

Abstract
Background
Bidirectional signalling between the brain and the gastrointestinal tract is regulated at neural, hormonal, and immunological levels. Recent studies have shown that helminth infections can alter the normal gut microbiota. Studies have also shown that the gut microbiota is instrumental in the normal development, maturation and function of the brain. The pathophysiological pathways by which helminth infections contribute to altered cognitive function remain poorly understood.
Discussion
We put forward the hypothesis that gastrointestinal infections with parasitic worms, such as helminths, induce an imbalance of the gut-brain axis, which, in turn, can detrimentally manifest in brain development. Factors supporting this hypothesis are: 1) research focusing on intelligence and school performance in school-aged children has shown helminth infections to be associated with cognitive impairment, 2) disturbances in gut microbiota have been shown to be associated with important cognitive developmental effects, and 3) helminth infections have been shown to alter the gut microbiota structure. Evidence on the complex interactions between extrinsic (parasite) and intrinsic (host-derived) factors has been synthesised and discussed.
Summary
While evidence in favour of the helminth-gut microbiota-central nervous system hypothesis is circumstantial, it would be unwise to rule it out as a possible mechanism by which gastrointestinal helminth infections induce childhood cognitive morbidity. Further empirical studies are necessary to test an indirect effect of helminth infections on the modulation of mood and behaviour through its effects on the gut microbiota.
Keywords: Central nervous system, Development, Gut microbiota, Helminths, Mental health, Microbiota-gut-brain axis
Background
Soil-transmitted helminth (STH) infections such as Ascaris lumbricoides, Trichuris trichiura and hookworms affect more than a third of the world’s population, with the heaviest worm burdens occurring in non-industrialized countries [1]. STH,infections are associated with significant morbidity, particularly in children; infection is recognised to cause nutritional deficits, with clinical and physical consequences including anaemia and reduced growth [2–5]. In addition to their nutritional effects, soil-transmitted helminth infections are also reported to impair cognitive function [6–8], limit educational advancement, and as a result, hinder economic development [9]. The effects of helminth infections, particularly by hookworms, on cognitive development of children were first reported by Waite and Neilson in 1919 [10]. Since then, there have been several studies that looked at the effects of STH infections on different domains of cognitive function. Nevertheless, at present, available evidence is conflicting. A recent update of the Cochrane review concluded that there is little or no evidence that cognitive function is affected by STH infections [11, 12], but some have argued that trials included in the Cochrane review were of poor quality to measure effects [13] and that STH effects on cognition could even be greater than initially suggested [3]. Several observational studies (some not included in the meta-analysis by the Cochrane review) reviewed in [13] reported that STH infections impair the efficiency of cognitive processes including memory, learning, verbal fluency and non-verbal intelligence [6–8, 14–17]. In addition, investigations into STH infections during pregnancy have also demonstrated an association with poor cognitive and motor development in infants [18, 19].
Multiple mechanisms have been demonstrated to explain STH-induced nutritional impairment in infected individuals: a loss of iron and protein through feeding on host tissues [20], an increasing malabsorption of nutrients [5], a competition for vitamin A in the intestine [21], or diarrhoea and dysentery [22]. Conversely, the pathophysiological pathways by which STH infections contribute to altered intellectual performance reported by some studies are still subject to significant debate. Some of the cognitive effects of STH infection can be partly explained by the direct effect of observed nutritional deficits on the brain and the indirect effect of pathophysiological events occurring in the gut environment where STH reside; the relative importance of these competing mechanisms remain unclear.
Main text
Interaction between gut microbial communities and the brain
There is a bidirectional functional communication between gut microbiota, the gastrointestinal (GI) tract and the central nervous system (CNS); these relationships have been recognised as the microbiota-gut-brain axis, which has been subject to substantial scientific enquiry in recent years [23–26]. The microbiota-gut-brain axis operates through a variety of physiological mechanisms, including neural, hormonal and immunological pathways [24].
