

Motor actions may result from minimization of the deflection of the actual body configuration from the centrally specified referent body configuration, in the limits of neuromuscular and environmental constraints. The minimization process may maintain reaching trajectory and accuracy regardless of the number of body segments involved (motor equivalence), as confirmed in this study of reaching from standing in young healthy individuals. Results suggest that the referent control process may underlie motor equivalence in reaching.
Keywords: motor control, arm movement, redundancy, adaptation, compensation, interjoint coordination, synergy
Abstract
Motor actions may result from central changes in the referent body configuration, defined as the body posture at which muscles begin to be activated or deactivated. The actual body configuration deviates from the referent configuration, particularly because of body inertia and environmental forces. Within these constraints, the system tends to minimize the difference between these configurations. For pointing movement, this strategy can be expressed as the tendency to minimize the difference between the referent trajectory (RT) and actual trajectory (QT) of the effector (hand). This process may underlie motor equivalent behavior that maintains the pointing trajectory regardless of the number of body segments involved. We tested the hypothesis that the minimization process is used to produce pointing in standing subjects. With eyes closed, 10 subjects reached from a standing position to a remembered target located beyond arm length. In randomly chosen trials, hip flexion was unexpectedly prevented, forcing subjects to take a step during pointing to prevent falling. The task was repeated when subjects were instructed to intentionally take a step during pointing. In most cases, reaching accuracy and trajectory curvature were preserved due to adaptive condition-specific changes in interjoint coordination. Results suggest that referent control and the minimization process associated with it may underlie motor equivalence in pointing.
NEW & NOTEWORTHY Motor actions may result from minimization of the deflection of the actual body configuration from the centrally specified referent body configuration, in the limits of neuromuscular and environmental constraints. The minimization process may maintain reaching trajectory and accuracy regardless of the number of body segments involved (motor equivalence), as confirmed in this study of reaching from standing in young healthy individuals. Results suggest that the referent control process may underlie motor equivalence in reaching.
there are abundant ways of combining different degrees of freedom (DFs) of the body to reach the same motor goal in changing environmental conditions, an important feature called motor equivalency, permitting flexibility of motor behavior in everyday life (Bernstein 1967; Feldman 2015; Lashley 1951; Latash et al. 2007; Yang et al. 2007). This flexibility contributes to the variability in endpoint trajectories and joint rotations when movements to the same target are repeated. Motor equivalency is associated with another fundamental question posed by Bernstein (1967) of how the nervous system chooses a particular action from many possible actions each time a movement is made in order to meet the task demand (the redundancy problem).
Typically, motor equivalence is associated with coordinated joint rotations (synergies) that allow the system to reach the same motor goal in different conditions (Bernstein 1967; Lashley 1951; Massion 1992). The accumulated evidence starting from the work of Matthews (1959) and Asatryan and Feldman (1965) shows that synergies are not selected by the nervous system as such but emerge in spatial frames of reference (FRs) predetermined by task-specific variables that represent the origin (referent) points of these frames. Once a spatial FR is predetermined, synergies are dynamically formed in this FR due to interactions between neuromuscular elements within certain environmental constraints. This conceptual framework (referent control of actions) is a departure from traditional notions of direct preprogramming of synergies or other kinematic features of actions or requisite motor commands to muscles [i.e., electromyographic (EMG) patterns]. A similar approach has been considered in recent studies (Mattos et al. 2011, 2015).
Optimality principles are often used to solve problems in physics and biology (Rosen 2013). Considering human motor behavior, such principles can be formulated within the context of referent control in which the nervous system orchestrates numerous muscles of the body as a coherent unit (Feldman et al. 1998, 2015; Foisy and Feldman 2006; Lestienne et al. 2000; St-Onge and Feldman 2004). Specifically, the system can predetermine the referent body configuration (R) such that the activity of muscles changes depending on the deviation of the emergent actual body configuration (Q) from the R. In the absence of coactivation of agonist and antagonist muscles, the R represents a common threshold configuration, i.e., the body position at which muscles begin to be activated or deactivated. This common threshold configuration can be changed by central reciprocal influences on motoneurons (MNs) of agonist and antagonist muscles. In addition, by means of central cofacilitation of agonist and antagonist MNs (referent C command), muscles can be coactivated at position R and at adjacent positions within a range (R−, R, R+; Asatryan and Feldman 1965). The role of cofacilitation (referent C command) is multifaceted. It increases stability of posture and movement. Applied in combination with reciprocal influences on agonist and antagonist MNs, it initially benefits agonist and later antagonist muscles in accelerating and decelerating motion, respectively, thus complementing the R command in increasing the speed of movement (Pilon and Feldman 2006; Zhang et al. 2016).
The activity of muscles depends not only on the deviation of Q from R but also on the speed (rate) of changes in this deviation. The Q depends, in particular, on the referent C command, the body's inertia, external and muscle forces (torques) that are also affected by the anatomical arrangement of muscles, and reflex interactions between them (Archambault et al. 2005). The referent body configuration is specified by the brain in a task-specific way. If the resulting action is not satisfactory, the referent configuration is modified to minimize the movement error. At the level of multi-muscle control, the optimality principle can be defined as a process that tends to minimize, in the limits of internal and external constraints, the deflection of Q from R:
| (1) |
where the asterisk implies the possible dependency of the R on the speed of the deflection from Q.
This approach predicts that to produce a pointing movement to a remembered target (i.e., in the absence of vision), the system uses a subset of referent body configurations allowing movement of the endpoint (the tip of index finger) to the target in each trial with small variations across trials. Changes in the referent body configuration from this subset will result in a referent trajectory (RT). Because of the inertia of body segments, external forces, and dynamical properties of neuromuscular elements, the actual endpoint trajectory (QT) may not coincide with the referent one. Interactions between neuromuscular elements mentioned above tend to minimize the deviation of QT from RT such that the residual difference (QT − RT) is responsible for compensating external forces. In other words, the optimality principle does not imply that the system always reaches the absolute minimum (zero) at the end of the movement: as a rule, external and internal constraints (e.g., gravitational forces, additional external loads, and stability requirements) may preclude the system from reaching absolute minimum (Foisy and Feldman 2006). The minimization process also does not imply that the final RT represents the exact targeted position: RT is just a tool for reaching the remembered target. Similarly, the difference (QT − RT) does not represent a movement error: if QT reaches the target, the movement error is zero, even if QT − RT is different from zero.
While minimizing the difference between QT and RT, the endpoint tends to reach an equilibrium position (eQT) at which muscle and external forces (torques) can be balanced, although an absolutely static position can be achieved only transiently because of varying properties of multiple muscles, central and afferent influences on MNs. Such variations occur even during quiet standing when the center of body mass drifts from one point of transient balance to another (Zatsiorsky and Duarte 2000). Within these limitations, the nervous system can adjust the rate, direction, and extent of changes in RT such that the endpoint trajectory (QT) reaches the target without an error. In dynamical terms, the referent trajectory represents a moving attractor to which actual movement tends to converge (Ghafouri and Feldman 2001; Won and Hogan 1995).
Thus, in the framework of referent control, pointing movement emerges following a minimization process:
| (2) |
In other words, to produce pointing motion, the system gradually shifts RT to a location near the target, whereas QT deviates from RT because of gravity (Fig. 1). Transmitted to MNs, the difference between the actual and referent body configurations and its speed influence muscle activation thresholds (λ), thus producing motion of body segments (for details see Feldman 2015). The emergent motion tends to bring the endpoint to the final equilibrium position eQT. If this position misses the remembered target, RT is adjusted until the endpoint reaches the target.
Fig. 1.

Referent and actual body configurations during pointing from standing. Muscles are activated depending on the difference between the referent (R) and actual (Q) body configurations. The shift in R results in a change in the referent trajectory of the endpoint (RT), where subscript T refers to the endpoint trajectory. Under the influence of gravity (vertical arrow), the final actual body configuration and the final endpoint position of the trajectory (QT) are below their respective referent values. Muscles are activated to minimize the gap between the actual and referent states for both the body configuration and the endpoint position. Muscle activation proceeds until a balance is achieved between muscle and external forces (e.g., gravity).
It is assumed that, if necessary, the minimization process can involve not only the arm but also other body segments. This occurs during reaching to a target located beyond the arm length when changes in the referent trajectory are provided by appropriate alterations in R, which include trunk bending, to contribute to the endpoint displacement (Horak and Nashner 1986; Kaminski and Simpkins 2001; Rossi et al. 2002). The minimization rule (Eq. 2) thus suggests that the same referent trajectory and, as a consequence, the actual pointing trajectory can be produced regardless of the number of DFs involved. For example, for reaching targets located within arm reach in sitting subjects, the endpoint trajectory remained invariant whether trunk movement was produced intentionally or suddenly blocked by an electromechanical device (Adamovich et al. 2001; Archambault et al. 1999; Shaikh et al. 2014). Trajectory invariance was maintained by changing the arm interjoint coordination without changing the referent endpoint trajectory.
