

Abstract
The coenzyme A (CoA) biosynthetic enzymes have been used to produce various CoA analogues, including mechanistic probes of CoA-dependent enzymes such as those involved in fatty acid biosynthesis. These enzymes are also important for the activation of the pantothenamide class of antibacterial agents, and of a recently reported family of antibiotic resistance inhibitors. Herein we report a study on the selectivity of pantothenate kinase, the first and rate limiting step of CoA biosynthesis. A robust synthetic route was developed to allow rapid access to a small library of pantothenate analogs diversified at the β-alanine moiety, the carboxylate or the geminal dimethyl group. All derivatives were tested as substrates of Escherichia coli pantothenate kinase (EcPanK). Four derivatives, all N-aromatic pantothenamides, proved to be equivalent to the benchmark N-pentylpantothenamide (N5-pan) as substrates of EcPanK, while two others, also with N-aromatic groups, were some of the best substrates reported for this enzyme. This collection of data provides insight for the future design of PanK substrates in the production of useful CoA analogues.
Keywords: enzyme ligand, kinase, coenzyme A, structure-activity relationship, inhibitor, substrate selectivity
Graphical abstract
The promiscuity of E. coli PanK has limits! We report the synthesis of pantothenate analogs diversified at the β-alanine moiety, the carboxylate or the geminal dimethyl group. Interestingly, some of these are among the best substrates reported for E. coli PanK.
1. Introduction
The biosynthesis of coenzyme A (CoA),a an essential cofactor, is a process that is ubiquitously present in all living organisms. It is most commonly a five-step process from pantothenate (vitamin B5). Pantothenate kinase (PanK, EC 2.7.1.33) is the first and rate-limiting step of this process and is characterized by the ATP-dependent transformation of pantothenate to phosphopantothenate,1 as shown in Fig. 1. Three main types of PanK have been identified, namely Types I, II and III, based on differences in structural fold, regulation, substrate affinity, and specificity.2 Type I PanK enzymes, typically associated with bacteria, have gained significant interest due to the low sequence similarity to their human counterparts.3 Their structural and amino acid sequence similarity is considered high (52–91% similarity), with many predicted active site residues conserved among all isoforms (Fig. S1). Numerous studies have focused on developing PanK inhibitors and substrates.4 Of these, pantothenamides are particularly interesting due to their antibacterial activity against S. aureus and E. coli.5–10 Although the bacteriostatic effect of pantothenamides has been recognized since the 1950s, their exact mechanism of action in bacteria has only been clarified in the last 15 years.5–15 Interestingly, pantothenamides are substrates of E. coli PanK (EcPanK). Their lethal effect on bacteria stems, at least in part, from their biotransformation by the CoA biosynthetic pathway into the corresponding CoA analogues. The resulting antimetabolite proceeds to inhibit downstream pathways, such as fatty acid biosynthesis.8,16 Other studies have suggested that pantothenamides may, in some strains, act as inhibitors of the CoA biosynthetic pathway.5,16
Figure 1.
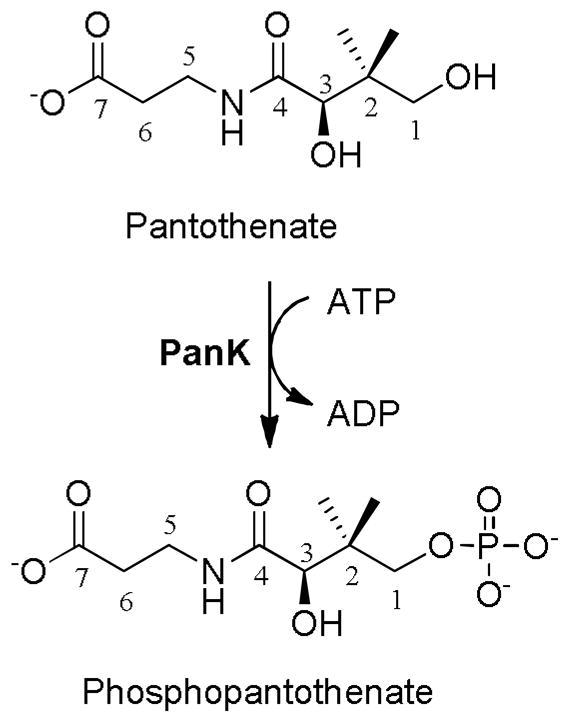
Reaction scheme for the PanK-catalyzed phosphorylation of pantothenate to phosphopantothenate.