Recent reports pointed to a crucial role of the microbiota-gut-brain axis in normal development, maturation and function of the brain [27–29], supported by emerging evidence that the disruption of the gut microbial community (i.e. dysbiosis) can affect emotional behaviour and related brain systems, which can lead to a range of abnormal or altered phenotypes, i.e. human brain diseases including autism spectrum disorder [30], anxiety, depression, and chronic pain [31, 32]. Clinical findings of these studies are supported by evidence primarily collected from experimental studies in rodents using various strategies: experiments using germ-free animals, experimental modification of the gut microbiota via antibiotics (downregulation) or prebiotics/probiotics (upregulation), or experimental infection with various pathogenic bacteria. Experimental studies on mice have shown how the microbial content of the GI tract influences feeding behaviours [33], stress-related behaviours [23, 34–36], pain perception pathways [37], and memory/learning development [36, 38].
Gut microbiota and brain development
Mammalian brain development is initiated in utero, with rapid changes in neuronal organization [29]. But a considerable amount of morphological development, cell differentiation and acquisition of function takes place during postnatal development, with a striking increase in brain growth during the first 2 years of life [39]. Internal and external environmental signals such as nutrition, infection, the environment, or stress (maternal or environmental) can affect brain development until adulthood. As an example of diet impact, prolonged and exclusive breastfeeding has been shown to improve children’s cognitive development [40, 41].
Interestingly, the maturation of the gut microbiota occurs during the first 2 to 3 years of postnatal life, coinciding with a critical window of early brain development [42]. Initial colonization of the GI tract is dictated by the mother’s microbes (during the delivery) and the hospital environment, and further influenced by a number of factors including antibiotic use, diet, mode of delivery, environmental factors, or the maternal microbiota [43–45]. The gut microbiota plays a fundamental role in key systems regulating CNS development, especially synaptogenesis and myelination [39, 46]; this entails that a sustained imbalance within the microbial ecosystem of the infant gut could impair the cognitive development in early life. Insufficient or disturbed colonization of an infant resulting from Caesarean section and/or inadequate nursing could thus have unexpected outcomes.
Interactions between helminths, gut microbiota and the host
One environmental factor impacting the gut microbiota is parasitic infection. The qualitative and quantitative alterations on the composition of the gut microbiota of the host upon helminth infection, and the underlying mechanisms leading to these changes have mainly been studied in animal models, specifically Heligmosomoides polygyrus bakeri infection in mice and Trichuris suis infection in pigs (reviewed in [47]). Such evidence in humans [48] or wild animals [49] under natural settings is scarce but also rarely investigated. A recent study on a population of wild mice naturally infected showed a modification in the diversity and composition of the gut microbiota, with evidence that the abundance of gut microbial taxa varies according to the helminth species colonizing the host [49].
A potential mechanism by which helminth infection could alter the gut microbiota composition is its effect upon the host immune system, which could disrupt the homeostatic relationship established between the gut microbiota and the host. Helminth effects on the host immune system can come about by direct competition for niche space in the GI tract, or primarily through host immunomodulation. The direct helminth-induced host immune responses include down-regulation of inflammation, mainly via a protective Th2 response [50, 51]. These changes have resistance and tolerance roles, establishing an environment that promotes parasite survival and a prolonged reproductive phase [50–53]. Helminth interaction with the host immune cells/molecules can also occur indirectly through the excretory/secretory products released by live worms [54]. These proteins with immunomodulation properties are involved in creating an anti-inflammatory (e.g. by induction of Th2 response) and immuno-tolerant environment (e.g. via host dendritic cells modulation) [55–57] that promote both helminth and host survival [51]. The host-helminth-microbiota interaction is thus a complex and dynamic relationship, and all three components must be considered to better understand helminth pathogenesis.
Testing the helminth-gut microbiota-CNS hypothesis
Available data suggests that helminths, the gut microbiota and the host should be viewed as a dynamic and integrated system [52]. In support of that view is current evidence pointing out that (1) helminth infection in infants is associated with significant cognitive impairment; (2) studies focusing on the gut-microbiota-brain axis demonstrate that the gut microbiota has a key role in early brain development and dysbiosis in the gut microbiota adversely affects cognitive abilities; and (3) STH infections result in gut microbiota dysbiosis. This suggests that any disturbance in homeostasis of the healthy gut microbiota, e.g. as a result of STH infection, is likely to impact on the host’s health (Fig. 1). We thus hypothesize that the effects on cognitive function associated with helminth infection can be partly explained by a secondary/indirect effect of the gut microbiota dysbiosis induced by infection.
