

Abstract
We describe a protocol for measuring ethanol self-administration in fruit flies (Drosophila melanogaster) as a proxy for changes in reward states. We demonstrate a simple way to tap into the fly reward system, modify experiences related to natural reward, and use voluntary ethanol consumption as a measure for changes in reward states. The approach serves as a relevant tool to study the neurons and genes that play a role in experience-mediated changes of internal state. The protocol is composed of two discrete parts: exposing the flies to rewarding and nonrewarding experiences, and assaying voluntary ethanol consumption as a measure of the motivation to obtain a drug reward. The two parts can be used independently to induce the modulation of experience as an initial step for further downstream assays or as an independent two-choice feeding assay, respectively. The protocol does not require a complicated setup and can therefore be applied in any laboratory with basic fly culture tools.
Keywords: Neuroscience, Issue 118, Drosophila, behavior, alcohol, addiction, experience, preference, consumption, social, courtship, courtship suppression, mating.
Introduction
Modification of behavior in response to experience allows animals to adjust their behavior to changes in their environment1. During this process, animals integrate their internal physiological state with the changing conditions of the external environment and subsequently choose one action over another to increase their chances of survival and reproduction. Reward systems evolved to motivate behaviors that are required for the survival of individuals and species by reinforcing behaviors that enhance immediate survival, such as eating or drinking, or those that ensure long-term survival, such as sexual behavior or caring for offspring2. Artificial compounds such as drugs of abuse also affect reward systems by co-opting neural pathways that mediate natural rewards2.
During the last two decades, the fruit fly Drosophila melanogaster has been established as a promising model for studying the molecular and neuronal mechanisms that are shaping the effects of ethanol on behavior3,4.
Previously, we have identified a subset of peptidergic neurons in flies (NPF/NPF receptor (R) neurons) that couple natural rewards, such as sexual experience, to the motivation of obtaining drug rewards5. NPF expression is sensitive to both sexual experiences and to drug rewards, such as ethanol intoxication. Changes in NPF expression levels are converted to alterations in ethanol self-administration5, where high NPF reduces and low NPF increases the preference to consume ethanol. Activating NPF neurons is rewarding for flies, as they display strong preference for an odor paired with the activation, which is also reflected by reduced ethanol consumption. More importantly, activation of NPF neurons interferes with the ability of flies to form a positive association between ethanol intoxication and an odor cue. The causal link between the NPF/R system, reward memory, and ethanol consumption suggests that one can use ethanol self-administration as a measure for changes in reward states5.
In this publication we demonstrate an integrated approach for tapping into the fly natural reward system and assaying changes in reward states. The approach consists of two separate parts, a training protocol for manipulating natural reward-related experiences, followed by a two-choice capillary feeder assay (CAFE) to assess ethanol self-administration as an estimate for changes in reward states. The CAFE assay is analogous to the two-bottle choice assays used in rodent studies for drug self-administration and has been shown to reflect certain properties of addiction-like behavior in flies6.
Protocol
Note: General overview of the experimental design: The experimental design includes an adapted protocol for courtship suppression7-9 in which male flies are exposed to rewarding and nonrewarding experiences in 3 consecutive training sessions over the course of 4 d. At the end of the experience phase, the flies are tested in a two-choice voluntary ethanol consumption assay for 3 - 4 d. The protocol herein includes several preparatory steps, some of which can be done in advance to be used in more than one experiment, while others should take place in a timely manner before the beginning of the experiment (Table 1).
1. Preparatory Steps
- Preparing the capillary feeder system
- Heat a 27 G needle and use it to puncture small holes along a vial (Figure 1).
- Cut the vial plug in half (crosswise) using a razor blade. Mark four points on the surface of the plug, creating a square shape to position the capillary holders into. Use an 18 G needle to make 4 holes through the plug (Figure 1b).
- Use a razor blade to cut micropipets so that they can firmly hold 5 µL glass capillaries, which serve as adaptors.
- Insert 4 capillary adaptors into the prepared plugs.
- Mark the position of the 2 ethanol- and two nonethanol-containing capillaries on the plug. Note: It is recommended to keep the positions of the 2 types of capillaries consistent across all vials. Pay attention to the size of the holes, as large holes allow the escape of flies out of the vials. In addition, it is recommended to use the smoother side of the plug (usually opposite the cut side) as the side that faces down when inserted into the vial, as cutting the plug creates some small, loose fibers that the flies can get tangled in.