The importance of CoA analogues not only stems from their antibacterial activity, but also from their demonstrated use as chemical biology tools and as mechanistic probes of CoA-dependent enzymes. These applications include both in vitro and in-cell labelling of carrier proteins in polyketide and nonribosomal peptide synthesis,17–19 as well as aminoglycoside resistance inhibitors activated by the CoA biosynthetic pathway.20
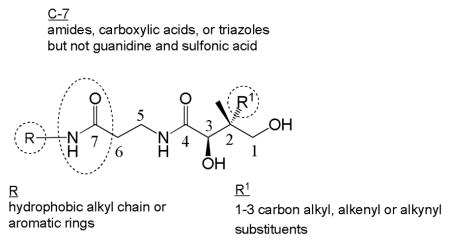
Although a wide range of pantothenamide derivatives have been reported over the years, most of these derivatives are modified at the amine moiety and less is known about the effects of modifications at other positions. Given the current and potential utility of pantothenate derivatives, a greater understanding of their effect on PanK is beneficial to future chemical biology and medicinal chemistry research. We report here on the synthesis of a library of pantothenate analogues modified at the β-alanine, the carboxylate or the geminal dimethyl group (Fig. 2), and on their structure-activity relationships as EcPanK substrates.
Figure 2.

Pantothenate analogues described in this work.
2. Results
2.1 Design and synthesis
Compounds 4-12
Compounds 4-12 are designed to probe the hydrogen bonds bridging residues Asn282 and Tyr240 to the C-7 carbonyl group of pantothenate in EcPanK. Their importance was suggested by studies with CoA analogues used for carrier protein labelling studies, where C-7 carbonyl group substitutions often showed detrimental effects.18 Synthesis of 4 and 5 (Scheme 1A) began with first obtaining the amine required for D-pantolactone ring opening. Coupling of 1-pentylamine (13) with Boc-glycine yielded 14, which was coupled with Boc-β-alanine to form the Boc protected amine 15. For the preparation of 5, the reaction of 13 with chloroacetyl chloride first produced 16, to which was added tert-butyl-3-aminopropylcarbamate to yield the Boc-protected amine 17. With 15 and 17 in hand, subsequent Boc-deprotection followed by D-pantolactone addition afforded the desired compounds 4 and 5. In the synthesis of guanidine-containing 6 (Scheme 1B), N,N′-di-Boc-thiourea (18) was first reacted with ethylenediamine to produce 19, followed by coupling to 20 as previously reported21 to yield 21. Final Boc and PMP deprotection afforded 6. For the p-chloroaniline N-linked guanidine-containing 7 (Scheme 1C), imidazole was first reacted with cyanogen bromide to form 22.22 Reaction of 22 with p-chloroaniline generated the guanidine derivative 23, which upon addition of Boc-protected propane-1,3-diamine yielded intermediate 24. Boc-deprotection of 24 and D-pantolactone addition finally afforded 7. The assembly of triazole-containing derivatives 8 and 9 (Scheme 1D) involved 1,3-dipolar cycloadditions with in situ azide formation to form intermediates 26 and 28, respectively. Deprotection and D-pantolactone addition then afforded 8 and 9. Carboxylate-containing pantothenamide 10 (Scheme 1E) was assembled by first reductive amination of 3-N-Boc protected 2,3-diaminopropanoic acid 29 with pentanal to afford 30, which was needed for nucleophilic ring opening of D-pantolactone following Boc deprotection. Synthesis of the known sulfonate-containing 11 used commercially available taurine (after NaOH treatment and precipitation) in the nucleophilic ring opening of D-pantolactone to yield 11, as previously reported.23 Finally, compound 12 (Scheme 1F) was prepared from two subsequent amide couplings of Boc-protected ethylenediamine (31) with butyric acid and with Boc-β-alanine to afford 33. Deprotection of the Boc group followed by D-pantolactone addition yielded 12.
Scheme 1.

