

Abstract
Although previous studies have shown that stem cells can be differentiated into Leydig cells by gene transfection, a simple, safe and effective induction method has not yet been reported. Therefore, the present study investigated novel methods for the induction of human umbilical cord mesenchymal stem cell (HUMSC) differentiation into Leydig-like, steroidogenic cells. HUMSCs were acquired using the tissue block culture attachment method, and the expression of MSC surface markers was evaluated by flow cytometry. Leydig cells were obtained by enzymatic digestion and identified by lineage-specific markers via immunofluorescence. Third-passage HUMSCs were cultured with differentiation-inducing medium (DIM) or Leydig cell-conditioned medium (LC-CM), and HUMSCs before induction were used as the control group. Following the induction of HUMSCs, Leydig cell lineage-specific markers (CYP11A1, CYP17A1 and 3β-HSD) were positively identified using immunofluorescence analysis. Additionally, reverse transcription-quantitative polymerase chain reaction and western blot analysis were performed to evaluate the expression levels of these genes and enzymes. In contrast, the control group cells did not show the characteristics of Leydig cells. Collectively, these results indicate that, under in vitro conditions, LC-CM can achieve a comparable effect to that of DIM on inducing HUMSCs differentiation into steroidogenic cells.
Keywords: Leydig cells, umbilical cord mesenchymal stem cells, steroidogenesis
Introduction
Environmental pollution, food additives, aging, injuries and trauma induced by cancer chemotherapy may all lead to androgen deficiency or nonexistence of testicular cases, which are increasing each year (1). The congenital abnormalities and other causative factors underlying these diseases are also common among children (2). One clinical treatment method involves the transplantation of Leydig cells (3). Leydig cells can secrete 95% of the body's testosterone in the adult testes and serves a crucial function in testosterone generation (4). Although this treatment avoids certain complications by using exogenous androgen therapy, it may lead to immunological rejection by the host body. Furthermore, the supply of testicular interstitial cells is limited, which restricts the transplantation of Leydig cells in clinical applications (5,6).
Mesenchymal stem cells (MSCs) have demonstrated marked self-renewal and pluripotency characteristics clinically and have been increasingly used in tissue engineering and regenerative medicine seed cell research in recent years (7). Bone marrow MSCs have been thoroughly characterized (8,9); however, their acquisition can be harmful, and their differentiation capability decreases with age (10). Thus, human umbilical cord MSCs (HUMSCs) have been investigated as an alternative due to the ease with which they are non-invasively obtained and their low immunogenicity and higher differentiation capacity (11).
Studies have shown that, under certain microenvironment conditions or through addition of special inducing factors, HUMSCs can be differentiated into cartilage cells, matrix cells, musculoskeletal and endothelial cells (12–15). However, the induction of HUMSC differentiation into Leydig cells by changing the culture conditions in vitro has not been reported. Apart from this, one study demonstrated that stem Leydig cells can differentiate into mature Leydig cells by the application of differentiation-inducing medium (DIM) (16). Currently, the differentiation of stem cells into Leydig cells with gene transfection technology is mature, but its reliability is controversial (17).
In the present study, HUMSC differentiation into steroidogenic cells was induced using DIM prepared by adding multiple inducible factors. However, as the DIM preparation process is complex, the efficacy of Leydig cell-derived conditioned medium (LC-CM) for HUMSC differentiation was also tested in order to simplify the experimental methods. The present data demonstrate that HUMSCs may differentiate into steroidogenic cells by adding cytokines in the absence of gene transfection conditions; moreover, LC-CM may induce HUMSC differentiation into steroidogenic cells.
Materials and methods
Ethics statement
In this study, all experiments involving human participants were approved by the institutional review board of the Chinese Academy of Medical Science and Medical School of Shanghai Jiaotong University (Shanghai, China). All subjects provided written informed consent for participation in the study and approved publication of their case details.
This study was conducted in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of Shanghai Jiaotong University and the protocol was approved by the Committee on the Ethics of Animal Experiments of Shanghai Jiaotong University. All surgical procedures on animals were performed under sodium pentobarbital anesthesia (P3767; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) and all efforts were made to minimize suffering.
Isolation and culture of HUMSCs
With parental consent, human umbilical cords were aseptically obtained from 12 full-term newborn male infants delivered by cesarean section at Shanghai Jiaotong University School of Medicine affiliated with Renji Hospital. Umbilical cords were preserved in cold low glucose Dulbecco's modified Eagle's medium (DMEM-LG; Invitrogen; Thermo Fisher Scientific, Inc., Carlsbad, CA, USA), and cellular isolation began within 3 h after delivery. HUMSC isolation was performed using the tissue block culture attachment method (18). The blood vessels were removed, and the cord was sheared into 2-3-mm3 pieces, which were transferred to petri dishes and cultured in a 37°C incubator with 5% CO2 in DMEM-LG containing 10% fetal bovine serum (FBS; Invitrogen), 1% penicillin and streptomycin (Gibco; Thermo Fisher Scientific, Inc.). The medium was changed every 2 days after planting and 10 days later, the tissue blocks were removed. When the cells reached 70–80% confluence, they were harvested and cultured at a density of 1×104 cells/cm2. Only cells from passages 3–5 were used for cell differentiation. The surface markers of HUMSCs were analyzed by flow cytometry.