- Prepare a large number of vials and adaptors to be used for the capillary feeder system (CAFE) experiments described in step 2.3.
- Preparing small glass food vials
- A day before collecting the flies (step 1.4), arrange 200 glass vials on microcentrifuge tube racks. Melt fly food (according to the cornmeal molasses recipe, see the Table of Materials) using a microwave. Pour 2 mL of fly food into the glass vials (try not to smear the food on the vial walls). Let the food harden, and cover the vials with plastic wrap. NOTE: The glass vials containing the food can be kept at 4 °C for several weeks.
- On the day of fly collection, take the vials out of the refrigerator and let them reach RT before using them to house flies. NOTE: If food vials are several weeks old, make sure that the food is still intact and not detached from the vial, as flies can be trapped in the cracks between the food and the glass.
- Making stock food solution for the capillary feeder assay
- To prepare the stock food solution (5% sucrose and 5% yeast extract), dissolve 5.88 g of yeast extract and 5.88 g of sucrose in 100 mL of distilled water. NOTE: The stock solution is highly prone to contamination. Therefore, it is recommended to autoclave it (121 °C, 30 min, and without a drying cycle) and aliquot it into 850 µL working aliquots to be kept at 4 °C until further use. The concentration of the ingredients in the stock solution is 5.88% and will reach the final concentration of 5% after the addition of 15% ethanol or water (150 µL into 850 µL of premade food solution) just before its use in step 2.3.
- Collecting experimental flies
- Collect 200 naïve male flies using a CO2 pad. Note: Use any lab strain relevant for the experimental question. 250 flies can be easily collected from five bottles of flies at their eclosion pick phase. When collecting the flies, pay careful attention to collect flies that have recently eclosed (virgin-looking flies: bright flies with a meconium spot on their abdomen). Experimental flies should be kept singly in small glass vials containing food and aged to 3 - 4 d old before being used in the experiment.
- Use soft plugs to close the vials. NOTE: The same soft plugs will be very useful for the training phase when using mouth aspiration to insert females into and out of the vials during the experience phase.
- Collecting trainer flies NOTE: All collected flies should be kept in a 12 h/12 h light/dark incubator at 25 °C and 70% Relative Humidity (RH). The 12 h/12 h light/dark cycle can be changed to a convenient starting time.
- Virgin female trainer flies
- Collect 500 virgin female trainer flies from a WT stock or a virginator stock for use as virgin receptive females. Keep the female flies in groups of 25 per regular-size food vial, and allow them to age to 3 - 4 d old before using them in the experiment.
- Mated female trainer flies
- Collect 300 virgin female flies from a WT stock or a virginator stock to undergo mating before the training phase (step 1.6.1). NOTE: Female flies are kept in groups of 10 females per regular-size food vial and aged to 3 - 4 d old before being used in the experiment.
- Male flies used to generate mated female trainer flies
- Collect 150 male flies (not necessarily virgin). NOTE: Flies are kept in groups of 15 males per regular-size food vial and aged to 3 - 4 d old before being paired with virgin female flies to create the mated trainer females.
- Generating mated female trainer flies
- To generate mated female flies to be used as trainers, pair groups of 10 virgin female flies with 15 male flies by flipping one male-containing vial into a female-containing vial 16 - 18 h before the training phase (step 2.1)10. NOTE: This should normally take place the evening before the training session. The 1.5:1 ratio between male flies to female flies assures that all females will be mated.
- On the day of the experiment, separate the mated females from the males by aspirating the males out of the vial using a mouth aspirator. Perform this right before the beginning of each training day.
2. Experimental Steps
- Setting up the experience phase
- Start the experiment as close as possible to the beginning of the light phase in the incubator. Place the trainer females (both virgin and mated) in the behavior chamber. NOTE: It is recommended to perform the experiment in a behavior chamber, or any secluded environment that is quiet and set to 25 °C and 60 - 65% RH. Keeping humidity over 50% is very important, since courtship and mating behaviors depend on pheromone sensing, which is known to be sensitive to humidity.
- Exposing male flies to rewarding and nonrewarding experiences
- Arrange the glass vials containing the single male flies on microcentrifuge racks, positioning the vials along the margins of the rack. NOTE: This arrangement allows the experimenter to easily observe the pairs of flies that undergo copulation without taking the vials out of the rack.