(a) Boc-glycine, EDC, HOBt, DIPEA, THF, rt, 16 h, 94%; (b) TFA, DCM, rt, 1 h; (c) Boc-β-alanine, EDC, HOBt, DIPEA, THF, rt, 16 h, 60% over 2 steps; (d) D-pantolactone, EtOH, TEA, 80°C, 16 h, 40% over 2 steps; (e) chloroacetyl chloride, THF, rt, 16 h, 89%; (f) tert-butyl 3-aminopropylcarbamate, K2CO3, DMF, 50°C, 16 h, 60%; (g) ethylenediamine, DMF, rt, 30 min, 82%; (h) EDC, DIPEA, HOBt, THF, rt, 16 h, 79%; (i) TFA, DCM, rt, 3 h; (j) 80% AcOH, rt, 16 h, 66% over 2 steps; (k) p-chloroaniline, THF, rt, 12 h, 88%; (l) Boc-propane-1,3-diamine, THF, 65°C, 24 h, 86%; (m) TFA, DCM, rt, 1 h; (n) D-pantolactone, EtOH, TEA, 75°C, 24 h, 59% over 2 steps; (o) 1-bromopentane, NaN3, CuSO4·5H2O, sodium ascorbate, DMF, H2O, 100°C, 16 h, 81%; (p) 1-pentyne, NaN3, CuSO4·5H2O, sodium ascorbate, DMF, H2O, 100°C, 16 h, 26%; (q) TFA, DCM, rt, 1 h; (r) D-pantolactone, EtOH, NEt3, 75°C, 16 h, 14–46% over 2 steps; (s) pentanal, 1% AcOH/MeOH, rt, 5 h; (t) NaBH4, MeOH, 0°C-rt, 1 h; (u) TFA, DCM, rt, 1 h; (v) D-pantolactone, EtOH, NEt3, 75°C, 24 h, 52% over 4 steps; (w) butyric acid, EDC, HOBt, DIPEA, THF, rt, 16 h, 53%; (x) TFA, DCM, rt, 1 h; (y) Boc-β-alanine, EDC, DMAP, DIPEA, THF, rt, 16 h, 64% over 2 steps; (z) D-pantolactone, EtOH, NEt3, 75°C, 16 h, 11% over 2 steps.
Compounds 35-52
The recently reported crystal structure of KpPanK bound to Np-Pan (34) suggests that PanK substrates may interact with the enzyme via π-π interactions with nearby Tyr180 and Phe259, and potentially with Tyr240, Tyr258, and Tyr262.24 As such, derivatives 35-52 were synthesized and used in this study to probe the potential of pi-pi interactions with PanK. Np-Pan (34) was also re-synthesized because no kinetic data were reported.24 For 34, 35-39, 41-42, 44-46, and 51-52 (Scheme 2A), synthesis started with the 1,3-diol protection of commercially available pantothenate as a p-methoxyphenyldimethylacetal to yield 53, followed by activation to a N-succinamyl pantothenic ester 54.25 The desired commercially available amines were then reacted separately with 54, followed by dilute acidic removal of the acetal protecting group to afford the N-linked derivatives 34, 35-37, 39, 41-42, 45, and 51-52 in good yield. Deactivated amines such as aniline, 4-aminopyrimidine, and 3-methylpridin-2-amine under standard reaction conditions resulted in lower yields (10–20%) of 38, 44, and 46 respectively. However, under microwave irradiation at 60°C for 30 min and using HATU as the coupling agent, higher yields (82–89%) of 38, 44, and 46 were achieved. In our hands, previous synthetic protocols for pantothenamide analogues using diphenylphosphorylazide to activate pantothenate were found to generate diphenylphosphonic acid as a byproduct which contaminated the final product.7 For the synthesis of 40, 43, and 49-50 (Scheme 2B), Boc-β-alanine (56) was first coupled to the desired commercially available amine to produce 57a-d. Subsequent deprotection followed by D-pantolactone addition then afforded the desired compounds 40, 43, and 49-50.
Scheme 2.

(a) PMPCH(OMe)2, CSA, DMF, rt, 16 h, 75%; (b) NHS, DCC, THF, rt, 6 h–24 h, 90%; (c) R-NH2, DCM, rt, 16 h, 69–94%; (d) 80% AcOH, rt, 16 h, 71–94%; (e) microwave, HATU, DIPEA, DMF, 60°C, 30 min; (f) 80% AcOH, rt, 16 h, 82–89% over 2 steps; (g) R-NH2, EDC, HOBt, DIPEA, THF, rt, 16 h, 60–98%; (h) TFA, DCM, rt, 1 h; (i) D-pantolactone, EtOH, NEt3, 75°C, 16 h, 10–53% over 2 steps.