Identification of MSC marker expression by flow cytometry
Third passage cells were collected and washed twice in phosphate-buffered saline (PBS; Hyclone; GE Healthcare Life Sciences, Logan, UT, USA). Cells were harvested using 0.25% trypsin (Invitrogen), washed in PBS and incubated for 30 min at 4°C in the dark with the following anti-human antibodies conjugated with fluorescein isothiocyanate (FITC), phycoerythrin (PE) or allophycocyanin (APC): Anti-CD31-PE (FAB3567P), -CD45-PE (FAB1430P), -CD34-PE (FAB72271P) and-CD105-APC (FAB10971A) from R&D Systems, Inc. (Minneapolis, MN, USA), and anti-CD44-FITC (ab27285) and -CD90-FITC (ab11155) from Abcam (Cambridge, UK). PE-conjugated IgG1 (IC002P; R&D Systems, Inc.) and FITC-conjugated IgG1 (sc-2078; Santa Cruz Biotechnology, Inc., Dallas, TX, USA) were used as isotype controls. After washing the cells with PBS, the expression of the markers on the cells was analyzed using flow cytometry, as previously described (16).
Differentiation potential of MSCs
The osteogenic, adipogenic and chondrogenic differentiation capacity of isolated HUMSCs were assessed using third passage MSCs.
For osteogenic differentiation, cells were plated at a density of 2×104 cells/cm2 and cultured in DMEM-LG supplemented with 10% FBS until reaching 80–90% confluence. The DMEM-LG was then removed and Human Umbilical Cord MSC Osteogenic Differentiation Medium (Cyagen Biosciences, Inc., Santa Clara, CA, USA) was added and cells were cultured for 28 days. To detect osteogenesis, Alizarin Red S staining (Cyagen Biosciences, Inc.) was performed after 28 days of culture. The cells were then washed with PBS and fixed with 4% formaldehyde solution for 30 min. The wells were rinsed twice with 1X PBS and cells were then stained with Alizarin Red working solution for 3–5 min.
For adipogenic differentiation, cells were plated at a density of 2×104 cells/cm2 and cultured in DMEM-LG supplemented with 10% FBS until reaching 100–120% confluence. After the DMEM-LG was removed, Human Umbilical Cord MSC Adipogenic Differentiation Medium A and Medium B (Cyagen Biosciences, Inc.) were used to culture cells successively for 28 days. Cells were processed for Oil Red O staining (Cyagen Biosciences, Inc.) to detect adipogenesis. The staining process was similar to that used for detection of osteogenesis.
For chondrogenic differentiation, 2.5×105 cells were seeded into 15 ml polypropylene culture tubes. The cells were centrifuged at 150 × g for 5 min at room temperature. The caps of the tubes were loosened by one half turn to allow gas exchange, and the tubes were incubated at 37°C, in a humidified atmosphere of 5% CO2. After 24 h, freshly prepared Complete Chondrogenic Medium (Cyagen Biosciences, Inc.) was to each tube. After 28 days culture, the chondrogenic pellets were harvested, formalin fixed and paraffin embedded for Alcian blue staining (Cyagen Biosciences, Inc.).
Isolation and culture of Leydig cells
Testes were obtained from 8-week-old mice, which were sacrificed by cervical dislocation. Leydig cells were isolated as described previously (19) digested by combining testes with 0.03% collagenase NB4 (SERVA Electrophoresis GmbH, Heidelberg, Germany) in a centrifuge tube. The supernatant was collected after digestion for 15 min in a 37°C shaker at 150 rpm. Cells in the supernatant were centrifuged and cultured in DMEM/F12 (Invitrogen) with 10% FBS. After 4 days, cells were harvested with 0.25% trypsin treatment. First passage cells were preserved at −80°C. The surface makers of Leydig cells were detected by immunofluorescence staining (20).
Preparation of Leydig cell-conditioned medium (LC-CM)
Second passage Leydig cells were plated on a Petri dish at a density of 1×104 cells/cm2. Leydig cells were cultured in DMEM/F12 containing 10% FBS, 1% penicillin and streptomycin. The medium was collected daily and replaced with fresh medium every 3 days. The medium was centrifuged at 500 × g for 5 min, and the supernatant was filtered with a 0.22 µm filter. The filtered medium and normal DMEM/F12 medium constitute the LC-CM, which was prepared by diluting the collected conditioned medium with an equal volume of DMEM/F12 containing 10% FBS.
Preparation of DIM
DIM (16) was prepared by adding several hormones to phenol red-free DMEM/F12 as follows: 5% FBS, 1 ng/ml luteinizing hormone (LH; American Research Products, Inc., Waltham, MA, USA), 1 nM thyroid hormone (Gibco), 70 ng/ml insulin-like growth factor 1 (IGF-1), 10 ng platelet-derived factor (PDGF)-BB (American Research Products, Inc.) and insulin-transferrin-selenium cell culture supplement containing 5 mg/l insulin, 5 mg/l transferrin and 5 µg/l selenium (17-838Z; Lonza Walkersville, Inc., Walkersville, MD, USA).
HUMSC differentiation into steroidogenic cells in vitro
For differentiation into steroidogenic cells, three experimental groups of HUMSCs were established and cultured initially at 2,000–4,000 cells/cm2 in DMEM-LG with 10% FBS for 24 h. After 24 h, media from the three groups were replaced with LC-CM, DIM or DMEM/F12 with 10% FBS (control group). The medium was changed daily.