- Prepare (as described in step 2.2.1) 2 groups of 100 glass vials containing single male flies on separate racks for males undergoing rewarding and nonrewarding mating experiences (i.e. rejected and mated).
- Aspirate trainer females and add them to each vial containing the male flies. Set the timer for 1 h. Starting with the rejected group, add a mated female to each vial. Then, move on to the males to be mated and add a virgin female to each vial.
- Watch the mating encounters closely to make sure that the males interacting with virgin females mate successfully within the first hour and that the males interacting with the previously-mated females do not mate. To monitor the rejected cohort over the course of 1 h, set a second timer for 15 min and watch the males every 15 min.
- Discharge the males from the rejected cohort that managed to mate and males from the mated cohort that did not end up mating by the end of the first training session.
- Finish the first training session by aspirating out the previously-mated female trainers into a regular fly food vial, and keep them for the next conditioning session. Follow this step by gently aspirating the females from the mated male group.
- Let the flies rest for 1 h.
- Repeat steps 2.2.3 - 2.2.7 twice.
- Repeat the training session in steps 2.2.3 - 2.2.8 each day for 3 more d.
- Recording consumption (CAFE)
- At the end of the last conditioning session (fourth day), aspirate the single males and group them into the capillary feeder vials. Use a soft plug to cover the capillary feeder vial while inserting the flies, and replace it quickly with the ready-made capillary feeder plug after all the flies have been inserted. Group 6 - 8 flies from each of the experimental groups into a vial, creating no less than 8 vials per condition.
- Wet the plugs by gently adding 5 mL of water to the top of the plug. Be careful while adding the water to avoid spillage into the vials. Add equal amount of water to each vial. NOTE: This step is done to reduce evaporation from the capillaries. Therefore, it is repeated every day for the rest of the experiment. Depending on the relative humidity, the following days may require less water to be added to the plugs.
- Prepare two separate food solutions by adding ethanol or water to the stock food aliquots to create a final concentration of 15%.
- To fill the capillaries with the two food solutions, create 5 µL liquid food drops on a laboratory film strip and allow the solution to fill the capillary until it reaches the 5 µL mark. Make sure to use the clear end of the capillary, which is close to the 5 µL black mark line. NOTE: To fill several capillaries at once, it is possible to connect 6 - 8 capillary adaptors together and use them as an insert to hold the capillaries.
- Tap the capillaries gently on the paraffin strip to position the food solution exactly at the black mark. Seal the capillaries by dipping the end of each capillary into small drops of mineral oil. This step minimizes potential evaporation of the food solution.
- Insert two ethanol-containing capillaries and two nonethanol containing capillaries through the 4 adaptors/holders in the plug until the exposed sides of the capillaries barely peek from the interior of the vial (1 mm or less from the plug surface). NOTE: Keeping the opening of the capillaries close to the wet plug reduces evaporation and allows easy access to the food, as flies normally sit on the plugs and climb down on the exposed capillary to feed.
- In addition to vials that contain experimental flies, prepare a mock vial without flies. This control serves to assess natural evaporation. NOTE: Try to maintain relatively equal positioning of the capillary opening compared to the plugs, as differences can lead to variable evaporation and to misleading consumption data. Maintain adequate humidity during the experiment and position the capillary openings as closely as possible to the wet plug, as low humidity and large distance from the wet plug can both result in uncontrolled evaporation.
- Record the relative position of the liquid in each of the capillaries with respect to the black mark line in mm using a ruler, and write these values down. NOTE: If the solution is positioned exactly at the black line, the value should equal zero, and if it is above or below, it should be positive or negative values, respectively. The initial levels of the liquid set the reference points for the final O/N consumption values.
- Set the vials back in the incubator for 24 h. Replace the capillaries at a constant time throughout the entire experiment.
- Remove the capillaries gently from the vials and measure the levels of liquid with respect to the black marking line.
- Carefully check the liquid level in the mock vial. NOTE: If the levels dropped more than 2 mm, it indicates that evaporation is high and the consumption levels are not reliable.
- Dampen the plugs gently with 2 mL of water and insert a new set of capillaries, as indicated in steps 2.3.3 - 2.3.10. Set the vials back in the incubator for 24 h.