Two approaches were envisaged to assemble the triclosan derivatives 47 and 48 (Scheme 3). Synthesis of the amide-linked 47 was achieved from amine 58 which was synthesized according to a literature protocol.26 Subsequent coupling of 58 to 53 afforded 59, followed by deprotection to yield 47. Synthesis of the ether-linked triclosan derivative 48 on the other hand began with PMP-protected D-pantothenol 60,25 which underwent a Mistunobu reaction with triclosan to afford 61. PMP deprotection yielded the desired product 48.
Scheme 3.

(a) 53, HATU, DMF, DIPEA, 80°C, 16 h, 72%; (b) 80% AcOH, rt, 16 h, 90%; (c) triclosan, Ph3P, DIAD, THF, rt, 24 h; (d) 80% AcOH, rt, 16 h, 50% over 2 steps.
Compound 62-67
Finally, a previous study with C-2 geminal dialkylpantothenamides suggests that they might be good PanK substrates.11 Crystal structure analysis points to possible interactions of the alkyl group with a hydrophobic pocket consisting of Val97, Leu201, and Ile290. Compounds 62-67 were synthesized to probe this pocket and study consequent effects on substrate activity. Synthesis of C-2 geminal dialkylpantothenamide derivatives 62-67 was achieved by adapting a protocol previously reported by our group (Scheme 4).11 The known diethyl ester 68,11 was first reacted in two successive alkylations using the method developed by Seebach and Frater27 to generate derivatives 70a-f via 69a-f. Reduction to the triol followed by selective 1,3-diol protection, left the terminal alcohol of 71a-f available for oxidation to the corresponding acids 72a-f, via Dess-Martin Periodinane oxidation of the alcohol to the aldehyde followed by Pinnick oxidation. The acids 72a-f are unstable and must be used right away. Two subsequent amide couplings then afforded the desired pantothenamides 62-67.
Scheme 4.

(a) LDA, THF, −78°C, 15 min; (b) R1-X, −78°C-rt, 16 h, 67–75%; (c) R2-X, 78°C-rt, 16 h, 29–58%; (d) LAH, THF, 70°C, 16 h; (e) PMPCH(OMe)2, CSA, DCM, rt, 4 h, 50–86% over 2 steps; (f) DMP, DCM, rt, 2 h; (g) NaClO2, NaHPO4, acetone, DCM, H2O, rt, 15 min; (h) piperidine, DMF, rt, 30 min; (i) EDC, DIPEA, HOBt, THF, rt, 16 h, 42–51% over 4 steps; (j) 80% AcOH, rt, 16h, 64–81%.
2.2. Biological studies
Following the synthesis of 4-12, 35-52, and 62-67, their substrate activity with EcPanK was measured and the results are compiled in Table 1.
Table 1.
EcPanK kinetic parameters.
