (2012). [DOI] [PubMed] [Google Scholar]
- Mazziotta J. et al. A probabilistic atlas and reference system for the human brain: International Consortium for Brain Mapping (ICBM). Philos Trans R Soc Lond B Biol Sci 356, 1293–1322, doi: 10.1098/rstb.2001.0915 (2001). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaufmann T. et al. Disintegration of Sensorimotor Brain Networks in Schizophrenia. Schizophr Bull 41, 1326–1335, doi: 10.1093/schbul/sbv060 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kelly R. E. Jr. et al. Visual inspection of independent components: defining a procedure for artifact removal from fMRI data. J Neurosci Methods 189, 233–245, doi: 10.1016/j.jneumeth.2010.03.028 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Friedman J., Hastie T. & Tibshirani R. Sparse inverse covariance estimation with the graphical lasso. Biostatistics 9, 432–441, doi: 10.1093/biostatistics/kxm045 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Politis D. N. & Romano J. P. A circular block-resampling procedure for stationary data. In Exploring the limits of bootstrap (eds LePage R & Billard L), 263–270 (Wiley-Interscience, 1992). [Google Scholar]
- Prichard D. & Theiler J. Generating Surrogate Data for Time-Series with Several Simultaneously Measured Variables. Phys Rev Lett 73, 951–954, doi: 10.1103/PhysRevLett.73.951 (1994). [DOI] [PubMed] [Google Scholar]
- Deco G., Jirsa V. K. & McIntosh A. R. Emerging concepts for the dynamical organization of resting-state activity in the brain. Nat Rev Neurosci 12, 43–56, doi: 10.1038/nrn2961 (2011). [DOI] [PubMed] [Google Scholar]
- Gollo L. L., Zalesky A., Hutchison R. M., van den Heuvel M. & Breakspear M. Dwelling quietly in the rich club: brain network determinants of slow cortical fluctuations. Philos Trans R Soc Lond B Biol Sci 370, doi: 10.1098/rstb.2014.0165 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Honey C. J., Kotter R., Breakspear M. & Sporns O. Network structure of cerebral cortex shapes functional connectivity on multiple time scales. Proc Natl Acad Sci USA 104, 10240–10245, doi: 10.1073/pnas.0701519104 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosenblum M. G., Pikovsky A. S. & Kurths J. Phase synchronization of chaotic oscillators. Phys Rev Lett 76, 1804–1807, doi: DOI 10.1103/PhysRevLett.76.1804 (1996). [DOI] [PubMed] [Google Scholar]
- Seabold S. & Perktold J. Statsmodels: Econometric and statistical modeling with python. In Proceedings of the 9th Python in Science Conference (eds van der Walt S. & Millman J.), 57–61 (SciPy, 2010).
- Van Rossum G. & Drake F. L. The python language reference manual. (Network Theory Ltd., 2011). [Google Scholar]
- Zalesky A., Fornito A. & Bullmore E. T. Network-based statistic: identifying differences in brain networks. Neuroimage 53, 1197–1207, doi: 10.1016/j.neuroimage.2010.06.041 (2010). [DOI] [PubMed] [Google Scholar]
- Peterson B. G. et al. PerformanceAnalytics: Econometric tools for performance and risk analysis R package version 1.4.3541. https://cran.r-project.org/package=PerformanceAnalytics (2014).
- Core Team R R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org/ (2015).
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.