In everyday activity, reaching often occurs in dynamic situations that involve recruitment of additional body segments to regain balance despite perturbations. Specifically, we investigated whether the minimization rule applies to whole body actions, such as pointing to a target placed beyond the arm reach during standing in the presence of perturbations that force subjects to take a step to prevent falling. The experimental question was whether or not, in the presence of perturbations, the system can call on referent body configurations that not only preserve the referent and, as a consequence, the actual endpoint trajectory but also body balance and stability by involving additional DFs. Thus we tested the prediction of the minimization rule that in a task that involves enhanced stability requirements, pointing trajectory and accuracy would be preserved by appropriate adaptive modifications of upper and/or lower limb interjoint coordination. Alternatively, stability requirements in the case of perturbations may become incompatible with reaching the target, resulting in a substantial arm movement error. Preliminary results have been reported in abstract form (Tomita et al. 2015).
METHODS
Participants
Ten right-hand dominant healthy young subjects (age, 24 ± 5.0 yr; height, 165.8 ± 8.0 cm; body weight, 61.3 ± 14.8 kg) participated in the study after signing an informed consent form approved by the Ethics Committee of the Centre for Interdisciplinary Research in Rehabilitation of Montreal, in accordance with the Declaration of Helsinki. Participants had no musculoskeletal or neurological pathology that could affect task performance.
Experimental Setup
Participants stood with their feet placed at the shoulder width (Fig. 2, initial position). The foot position was marked on the floor to maintain the body location relative to the target throughout the experiments. The target light-emitting diode (LED) was placed in the participant's midsagittal plane at a distance of 130% of arm length measured from the acromion process to the tip of the index finger (endpoint) with the arm extended, at the height of the anterior superior iliac spine (ASIS) on the tested side (Fig. 2). The endpoint starting position was at the upper edge of the greater trochanter on the same side. In this position, the shoulder was extended to 30° and the elbow was flexed to about 90°.
Fig. 2.

Experimental conditions. In the initial position, participants reached a remembered target (open circle) placed beyond the arm's reach with the eyes closed before the arm movement onset. In most of the trials, no perturbation was applied because the board that could prevent hip flexion was moved away (free-hip condition). In 30% of randomly chosen trials, hip flexion was unexpectedly prevented by an electromagnetic device (blocked-hip condition), forcing subjects to take a step to prevent falling. In yet another experiment, subjects made an intentional step during reaching in the absence of perturbation (intentional-step condition).
Postural stability during pointing movements was challenged in randomly selected trials by preventing the backward motion of the buttocks during pointing, thus forcing subjects to take a step forward to regain balance. This was done with a perturbation device consisting of an electromagnet attached to the wall behind the subject at an adjustable height. The electromagnet was locked to an electromagnetic plate attached to a concave-shaped board just behind the subject. In the initial position, there was about a 1-cm gap between the buttocks and the board (Fig. 2). In 70% of trials, the electromagnet was switched off simultaneously with a “go” signal (beep), and the board swung backward without contacting the buttocks (Fig. 2, free-hip condition). In the remaining 30% of trials, the electromagnet remained locked, preventing the buttocks from moving backward and forcing an involuntary step forward to regain balance (Fig. 2, blocked-hip condition).
Experimental Procedures
Two experiments were done in a random order across subjects to determine the effects of involuntary or voluntary recruitment of additional degrees of freedom on trajectory characteristics. Before each trial, subjects visualized the target LED located on a retractable rod. They then closed their eyes and reached with their right arm to the remembered target at the go signal. The target was retracted with the go signal such that touching the target was avoided. Because the target was placed beyond arm length, subjects were forced to lean the trunk forward to reach the target. To accomplish this task, balance could be maintained either by flexing the knees or by moving the lower part of the body (buttocks) backward by hip flexion (hip strategy; Horak and Nashner 1986; Winter 1995). In experiment 1 (50 trials), reaching was done with eyes closed in two conditions: free hip (35 trials) and blocked hip (15 trials) using the electromagnet perturbation device as described above. Free-hip and blocked-hip trials were randomized. To ensure consistency, subjects were instructed to use the hip strategy to lean the trunk. In experiment 2 (15 trials), the electromagnetic device was not used, but instead subjects had to intentionally step forward in each trial (Fig. 2, intentional-step condition) while reaching the target with eyes closed. Rest periods of 5 min were provided between experiments to avoid fatigue. Before each experiment, subjects practiced reaching in all conditions with their eyes closed, and feedback on reaching accuracy was given on each trial (7–10 trials for each condition) to stabilize performance and minimize effects of learning.
Data Recording and Analysis
Arm, trunk, and leg movements were recorded using an optoelectronic, three-dimensional motion analysis system (Optotrak; NDI, Waterloo, ON, Canada; sampling rate, 100 Hz; trial duration, 5 s). Markers were placed on bony landmarks at the tip of the index finger (endpoint), the head of the ulna (wrist), lateral epicondyle (elbow), right and left acromion processes (shoulders), midsternum (trunk), ASIS (hip), patella (knee), and lateral malleolus (foot).
Raw data were interpolated for missing segments (3rd-order spline function) and filtered with a low-pass Butterworth filter (10 Hz). Because the endpoint displacement primarily occurred in the sagittal plane, we focused on sagittal components of displacements and velocities of the endpoint, trunk, and foot movements. Movement onsets and offsets were determined when the velocity of the each marker (endpoint, right acromion, foot) exceeded or fell below 5% of their peak velocities, respectively. In each trial, the timing of all kinematic events was determined relative to the endpoint movement onset. Trials in which subjects anticipated the forthcoming condition, for example, by taking a step in a free-hip trial, were excluded (1–2 trials/subject).
Primary outcome variables were the endpoint trajectory characteristics and final position accuracy. Trajectories were characterized by spatial (index of curvature, IC) and temporal variables (time to peak velocity, deceleration time). Secondary outcome variables characterized the kinematics of different segments (trunk, foot) and joints. In addition, the time at which the earliest change in joint angles occurred in response to the perturbation was determined (divergence point).
The endpoint trajectory shape was evaluated by computing the IC, defined as the ratio of the total length of the endpoint displacement to the shortest distance between its initial and final position. IC = 1 when the path is straight, and IC = 1.57 when the path is a perfect semicircle (Atkeson and Hollerbach 1985). Final endpoint position accuracy was computed by calculating the square root of the sagittal and vertical positions of the endpoint at movement offset in each condition (blocked hip, intentional step) and was compared with those in the free-hip condition.
To characterize changes in the body configuration, several joint angles were computed at two time points: 1) at endpoint movement offset and 2) after endpoint movement offset at the time of trunk movement offset. Shoulder flexion angle was computed as the sagittal angle between the trunk midline (determined from the line between the midpoint between the 2 shoulder markers and the sternal marker) and the vector between the right shoulder and elbow markers, where the arm alongside the body was 0°. The elbow angle was determined based on the dot product of vectors defined by the wrist, elbow, and shoulder markers, where 0° indicated full elbow extension. The hip angle was defined by the dot product of vectors defined by the right shoulder, hip, and knee markers, where the hip position in upright standing was defined as 0° and forward hip motion was positive.
Trunk displacement in space in the sagittal and vertical directions was computed from the absolute movement of the right acromial marker, where the initial position was 0 cm. Trunk motion usually continued after the endpoint movement offset. To characterize this additional movement, we computed the amount of shoulder, elbow, and hip displacement between the endpoint movement offset and the trunk movement offset.
To determine when adaptations occurred in joint space during blocked-hip trials, we identified the earliest time when shoulder and/or elbow movement began to diverge from the control free-hip condition. Individual shoulder flexion and elbow flexion velocity/angle (phase) diagrams (mean ± 95% confidence intervals) were obtained for both free-hip and blocked-hip trials. Joint angles for each condition were aligned, averaged, and superimposed, and the point at which the blocked-hip phase diagram of the earliest joint (shoulder or elbow) deviated from the 95% confidence interval of that of the free-hip trials was identified. Because the perturbation did not coincide with arm movement onset, the time of perturbation was subtracted from the divergence point to identify the earliest time of angular divergence. Data analysis was performed using custom-made programs (MATLAB 2012b; The MathWorks, Natick, MA).
Statistical Analysis
The normality of data distribution was tested for each outcome with Shapiro-Wilk tests. One-way repeated-measures ANOVAs and paired t-tests with Bonferroni corrections (α/2) were used for the outcomes with normal distributions (accuracy, IC, time to peak velocity, deceleration time, trunk displacement, joint angle displacements) to estimate pairwise differences between conditions (free hip, blocked hip, intentional step). For outcomes with nonnormal distributions, Friedman and Wilcoxon signed rank tests with Bonferroni corrections (α/2) were used. Hodges-Lehman tests were used to estimate 97.5% (significance level corrected) confidence intervals (CI) of median values of the condition-dependent differences. The possible presence of learning effects was tested by comparing trajectory curvature (IC) and movement accuracy between the first and subsequent trials for both blocked-hip and intentional-step conditions.