Fig. 1.
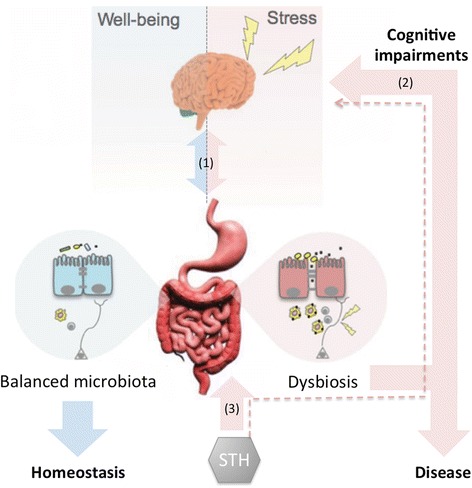
The microbiota-gut-brain axis and its interactions with soil-transmitted helminths (STH): (1) shows the bidirectional communication between the gut and the brain, which occurs through multiple pathways that include hormonal, neural and immune mediators; (2) shows the impact of gut microbiota dysbiosis on cognition; (3) shows the impact of helminth infection on the gut microbiota. The dotted arrow shows the hypothesized pathway leading from STH infection to cognitive impairments, potentially through its impact on the gut microbiota (i.e. dysbiosis). Adapted from De Palma et al. 2014 [65]
While there is evidence on the interactions between different players in this complex system, the quantitative rates by which the players interact with each other cannot be informed from current literature. The helminth-gut microbiota-CNS hypothesis needs support from further experimental and epidemiological studies. Some reliable measurement tools exist to assess anxiety behaviours, working memory or pain perception, and they have been used in some studies investigating the impact of helminth-microbiota induced changes on those parameters. The specific micro-organisms and components involved in this process, the clinical relevance and the mechanisms underlying possible alterations of the microbiota composition in helminth-infected children require further population-based studies. These investigations would bring a novel and refreshing approach to scientific enquiry into the role of STH on cognitive function of children, with important implications for clinical practice by offering a range of potential therapeutic opportunities to target CNS developmental and behavioural disorders.
There is strong evidence linking parasite infections, particularly hookworms, to anemia [58], and many studies which link anemia to disturbances in social, emotional, and cognitive development (e.g. [59, 60]). Thus investigations could be extended to examine the role of parasite infection on cognitive impairment as an indirect result of anemia. Furthermore, recent studies showed that, unlike gut microbiota from healthy children, microbiota from undernourished children is immature [61–64] and can transmit growth impairment, as shown by faecal transplant experiments [61]. So it would be worth testing the potential impact of helminth infection on children growth via infection-induced microbiota dysbiosis through a similar loop analysis.
Summary
We put forward the hypothesis that changes in the gut microbiota induced by helminth infections play an important role in cognitive morbidity of children (the helminth-gut microbiome-CNS axis). Factors supporting this hypothesis are: 1) the role that gut microbiota has on cognitive development; 2) the ability of helminth infections to change gut microbiota composition and diversity; and 3) the observed effect of helminth infection on cognitive development indicators. The hypothesis should be further tested using experimental and epidemiological studies.
Conclusions
While available evidence in favour of the helminth-microbiota-CNS hypothesis is circumstantial, the recent debate around helminth associated morbidity indicate the need for further research to elucidate the mechanisms through which gastrointestinal helminth infections induce cognitive developmental morbidity. Future studies looking at the effect on STH on childhood cognitive developmental domains should be adequately powered to measure effects that are likely to be subtle. In addition future studies should considered validated tools for measuring cognitive morbidity effects; these need to be sensitive enough to detect quantitative changes in the microbiota and longitudinal study designs will be paramount to quantify the effects on each element of the system. Combining metagenomic output with comprehensive psychometric tests constitutes an important direction for future work.
Acknowledgements
We thank the editorial advice from Prof. Gregory O’Brien MD MRCPsych II PhD and Dr. Jennifer Galstuch Leon MD MRCPsych II on the very early draft of the manuscript. We sincerely thank the editor and the two reviewers for constructive criticisms and valuable comments on a previous version of this manuscript.