- Determining preference
- Calculate the total consumption of each capillary by subtracting the final level from the initial level at the starting point.
- Combine the values of both capillaries with the same food (i.e. nonethanol capillaries separately from ethanol-containing capillaries).
- Calculate ethanol preference by subtracting the amount consumed from the non-ethanol capillaries from the amount consumed from the ethanol-containing capillaries and divide the value by the total consumption values from all the capillaries of the same vial.
- Average the preference indexes of all vials of the same experimental group.
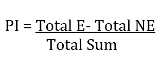 NOTE: Nonethanol 1 (NE1), Nonethanol 2 (NE2), Ethanol 1 (E1), Ethanol 2 (E2), Total nonethanol (Total NE) = NE1 + NE2, Total ethanol (Total E) = E1 + E2, Total sum consumption (Total Sum) = Total NE + Total E, and Ethanol preference Index (PI)
NOTE: Nonethanol 1 (NE1), Nonethanol 2 (NE2), Ethanol 1 (E1), Ethanol 2 (E2), Total nonethanol (Total NE) = NE1 + NE2, Total ethanol (Total E) = E1 + E2, Total sum consumption (Total Sum) = Total NE + Total E, and Ethanol preference Index (PI)
Representative Results
FPreviously, Devineni et al. showed that when fruit flies are given the choice to consume ethanol-containing food, they display a strong preference for ethanol-containing food over nonethanol containing food6. Shown here are some representative results we obtained when assaying the innate ethanol preference of naïve male flies that did not undergo the training protocol.
Naïve Canton S male flies were collected upon eclosion, aged until 4-day-old, and assayed for their innate preference to consume ethanol over the course of 4 d (Figure 2). Analyzing the amounts consumed from ethanol- and nonethanol-containing solutions shows that flies exhibit a robust preference for consuming 15% ethanol food over nonethanol food (Figure 2).
Using opposing sexual experiences, we have previously demonstrated that male flies that were allowed to mate in groups ("mated-grouped") display lower ethanol preference than singly-housed males that were rejected by previously-mated females ("rejected-isolated"). We used several controls to uncouple the housing experience from the mating-related experience and showed that the differential preference results from the mating experience5.
Using the training protocol described here, we generated singly-mated and rejected male flies that underwent similar housing and mating regimes. Single male Canton S flies were subjected over the course of 4 d to disparate mating experiences. One cohort of males was allowed to interact and mate with virgin female flies (single mated cohort), and the other cohort was trained with previously-mated female flies (single rejected cohort). Following the training phase, the single males from the two cohorts were grouped (8/vial) and placed in the capillary feeder vials; their voluntary ethanol consumption was recorded for 4 d (Figure 3A). The preference to consume ethanol was calculated from the consumption data using the formula in step 2.4 (Figure 3B). Positive values indicate preference for ethanol-containing food, and negative values indicate aversion to ethanol-containing food.
The present experiment supports our previous findings, suggesting that sexual experience, and not housing condition, modulates ethanol consumption. Successful mating increases internal reward levels, which in turn lowers ethanol consumption. Rejections, perceived as a lack of reward, lead to an increase in reward-seeking behavior (Figure 3B).
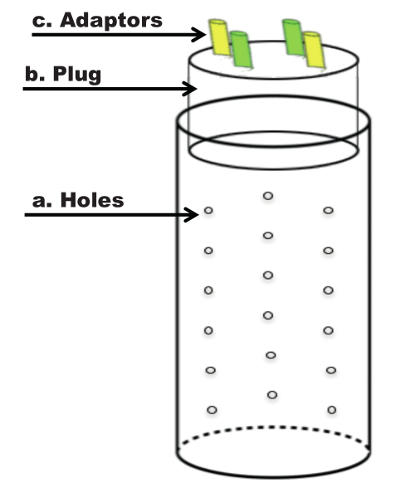 Figure 1. Schematic of the Capillary Feeder System (CAFE). (a) Schematics of the perforated vial with holes to equilibrate pressure and humidity. (b) Schematics of the cut-in-half plug that serves as the base for the insertion of the capillaries. (c) Schematics of shape and location of the adaptors serving to hold the capillaries. Please click here to view a larger version of this figure.