For accuracy, we considered deviations of endpoint position in blocked-hip or intentional-step compared with free-hip conditions >5 cm as meaningful for the following reason. An involuntary or voluntary step resulted in additional trunk motion of 30–50 cm. If not compensated, this motion would have produced an equal amount of endpoint movement error. Deviations of up to 5 cm in the blocked-hip condition would represent 0–10% of the total potential deviation in the other two conditions. Thus deviations >5 cm were considered to be greater than minimal levels. SPSS (version 20) software was used for statistical analyses. Initial statistical significance levels of P < 0.05 were used for all tests.
RESULTS
Trajectory Characteristics and Movement Accuracy (Group Analysis)
Blocked-hip compared with free-hip condition.
Group mean values for spatial (IC) and temporal (time to peak velocity, deceleration time) endpoint trajectory characteristics did not significantly differ between blocked-hip and free-hip conditions (Tables 1 and 2). In addition, ICs did not exceed the functional criteria of greater than 3% deviation between conditions. Although there was a statistical difference in the endpoint accuracy in the blocked-hip condition (3.6 cm compared with free hip; Table 2), this difference was small and did not exceed a functionally significant 10% level.
Table 1.
Mean endpoint, trunk displacement, and joint angle variables for each condition
| Condition |
||||
|---|---|---|---|---|
| Free hip | Blocked hip | Intentional step | Significance | |
| Trajectory curvature, IC | 1.06 ± 0.03 | 1.07 ± 0.03 | 1.04 ± 0.03 | χ2 = 7.400† |
| Time to peak velocity, ms | 358.1 ± 13.1 | 382.5 ± 67.9 | 369.9 ± 21.9 | F2,18 = 6.305* |
| Deceleration time, ms | 446.4 ± 44.5 | 596.7 ± 79.3 | 734.2 ± 64.2 | F2,18 = 14.664* |
| Accuracy, cm | 140.4 ± 13.6 | 144.1 ± 11.4 | 144.6 ± 10.9 | F2,18 = 5.582* |
| Endpoint offset | ||||
| Trunk displacement, cm | 29.2 ± 14.2 | 45.9 ± 14.2 | 52.9 ± 15.0 | F2,18 = 91.993* |
| Shoulder flexion, ° | 126.7 ± 13.5 | 83.0 ± 17.0 | 31.5 ± 9.1 | F2,18 = 145.591* |
| Elbow flexion, ° | 33.3 ± 16.0 | 54.6 ± 22.4 | 58.6 ± 28.9 | F2,18 = 10.499* |
| Hip flexion, ° | 63.1 ± 9.8 | 54.8 ± 16.9 | 13.4 ± 12.9 | F2,18 = 42.100* |
| After endpoint offset | ||||
| Trunk displacement, cm | 0.6 ± 0.9 | 24.9 ± 5.8 | 27.8 ± 8.6 | F2,18 = 78.187* |
| Shoulder flexion, ° | 1.4 ± 0.6 | 20.6 ± 11.8 | 4.6 ± 3.5 | F2,18 = 3.794 |
| Elbow flexion, ° | 0.7 ± 0.8 | 12.6 ± 8.4 | 1.0 ± 1.4 | F2,18 = 3.593 |
| Hip flexion, ° | 0.2 ± 0.5 | 10.6 ± 5.3 | 3.9 ± 1.3 | F2,18 = 11.057* |
Values are means ± SD of endpoint, trunk displacement, and joint angle variables for each condition. IC, index of curvature. Overall results of multiple comparison analyses (ANOVA, Friedman's test) are shown at far right. Results of post hoc comparisons are shown in Table 2.
P < 0.05, 1-way repeated-measures ANOVA (F).
P < 0.05, Friedman test (χ2).
Table 2.
Mean and median differences and 97.5% CI of endpoint, trunk displacement, and joint angles in blocked-hip and intentional-step compared with free-hip conditions
| Blocked Hip − Free Hip | Intentional Step − Free Hip | |
|---|---|---|
| Trajectory curvature, IC | 0.8 (−0.9, 2.8) | −1.4† (−3.7, −0.4) |
| Time to peak velocity, ms | 24.4 (−19.9, 68.8) | 11.8* (2.2, 21.4) |
| Deceleration time, ms | 150.3 (−19.8, 320.4) | 287.8* (138.0, 437.6) |
| Accuracy, cm | 3.6* (0.2, 7.1) | 4.2* (0.5, 7.8) |
| Endpoint offset | ||
| Trunk displacement, cm | 12.0* (5.9, 18.0) | 14.1* (6.4, 21.8) |
| Shoulder flexion, ° | −43.7* (−54.7, −32.8) | −95.2* (−113.1, −77.3) |
| Elbow flexion, ° | 21.3* (7.1, 35.4) | 25.3* (5.8, 44.7) |
| Hip flexion, ° | −8.3 (−20.3, 3.6) | −49.7* (−65.8, −33.6) |
| After endpoint offset | ||
| Trunk displacement, cm | 20.4* (14.6, 26.2) | 17.4* (8.8, 26.0) |
| Shoulder flexion, ° | 18.0 (−1.9, 38.0) | 5.5 (−0.8, 11.8) |
| Elbow flexion, ° | 10.8 (−0.3, 22.0) | 1.5 (−1.5, 4.5) |
| Hip flexion, ° | 9.5 (−0.5, 19.6) | 6.7* (2.2, 11.1) |
Values are mean and median differences as well as their 97.5% confidence intervals (CI; corrected for multiple comparisons) of endpoint, trunk displacement, and joint angles in blocked-hip and intentional step compared with free-hip conditions. Results of post hoc comparisons are indicated.
P < 0.025, post hoc paired t-test with Bonferroni correction (α = 0.025): mean difference (97.5% CI).
P < 0.025, post hoc Wilcoxon signed rank test with Bonferroni correction (α = 0.025): median difference (97.5% CI).
Intentional-step compared with free-hip condition.
When the use of additional DFs was caused by taking an intentional step, spatial endpoint trajectory and accuracy characteristics of the group differed by small amounts between conditions. Paired comparisons showed that endpoint trajectories were less curved by 1.4% (Table 2) and that accuracy differed by 4.2 cm in intentional-step compared with free-hip conditions. Differences in both measures did not exceed the 3% and 10% functionally meaningful criteria. Taking an intentional step, however, led to significantly longer times to peak velocity (11.8 ms) and deceleration times (287.8 ms) of the endpoint trajectory.
Trajectory Characteristics and Movement Accuracy (Individual Analysis)
Endpoint trajectory invariance is shown for two subjects in Figs. 3 and 4. Trajectory invariance was maintained despite condition-specific differences in spatial and temporal recruitment of different DFs. For example, in both subjects, the trunk and foot movements differed, whereas spatial (Figs. 3A and 4A) and temporal (Figs. 3B and 4B) characteristics of the trajectory were invariant.
Fig. 3.

Spatiotemporal profiles of motion of body segments in different conditions in a representative subject (S1). The endpoint trajectories (A) and velocity profiles (B) remained similar, regardless of substantial motion of the trunk (shoulder marker) and foot. In A, circles indicate the final endpoint position and the instantaneous trunk position at that moment. In B, the circle and the vertical line indicate velocities of different body segments at the time of the endpoint movement offset. B shows that trunk motion exceeded the minimum (horizontal line) required for reaching the target; trunk position at the endpoint movement offset was more anterior in the blocked-hip and intentional-step conditions than the in free-hip condition. In B, the trunk continued to move after the endpoint movement offset, especially in the blocked-hip and intentional-step conditions.
Fig. 4.

Spatiotemporal profiles of motion of body segments in different conditions in another subject (S2). Like in S1 (Fig. 3), endpoint trajectories and velocity profiles in S2 were preserved across conditions (A). The velocity profile was symmetrical in S1. In contrast to S1, deceleration phases in S2 were prolonged in all conditions, resulting in an asymmetrical velocity profile (B). Trunk displacement after the endpoint movement offset (vertical line) in S2 was smaller than in S1.
There were no condition-dependent changes in trajectory characteristics in blocked-hip compared with free-hip conditions in any of the subjects. The small group difference in trajectory characteristics in intentional-step compared with free-hip conditions may have been due to differences observed in 2 of the 10 subjects. In one subject, represented in Fig. 5, trajectory IC differed by 3.9% in intentional-step compared with free-hip conditions, which exceeded the 3% threshold. In the second subject, the difference was 6.6%.