Immunofluorescence staining
After 14 days of exposure to LC-CM or DIM, the HUMSCs were fixed with 4% polyoxymethylene for 20 min at room temperature and permeabilized with 0.25% Triton X-100 (Sigma-Aldrich) and blocked with 0.1% bovine serum albumin (Roche, Basel, Switzerland). Subsequently, cells were incubated overnight at 4°C with the following primary antibodies: Cholesterol side-chain cleavage enzyme CYP17A1 (1:200; ABC392; Merck Millipore, Merck KGaA), CYP11A1 (1:200; ab75497; Abcam) and 3β-hydroxysteroid dehydrogenase (3β-HSD; 1:200; ab154385; Abcam). After washing three times with PBS, the cells were reacted with the appropriate fluorescent dye-conjugated secondary antibody for 30 min at 37°C. The cell nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI; Invitrogen) and visualized by fluorescence microscopy (Leica Camera AG, Wetzlar, Germany). The untreated HUMSCs were used as negative controls. All antibodies were diluted in PBS.
Reverse transcription-polymerase chain reaction (RT-PCR) analysis
Total cellular RNA was extracted from LC-CM and DIM induced HUMSCs at days 3, 7 and 10 using TRIzol reagent (Invitrogen) according to the manufacturer's recommendations. Phase Lock Gel (Tiangen Biotech Co., Ltd., Beijing, China) was used to remove genomic DNA according to the manufacturer's protocol. The concentration of RNA was measured by using a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Inc.) and then 900 ng RNA was reversely transcribed using a reverse transcriptase kit (Takara Bio, Inc., Shiga, Japan) for 15 min at 42°C and 5 sec at 85°C. The cDNA was amplified using a PCR kit (Takara Bio, Inc.) with the following primers:: Luteinizing hormone receptor (LHR), forward 5′-GTGGCTGGGACTATGAATATGGTTT-3′ and reverse 5′-CTAGAGTGATGACGGTGAGGGTGTA-3′; Steroidogenic acute regulatory protein (StAR), forward 5′-ATGGGTGGGGTTCGTGTTTAGA-3′ and reverse 5′-TGGGGTGCTGCTTGTTCTGTG-3′; HSD3B2, forward 5′-TCCAAGCCAGTGTGCCAGTCT-3′ and reverse 5′-CTTTGGTGAGGCGTGTCATCTG-3′; CYP17A1, forward 5′-GCAAGAGATAACACAAAGTCAAGGT-3′ and reverse 5′-TGCCCATACGAACCGAATAGAT-3′; CYP11A1, forward 5′-CCAACCCAGAACGATTCCTCAT-3′ and reverse 5′-CACTACTTCCTCCAGCATCCCCT-3′; and β-actin, forward 5′-CGGGAAATCGTGCGTGACAT-3′ and reverse 5′-CGGACTCGTCATACTCCTGCTTG-3′. PCR assays were conducted by denaturation at 95°C for 30 sec followed by 30 cycles of annealing at 58°C for 30 sec and extension at 72°C for 60 sec. Finally, PCR products were separated by 2% agarose gel electrophoresis and observed under ultraviolet illumination. The expected product sizes from these primers were 454, 334, 408, 332, 387 and 481 bp for HSD3B2, CYP17A1, LHR, CYP11A1, StAR and β-actin, respectively.
Protein extraction and western blot analysis
After 3 weeks of exposure to LC-CM or DIM, cells were washed twice with cold PBS and harvested by trypsinization. For total protein isolation, cells were suspended in cell lysis buffer (Invitrogen) and placed on ice for 30 min. The suspension was collected after centrifugation at 15,000 × g for 30 min at 4°C. Protein concentrations were measured using a bicinchoninic acid protein assay kit (Bio-Rad Laboratories, Inc., Hercules, CA, USA) according to the manufacturer's instruction. Proteins (20 µg) were denatured by boiling at 100°C for 5 min after the addition of a loading buffer (CWbio, Beijing, China). Equivalent quantities of protein were loaded and electrophoresed in 6–15% SDS-PAGE gels. After electrophoresis, gels were transferred to nitrocellulose (NC; Merck Millipore) membranes, and membranes were blocked with 5% non-fat milk in Tris-buffered saline containing 0.1% Tween 20 for 1 h at room temperature. Membranes were incubated with primary antibodies targeting CYP17A1 (1:1,000; ABC392), CYP11A1 (1:1,000; ab75497), 3β-HSD (1:1,000; ab154385) or β-actin (1:1,000; ab8227; Abcam) overnight at 4°C, then with IRDye 800-conjugated goat anti-rabbit IgG secondary antibody (1:20,000; A50984P; Rockland Immunochemicals, Inc., Pottstown, PA, USA) for 2 h at room temperature. Immunoreactive bands were visualized using a LI-COR Odyssey (LI-COR Biosciences, Lincoln, NE, USA).
Results
Isolation and morphological features of HUMSCs
After three days in culture, the cells isolated from the human umbilical cord blocks were scarce and scattered, principally exhibiting a spindle-shaped morphology (Fig. 1A). After 10 days, the number of cells increased, and they grew in a parallel arrangement (Fig. 1B). Once passaged, the cells expanded rapidly with no significant changes in morphology (Fig. 1C).
Figure 1.

Morphological appearance of human umbilical cord mesenchymal stem cells (HUMSCs). (A) The appearance of primary HUMSCs cultured for (A) 3 days and (B) 10 days. (C) The appearance of HUMSCs after passaging (scale bar, 100 µm).