Figure 1. Schematic of the Capillary Feeder System (CAFE). (a) Schematics of the perforated vial with holes to equilibrate pressure and humidity. (b) Schematics of the cut-in-half plug that serves as the base for the insertion of the capillaries. (c) Schematics of shape and location of the adaptors serving to hold the capillaries. Please click here to view a larger version of this figure.
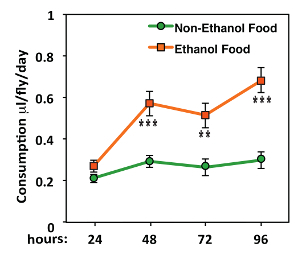 Figure 2. An Example of innate Preference to Consume Ethanol. Naïve WT males were grouped in groups of 8 males/vial, and their consumption of ethanol- and nonethanol-containing solutions were recorded over the course of 96 h. The flies consumed a larger amount of 15% ethanol containing food (**P <0.01, ***P <0.001, two-way repeated measures ANOVA with Bonferroni post-tests, n = 8). Data shown are the mean + SEM or mean - SEM. Please click here to view a larger version of this figure.
Figure 2. An Example of innate Preference to Consume Ethanol. Naïve WT males were grouped in groups of 8 males/vial, and their consumption of ethanol- and nonethanol-containing solutions were recorded over the course of 96 h. The flies consumed a larger amount of 15% ethanol containing food (**P <0.01, ***P <0.001, two-way repeated measures ANOVA with Bonferroni post-tests, n = 8). Data shown are the mean + SEM or mean - SEM. Please click here to view a larger version of this figure.
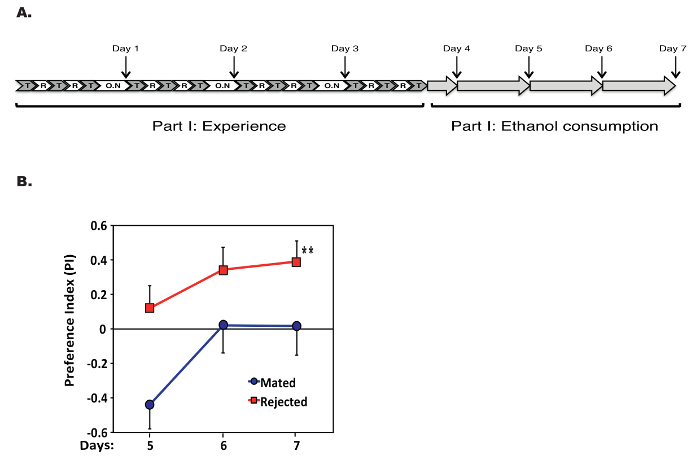 Figure 3. Opposing Mating Experiences mModulate Ethanol Preference. (A) Schematic of the combined protocol. Single virgin WT males are allowed to mate with virgin females or are subjected to 3x 1h courtship-suppression training sessions (depicted as "T") spaced by 1 h of rest (depicted as "R"). Training is repeated for 4 d. At the end of each day, the flies are placed in the incubator (depicted as "O.N"). After 4 d of training, the males were placed in vials where they could choose to feed from capillaries containing food solutions with or without 15% ethanol. (B) Single rejected males exhibited higher ethanol preference than single mated males (**P <0.01, two-way repeated-measures ANOVA with Bonferroni post-tests, n = 9, comparisons are between treatment groups across 3 d after the end of training). Data shown are the mean + SEM or mean - SEM. Please click here to view a larger version of this figure.
Figure 3. Opposing Mating Experiences mModulate Ethanol Preference. (A) Schematic of the combined protocol. Single virgin WT males are allowed to mate with virgin females or are subjected to 3x 1h courtship-suppression training sessions (depicted as "T") spaced by 1 h of rest (depicted as "R"). Training is repeated for 4 d. At the end of each day, the flies are placed in the incubator (depicted as "O.N"). After 4 d of training, the males were placed in vials where they could choose to feed from capillaries containing food solutions with or without 15% ethanol. (B) Single rejected males exhibited higher ethanol preference than single mated males (**P <0.01, two-way repeated-measures ANOVA with Bonferroni post-tests, n = 9, comparisons are between treatment groups across 3 d after the end of training). Data shown are the mean + SEM or mean - SEM. Please click here to view a larger version of this figure.
| Reference section | Notes |
| 1.1 | Can be used in many experiments |
| 1.2 | Can be kept in 4 °C for several weeks |
| 1.3 | Keep in 4 °C after autoclave |
| 1.4 | Aged to 3 - 4 d before being used in the experiment |
| 1.5 | This step should take place the evening before each training session |
| 2.1 | This step should take place the morning of each training session |
| 2.2 | To be repeated in 3 consecutive training sessions over the course of 4 d |
| 2.3 | Dump the plugs with water, measure consumption and replace capillaries over the course of 4 d |
| 2.4 | This step is done each time before replacing the capillaries during the two-choice assay |
Table 1. An overview of the protocol depicting the order and time flow for the different steps.