Fig. 5.
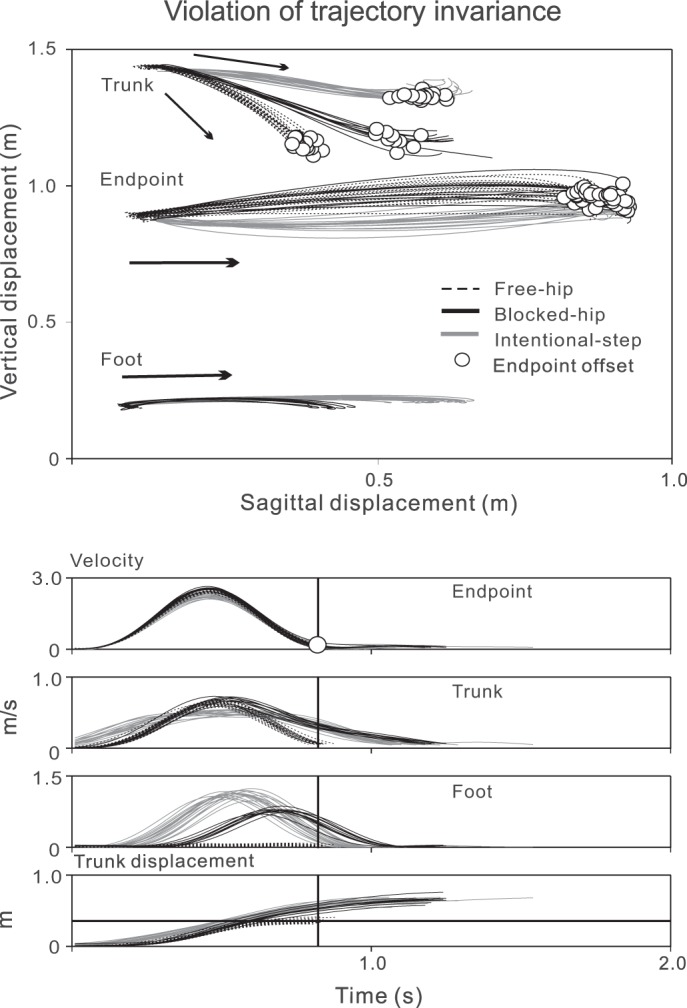
An example of violations of trajectory invariance. The endpoint trajectories in the free-hip and blocked-hip conditions were similar but different from those in the intentional-step condition, although the endpoint movement accuracy was preserved in all conditions (subject S3).
Mean endpoint error did not exceed 5 cm (Table 2) in 8 of the 10 subjects for blocked-hip conditions and in 6 of 10 subjects for the intentional-step conditions. The mean error was 8.8 cm for 2 subjects in blocked-hip conditions and 7.1 cm for 4 subjects in intentional-step conditions.
Trajectory curvature and movement accuracy did not change between the first and subsequent trials for the blocked-hip and intentional-step conditions. For the blocked-hip condition, curvature and accuracy in the first trial were respectively 1.07 and 145.0 cm, which were within the means ± 2SD of subsequent trials (1.07 ± 0.06 and 144.0 ± 5.4 cm, respectively). Similarly, trajectory curvature and movement accuracy in the first trial of the intentional step condition were respectively 1.04 and 143.4 cm, which were within the means ± 2SD of the subsequent trials (1.04 ± 0.03 and 144.7 ± 5.9 cm, respectively).
Temporal and Spatial Recruitment of Different Segments
Condition-specific differences in temporal and spatial recruitment of different DFs were greater for intentional-step compared with free-hip conditions than for blocked-hip compared with free-hip conditions for the whole group. The endpoint and trunk started to move simultaneously (Fig. 6), the foot started to move earlier in intentional-step compared with blocked-hip trials (Figs. 3B and 4B), and the trunk and foot continued to move after endpoint movement offset (Figs. 3B and 4B, vertical lines). Trunk movement time was significantly prolonged for blocked-hip and intentional-step compared with free-hip conditions (Fig. 6; F2,18 = 40.757, P < 0.001).
Fig. 6.

Timing of movement events (the onset, peak velocity, and offset) for different body segments during pointing (group data). For the endpoint, the timing (mean ± SD) remained similar across conditions. The trunk and endpoint movement onsets were not always synchronized, but in all cases the trunk continued to move after the endpoint movement offset, especially in the blocked-hip and intentional-step conditions. Foot movement was initiated earlier in the intentional-step condition compared with the blocked-hip condition.
Perturbation and intentional stepping led to additional trunk displacement, which exceeded 30 cm (32.4 ± 11.5 cm in blocked hip; 31.5 ± 17.3 cm in intentional step). This trunk movement exceeded that in the free-hip condition for the whole group and was greater than that needed for the hand to reach the target (Figs. 3B and 4B, bottom; horizontal lines). Trunk movement extent at endpoint movement offset was greater by 12.0 and 14.1 cm in blocked-hip and intentional step compared with free-hip conditions, respectively. The trunk moved about 20.4 cm farther in blocked-hip and about 17.4 cm farther in intentional step compared with free-hip conditions after endpoint movement offset (Figs. 3 and 4).
Condition-Specific Changes in Interjoint Coordination
Figures 7 and 8 show coordinated changes in shoulder, elbow, and hip angles (A) as well as individual changes of these angles in time (B) in two subjects, S1 and S2. In all conditions, the angles changed monotonically until the endpoint movement offset but the synergetic pattern varied depending on condition. This occurred whether or not movement trajectories and accuracy remained invariant. The coordination (Figs. 7A and 8A) between movements of the shoulder, elbow, and hip was initially the same in blocked-hip and free-hip conditions and later started to differ, whereas that between intentional-step and free-hip conditions differed from the very beginning of the movement. The coordination continued to change after the endpoint movement offset (circles in Figs. 7 and 8). S1 showed a large joint displacement in elbow, shoulder, and hip joints after the endpoint movement offset (Fig. 7), whereas joint angle changes in S2 were small (Fig. 8). Reversals in movement direction in specific joints (i.e., elbow) after endpoint offset could also occur in blocked-hip and intentional-step conditions (Figs. 7 and 8).
Fig. 7.

Condition-dependent interjoint coordination (A) and individual changes in the joint angles (B) during pointing (subject S1). In A, for better comparison, the initial points in the angular trajectories were adjusted to be matched. In B, each solid line indicates the mean and dotted lines the 95% confidence interval for changes in elbow, shoulder, and hip joint angles. In the free-hip and blocked-hip conditions, the interjoint coordinations began to diverge some time after the motion onset (A). In A, the interjoint coordination during the intentional-step condition began to diverge from the very beginning from coordinations during the other conditions. Interjoint coordinations at the endpoint movement offset (circles) were substantially different for all 3 conditions. In other words, the invariance of endpoint trajectories and movement accuracy were maintained due to condition-dependent changes in the interjoint coordination. Interjoint coordination continued to change after the endpoint movement offset (circles), indicating that the changes in the joint angles during this period neutralized the influence of continuous body motion on the endpoint position. Similarly, the divergence of individual joint displacement occurred before the endpoint movement offset (B).
Fig. 8.

Interjoint coordination during pointing in another subject (S2). Conventions are as described in Fig. 7. Changes in elbow, shoulder, and hip joint angles after the endpoint movement offset (A, circles) were smaller than in S1. The influence of trunk displacement on endpoint was not completely neutralized by changing interjoint coordination, resulting in prolonged deceleration time of the endpoint (B).
The final joint angles at endpoint movement offset were different across conditions (Figs. 7B and 8B; Table 2). Compared with movement in the free-hip condition, the shoulder was more extended by 43.7° (P < 0.01) and the elbow was more flexed by 21.3° (P < 0.01) in the blocked-hip condition (Table 2), suggesting that the potential influence of the unexpected involvement of additional DFs on the endpoint motion was neutralized (Table 2).
For the group, based on the time of divergence of arm joint phase diagrams between blocked-hip and free-hip conditions, changes in arm joints (i.e., divergence point) started to occur at 178.0 ± 66.8 ms after the perturbation onset in either the elbow or shoulder joint. The earliest divergence occurred at the shoulder joint in 60% of subjects. The divergence point corresponded to 42.6 ± 24.8% of the total endpoint displacement and to 99.0 ± 18.1% of the time to endpoint peak velocity.
DISCUSSION
Basic Findings
The experiments were designed to test the hypothesis that pointing results from minimization of the deviation of the emergent actual endpoint position from the referent position specified by neural control levels. According to this hypothesis, the system can use referent body configurations that preserve the referent endpoint trajectory while producing condition-specific changes in interjoint coordination to minimize changes in endpoint kinematics (trajectory curvature, accuracy, and velocity profile) despite substantial changes in the number of DFs elicited by a step. We compared reaching trajectories, accuracy, velocity profiles of body segments, and interjoint coordination in these conditions.