Funding
The work was funded through a University of Queensland New Staff Start-up Grant; the funding source had no involvement in study design, data collection, analysis and interpretation, in the writing of the report and in the decision to submit the paper for publication.
Availability of data and material
Data sharing not applicable to this article as no datasets were generated or analysed during the current study.
Author’s contributions
RSM raised the original idea for the paper. BB did the initial literature search and BB and VG collated and reviewed studies. RSM structured the initial draft and BB wrote the initial draft. RSM, GM and LY provided supervision. GM and VG drafted the figures. VG, BB, RSM, LY, GM and AC wrote sections of the draft and revised subsequent drafts. All authors have contributed to revision the manuscript.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Ethics approval and consent to participate
Not applicable.
Abbreviations
- CNS
Central nervous system
- GI
Gastrointestinal
- STH
Soil-transmitted helmiths
References
- 1.de Silva NR, Brooker S, Hotez PJ, Montresor A, Engels D, Savioli L. Soil-transmitted helminth infections: updating the global picture. Trends Parasitol. 2003;19(12):547–551. doi: 10.1016/j.pt.2003.10.002. [DOI] [PubMed] [Google Scholar]
- 2.Stephenson LS, Latham MC, Ottesen EA. Malnutrition and parasitic helminth infections. Parasitology. 2000;121(Suppl):S23–38. doi: 10.1017/S0031182000006491. [DOI] [PubMed] [Google Scholar]
- 3.Bethony J, Brooker S, Albonico M, Geiger SM, Loukas A, Diemert D, Hotez PJ. Soil-transmitted helminth infections: ascariasis, trichuriasis, and hookworm. Lancet. 2006;367(9521):1521–1532. doi: 10.1016/S0140-6736(06)68653-4. [DOI] [PubMed] [Google Scholar]
- 4.Pullan RL, Smith JL, Jasrasaria R, Brooker SJ. Global numbers of infection and disease burden of soil transmitted helminth infections in 2010. Parasit Vectors. 2014;7:37. doi: 10.1186/1756-3305-7-37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Hall A, Hewitt G, Tuffrey V, de Silva N. A review and meta-analysis of the impact of intestinal worms on child growth and nutrition. Matern Child Nutr. 2008;4(Suppl 1):118–236. doi: 10.1111/j.1740-8709.2007.00127.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ezeamama AE, Friedman JF, Acosta LP, Bellinger DC, Langdon GC, Manalo DL, Olveda RM, Kurtis JD, McGarvey ST. Helminth infection and cognitive impairment among Filipino children. Am J Trop Med Hyg. 2005;72(5):540. [PMC free article] [PubMed] [Google Scholar]
- 7.Kvalsvig JD, Cooppan RM, Connolly KJ. The effects of parasite infections on cognitive processes in children. Ann Trop Med Parasitol. 1991;85(5):551–568. doi: 10.1080/00034983.1991.11812608. [DOI] [PubMed] [Google Scholar]
- 8.Nokes C, Grantham-McGregor SM, Sawyer AW, Cooper ES, Bundy DA. Parasitic helminth infection and cognitive function in school children. Proc Biol sci/Roy Soc. 1992;247(1319):77–81. doi: 10.1098/rspb.1992.0011. [DOI] [PubMed] [Google Scholar]
- 9.Hotez PJ, Bundy DAP, Beegle K, Brooker S, Drake L, de Silva N, Montresor A, Engels D, Jukes M, Chitsulo L, et al. Helminth Infections: Soil-transmitted Helminth Infections and Schistosomiasis. In: Jamison DT, Breman JG, Measham AR, Alleyne G, Claeson M, Evans DB, Jha P, Mills A, Musgrove P, editors. Disease Control Priorities in Developing Countries. 2nd edn. Washington (DC): The International Bank for Reconstruction and Developement/ The World Bank; New York: Oxford University Press; 2006.