Discussion
Here, we illustrate the details of an integrated approach to measure alterations in reward-seeking behavior, based on previous work described by Devineni et al.6 and Shohat-Ophir et al5. The first section of the protocol uses different types of sexual interactions as the experience input, and the second section uses a two-choice feeding assay to assess the effect of experience on the preference to consume ethanol.
As shown by Devineni et al.6, ethanol self-administration in flies is a robust behavior, characterized at first by low and variable preference that increases and becomes more consistent over time. The developing preference may reflect a process through which flies learn to associate the pharmacological effect of ethanol with its position, in spite of its unpleasant taste6 and the aversive responses known to occur during initial exposure to ethanol vapors11. Changing the position of the ethanol-containing capillary on a daily basis results in a constant level of preference, strongly demonstrating the role of learning and suggesting that the flies leave chemical cues next to the preferred capillaries12. In addition, the shape, the slope, and the starting point of the preference curve can be slightly different between experiments, but the overall magnitude of difference between the groups of flies that are exposed to rewarding and non-rewarding experiences is preserved across experiments.
The question of whether flies display preference to consume ethanol for its caloric value or pharmacological properties is still under debate and was examined in the last few years by several independent studies (for a review, see Devineni et al.)13. Taking into account the results obtained from these studies, it is likely that the caloric value of ethanol contributes to the pharmacological effect of ethanol and is not the main reason for the development of preference to consume ethanol.
As this protocol reflects voluntary consumption in groups of flies, the technique is limited to cumulative changes in consumption over at least several hours and is not sensitive enough to reflect short temporal changes in consumption over short time frames (minutes). Other feeding assays, such as FLIC14, flyPAD15, and EXPRESSO16, that can measure the frequency of feeding events and their bout length are technologically superior to the CAFE assay and can therefore provide high-content temporal data of feeding and ethanol preference. Still, compared to other simple two-choice assays that are based on measuring the ingestion of food mixed together with food dyes, the CAFE approach allows the accurate and uninterrupted monitoring of consumption data over several days17,18.
Although described together, each section can be used separately in future applications. The training phase can be used for further downstream analyses of plasticity-related events using real-time PCR, RNAseq, proteomic, and metabolomic screens. Following the training phase, it is also possible to use FLIC14, flyPAD15, or EXPRESSO16 to achieve higher resolution in the consumption data. The two-choice ethanol consumption assay can also serve to study the effect of other external stimuli, such as stress, on the internal state19,20. Additionally, using neurogenetic tools that control neuronal activity as an input experience and ethanol consumption as an output behavior, it can be used to dissect the neuronal circuits that encode internal reward.
Even though the described protocol is relatively simple and does not require sophisticated equipment, there are critical steps that require the most attention to ensure reproducible results. First, it is important to start the experiment with large male flies that eclosed from healthy fly bottles. Secondly, it is essential to make sure that the mated cohort indeed undergo mating and that the rejected cohort do not manage to mate. Lastly, it is important to maintain a well-humidified environment for the entire experiment, with an emphasis on the consumption part. This includes watering the plugs every day with equal amounts of water and keeping the capillary ends in close proximity to the wet plugs.
Disclosures
The authors have nothing to disclose.
Acknowledgments
We thank U. Heberlein and A. Devineni for long-lasting discussions and technical advice. We also thank the Shohat-Ophir lab members, A. Benzur, L. Kazaz, and O. Shalom, for the help with demonstrating the method. Special appreciation goes to Eliezer Costi for establishing the fly systems in the lab. This work was supported by the Israel Science Foundation (384/14) and the Marie Curie Career Integration Grants (CIG 631127).