The experimental question was whether or not, in the presence of perturbations, the system can call upon referent body configurations that not only preserve body balance by involving additional DFs but also preserve the endpoint trajectory. Results support the hypothesis, because substantial changes in interjoint coordination that tended to minimize condition-dependent modifications in endpoint kinematics were observed in all subjects. In other words, the minimization strategy allowed the system to reach the motor goal even in cases when body stability was challenged and stepping occurred. Indeed, this result does not imply that the capacity to accommodate different task constraints in reaching the same motor goal (motor equivalency) is limitless; in everyday life, stumbling can elicit a fall, preventing reaching for the desired object.
We first discuss our findings in the context of referent control and then address the question of whether alternative theoretical approaches to motor control can explain the present findings.
Unperturbed Motion to a Target Placed Beyond the Arm Reach
Similar to our findings of reaching from standing (Fig. 6), the trunk and endpoint moved practically simultaneously when sitting subjects reached for a target placed beyond the arm reach (Saling et al. 1996). However, using electromagnet trunk perturbation methods during reaching from sitting, Rossi et al. (2002) showed that the potential influence of the trunk and shoulder motion on the endpoint position was initially neutralized by appropriate changes in arm joint angles: by an adaptive arm-trunk synergy (Ma and Feldman 1995). The adaptation occurred up until approximately the endpoint peak velocity, after which the trunk began to contribute to the endpoint displacement. The time of the endpoint peak velocity is also the time at which referent shifts underlying endpoint control of arm DFs are completed (Ghafouri and Feldman 2001), even though actual endpoint movement can continue for some time. One can assume that the full extent of referent endpoint shifts by the arm control system resembles the anatomical limit of arm extension (Rossi et al. 2002; Shaikh et al. 2014). Thus, at the time when the endpoint velocity reaches its maximum, referent guidance of endpoint shifts is relayed from the arm to the trunk control system. In other words, although the arm and trunk move in parallel, the contributions of these body segments to the endpoint movement extent at a control, referent level are accomplished sequentially.
A strategy of sequential referent control similar to that described during pointing in sitting subjects (Rossi et al. 2002) was likely used during pointing in standing subjects in our study. In the case of standing reaches beyond arm length, the movement duration and endpoint velocity profiles in free-hip trials could depend on the duration of the transfer of referent control from the arm to the trunk: an increase in the time of transfer could prolong the endpoint movement duration and velocity profile, which can result in subject-dependent differences in velocity characteristics (Figs. 3 and 4).
In our experiments, as in sitting subjects, the trunk continued to move after the endpoint reached its final position (Fig. 6), showing that adaptive changes in the arm joint angles in free-hip trials (adaptive arm-trunk synergy) were also accomplished after the endpoint stopped moving. The influence of trunk displacement on the endpoint position during this period was neutralized by appropriate changes in interjoint coordination (Figs. 7 and 8). Thus the trunk motion contributed to the endpoint motion extent during the period between the hand peak velocity and the hand motion offset, and was neutralized by changes in interjoint coordination before and after this period.
Maintaining Endpoint Trajectory and Accuracy When Additional DFs are Involved in the Task
Unexpectedly preventing hip flexion challenged postural stability. In both blocked-hip and intentional-step conditions, a substantial sagittal translation of the whole body occurred (Figs. 3 and 4), requiring adaptive interjoint coordination to maintain reaching accuracy and trajectory curvature. Specifically, in trials involving a step (Fig. 7), additional forward trunk displacement occurred exceeding 30 cm. Transmitted to the endpoint, this displacement would result in a substantial movement error and a change in the endpoint velocity profile. These potential modifications of the endpoint kinematics were, however, largely neutralized by appropriate changes in the arm joint angles (Figs. 7 and 8). In all conditions, trunk motion continued after the endpoint movement offset (Figs. 3, 4, and 6), as did motion in the shoulder, elbow, and hip joints (Figs. 7 and 8; Table 1). Changes in interjoint coordination neutralized the influence of the continued trunk motion on the endpoint after it reached a final position.
Parametric Control of Motor Actions
The role of parameters in predetermining the system equilibrium position can be illustrated by a pendulum that oscillates, whereas its virtual, equilibrium position (the lowest point traversed by the pendulum mass) remains the same because it is defined by system parameters (the direction of gravity, coordinates of the pivot point, and the pendulum length), regardless of the current position and velocity of the pendulum mass. The equilibrium position is a dynamic concept: it plays the role of an attractor in the system behavior. To change the place where a pendulum oscillates, we do not apply forces to the pendulum mass but simply move its suspension point. Physically, this means that we change the pendulum's parameters: the coordinates of its pivot point. The forces that compel the pendulum to move and oscillate in the new place are not a matter of our concern; they emerge automatically.
Parametric control defined in the dynamic system theory states that although stable body postures are associated with a tendency to balance torques, the choice between different stable postures is not defined by torque, forces, or other kinetic or kinematic variables describing the motor outcome. Rather, it is defined by system parameters characterizing the intrinsic state of the neuromuscular system and can be changed independently of these variables. Because EMG levels depend on afferent feedback related, in particular, to muscle forces, they cannot be considered as parameters that predetermine the choice between different postures. This point is illustrated by findings that in near-isotonic conditions, tonic EMG levels of muscles can be similar at different arm postures (Foisy and Feldman 2006; Raptis et al. 2010).
Experimental evidence justifies the suggestion that changes in mechanical balance inherent in active human movements, including small movements during quiet standing (Zatsiorski and Duarte 2000), result from parametric control. It was initially Matthews (1959) who found that central and reflex muscle activation is controlled parametrically, by shifting the threshold lengths (λ) at which muscles begin to be recruited. Because this was demonstrated in decerebrated cats, validity of the results for intact animals and humans could not be assumed. In intact humans, Asatryan and Feldman (1965) found that at the single-joint level, muscle activation is controlled by setting and resetting the common threshold joint angle (R) and respective threshold muscle lengths at which MNs begin to be recruited such that muscles generate forces depending on the deflection of the muscle length from the threshold length (force-length characteristics). By shifting thresholds and thus the force-length characteristics in the spatial domain, the nervous system shifts the equilibrium point (EP) in the interaction between muscles and external forces. These findings motivated the formulation of the EP hypothesis (Asatryan and Feldman 1965) and later were confirmed in several laboratories (Capaday 1995; Davis and Kelso 1982; Feldman and Orlovsky 1972; Nichols and Steeves 1986), most recently by Turpin et al. (2016). In elegant experiments, Bizzi et al. (1982) demonstrated in monkeys that EP is shifted gradually, rather than in a steplike manner, during goal-directed motion. These experimental data also imply that the nervous system can change system parameters to shift balance in the spatial domain and thus initiate and control the motor action studied in the current task.
An explicit description of spinal circuitry underlying threshold control of muscle activation with resulting EP shifts has been proposed in several publications, starting with Feldman (1986). The proposed mechanism has not, as yet, been experimentally verified. However, it has been experimentally confirmed that as a threshold of muscle activation, R is modified by changing the subthreshold state of MNs via pre- and/or postsynaptic influences on them prior to changes in EMG signals (Capaday 1995; Feldman and Orlovsky 1972; Matthews 1959).
Being parametric, referent control guides the well-organized structure (plant) of interactions of neuromuscular elements between themselves and the environment. It has some features of a predictive, feedforward mechanism relying on existing properties of the plant in the absence of any internal model of the plant. It is assumed that the patterns of referent control can be memorized together with the associated conditions in which these patterns were successful in reaching the motor goal and reproduced during movement repetition. If conditions change, the patterns might become inadequate and require corrections. It has been empirically shown (Ghafouri and Feldman 2001) that shifts in R underlying fast arm pointing movements (peak velocity about 1 m/s) are completed about 200 ms before the offset of the resulting arm movement. The time gap between the ends of R and Q is sufficient to decide, based on sensory feedback, whether or not the R needs to be changed to prevent a possible error or initiate a new movement without waiting for the end of the previous one, which is helpful in generating rapid motor sequences, e.g., in piano playing and speech production. Such a feedforward mechanism is likely responsible for the ability of pianists to recognize and prevent an error before hitting a wrong key (Ruiz et al. 2009). Referent, threshold control can also be modified to accommodate changes in muscle force-length characteristics depending on temperature, fatigue, tonic state, and changes in the recruitment order of motor units, for example, during training in athletes. Adaptation can also be achieved by compensatory changes in the activation thresholds of other muscles involving other degrees of freedom (see Côte et al. 2002).