- 10.Waite JH, Neilson IL. A study of the effects of hookworm infection upon mental development of North Queensland school children. Med J Aust. 1919;1:1–7. [Google Scholar]
- 11.Taylor-Robinson DC, Maayan N, Soares-Weiser K, Donegan S, Garner P. Deworming drugs for soil-transmitted intestinal worms in children: effects on nutritional indicators, haemoglobin, and school performance. Cochrane Database Syst Rev. 2015;7:CD000371. doi: 10.1002/14651858.CD000371.pub6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Dickson R, Awasthi S, Williamson P, Demellweek C, Garner P. Effects of treatment for intestinal helminth infection on growth and cognitive performance in children: systematic review of randomised trials. BMJ. 2000;320(7251):1697–1701. doi: 10.1136/bmj.320.7251.1697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Kvalsvig J, Albonico M. Effects of geohelminth infections on neurological development. Handb Clin Neurol. 2013;114:369–379. doi: 10.1016/B978-0-444-53490-3.00029-7. [DOI] [PubMed] [Google Scholar]
- 14.Liu C, Luo R, Yi H, Zhang L, Li S, Bai Y, Medina A, Rozelle S, Smith S, Wang G, et al. Soil-transmitted helminths in southwestern china: a cross-sectional study of links to cognitive ability, nutrition, and school performance among children. PLoS Negl Trop Dis. 2015;9(6):e0003877. doi: 10.1371/journal.pntd.0003877. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Sakti H, Nokes C, Hertanto WS, Hendratno S, Hall A, Bundy DA. Satoto: evidence for an association between hookworm infection and cognitive function in Indonesian school children. Tropical med Int health. 1999;4(5):322–334. doi: 10.1046/j.1365-3156.1999.00410.x. [DOI] [PubMed] [Google Scholar]
- 16.Taylor-Robinson DC, Maayan N, Soares-Weiser K, Donegan S, Garner P. Deworming drugs for soil-transmitted intestinal worms in children: effects on nutritional indicators, haemoglobin and school performance. Cochrane Database Syst Rev. 2012;7:CD000371. doi: 10.1002/14651858.CD000371.pub4. [DOI] [PubMed] [Google Scholar]
- 17.Nokes C, Bundy DA. Does helminth infection affect mental processing and educational achievement? Parasitol Today. 1994;10(1):14–18. doi: 10.1016/0169-4758(94)90348-4. [DOI] [PubMed] [Google Scholar]
- 18.Mireku MO, Boivin MJ, Davidson LL, Ouedraogo S, Koura GK, Alao MJ, Massougbodji A, Cot M, Bodeau-Livinec F. Impact of helminth infection during pregnancy on cognitive and motor functions of one-year-old children. PLoS Negl Trop Dis. 2015;9(3):e0003463. doi: 10.1371/journal.pntd.0003463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Nampijja M, Apule B, Lule S, Akurut H, Muhangi L, Webb EL, Lewis C, Elliott AM, Alcock KJ. Effects of maternal worm infections and anthelminthic treatment during pregnancy on infant motor and neurocognitive functioning. J Int Neuropsychol Soc. 2012;18(6):1019–1030. doi: 10.1017/S1355617712000768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Solomons NW. Pathways to the impairment of human nutritional status by gastrointestinal pathogens. Parasitology. 1993;107(Suppl):S19–35. doi: 10.1017/S003118200007548X. [DOI] [PubMed] [Google Scholar]
- 21.Ahmed F, Mohiduzzaman M, Jackson AA. Vitamin a absorption in children with ascariasis. Br J Nutr. 1993;69(3):817–825. doi: 10.1079/BJN19930082. [DOI] [PubMed] [Google Scholar]
- 22.Callender JE, Walker SP, Grantham-McGregor SM, Cooper ES. Growth and development four years after treatment for the Trichuris dysentery syndrome. Acta Paediatr. 1998;87(12):1247–1249. doi: 10.1111/j.1651-2227.1998.tb00946.x. [DOI] [PubMed] [Google Scholar]