Identification of HUMSCs
After two passages, the cells exhibited positive expression of the MSC surface markers CD44, CD90 and CD105. However, the cells were negative for hematopoietic lineage markers (CD34, CD45 and CD31). These results indicated that the isolated cells in this study were HUMSCs (Fig. 2A). After 28 days of osteogenic induction, the deposition of calcium of cells could be visualized by Alizarin Red S staining (Fig. 2B). When cells were cultured in adipogenic medium for 28 days, numerous lipid droplets were observed following staining with Oil Red O solution, which used to identify adipocytes (Fig. 2C). Blue staining indicates synthesis of proteoglycans by chondrocytes after 28 days of induction (Fig. 2D).
Figure 2.

Identification of human umbilical cord mesenchymal stem cells. (A) The third-passage adherent cells were positive for CD44, CD90 and CD105 and negative for CD31, CD34 and CD45. (B) Alizarin Red S staining shows the deposition of calcium of cells. (C) Oil Red O stained the lipid droplets of the cells. (D) Alcian blue staining indicates synthesis of proteoglycans by chondrocytes (magnification, ×200).
Isolation and morphological features of Leydig cells
Leydig cells predominantly exhibited round shapes with large lipid drops and tended to be arranged in clusters (Fig. 3), as previously reported (21). Immunofluorescence staining of Leydig cells for CYP17A1 and 3β-HSD was positive (Fig. 4).
Figure 3.
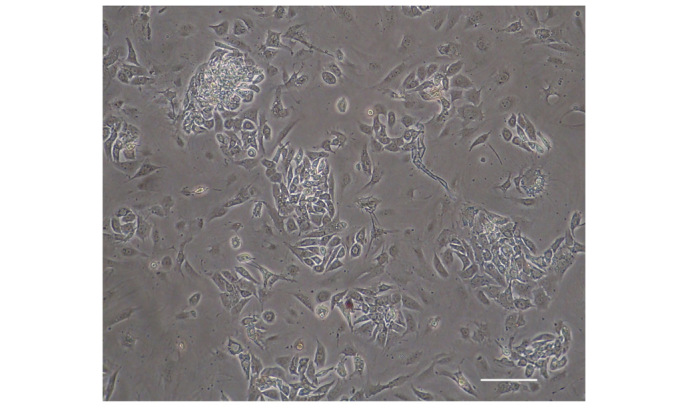
Primary culture of Leydig cells. Leydig cells predominantly exhibited round shapes with large lipid droplets present in the cytoplasm. Cells tended to group together to form clusters (scale bar, 100 µm).
Figure 4.

Enzymes for steroidogenesis expressed in Leydig cells. Positive immunofluorescence for 3β-HSD: (A) Green, 3β-HSD in cytoplasm; (B) blue, nuclei; (C) merged. Positive immunofluorescence for CYP17A1: (D) Green, CYP17A1 in cytoplasm; (E) blue, nuclei; (F) merged. (Scale bar, 100 µm.) 3β-HSD, 3β-hydroxysteroid dehydrogenase.
Expression of Leydig cell lineage markers in induced HUMSCs analyzed by immunofluorescence staining
To detect the Leydig cell phenotype in induced HUMSCs, immunofluorescence staining for CYP11A1, CYP17A1 and 3β-HSD was performed on cells from all three groups following 14 days of culture. Positive staining for CYP11A1, CYP17A1 and 3β-HSD was observed in the HUMSCs treated with either LC-CM or DIM. However, no positive staining was observed in the control cells (Fig. 5).
Figure 5.

Immunofluorescence staining of Leydig cell differentiation of human umbilical cord mesenchymal stem cells (HUMSCs) induced in DIM and LC-CM. HUMSCs induced by DIM and LC-CM were positively stained for CYP11A1, CYPY17A1 and 3β-HSD. Immunofluorescence for 3β-HSD was (A) negative in control cells and positive in (B) LC-CM and (C) DIM-induced cells. Blue, nuclei; red, 3β-HSD in cytoplasm. Immunofluorescence for CYP11A1 was (D) negative in control cells and positive in (E) LC-CM and (F) DIM-induced cells: Blue, nuclei; red CYP11A1 in cytoplasm. Immunofluorescence for CYP17A1 was (G) negative in control cells and positive in (H) LC-CM and (I) DIM-induced cells: Blue, nuclei; red, CYP17A1 in cytoplasm. (Scale bar, 100 µm.) CM, conditioned medium; DIM, differentiation-inducing medium; 3β-HSD, 3β-hydroxysteroid dehydrogenase.
Expression of Leydig cell lineage markers in induced HUMSCs analyzed by RT-PCR
To assess the possibility of Leydig cell differentiation, HUMSCs were evaluated by RT-PCR analysis of specific testosterone-producing genes. After 7 days of culture, the results demonstrated that HUMSCs induced with LC-CM or DIM positively expressed Leydig cell lineage marker genes (Fig. 6).
Figure 6.