The feedforward feature of referent control allows the system to transfer body balance in space by shifting, in advance, the base of support (BOS) in the environment such that stability is not lost and falling is avoided when a step is made despite the COM moving outside of the previous BOS (Feldman et al. 2011). Physiologically, the interjoint adaptation is likely mediated by vestibulospinal and corticospinal pathways, given it is substantially reduced in subjects with vestibular deficiency (Sibindi et al. 2013) and in patients with stroke (Sambasivan et al. 2015). The latency of the adaptive reaction in joint space (178.0 ± 66.8 ms) in our study is consistent with a previous report that the latency of adaptation produced through vestibulospinal pathways in healthy subjects was 60–180 ms (Raptis et al. 2007).
The referent control strategy and the minimization rule associated with it only suggest that the system tends to neutralize the influence of additional DFs on the reaching arm, but it does not predict that minimization is always associated with complete neutralization of the influence of additional DFs on endpoint kinematics. The system only specifies, with trial-to-trial variations, a referent trajectory (RT) and allows MNs to generate muscle activity that tends to diminish the deviation of the actual endpoint trajectory (QT) from RT. Other factors, such as variations in the specification of RT, external forces, inertia of body segments, and history-dependent properties of neuromuscular elements not directly controlled by the nervous system can also change QT so that it does not coincide exactly with RT (Fig. 1). Therefore, the minimization rule postulates a tendency to maintain invariant endpoint kinematics, but this process may not be absolute. Indeed, we observed violations of invariance of endpoint kinematics in some conditions (Fig. 5). However, condition-dependent changes in interjoint coordination that tended to maintain the endpoint kinematics (Figs. 7 and 8) were apparent even in these cases.
Time to peak velocity changes according to the anticipated reaching extent and direction (e.g., Messier and Kalaska 1999). We observed longer times to peak velocity in the intentional-step condition compared with the free-hip condition, whereas it remained unchanged in the blocked-hip condition (Table 2). Our results suggest that compared with the planned and specified referent movement extent (Rt), the rate of change and direction of the endpoint movement were identical in free-hip and blocked-hip conditions but differed in intentional-step conditions. On the other hand, we observed prolonged endpoint deceleration time in the intentional-step compared with the free-hip condition (Table 2), suggesting greater adaptive movements when additional DFs associated with taking a step were recruited. Interestingly, the onset of adaptation in joint space occurred at around endpoint peak velocity, which is similar to the termination time of the shift of RT (Ghafouri and Feldman 2001).
Parametric control implies that referent variables are used as tools for action productions. For comparison, when changing the direction of car motion, one just turns the steering wheel to move the car in the desired direction without any computations. In this example, the steering wheel is a practical tool for reaching a specific goal. Similarly, the nervous system can choose appropriate patterns of multi-muscle coordination of thresholds as tools to produce motor actions. The referent control theory defines several such tools or forms of threshold (referent) control, particularly the referent body configuration, referent position of the hand in space, referent hand shape and aperture for grasping, referent body orientation in gravity, referent coactivation commands, etc.; these are practical tools employed in action-specific ways (Feldman et al. 2011; Feldman 2015). By selecting a subset of referent body configurations, the system can gradually change the referent position of the endpoint such that the actual endpoint will be driven to the desired object in space. The referent control theory relies on natural properties of system without inversing the causality inherent in physiological and physical laws.
It should be emphasized that referent patterns are condition and task specific, and therefore they are memorized, reproduced, and corrected by taking into account the external condition in which the motor task is produced. However, this strategy relies on a qualitative representation of external forces without building an internal computational model of them. The system can use referent variables as tools to reach the motor goal in a given force field, even if the nature of the field is unknown during movement repetition. Consider the effects of a velocity-dependent Coriolis force field on pointing movements (DiZio and Lackner 1995). During motion in such a field, the arm is forcefully deflected from the intended trajectory. A natural reaction to this unknown destabilizing force was to stop this deflection before the end of intended motion, resulting in errors in the movement extent and direction that were gradually corrected in subsequent trials. Suppose that the system could recognize that the force is velocity dependent, i.e., transient, and that consequently no active reaction is needed to let the hand eventually arrive at the intended final position. If this were the case, the optimal strategy would be to suppress active reactions to the force during learning trials to produce movements without errors. This passive strategy could also guarantee no movement error after the Coriolis force was removed. This was not the case: in the first trials in the absence of the force, subjects made a movement error in the opposite direction, showing that despite numerous repetitions of movements in the presence of the Coriolis force, the system was unable to recognize that the force was transient. The system simply dealt with the force in the absence of any adequate model of it. Indeed, basic effects of velocity-dependent Coriolis force fields on pointing movements have been explained and numerically simulated in the framework of the EP hypothesis (Feldman et al. 1998). In contrast, DiZio and Lackner (1995) and others (e.g., Hwang and Shadmehr 2005) assumed that the system builds an internal model of the force field in repeated trials and did not consider possible alternatives such as that described above. Unfortunately, the findings by Dizio and Lackner, although interesting, were mistakenly perceived as refuting the EP hypothesis. Indeed, criticisms of the referent control theory are welcome, especially if they could help advance the theory beyond its present boundaries, for example, by addressing the challenging question of global organization of referent control in the brain. Hitherto, criticisms just required some clarifications of the existing aspects of the theory. These criticisms, including those that mistakenly claimed rejection of the theory (DiZio and Lackner 1995), have been addressed in several publications (Feldman 1986; Feldman et al. 1998; Feldman and Latash 2005; Gribble et al. 1998; Ostry and Feldman 2003).
Can Our Findings be Explained by Computational Approaches to the Problem of Motor Equivalence?
A major alternative to the referent control theory is the computational approach that has its origins in robotics (Hollerbach 1982; Kawato 1999). This approach is based on the notion that by using internal inverse and forward models of the interaction of the body with the environment, the nervous system can precompute (preprogram) and directly specify motor outcomes in terms of mechanical variables and requisite muscle forces. This output is estimated from current and predicted sensory signals derived from an internal forward model. If the motor outcome is unsatisfactory, corrections of muscle forces are made based, in particular, on some optimality criteria defined by a cost function of kinematic and kinetic variables (Kawato 1999; Kuo 1995; Todorov and Jordan 2002; Wolpert and Ghahramani 2000). The optimal control theory by Todorov and Jordan (2002) only uses forward models, thus avoiding inverse-dynamic models of mechanical laws.
In computational approaches, muscle force is assumed to be a major control signal preprogrammed by the nervous system according to task demands. This assumption conflicts with principles of parametric control (see above). Computational models presume that, instead, the system directly produces muscle forces. However, forces do not change the posture to which the system is attracted. Therefore, when subjects are changing posture from standing to leaning forward and pointing in our task, in the absence of changing parameters, the stretch reflex and other posture-stabilizing mechanisms would resist deviations from the initial posture elicited by direct generation of muscle forces, making even optimal computations of muscle forces suboptimal. This also means that computational approaches fail to explain a basic aspect of behavior in our study: how the system transfers balance and stability from the initial to the final posture, as well as the observation of motor equivalency.
The resistance to motion that would result if muscle forces were directly specified by the nervous system illustrates the classical posture-movement problem that was originally formulated by von Holst (von Holst and Mittelstaedt 1950; von Holst 1954) and later discussed in several publications (Feldman and Latash 2005; Ostry and Feldman 2003; Stuart 2005). The posture-movement problem thus arises and cannot be solved in the computational approaches or in any theory postulating direct preprogramming of motor outcome by the nervous system.
Computational approaches do not account for empirical data in which the physiological parameters underlying the control of posture and movement have been identified (see above). In other words, internal model approaches do not take into account empirical data on how MNs, reflexes, posture, and movement are controlled, allowing the system to convert the posture-stabilizing to movement-producing mechanisms without any posture-movement problem.
There are additional drawbacks of computational approaches. First, some of them rely on inverse dynamics computations of muscle forces. They thus invert the causality inherent in mechanical laws in which forces cause changes in movement kinematics, not the other way around, conflicting with elementary physics (Ostry and Feldman 2003). Second, some computational models avoid inverse dynamics transformations of mechanical laws (Kuo 1995; Todorov and Jordan 2002), but they implicitly require inverse dynamics transformations of physiological laws. Specifically, such models compute forces on the basis of some optimality criteria without considering how the computed forces can be actualized by the nervous system. Forces are outputs of MNs and muscles, and to actualize the computed forces, they should be converted into requisite input signals to MNs, thus inverting the causality of physiological laws. Such a treatment of physiological laws is prohibited, because motoneuronal input/output functions are fundamentally irreversible due to such properties of MNs as thresholds, plateau potentials, and the ability for self-supported generation of spikes.