- 23.Bravo JA, Julio-Pieper M, Forsythe P, Kunze W, Dinan TG, Bienenstock J, Cryan JF. Communication between gastrointestinal bacteria and the nervous system. Curr Opin Pharmacol. 2012;12(6):667–672. doi: 10.1016/j.coph.2012.09.010. [DOI] [PubMed] [Google Scholar]
- 24.Chen X, D’Souza R, Hong ST. The role of gut microbiota in the gut-brain axis: current challenges and perspectives. Protein Cell. 2013;4(6):403–414. doi: 10.1007/s13238-013-3017-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Cryan JF, Dinan TG. Mind-altering microorganisms: the impact of the gut microbiota on brain and behaviour. Nat Rev Neurosci. 2012;13(10):701–712. doi: 10.1038/nrn3346. [DOI] [PubMed] [Google Scholar]
- 26.Rhee SH, Pothoulakis C, Mayer EA. Principles and clinical implications of the brain-gut-enteric microbiota axis. Nat Rev Gastroenterol Hepatol. 2009;6(5):306–314. doi: 10.1038/nrgastro.2009.35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Al-Asmakh M, Anuar F, Zadjali F, Rafter J, Pettersson S. Gut microbial communities modulating brain development and function. Gut microbes. 2012;3(4):366–373. doi: 10.4161/gmic.21287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Sudo N, Chida Y, Aiba Y, Sonoda J, Oyama N, Yu XN, Kubo C, Koga Y. Postnatal microbial colonization programs the hypothalamic-pituitary-adrenal system for stress response in mice. J Physiol. 2004;558(Pt 1):263–275. doi: 10.1113/jphysiol.2004.063388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Diaz Heijtz R, Wang S, Anuar F, Qian Y, Bjorkholm B, Samuelsson A, Hibberd ML, Forssberg H, Pettersson S. Normal gut microbiota modulates brain development and behavior. Proc Natl Acad Sci U S A. 2011;108(7):3047–3052. doi: 10.1073/pnas.1010529108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Finegold SM, Downes J, Summanen PH. Microbiology of regressive autism. Anaerobe. 2012;18(2):260–262. doi: 10.1016/j.anaerobe.2011.12.018. [DOI] [PubMed] [Google Scholar]
- 31.Mayer EA, Knight R, Mazmanian SK, Cryan JF, Tillisch K. Gut microbes and the brain: paradigm shift in neuroscience. J neuro. 2014;34(46):15490–15496. doi: 10.1523/JNEUROSCI.3299-14.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Foster JA, McVey Neufeld KA. Gut-brain axis: how the microbiome influences anxiety and depression. Trends Neurosci. 2013;36(5):305–312. doi: 10.1016/j.tins.2013.01.005. [DOI] [PubMed] [Google Scholar]
- 33.Hsiao EY, McBride SW, Hsien S, Sharon G, Hyde ER, McCue T, Codelli JA, Chow J, Reisman SE, Petrosino JF, et al. Microbiota modulate behavioral and physiological abnormalities associated with neurodevelopmental disorders. Cell. 2013;155(7):1451–1463. doi: 10.1016/j.cell.2013.11.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Goehler LE, Park SM, Opitz N, Lyte M, Gaykema RP. Campylobacter jejuni infection increases anxiety-like behavior in the holeboard: possible anatomical substrates for viscerosensory modulation of exploratory behavior. Brain Behav Immun. 2008;22(3):354–366. doi: 10.1016/j.bbi.2007.08.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Lyte M, Li W, Opitz N, Gaykema RP, Goehler LE. Induction of anxiety-like behavior in mice during the initial stages of infection with the agent of murine colonic hyperplasia Citrobacter rodentium. Physiol Behav. 2006;89(3):350–357. doi: 10.1016/j.physbeh.2006.06.019. [DOI] [PubMed] [Google Scholar]
- 36.Gareau MG, Wine E, Rodrigues DM, Cho JH, Whary MT, Philpott DJ, Macqueen G, Sherman PM. Bacterial infection causes stress-induced memory dysfunction in mice. Gut. 2011;60(3):307–317. doi: 10.1136/gut.2009.202515. [DOI] [PubMed] [Google Scholar]
- 37.Forsythe P, Sudo N, Dinan T, Taylor VH, Bienenstock J. Mood and gut feelings. Brain Behav Immun. 2010;24(1):9–16. doi: 10.1016/j.bbi.2009.05.058. [DOI] [PubMed] [Google Scholar]