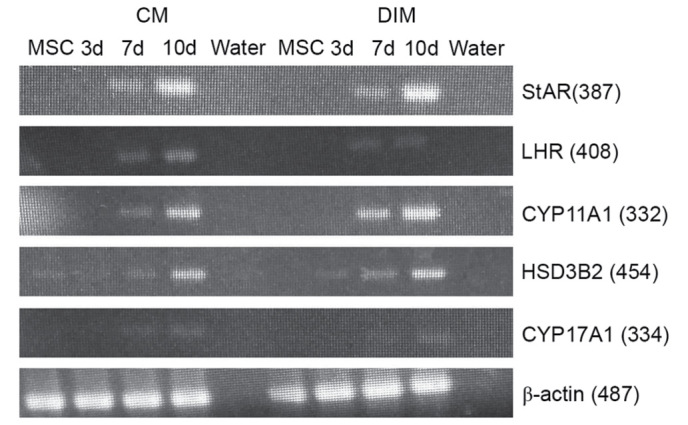
Reverse transcription-polymerase chain reaction analysis results for LC-CM and DIM-treated HUMSCs. LC-CM and DIM-treated HUMSCs were positive for StAR, LHR, 3β-HSD, CYP11A1, CYP17A1 and β-actin while control cells were negative. CM, conditioned medium; DIM, differentiation-inducing medium; StAR, steroidogenic acute regulatory protein; LHR, luteinizing hormone receptor; 3β-HSD, 3β-hydroxysteroid dehydrogenase; HUMSC, human umbilical cord mesenchymal stem cell; d, day.
Expression of Leydig cell lineage markers in induced HUMSCs analyzed by western blot analysis
Next, whether whether Leydig cell lineage enzymes can be expressed after induction was examined. 3β-HSD, CYP11A1 and CYP17A1 were induced 3 weeks after LC-CM or DIM treatment. In the testes, the production of testosterone is dependent on 3β-HSD activity (22). Additionally, 3β-HSD enzymes are essential for the production of progesterone, and CYP17A1 is required for the production of testosterone (23). These results demonstrate that HUMSCs have properties indicating their differentiation into steroidogenic cells, which are similar to those of Leydig cells (Fig. 7).
Figure 7.

Western blot analysis was performed with LC-CM and DIM-treated human umbilical cord mesenchymal stem cells (HUMSCs). LC-CM and DIM-treated HUMSCs were positive for 3β-HSD, CYP11A1 and CYP17A1. CM, conditioned medium; DIM, differentiation-inducing medium; 3β-HSD, 3β-hydroxysteroid dehydrogenase.
Discussion
MSCs have a great capacity for self-renewal while maintaining multipotency. Previous experiments have shown that HUMSCs have the advantages of easy accessibility, high multipotency and low immunogenicity, and they are gradually replacing bone marrow MSCs as an ideal clinical source of MSCs for cell therapy and tissue engineering (24). The tissue block attachment method and mechanical removal of the epithelia and vessels may avoid contamination while maintaining cellular viability and function (25). The present study was conducted under strict quality control to ensure proper expression of multiple markers of MSCs via flow cytometry. Positive expression of the MSC markers CD44, CD90 and CD105 was detected, and negative expression of the hematopoietic stem cell markers CD31, CD34 and CD45. As a result, the adherent cells, which were isolated and cultured, were considered to be HUMSCs.
During development, Leydig cells arise sequentially, first as stem Leydig cells (SLC) then as progenitor Leydig cells (PLC), immature Leydig cells (ILC) and mature Leydig cells (MLCs). Various cytokines, such as insulin-like factor 3 (INSL3), stem cell factor (SCF), platelet-derived factor (PDGFAA) and Müllerian inhibiting substance can regulate Leydig cell development (26–29). Cellular synthesis of testosterone also requires the application of numerous enzymes. StAR plays an important role in the synthesis of testosterone, which is a rate-limiting enzyme for steroid hormone synthesis (30). LHRs first appear as PLCs differentiate, and the development of the steroidogenic capacity of PLCs needs stimulation by LH (31). Leydig cell lineage specific markers, such as CYP11A1, CYP17A1 and 3β-HSD, are essential for steroid synthesis in adrenal gland and gonads. In this study, the positive expression of Leydig cell lineage markers indicate that HUMSCs could differentiate into Leydig cells (32).
MLCs can be considered a terminally differentiated state; however, previous studies have reported that, in an appropriate microenvironment, certain cytokines can regulate Leydig cell proliferation (33). Ge et al cultured SLCs in DIM and successfully induced their differentiation into ALCs (16,34). Microarray analysis was conducted using purified SLCs, PLCs, ILCs, MLCs and bone marrow stem cells (BSCs); highly correlated gene expression patterns were seen between BSCs and SLCs, and greater differences were seen in gene expression between SLCs and PLCs and between SLCs and MLCs, indicating that the transcriptome of the SLCs is similar to that of stem cells (35). In view of these results, the same inducible factors were used in this study to induce HUMSC differentiation into Leydig cells.
DIM contains several cytokines important for Leydig cell development. LH serves a crucial function in inducing differentiation of ILCs into MLCs. LH is also crucial for MLC proliferation, as binding of LH to LHR increases testosterone synthesis by Leydig cells (36). Thyroid hormone T3 is critical to initiate the onset of mesenchymal cell differentiation into adult Leydig cells, while IGF-I and PDGF-BB can promote cell proliferation and differentiation (31,37,38).
In the present study, under the effect of DIM, HUMSCs demonstrate the ability to synthesize steroid hormones. However, this approach has certain disadvantages, as the cytokine concentration and media preparation method is complicated and the cost of purchasing inducible factors is high. Therefore, LC-CM use as a new induction method was investigated.