One can suggest that muscle activation thresholds required for reaching a specific endpoint position (a mapping problem) also require some computations associated with inverse dynamics problems characteristic of internal model approaches we criticize. To explain why this is not the case, consider, say, elbow flexion against some unknown spring load until a specific joint angle or/and torque is reached. Although R and Q at which equilibrium is established are different variables, they operate in the same spatial domain (positions of body segments). Unless movement is prevented (in isometric conditions), shifts in R cause shifts in equilibrium Q to which the system is attracted. Although Q depends on load characteristics (increasing, decreasing, or nonmonotonic), it follows the changes in R and is therefore monotonically related to R (Fig. 9), although the sensitivity (“gain”) of this relationship indeed depends on the load (load specific). Analytically, let f(Q) be the load and F(Q − R) the muscle torque-angle characteristic. At equilibrium, F(Q − R) = f(Q). By differentiating this equation, one can find that at equilibrium points, dQ/dR = (K − k)/K, where K and k are slopes (stiffness) of muscle and load characteristics, respectively. Stable balance necessitates K > k (Feldman 1986). Therefore, for stable equilibrium points, dQ/dR is always positive, regardless of the load characteristic. The nervous system can thus gradually change R in an open-loop way until sensory feedback signals that the desirable equilibrium joint angle and/or torque is achieved, even if the load characteristic is unknown. In other words, the system can specify the thresholds required for reaching the goal empirically, without any forward or inverse dynamics computations. Because, in fast movements, these shifts terminate long before the end of the resulting movement (Ghafouri and Feldman 2001), the system can initiate additional shifts in the R if the motion is going to be in error. This strategy is sufficient to reach the goal even in the first trial with the unknown spring load. The total changes in R can be memorized and quickly reproduced in subsequent trials to make movements without errors (one-trial learning; Richter et al. 2004; Weeks et al. 1996). Thus, because of their inconsistencies with physical and physiological laws, current computation-based approaches do not provide adequate explanations of how changes in posture are organized and controlled.
Fig. 9.

With referent control, the system can reach the desired goal even if external force characteristics are not known. Regardless of load characteristic (a, increasing; b, nonmonotonic; c, decreasing), changes in the equilibrium joint angle (ΔQ) are monotonically related to shifts of the referent angle (ΔR) at which active muscle torque (thick curves) begin to be generated. Filled circles are equilibrium points at which muscle and load forces are balanced. The nervous system can thus monotonically shift the referent position until the desired final torque and/or position are reached.
Optimality in Referent Control
The notion that referent control only relies on empirical neuromuscular processes results in a quite different view of behavioral optimality, compared with computational approaches. To clarify, consider laws of mechanics. They are associated with the known optimality “principle of least action.” It would be meaningless to say that “to move, stones compute their optimal trajectories on the basis of this principle” or that “the law of conservation of energy is based on hidden computations.” Similarly, there is the smoothness criterion characteristic of movements of humans and animals (Hogan and Flash 1987). Mathematically, it is described as minimization of a function of high-order derivatives of hand displacement (“jerk”). Consistent with physiological findings, referent control implies that smoothness results from gradual muscle contractions complemented by gradual recruitment of MNs and neurons at all hierarchical levels without any computations. This means that the action optimality is a natural feature of actions; i.e., no specific effort or evaluation of cost functions by neural control levels is required to make actions optimal. The minimization principle in referent control theory reflects this view of behavioral optimality: physiologically, the neuromuscular system tends to minimize the difference between actual and referent values of variables at all levels empirically, without any computations. By means of threshold position control, the nervous system can only parametrically modulate this optimality without changing its nature.
Conclusions
Our results suggest that the minimization principle in the framework of referent control may be responsible for maintaining the invariance of the pointing trajectory and accuracy despite perturbations involving multiple DFs and affecting stability of posture and movement. Referent control may play a fundamental role in providing motor equivalence in pointing and other motor actions. In future studies, one can determine when the requirement to maintain balance after the perturbation may take priority over reaching the motor goal. It is possible that individuals with balance deficits may have impaired reaching accuracy when balance is threatened, due to prioritization of the maintenance of balance. Our observations are limited to the reaching task studied, but the validity of the minimization principle for other tasks can be determined in future studies.
GRANTS
This project was supported by National Sciences and Engineering Research Council (Canada). The doctoral studies of Y. Tomita are supported by a fellowship from the Japanese Student Support Organization and the Richard and Edith Strauss Musculoskeletal Doctoral Fellowship awarded by McGill University. M. F. Levin holds a Tier 1 Canada Research Chair in Motor Recovery and Rehabilitation.
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the authors.
AUTHOR CONTRIBUTIONS
Y.T. and A.G.F. performed experiments; Y.T. and A.G.F. analyzed data; Y.T., A.G.F., and M.F.L. interpreted results of experiments; Y.T., A.G.F., and M.F.L. prepared figures; Y.T., A.G.F., and M.F.L. drafted manuscript; Y.T., A.G.F., and M.F.L. edited and revised manuscript; Y.T., A.G.F., and M.F.L. approved final version of manuscript.
ACKNOWLEDGMENTS
We thank Valeri Goussev and Christian Beaudoin for technical support and Fang Yang for help at the initial stage of this project.
REFERENCES
- Adamovich SV, Archambault PS, Ghafouri M, Levin MF, Poizner H, Feldman AG. Hand trajectory invariance in reaching movements involving the trunk. Exp Brain Res 138: 288–303, 2001. [DOI] [PubMed] [Google Scholar]
- Archambault P, Pigeon P, Feldman AG, Levin MF. Recruitment and sequencing of different degrees of freedom during pointing movements involving the trunk in healthy and hemiparetic subjects. Exp Brain Res 126: 55–67, 1999. [DOI] [PubMed] [Google Scholar]
- Archambault PS, Mihaltchev P, Levin MF, Feldman AG. Basic elements of arm postural control analyzed by unloading. Exp Brain Res 164: 225–241, 2005. [DOI] [PubMed] [Google Scholar]
- Asatryan DG, Feldman AG. Functional tuning of the nervous system with control of movements or maintenance of a steady posture: I. Mechanographic analysis for the work of the joint on execution of a postural task. Biophysics 10: 925–935, 1965. [Google Scholar]
- Atkeson CG, Hollerbach JM. Kinematic features of unrestrained vertical arm movements. J Neurosci 5: 2318–2330, 1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bernstein NA. The Coordination and Regulation of Movements. London: Pergamon, 1967. [Google Scholar]
- Bizzi E, Accornero N, Chapple W, Hogan N. Arm trajectory formation in monkeys. Exp Brain Res 46: 139–143, 1982. [DOI] [PubMed] [Google Scholar]
- Capaday C. The effects of baclofen on the stretch reflex parameters of the cat. Exp Brain Res 10: 287–296, 1995. [DOI] [PubMed] [Google Scholar]
- Côté JN, Mathieu PA, Levin MF, Feldman AG. Movement reorganization to compensate for fatigue during sawing. Exp Brain Res 146: 394–398, 2002. [DOI] [PubMed] [Google Scholar]
- Davis WA, Kelso JA. Analysis of “invariant characteristics” in the motor control of Down's syndrome and normal subjects. J Mot Behav 14: 194–211, 1982. [DOI] [PubMed] [Google Scholar]
- DiZio P, Lackner JR. Motor adaptation to Coriolis force perturbations of reaching movements: endpoint but not trajectory adaptation transfers to the nonexposed arm. J Neurophysiol 74: 1787–1792, 1995. [DOI] [PubMed] [Google Scholar]
- Feldman AG. Once more on the equilibrium-point hypothesis (lambda model) for motor control. J Mot Behav 18: 17–54, 1986. [DOI] [PubMed] [Google Scholar]
- Feldman AG. Referent Control of Action and Perception. New York: Springer, 2015. [Google Scholar]