- 38.Li W, Dowd SE, Scurlock B, Acosta-Martinez V, Lyte M. Memory and learning behavior in mice is temporally associated with diet-induced alterations in gut bacteria. Physiol Behav. 2009;96(4-5):557–567. doi: 10.1016/j.physbeh.2008.12.004. [DOI] [PubMed] [Google Scholar]
- 39.Diaz Heijtz R. Fetal, neonatal, and infant microbiome: Perturbations and subsequent effects on brain development and behavior. Semin Fetal Neonatal Med. 2016;21(6):410–417. doi: 10.1016/j.siny.2016.04.012. [DOI] [PubMed] [Google Scholar]
- 40.Kramer MS, Aboud F, Mironova E, Vanilovich I, Platt RW, Matush L, Igumnov S, Fombonne E, Bogdanovich N, Ducruet T, et al. Breastfeeding and child cognitive development: new evidence from a large randomized trial. Arch Gen Psychiatry. 2008;65(5):578–584. doi: 10.1001/archpsyc.65.5.578. [DOI] [PubMed] [Google Scholar]
- 41.Guxens M, Mendez MA, Molto-Puigmarti C, Julvez J, Garcia-Esteban R, Forns J, Ferrer M, Vrijheid M, Lopez-Sabater MC, Sunyer J. Breastfeeding, long-chain polyunsaturated fatty acids in colostrum, and infant mental development. Pediatrics. 2011;128(4):e880–889. doi: 10.1542/peds.2010-1633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Yatsunenko T, Rey FE, Manary MJ, Trehan I, Dominguez-Bello MG, Contreras M, Magris M, Hidalgo G, Baldassano RN, Anokhin AP, et al. Human gut microbiome viewed across age and geography. Nature. 2012;486(7402):222–227. doi: 10.1038/nature11053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Koenig JE, Spor A, Scalfone N, Fricker AD, Stombaugh J, Knight R, Angenent LT, Ley RE. Succession of microbial consortia in the developing infant gut microbiome. Proc Natl Acad Sci U S A. 2011;108(Suppl 1):4578–4585. doi: 10.1073/pnas.1000081107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Marques TM, Wall R, Ross RP, Fitzgerald GF, Ryan CA, Stanton C. Programming infant gut microbiota: influence of dietary and environmental factors. Curr Opin Biotechnol. 2010;21(2):149–156. doi: 10.1016/j.copbio.2010.03.020. [DOI] [PubMed] [Google Scholar]
- 45.Dominguez-Bello MG, Costello EK, Contreras M, Magris M, Hidalgo G, Fierer N, Knight R. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc Natl Acad Sci U S A. 2010;107(26):11971–11975. doi: 10.1073/pnas.1002601107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Yang I, Corwin EJ, Brennan PA, Jordan S, Murphy JR, Dunlop A. The infant microbiome: implications for infant health and neurocognitive development. Nurs Res. 2016;65(1):76–88. doi: 10.1097/NNR.0000000000000133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Glendinning L, Nausch N, Free A, Taylor DW, Mutapi F. The microbiota and helminths: sharing the same niche in the human host. Parasitology. 2014;141(10):1255–1271. doi: 10.1017/S0031182014000699. [DOI] [PubMed] [Google Scholar]
- 48.Lee SC, Tang MS, Lim YA, Choy SH, Kurtz ZD, Cox LM, Gundra UM, Cho I, Bonneau R, Blaser MJ, et al. Helminth colonization is associated with increased diversity of the gut microbiota. PLoS Negl Trop Dis. 2014;8(5):e2880. doi: 10.1371/journal.pntd.0002880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Kreisinger J, Bastien G, Hauffe HC, Marchesi J, Perkins SE. Interactions between multiple helminths and the gut microbiota in wild rodents. Philos Trans R Soc London Ser B. 2015;370(1675). [DOI] [PMC free article] [PubMed]
- 50.Pearson MS, Tribolet L, Cantacessi C, Periago MV, Valero MA, Jariwala AR, Hotez P, Diemert D, Loukas A, Bethony J. Molecular mechanisms of hookworm disease: stealth, virulence, and vaccines. J Allergy Clin Immunol. 2012;130(1):13–21. doi: 10.1016/j.jaci.2012.05.029. [DOI] [PubMed] [Google Scholar]