| Km (μM) | kcat(s−1) | kcat/Km (mM−1 s−1) | |
|---|---|---|---|
| Pantothenate (1) | |||
| exp | 21 ± 3 | 1.07 ± 0.05 | 50 ± 7 |
| lit6 | 34 ± 6 | 0.76 ± 0.03 | 21 ± 2 |
| Pantetheine (2) | |||
| exp | 60 ± 10 | 1.03 ± 0.06 | 17 ± 3 |
| lit17 | 91 ± 10 | 0.32 ± 0.02 | 3.5 ± 0.4 |
| N5-Pan (3) | 36 ± 3 | 0.93 ± 0.04 | 26 ± 2 |
| 4 | 120 ± 10 | 1.12 ± 0.05 | 10 ± 1 |
| 5 | 1200 ± 400 | 0.13 ± 0.03 | 0.1 ± 0.1 |
| 6 | NA[a] | ||
| 7 | NA | ||
| 8 | 190 ± 20 | 0.96 ± 0.04 | 5.1 ± 0.6 |
| 9 | 1000 ± 200 | 0.49 ± 0.06 | 0.5 ± 0.1 |
| 10 | 700 ± 300 | 0.23 ± 0.05 | 0.3 ± 0.2 |
| 11 | NA | ||
| 12 | 240 ± 30 | 1.09 ± 0.07 | 4.5 ± 0.6 |
| 34 | 33 ± 4 | 1.24 ± 0.06 | 38 ± 5 |
| 35 | >1000 | 1.19 ± 0.11 | 0.5 ± 0.1 |
| 36 | >1000 | 0.75 ± 0.18 | 0.3 ± 0.1 |
| 37 | 350 ± 20 | 1.03 ± 0.05 | 2.9 ± 0.3 |
| 38 | 43 ± 4 | 1.02 ± 0.05 | 24 ± 3 |
| 39 | 67 ± 6 | 1.15 ± 0.05 | 17 ± 2 |
| 40 | 32 ± 3 | 1.13 ± 0.05 | 35 ± 4 |
| 41 | 22 ± 7 | 0.46 ± 0.03 | 21 ± 6 |
| 42 | 50 ± 7 | 1.02 ± 0.05 | 20 ± 3 |
| 43 | 130± 30 | 0.94 ± 0.08 | 7 ± 2 |
| 44 | 39 ± 5 | 0.78 ± 0.04 | 20 ± 3 |
| 45 | 110 ± 10 | 1.15 ± 0.06 | 11 ± 1 |
| 46 | 450 ± 50 | 1.44 ± 0.09 | 3.2 ± 0.4 |
| 47 | NA | ||
| 48 | NA | ||
| 49 | 200 ± 100 | 0.45 ± 0.09 | 1.9 ± 0.9 |
| 50 | 400 ± 200 | 0.18 ± 0.05 | 0.4 ± 0.2 |
| 51 | NA | ||
| 52 | NA | ||
| 62 | 230 ± 20 | 1.17 ± 0.06 | 5.1 ± 0.5 |
| 63 | 1200 ± 400 | 0.28 ± 0.06 | 0.2 ± 0.1 |
| 64 | 590 ± 60 | 0.89 ± 0.06 | 1.5 ± 0.2 |
| 65 | 110 ± 10 | 1.04 ± 0.06 | 9 ± 1 |
| 66 | NA | ||
| 67 | NA | ||
NA – no observable activity up to 1 μM
While transformation of compounds 6-7, 11, 47-48, 51-52, and 66-67 by EcPanK is below our detection limit, complementary studies show that they did not inhibit the enzyme either (data not shown), suggesting that the compounds are likely not binding EcPanK. All other derivatives are substrates of EcPanK, yet their overall activity (kcat/Km) vary greatly from one to the other. A few compounds (3, 38, 39, 41, 42, 44 and 45) show comparable activity to that of pantetheine, while 34 and 40 are some of the rare compounds to rival the preferred substrate pantothenate.
3. Discussion
Crystal structures are reported for Type I PanK isoforms from four species: Escherichia coli (EcPanK),14,28 Mycobacterium tuberculosis (MtPanK),29–31 Coxiella burnetii (CbPanK; PDB 3TQC), and Klebsiella pneumoniae (KpPanK).24 The reported crystal structure of EcPanK in complex with pantothenate predicts several amino acids involved in stabilizing pantothenate binding.28 These include Asp127 and His177 forming hydrogen bonds with the C1 and C3 hydroxyl groups of pantothenate, respectively; Tyr175 hydrogen bonding with the C-4 carbonyl oxygen via H2O; and Asn282 and Tyr240 hydrogen bonding with the terminal C-7 carboxylate of pantothenate (Fig. 3A). Recently, KpPanK was also crystallized with 3 and 34,24 where analysis confirms similar roles of Asp127, His177, Tyr175, Asn282, and Tyr240 for substrate binding, whereas Asn282 and Tyr240 may hydrogen bond not only with the C-7 carboxylate of pantothenate but also with the C-7 amides of 3 and 34 (Fig. 3B–D). It is also interesting to note that 34 is able to adopt two different substrate conformations in the enzyme. One conformation has the pyridine moiety either positioned parallel to Tyr180 forming π-π stacking interactions (Fig. 3C), and the second conformation has the pyridine moiety positioned perpendicular to Phe259 forming T-shaped π-π interactions (Fig. 3D).
Figure 3.