- Feldman AG, Krasovsky T, Baniña MC, Lamontagne A, Levin MF. Changes in the referent body location and configuration may underlie human gait, as confirmed by findings of multi-muscle activity minimizations and phase resetting. Exp Brain Res 210: 91–115, 2011. [DOI] [PubMed] [Google Scholar]
- Feldman AG, Latash ML. Testing hypotheses and the advancement of science: recent attempts to falsify the equilibrium point hypothesis. Exp Brain Res 161: 91–103, 2005. [DOI] [PubMed] [Google Scholar]
- Feldman AG, Orlovsky GN. The influence of different descending systems on the tonic stretch reflex in the cat. Exp Neurol 37: 481–494, 1972. [DOI] [PubMed] [Google Scholar]
- Feldman AG, Ostry DJ, Levin MF, Gribble PL, Mitnitski AB. Recent tests of the equilibrium- point hypothesis (lambda model). Motor Control 2: 189–205, 1998. [DOI] [PubMed] [Google Scholar]
- Foisy M, Feldman AG. Threshold control of arm posture and movement adaptation to load. Exp Brain Res 175: 726–744, 2006. [DOI] [PubMed] [Google Scholar]
- Ghafouri M, Feldman AG. The timing of control signals underlying fast point-to-point arm movements. Exp Brain Res 137: 411–423, 2001. [DOI] [PubMed] [Google Scholar]
- Gribble PL, Ostry DJ, Sanguineti V, Laboissière R. Are complex control signals required for human arm movement? J Neurophysiol 79: 1409–1424, 1998. [DOI] [PubMed] [Google Scholar]
- Hogan N, Flash T. Moving gracefully: quantitative theories of motor control. Trends Neurosci 10: 170–174, 1987. [Google Scholar]
- Hollerbach JM. Computers, brains, and the control of movement. Trends Neurosci 5: 189–192, 1982. [Google Scholar]
- Horak FB, Nashner LM. Central programming of postural movements: adaptation to altered support-surface configurations. J Neurophysiol 55: 1369–1381, 1986. [DOI] [PubMed] [Google Scholar]
- Hwang EJ, Shadmehr R. Internal models of limb dynamics and the encoding of limb state. J Neural Eng 2: 266–278, 2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaminski TR, Simpkins S. The effects of stance configuration and target distance on reaching. I. Movement preparation. Exp Brain Res 136: 439–446, 2001. [DOI] [PubMed] [Google Scholar]
- Kawato M. Internal models for motor control and trajectory planning. Curr Opin Neurobiol 9: 718–727, 1999. [DOI] [PubMed] [Google Scholar]
- Kuo AD. An optimal control model for analyzing human postural balance. IEEE Trans Biomed Eng 42: 87–101, 1995. [DOI] [PubMed] [Google Scholar]
- Lashley KS. The problem of serial order in behavior. In: Cerebral Mechanisms in Behavior, edited by Jeffress LA. New York: Wiley, 1951. [Google Scholar]
- Latash ML, Scholz JP, Schöner G. Toward a new theory of motor synergies. Motor Control 11: 276–308, 2007. [DOI] [PubMed] [Google Scholar]
- Lestienne FG, Thullier F, Archambault P, Levin MF, Feldman AG. Multi-muscle control of head movements in monkeys: the referent configuration hypothesis. Neurosci Lett 283: 65–68, 2000. [DOI] [PubMed] [Google Scholar]
- Ma S, Feldman AG. Two functionally different synergies during arm reaching movements involving the trunk. J Neurophysiol 73: 2120–2122, 1995. [DOI] [PubMed] [Google Scholar]
- Massion J. Movement, posture and equilibrium: interaction and coordination. Prog Neurobiol 38: 35–56, 1992. [DOI] [PubMed] [Google Scholar]
- Matthews PB. A study of certain factors influencing the stretch reflex of the decerebrated cat. J Physiol 147: 547–564, 1959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mattos D, Schöner G, Zatsiorsky VM, Latash ML. Motor equivalence during multi-finger accurate force production. Exp Brain Res 233: 487–502, 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mattos DJ, Latash ML, Park E, Kuhl J, Scholz JP. Unpredictable elbow joint perturbation during reaching results in multijoint motor equivalence. J Neurophysiol 106: 1424–1436, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Messier J, Kalaska JF. Comparison of variability of initial kinematics and endpoints of reaching movements. Exp Brain Res 125: 139–152, 1999. [DOI] [PubMed] [Google Scholar]
- Nichols TR, Steeves JD. Resetting of resultant stiffness in ankle flexor and extensor muscles in the decerebrated cat. Exp Brain Res 62: 401–410, 1986. [DOI] [PubMed] [Google Scholar]
- Ostry DJ, Feldman AG. A critical evaluation of the force control hypothesis in motor control. Exp Brain Res 153: 275–288, 2003. [DOI] [PubMed] [Google Scholar]
- Pilon JF, Feldman AG. Threshold control of motor actions prevents destabilizing effects of proprioceptive delays. Exp Brain Res 174: 229–239, 2006. [DOI] [PubMed] [Google Scholar]
- Raptis H, Burtet L, Forget R, Feldman AG. Control of wrist position and muscle relaxation by shifting spatial frames of reference for motoneuronal recruitment: possible involvement of corticospinal pathways. J Physiol 588: 1551–1570, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raptis HA, Dannenbaum E, Paquet N, Feldman AG. Vestibular system may provide equivalent motor actions regardless of the number of body segments involved in the task. J Neurophysiol 97: 4069–4078, 2007. [DOI] [PubMed] [Google Scholar]
- Richter S, Jansen-Osmann P, Konczak J, Kalveram KT. Motor adaptation to different dynamic environments is facilitated by indicative context stimuli. Psychol Res 68: 245–251, 2004. [DOI] [PubMed] [Google Scholar]
- Rosen R. Optimality Principles in Biology. New York: Springer, 2013. [Google Scholar]
- Rossi E, Mitnitski A, Feldman AG. Sequential control signals determine arm and trunk contributions to hand transport during reaching in humans. J Physiol 538: 659–671, 2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruiz MH, Jabusch HC, Altenmüller E. Detecting wrong notes in advance: neuronal correlates of error monitoring in pianists. Cereb Cortex 19: 2625–2639, 2009. [DOI] [PubMed] [Google Scholar]
- Saling M, Stelmach GE, Mescheriakov S, Berger M. Prehension with trunk assisted reaching. Behav Brain Res 80: 153–160, 1996. [DOI] [PubMed] [Google Scholar]
- Sambasivan K, Haentjens K, Khanafer S, Subramanian SK, Banina MC, Feldman AG, Sveistrup H, Levin MF. Coordination of arm-trunk reaching movement in stroke. Int J Stroke 10, Suppl 4: 99, 2015.25635277 [Google Scholar]
- Shaikh T, Goussev V, Feldman AG, Levin MF. Arm-trunk coordination for beyond-the-reach movements in adults with stroke. Neurorehabil Neural Repair 28: 355–366, 2014. [DOI] [PubMed] [Google Scholar]
- Sibindi TM, Krasovsky T, Feldman AG, Dannenbaum E, Zeitouni A, Levin MF. Arm-trunk coordination as a measure of vestibulospinal efficiency. J Vestib Res 23: 237–247, 2013. [DOI] [PubMed] [Google Scholar]
- St-Onge N, Feldman AG. Referent configuration of the body: a global factor in the control of multiple skeletal muscles. Exp Brain Res 155: 291–300, 2004. [DOI] [PubMed] [Google Scholar]
- Stuart DG. Integration of posture and movement: contributions of Sherrington, Hess, and Bernstein. Hum Mov Sci 24: 621–643, 2005. [DOI] [PubMed] [Google Scholar]
- Todorov E, Jordan MI. Optimal feedback control as a theory of motor coordination. Nat Neurosci 5: 1226–1235, 2002. [DOI] [PubMed] [Google Scholar]
- Tomita Y, Feldman AG, Levin MF. Motor equivalence during whole body reaching in healthy young adults. Program No. 67.20. 2015 Neuroscience Meeting Planner. Chicago, IL: Society for Neuroscience, 2015. Online. [Google Scholar]
- Turpin NA, Levin MF, Feldman AG. Implicit learning and generalization of stretch response modulation in humans. J Neurophysiol 115: 3186–3194, 2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- von Holst E. Relations between the central nervous system and the peripheral organs. Br J Anim Behav 2: 89–94, 1954. [Google Scholar]
- von Holst E, Mittelstaedt H. Das Reafferezprincip. Wechselwirkungen zwischen Zentralnerven-system und Peripherie. Naturwissenschaften 37: 467–476, 1950. The reafference principle. In: The Behavioral Physiology of Animals and Man. The Collected Papers of Erich von Holst, edited and translated by Martin R. Coral Gables, FL: University of Miami Press, 1973, p. 139–173, 176–209. [Google Scholar]
- Weeks DL, Aubert MP, Feldman AG, Levin MF. One-trial adaptation of movement to changes in load. J Neurophysiol 75: 60–74, 1996. [DOI] [PubMed] [Google Scholar]
- Winter DA. Human balance and posture control during standing and walking. Gait Posture 3: 193–214, 1995. [Google Scholar]
- Wolpert DM, Ghahramani Z. Computational principles of movement neuroscience. Nat Neurosci 3: 1212–1217, 2000. [DOI] [PubMed] [Google Scholar]
- Won J, Holgan N. Stability properties of human reaching movements. Exp Brain Res 10: 125–136, 1995. [DOI] [PubMed] [Google Scholar]
- Yang JF, Scholz JP, Latash ML. The role of kinematic redundancy in adaptation of reaching. Exp Brain Res 176: 54–69, 2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang L, Straube A, Eggert T. Under threshold position control, peripheral mechanisms compensate efficiently for small perturbations of arm movements. Motor Control 20: 87–108, 2016. [DOI] [PubMed] [Google Scholar]
- Zatsiorsky VM, Duarte M. Rambling and trembling in quiet standing. Motor Control 4: 185–200, 2000. [DOI] [PubMed] [Google Scholar]