References
- Robinson GE, Fernald RD, Clayton DF. Genes and social behavior. Science. 2008;322:896–900. doi: 10.1126/science.1159277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koob GF. Neurobiological substrates for the dark side of compulsivity in addiction. Neuropharmacol. 2009;56:18–31. doi: 10.1016/j.neuropharm.2008.07.043. Suppl 1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaun KR, Devineni AV, Heberlein U. Drosophila melanogaster as a model to study drug addiction. Hum Genet. 2012;131:959–975. doi: 10.1007/s00439-012-1146-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Devineni AV, Heberlein U. Addiction-like behavior in Drosophila. Commun Integr Biol. 2010;3:357–359. doi: 10.4161/cib.3.4.11885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shohat-Ophir G, Kaun KR, Azanchi R, Mohammed H, Heberlein U. Sexual deprivation increases ethanol intake in Drosophila. Science. 2012;335:1351–1355. doi: 10.1126/science.1215932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Devineni AV, Heberlein U. Preferential ethanol consumption in Drosophila models features of addiction. Curr Biol : CB. 2009;19:2126–2132. doi: 10.1016/j.cub.2009.10.070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McBride SM, et al. Mushroom body ablation impairs short-term memory and long-term memory of courtship conditioning in Drosophila melanogaster. Neuron. 1999;24:967–977. doi: 10.1016/s0896-6273(00)81043-0. [DOI] [PubMed] [Google Scholar]
- Ejima A, Smith BP, Lucas C, Levine JD, Griffith LC. Sequential learning of pheromonal cues modulates memory consolidation in trainer-specific associative courtship conditioning. Curr Biol. 2005;15:194–206. doi: 10.1016/j.cub.2005.01.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ejima A, et al. Generalization of courtship learning in Drosophila is mediated by cis-vaccenyl acetate. Curr Biol. 2007;17:599–605. doi: 10.1016/j.cub.2007.01.053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keleman K, Kruttner S, Alenius M, Dickson BJ. Function of the Drosophila CPEB protein Orb2 in long-term courtship memory. Nat Neurosci. 2007;10:1587–1593. doi: 10.1038/nn1996. [DOI] [PubMed] [Google Scholar]
- Kaun KR, Azanchi R, Maung Z, Hirsh J, Heberlein U. A Drosophila model for alcohol reward. Nat Neurosci. 2011;14:612–619. doi: 10.1038/nn.2805. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu S, et al. The propensity for consuming ethanol in Drosophila requires rutabaga adenylyl cyclase expression within mushroom body neurons. Genes Brain Behav. 2012;11:727–739. doi: 10.1111/j.1601-183X.2012.00810.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Devineni AV, Heberlein U. The evolution of Drosophila melanogaster as a model for alcohol research. Annu Rev Neurosci. 2013;36:121–138. doi: 10.1146/annurev-neuro-062012-170256. [DOI] [PubMed] [Google Scholar]
- Ro J, Harvanek ZM, Pletcher SD. FLIC: high-throughput, continuous analysis of feeding behaviors in Drosophila. PloS one. 2014;9:101107. doi: 10.1371/journal.pone.0101107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Itskov PM, et al. Automated monitoring and quantitative analysis of feeding behaviour in Drosophila. Nat Commun. 2014;5:4560. doi: 10.1038/ncomms5560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yapici N, Cohn R, Schusterreiter C, Ruta V, Vosshall LB. A Taste Circuit that Regulates Ingestion by Integrating Food and Hunger Signals. Cell. 2016;165:715–729. doi: 10.1016/j.cell.2016.02.061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peru YC, et al. Long-lasting, experience-dependent alcohol preference in Drosophila. Addict Biol. 2014;19:392–401. doi: 10.1111/adb.12105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dus M, et al. Nutrient Sensor in the Brain Directs the Action of the Brain-Gut Axis in Drosophila. Neuron. 2015;87:139–151. doi: 10.1016/j.neuron.2015.05.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson DJ, Adolphs R. A framework for studying emotions across species. Cell. 2014;157:187–200. doi: 10.1016/j.cell.2014.03.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gibson WT, et al. Behavioral responses to a repetitive visual threat stimulus express a persistent state of defensive arousal in Drosophila. Curr Biol. 2015;25:1401–1415. doi: 10.1016/j.cub.2015.03.058. [DOI] [PMC free article] [PubMed] [Google Scholar]