Highlight of the conserved active site residues involved in substrate binding; A) EcPanK (PDB 1SQ5) in complex with pantothenate (1); B) KpPanK (PDB 4F7W) in complex with 3; C) KpPanK (PDB 4GI7) in complex with 34 (yellow) and in a conformation where sandwich π-π stacking interactions form with Tyr180, or D) KpPanK (PDB 4GI7) in complex with 34 (turquoise) in a conformation where T-shaped π-π interactions form with Phe259 and water-mediated hydrogen bonding between the pyridyl nitrogen and Arg243, Phe244, and Tyr240.
Based on the kinetic data obtained with EcPanK and crystal structure analysis, we were able to identify and verify several aspects of the enzyme binding pocket that are crucial for influencing the binding affinity and activity of PanK substrates. As mentioned above, the 3D structures of PanK suggest that the hydrogen bonds between the C-7 carboxylate of pantothenate and both Asn282 and Tyr240 can be sustained by an amide group (C-7 amide of N5-Pan). This is supported by very similar Km value (21 vs. 36 μM). The importance of the C-7 carbonyl group for binding EcPanK was confirmed with the 10 times difference in binding affinity between the amide-containing 4 (kcat/Km = 10 mM−1 s−1) compared to its amide-deficient counterpart, 5 (kcat/Km = 0.1 mM−1 s−1). This is also supported by modeling (Fig. 4A).
Figure 4.

Highlight of the predicted hydrogen bonding interactions of Tyr240 and Asn282 from EcPanK (green) with A) 4 (yellow) and 5 (turquoise), B) 6 (turquoise) and 1 (orange), and C) 8 (yellow) and 9 (grey).
The next series of molecules probed whether the hydrogen bonding acceptor and donor nature of the C-7 carboxylate/amide could be mimicked with triazoles,32,33 guanidines, or sulfonate. The results of the kinetic studies imply that guanidine groups (as in 6 and 7) are inadequate mimics in this context. Docking 6 into EcPanK suggest that the guanidine group might be better positioned near the hydroxyl group of Tyr240 (Fig. 4B). The aromatic group of 7 may however compete for Tyr240 thus preventing interaction between the guanine group of 7 and Tyr240. Triazole 8 on the other hand was a better mimic but still suffered a loss of activity (kcat/Km = 5.1 mM−1s−1) compared to that of 3 (kcat/Km = 26 mM−1 s−1). Docking studies predict that hydrogen bonding with Asn282 and Tyr240 should be preserved (Fig. 4C), and hence steric effects might explain the observed loss. This hypothesis was verified with compound 9, which has similar steric bulk as that of 8 but with a triazole moiety not positioned to hydrogen bond with Asn282 and Tyr240 (Fig. 4C). As expected 9 showed vastly reduced activity (kcat/Km = 0.5 mM−1 s−1) compared to 8.
Mimicking the carboxylate group was attempted with a sulfonate, however the sulfonate-containing 11 was not a substrate of PanK. A possible explanation for this observation is that the enzyme binding pocket may not tolerate the bulky tetrahedral geometry of the sulfonate group. This is further supported by the loss of activity for 10(kcat/Km = 0.3 mM−1 s−1), which is a derivative of pantothenate with a side chain at C-6.
Compounds 4, 5, and 12 were synthesized with either C-9 or C-10 amide groups. All of these compounds (kcat/Km = 0.1–1 mM−1 s−1) displayed reduced activity compared to 3 (kcat/Km = 26 mM−1 s−1). This is consistent with the lack of hydrogen bond acceptors/donors in the pantothenate binding pocket besides those aligned with C-7. Indeed the majority of residues in this area are hydrophobic, aromatic residues.
A recent study shows that 34 may adopt two different substrate conformations in KpPanK,24 hinting that aromatic rings might fit better in EcPanK. In fact, turnover rates were better for 34 (kcat/Km = 38 mM−1 s−1) than for 3 (kcat/Km = 26 mM−1 s−1). Given these results, potential π-π interactions between the enzyme and substrates were probed with additional pantothenamides containing various aromatic substituents. There was little change among binding affinities for substrates with differing chain lengths (0–3 carbons) linking the aromatic substituents to the C-7 amide, with a slight preference for 3 carbons (see 38-40). A possible explanation for this result is that the high proportion of aromatic residues in the substrate binding pocket (e.g. Tyr180, Phe259, Tyr240, Tyr258, and Tyr262) allows a high degree of flexibility for π-π interactions to form. Overall, imidazole (e.g. 41), pyridine (e.g. 34, 42-45), and phenyl groups (e.g. 38-40) were all effective for this purpose. Multi-ring aromatics (e.g. 47-48) and bulky aromatic substituents (e.g. 49-50), however had a detrimental effect. This suggests that the binding pocket can only accommodate one aromatic ring.