Previous studies have shown that cells and their secreted proteins can form a functional niche in vivo, and the balance of self-renewal and differentiation of all stem cells is affected under the influence of this niche. Similarly, all functional factors contained in the conditioned medium, including proteins or ions, may also determine the direction of stem cell differentiation (39–42). The present experimental data show that provision of a suitable microenvironment from LC-CM may directly promote HUMSC differentiation into Leydig cells.
Previous research has shown that two methods can induce MSCs differentiation towards Leydig cells: Infection of MSCs with an adenovirus-containing steroidogenesis-related gene in vitro or interstitial testicular transplantation of MSCs in vivo (43–46). However, both methods are limited by the difficultly of obtaining a large number of safe and effective seed cells for tissue reconstruction. The present experiments for the first time induced HUMSCs differentiation into steroidogenic cells, which may have the ability to synthesize testosterone in vitro. Nevertheless, further studies are required to investigate the underlying mechanism of DIM and LC-CM-mediated induction of HUMSC differentiation into Leydig cells.
In summary, the present results suggest that HUMSCs are able to differentiate into steroidogenic cells in an appropriate microenvironment in vitro. DIM and LC-CM may constitute a functional niche to promote the differentiation of HUMSCs.
Acknowledgements
This study was supported by National Natural Science Foundation of China (grant no. 81270689) and Research Program of Science and Technology Commission of Shanghai Municipality (grant no. 12ZR1419200).
References
- 1.Wohlfahrt-Veje C, Main KM, Skakkebaek NE. Testicular dysgenesis syndrome: Foetal origin of adult reproductive problems. Clin Endocrinol (Oxf) 2009;71:459–465. doi: 10.1111/j.1365-2265.2009.03545.x. [DOI] [PubMed] [Google Scholar]
- 2.Meikle AW. The interrelationships between thyroid dysfunction and hypogonadism in men and boys. Thyroid. 2004;14:S17–S25. doi: 10.1089/105072504323024552. (Suppl 1) [DOI] [PubMed] [Google Scholar]
- 3.Sun J, Xi YB, Zhang ZD, Shen P, Li HY, Yin MZ, Li WY, Shi CR. Leydig cell transplantation restores androgen production in surgically castrated prepubertal rats. Asian J Androl. 2009;11:405–409. doi: 10.1038/aja.2009.22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Zirkin BR. Where do adult Leydig cells come from? Biol Reprod. 2010;82:1019–1020. doi: 10.1095/biolreprod.110.084111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Jahnukainen K, Ehmcke J, Hou M, Schlatt S. Testicular function and fertility preservation in male cancer patients. Best Pract Res Clin Endocrinol Metab. 2011;25:287–302. doi: 10.1016/j.beem.2010.09.007. [DOI] [PubMed] [Google Scholar]
- 6.Lo KC, Lei Z, Rao ChV, Beck J, Lamb DJ. De novo testosterone production in luteinizing hormone receptor knockout mice after transplantation of leydig stem cells. Endocrinology. 2004;145:4011–4015. doi: 10.1210/en.2003-1729. [DOI] [PubMed] [Google Scholar]
- 7.Mobasheri A, Csaki C, Clutterbuck AL, Rahmanzadeh M, Shakibaei M. Mesenchymal stem cells in connective tissue engineering and regenerative medicine: Applications in cartilage repair and osteoarthritis therapy. Histol Histopathol. 2009;24:347–366. doi: 10.14670/HH-24.347. [DOI] [PubMed] [Google Scholar]
- 8.Charbord P. Bone marrow mesenchymal stem cells: Historical overview and concepts. Hum Gene Ther. 2010;21:1045–1056. doi: 10.1089/hum.2010.115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Lee OK, Kuo TK, Chen WM, Lee KD, Hsieh SL, Chen TH. Isolation of multipotent mesenchymal stem cells from umbilical cord blood. Blood. 2004;103:1669–1675. doi: 10.1182/blood-2003-05-1670. [DOI] [PubMed] [Google Scholar]
- 10.Stenderup K, Justesen J, Clausen C, Kassem M. Aging is associated with decreased maximal life span and accelerated senescence of bone marrow stromal cells. Bone. 2003;33:919–926. doi: 10.1016/j.bone.2003.07.005. [DOI] [PubMed] [Google Scholar]
- 11.Wang HS, Hung SC, Peng ST, Huang CC, Wei HM, Guo YJ, Fu YS, Lai MC, Chen CC. Mesenchymal stem cells in the Wharton's jelly of the human umbilical cord. Stem Cells. 2004;22:1330–1337. doi: 10.1634/stemcells.2004-0013. [DOI] [PubMed] [Google Scholar]
- 12.Hsieh JY, Fu YS, Chang SJ, Tsuang YH, Wang HW. Functional module analysis reveals differential osteogenic and stemness potentials in human mesenchymal stem cells from bone marrow and Wharton's jelly of umbilical cord. Stem Cells Dev. 2010;19:1895–1910. doi: 10.1089/scd.2009.0485. [DOI] [PubMed] [Google Scholar]