- 51.Harnett W, Harnett MM. Molecular basis of worm-induced immunomodulation. Parasite Immunol. 2006;28(10):535–543. doi: 10.1111/j.1365-3024.2006.00893.x. [DOI] [PubMed] [Google Scholar]
- 52.Bancroft AJ, Hayes KS, Grencis RK. Life on the edge: the balance between macrofauna, microflora and host immunity. Trends Parasitol. 2012;28(3):93–98. doi: 10.1016/j.pt.2011.12.001. [DOI] [PubMed] [Google Scholar]
- 53.Su CW, Cao Y, Kaplan J, Zhang M, Li W, Conroy M, Walker WA, Shi HN. Duodenal helminth infection alters barrier function of the colonic epithelium via adaptive immune activation. Infect Immun. 2011;79(6):2285–2294. doi: 10.1128/IAI.01123-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Hewitson JP, Grainger JR, Maizels RM. Helminth immunoregulation: the role of parasite secreted proteins in modulating host immunity. Mol Biochem Parasitol. 2009;167(1):1–11. doi: 10.1016/j.molbiopara.2009.04.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.White RR, Artavanis-Tsakonas K. How helminths use excretory secretory fractions to modulate dendritic cells. Virulence. 2012;3(7):668–677. doi: 10.4161/viru.22832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Mendlovic F, Flisser A. Dendritic cells in the gut: interaction with intestinal helminths. J Biomed Biotechnol. 2010;2010:250563. doi: 10.1155/2010/250563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Dzik JM. Molecules released by helminth parasites involved in host colonization. Acta Biochim Pol. 2006;53(1):33–64. [PubMed] [Google Scholar]
- 58.Stoltzfus RJ, Chwaya HM, Tielsch JM, Schulze KJ, Albonico M, Savioli L. Epidemiology of iron deficiency anemia in Zanzibari schoolchildren: the importance of hookworms. Am J Clin Nutr. 1997;65(1):153–159. doi: 10.1093/ajcn/65.1.153. [DOI] [PubMed] [Google Scholar]
- 59.Lozoff B, Clark KM, Jing Y, Armony-Sivan R, Angelilli ML, Jacobson SW. Dose-response relationships between iron deficiency with or without anemia and infant social-emotional behavior. J Pediatr. 2008;152(5):696-702–702 631-693. doi: 10.1016/j.jpeds.2007.09.048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Olney DK, Kariger PK, Stoltzfus RJ, Khalfan SS, Ali NS, Tielsch JM, Sazawal S, Black R, Allen LH, Pollitt E. Development of nutritionally at-risk young children is predicted by malaria, anemia, and stunting in Pemba Zanzibar. J Nutr. 2009;139(4):763–772. doi: 10.3945/jn.107.086231. [DOI] [PubMed] [Google Scholar]
- 61.Blanton LV, Charbonneau MR, Salih T, Barratt MJ, Venkatesh S, Ilkaveya O, Subramanian S, Manary MJ, Trehan I, Jorgensen JM et al. Gut bacteria that prevent growth impairments transmitted by microbiota from malnourished children. Sci .2016;351(6275). [DOI] [PMC free article] [PubMed]
- 62.Gordon JI, Dewey KG, Mills DA, Medzhitov RM. The human gut microbiota and undernutrition. Sci Transl Med. 2012;4(137):137ps112. doi: 10.1126/scitranslmed.3004347. [DOI] [PubMed] [Google Scholar]
- 63.Subramanian S, Huq S, Yatsunenko T, Haque R, Mahfuz M, Alam MA, Benezra A, DeStefano J, Meier MF, Muegge BD, et al. Persistent gut microbiota immaturity in malnourished Bangladeshi children. Nature. 2014;510(7505):417–421. doi: 10.1038/nature13421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Goyal MS, Venkatesh S, Milbrandt J, Gordon JI, Raichle ME. Feeding the brain and nurturing the mind: Linking nutrition and the gut microbiota to brain development. Proc Natl Acad Sci U S A. 2015;112(46):14105–14112. doi: 10.1073/pnas.1511465112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.De Palma G, Collins SM, Bercik P, Verdu EF. The microbiota-gut-brain axis in gastrointestinal disorders: stressed bugs, stressed brain or both? J Physiol. 2014;592(14):2989–2997. doi: 10.1113/jphysiol.2014.273995. [DOI] [PMC free article] [PubMed] [Google Scholar]