Another interesting observation with 34 was the position of the pyridyl nitrogen and its water-mediated hydrogen bonding with Tyr240, Phe244, and Arg243 of KpPanK.24 Our studies show that the pyridyl nitrogen brings a sizable contribution (e.g. 39, kcat/Km = 17 mM−1 s−1), and its position matters (e.g. 42, kcat/Km = 20 mM−1 s−1; 43, kcat/Km = 7.1 mM−1 s−1). The importance of the aromatic ring was further confirmed using the amine-containing, six membered ring substituted 35, 36, and 37 which all showed a loss of activity with EcPanK (kcat/Km = 0.3–2.9 mM−1 s−1) compared to 38-45. As an extra control for the increase bulk of sp3 over sp2 hybridized carbons, the less bulky linear polyamine-containing 51 and 52 were also tested, and, as predicted they were not significantly transformed by EcPanK.
Analysis of the various PanK crystal structures14,24,28–31 reveals a cluster of residues (Val97, Leu201, and Ile290) forming a small hydrophobic pocket in the enzyme active site (Fig. 5A). To explore whether substrates can interact with this pocket via van der Waals forces, a series of 3 derivatives modified at the C-2 geminal dialkyl position were prepared. Emphasis was placed on modifying the C-2 alkyl substituent anti to the C-3 hydroxyl group, which is the position on the 3 backbone that is best suited to reach into the pocket. Of these, the propynyl derivative 65 proved to be the most promising, consistent with a previous study reporting the allyl group as the best among a variety of short alkyl groups.11 A combination of reduced bulk of the propynyl substituent and stronger van der Waals forces that alkynes experience due to the increased polarity of its π-elections as a result of its linearity, and might explain this result (Fig. 5B).
Figure 5.

Highlight of the hydrophobic pocket consisting of residues Val97, Leu201, and Ile290 from EcPanK (green) in complex with 3 (yellow), and a modeled conformation of 65 (turquoise) and its potential reach into the hydrophobic binding pocket of EcPanK.
4. Conclusion
We have developed the synthetic route necessary to access pantothenate analogs diversified not only at the amine moiety but also at the β-alanine moiety, at the carboxylate, or at the geminal dimethyl group. Kinetic studies of these analogs with EcPanK revealed a number of structure-activity relationships. While some of our compounds proved to be equivalent to the benchmark compound 3, others were some of the best substrates reported for EcPanK. We believe that these results will be useful to guide the future design PanK substrates for the biosynthetic generation of CoA analogues as probes or drug candidates.
EXPERIMENTAL PROCEDURES
Docking studies
Docking was performed using FITTED via the web-based platform FORECASTER.34
Expression and purification of EcPanK
The expression vector pET28a/PanK was generously provided by Dr. Gerard D. Wright. Purification of EcPanK followed a protocol previously described.35
Kinetic assay of EcPanK
The enzyme assay couples the production of ADP to the consumption of NADH through the activity of pyruvate kinase and lactic dehydrogenase as previously described.13 Reactions were performed in 96-well microtiter plates, and the decrease in NADH concentration was monitored at 340 nm every 2 seconds using a Molecular Devices Spectramax190 plate reader. Each reaction mixture (200 μl) contained ATP (1.5 mM), NADH (0.3 mM), phosphor(enol)pyruvate (0.5 mM), MgCl2 (10 mM), KCl (20 mM), pyruvate kinase (8 units), lactic dehydrogenase (8 units) and pantothenate kinase (1.57 μM) in 50 mM Tris-HCl buffer (pH 7.6). The reaction was initiated with the addition of the desired substrate at variable concentrations (62.5–1000 μM). Kinetic parameters were determined by fitting the rate data into the Michaelis-Menten equation using GraphPad Prism 4.0 with non-linear regressions. All data points are from duplicate experiments. For inhibition studies the same conditions were used but in the presence of pantothenate (1 mM) and at varying concentration (0, 0.625, 1.25, 2.5, 5, 10, and 20 mM) of the potential inhibitor.