- 13.Mitchell KE, Weiss ML, Mitchell BM, Martin P, Davis D, Morales L, Helwig B, Beerenstrauch M, Abou-Easa K, Hildreth T, et al. Matrix cells from Wharton's jelly form neurons and glia. Stem Cells. 2003;21:50–60. doi: 10.1634/stemcells.21-1-50. [DOI] [PubMed] [Google Scholar]
- 14.Wang L, Ott L, Seshareddy K, Weiss ML, Detamore MS. Musculoskeletal tissue engineering with human umbilical cord mesenchymal stromal cells. Regen Med. 2011;6:95–109. doi: 10.2217/rme.10.98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Chen MY, Lie PC, Li ZL, Wei X. Endothelial differentiation of Wharton's jelly-derived mesenchymal stem cells in comparison with bone marrow-derived mesenchymal stem cells. Exp Hematol. 2009;37:629–640. doi: 10.1016/j.exphem.2009.02.003. [DOI] [PubMed] [Google Scholar]
- 16.Ge RS, Dong Q, Sottas CM, Papadopoulos V, Zirkin BR, Hardy MP. In search of rat stem Leydig cells: Identification, isolation, and lineage-specific development. Proc Natl Acad Sci USA. 2006;103:2719–2724. doi: 10.1073/pnas.0507692103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Wegman F, Oner FC, Dhert WJ, Alblas J. Non-viral gene therapy for bone tissue engineering. Biotechnol Genet Eng Rev. 2013;29:206–220. doi: 10.1080/02648725.2013.801227. [DOI] [PubMed] [Google Scholar]
- 18.Weiss ML, Medicetty S, Bledsoe AR, Rachakatla RS, Choi M, Merchav S, Luo Y, Rao MS, Velagaleti G, Troyer D. Human umbilical cord matrix stem cells: Preliminary characterization and effect of transplantation in a rodent model of Parkinson's disease. Stem cells. 2006;24:781–792. doi: 10.1634/stemcells.2005-0330. [DOI] [PubMed] [Google Scholar]
- 19.Sun J, Zhong L, Zhu Y, Liu G. Research on the isolation of mouse Leydig cells using differential digestion with a low concentration of collagenase. J Reprod Dev. 2011;57:433–436. doi: 10.1262/jrd.10-123N. [DOI] [PubMed] [Google Scholar]
- 20.Du T, Cheng J, Zhong L, Zhao XF, Zhu J, Zhu YJ, Liu GH. The alleviation of acute and chronic kidney injury by human Wharton's jelly-derived mesenchymal stromal cells triggered by ischemia-reperfusion injury via an endocrine mechanism. Cytotherapy. 2012;14:1215–1227. doi: 10.3109/14653249.2012.711471. [DOI] [PubMed] [Google Scholar]
- 21.Teerds KJ, Huhtaniemi IT. Morphological and functional maturation of Leydig cells: From rodent models to primates. Hum Reprod Update. 2015;21:310–328. doi: 10.1093/humupd/dmv008. [DOI] [PubMed] [Google Scholar]
- 22.Klinefelter GR, Hall PF, Ewing LL. Effect of luteinizing hormone deprivation in situ on steroidogenesis of rat Leydig cells purified by a multistep procedure. Biol Reprod. 1987;36:769–783. doi: 10.1095/biolreprod36.3.769. [DOI] [PubMed] [Google Scholar]
- 23.Shima Y, Miyabayashi K, Haraguchi S, Arakawa T, Otake H, Baba T, Matsuzaki S, Shishido Y, Akiyama H, Tachibana T, et al. Contribution of Leydig and Sertoli cells to testosterone production in mouse fetal testes. Mol Endocrinology. 2013;27:63–73. doi: 10.1210/me.2012-1256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Li X, Bai J, Ji X, Li R, Xuan Y, Wang Y. Comprehensive characterization of four different populations of human mesenchymal stem cells as regards their immune properties, proliferation and differentiation. Int J Mol Med. 2014;34:695–704. doi: 10.3892/ijmm.2014.1821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.De Bruyn C, Najar M, Raicevic G, Meuleman N, Pieters K, Stamatopoulos B, Delforge A, Bron D, Lagneaux L. A rapid, simple, and reproducible method for the isolation of mesenchymal stromal cells from Wharton's jelly without enzymatic treatment. Stem Cells Dev. 2011;20:547–557. doi: 10.1089/scd.2010.0260. [DOI] [PubMed] [Google Scholar]
- 26.Chen H, Ge RS, Zirkin BR. Leydig cells: From stem cells to aging. Mol Cell Endocrinol. 2009;306:9–16. doi: 10.1016/j.mce.2009.01.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Tsuchida J, Dohmae K, Kitamura Y, Nishimune Y. The role of the c-kit receptor in the regenerative differentiation of rat Leydig cells. Int J Androl. 2003;26:121–125. doi: 10.1046/j.1365-2605.2003.00412.x. [DOI] [PubMed] [Google Scholar]
- 28.Ferlin A, Foresta C. Insulin-like factor 3: A novel circulating hormone of testicular origin in humans. Ann N Y Acad Sci. 2005;1041:497–505. doi: 10.1196/annals.1282.074. [DOI] [PubMed] [Google Scholar]
- 29.Qu JH, Fei J, Hong X, Chen JF, Gu AH, Sun H, Xu XL, Song L, Wang SL, Wang XR. Involvement of IGF-I signaling pathway in the regulation of steroidogenesis in mouse Leydig cells treated with fenvalerate. Toxicology. 2012;292:151–155. doi: 10.1016/j.tox.2011.12.007. [DOI] [PubMed] [Google Scholar]
- 30.Luo L, Chen H, Zirkin BR. Leydig cell aging: Steroidogenic acute regulatory protein (StAR) and cholesterol side-chain cleavage enzyme. J Androl. 2001;22:149–156. [PubMed] [Google Scholar]
- 31.Ariyaratne HB, Mills N, Mason JI, Mendis-Handagama SM. Effects of thyroid hormone on Leydig cell regeneration in the adult rat following ethane dimethane sulphonate treatment. Biol Reprod. 2000;63:1115–1123. doi: 10.1095/biolreprod63.4.1115. [DOI] [PubMed] [Google Scholar]
- 32.Luu-The V. Assessment of steroidogenesis and steroidogenic enzyme functions. J Steroid Biochem Mol Biol. 2013;137:176–182. doi: 10.1016/j.jsbmb.2013.05.017. [DOI] [PubMed] [Google Scholar]
- 33.Wu X, Zhang N, Lee MM. Mullerian inhibiting substance recruits ALK3 to regulate Leydig cell differentiation. Endocrinology. 2012;153:4929–4937. doi: 10.1210/en.2012-1168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Svechnikov K, Landreh L, Weisser J, Izzo G, Colón E, Svechnikova I, Söder O. Origin, development and regulation of human Leydig cells. Horm Res Paediatr. 2010;73:93–101. doi: 10.1159/000277141. [DOI] [PubMed] [Google Scholar]
- 35.Stanley EL, Johnston DS, Fan J, Papadopoulos V, Chen H, Ge RS, Zirkin BR, Jelinsky SA. Stem Leydig cell differentiation: Gene expression during development of the adult rat population of Leydig cells. Biol Reprod. 2011;85:1161–1166. doi: 10.1095/biolreprod.111.091850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Stocco DM, Wang X, Jo Y, Manna PR. Multiple signaling pathways regulating steroidogenesis and steroidogenic acute regulatory protein expression: More complicated than we thought. Mol Endocrinol. 2005;19:2647–2659. doi: 10.1210/me.2004-0532. [DOI] [PubMed] [Google Scholar]
- 37.Teerds KJ, Rijntjes E, Veldhuizen-Tsoerkan MB, Rommerts FF, de Boer-Brouwer M. The development of rat Leydig cell progenitors in vitro: How essential is luteinising hormone? J Endocrinol. 2007;194:579–593. doi: 10.1677/JOE-06-0084. [DOI] [PubMed] [Google Scholar]
- 38.Bjelic MM, Stojkov NJ, Baburski AZ, Sokanovic SJ, Mihajlovic AI, Janjic MM, Kostic TS, Andric SA. Molecular adaptations of testosterone-producing Leydig cells during systemic in vivo blockade of the androgen receptor. Mol Cell Endocrinol. 2014;396:10–25. doi: 10.1016/j.mce.2014.08.007. [DOI] [PubMed] [Google Scholar]
- 39.Wu S, Cheng Z, Liu G, Zhao X, Zhong L, Zhu Y, Zhu J. Urothelial differentiation of human umbilical cord-derived mesenchymal stromal cells in vitro. Anal Cell Pathol (Amst) 2013;36:63–69. doi: 10.1155/2013/274640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Li TX, Yuan J, Chen Y, Pan LJ, Song C, Bi LJ, Jiao XH. Differentiation of mesenchymal stem cells from human umbilical cord tissue into odontoblast-like cells using the conditioned medium of tooth germ cells in vitro. Biomed Res Int. 2013;2013:218543. doi: 10.1155/2013/218543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Fuchs E, Tumbar T, Guasch G. Socializing with the neighbors: Stem cells and their niche. Cell. 2004;116:769–778. doi: 10.1016/S0092-8674(04)00255-7. [DOI] [PubMed] [Google Scholar]
- 42.Moore KA, Lemischka IR. Stem cells and their niches. Science. 2006;311:1880–1885. doi: 10.1126/science.1110542. [DOI] [PubMed] [Google Scholar]
- 43.Wei X, Peng G, Zheng S, Wu X. Differentiation of umbilical cord mesenchymal stem cells into steroidogenic cells in comparison to bone marrow mesenchymal stem cells. Cell Prolif. 2012;45:101–110. doi: 10.1111/j.1365-2184.2012.00809.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Sonoyama T, Sone M, Honda K, Taura D, Kojima K, Inuzuka M, Kanamoto N, Tamura N, Nakao K. Differentiation of human embryonic stem cells and human induced pluripotent stem cells into steroid-producing cells. Endocrinology. 2012;153:4336–4345. doi: 10.1210/en.2012-1060. [DOI] [PubMed] [Google Scholar]
- 45.Gondo S, Okabe T, Tanaka T, Morinaga H, Nomura M, Takayanagi R, Nawata H, Yanase T. Adipose tissue-derived and bone marrow-derived mesenchymal cells develop into different lineage of steroidogenic cells by forced expression of steroidogenic factor 1. Endocrinology. 2008;149:4717–4725. doi: 10.1210/en.2007-1808. [DOI] [PubMed] [Google Scholar]
- 46.Lue Y, Erkkila K, Liu PY, Ma K, Wang C, Hikim AS, Swerdloff RS. Fate of bone marrow stem cells transplanted into the testis: Potential implication for men with testicular failure. Am J Pathol. 2007;170:899–908. doi: 10.2353/ajpath.2007.060543. [DOI] [PMC free article] [PubMed] [Google Scholar]