Synthesis
Detailed synthetic protocols and compound characterizations are provided as supporting information.
Supplementary Material
Acknowledgments
This work was supported by research grants from the Canadian Institute of Health Research (CIHR) and the National Science and Engineering Research Council of Canada (NSERC) to KA. CIHR provided funding as part of the Strategic Training Initiative in Chemical Biology (to K.V.), the Drug Development Training Program (to E.A. and A.H.), and a graduate scholarship (to A.H.). The Fonds québécois de la recherche sur la nature et les technologies (FQRNT, to K.V.) also provided a scholarship. The authors are grateful to G. D. Wright at McMaster University for sharing his PanK expression plasmid.
Footnotes
Abbreviations: CoA, coenzyme A; PanK, pantothenate kinase; EcPanK, PanK isoform from Escherichia coli; MtPanK, PanK isoform from Mycobacterium tuberculosis; CbPanK, PanK isoform from Coxiella burnetii; and KpPanK, PanK isoform from Klebsiella pneumonia.
References
- 1.Song WJ, Jackowski S. Cloning, sequencing, and expression of the pantothenate kinase (coaA) gene of Escherichia coli. J Bacteriol. 1992;174:6411–6417. doi: 10.1128/jb.174.20.6411-6417.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Strauss E. Coenzyme A Biosynthesis and Enzymology. In: Mander L, Liu HW, editors. Comprehensive Natural Products II. Elsevier; Oxford: 2010. pp. 351–410. [Google Scholar]
- 3.Gerdes SY, Scholle MD, D’Souza M, Bernal A, Baev MV, Farrell M, Kurnasov OV, Daugherty MD, Mseeh F, Polanuyer BM, Campbell JW, Anantha S, Shatalin KY, Chowdhury SAK, Fonstein MY, Osterman AL. From genetic footprinting to antimicrobial drug targets: Examples in cofactor biosynthetic pathways. J Bacteriol. 2002;184:4555–4572. doi: 10.1128/JB.184.16.4555-4572.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Strauss E, de Villiers M, Rootman I. Biocatalytic production of coenzyme A analogues. ChemCatChem. 2010;2:929–937. [Google Scholar]
- 5.Virga KG, Zhang YM, Leonardi R, Ivey RA, Hevener K, Park HW, Jackowski S, Rock CO, Lee RE. Structure-activity relationships and enzyme inhibition of pantothenamide-type pantothenate kinase inhibitors. Bioorg Med Chem. 2006;14:1007–1020. doi: 10.1016/j.bmc.2005.09.021. [DOI] [PubMed] [Google Scholar]
- 6.Mercer AC, Meier JL, Hur GH, Smith AR, Burkart MD. Antibiotic evaluation and in vivo analysis of alkynyl coenzyme A antimetabolites in Escherichia coli. Bioorg Med Chem Lett. 2008;18:5991–5994. doi: 10.1016/j.bmcl.2008.07.078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Van Wyk M, Strauss E. Development of a method for the parallel synthesis and purification of N-substituted pantothenamides, known inhibitors of coenzyme A biosynthesis and utilization. Org Biomol Chem. 2008;6:4348–4355. doi: 10.1039/b811086g. [DOI] [PubMed] [Google Scholar]
- 8.Zhang YM, Frank MW, Virga KG, Lee RE, Rock CO, Jackowski S. Acyl carrier protein is a cellular target for the antibacterial action of the pantothenamide class of pantothenate antimetabolites. J Biol Chem. 2004;279:50969–50975. doi: 10.1074/jbc.M409607200. [DOI] [PubMed] [Google Scholar]
- 9.Clifton G, Bryant SR, Skinner CG. N1-(substituted) pantothenamides, antimetabolites of pantothenic acid. Arch Biochem Biophys. 1970;137:523–528. doi: 10.1016/0003-9861(70)90470-4. [DOI] [PubMed] [Google Scholar]
- 10.Spry C, Kirk K, Saliba KJ. Coenzyme A biosynthesis: An antimicrobial drug target. FEMS Microbiol Rev. 2008;32:56–106. doi: 10.1111/j.1574-6976.2007.00093.x. [DOI] [PubMed] [Google Scholar]
- 11.Akinnusi TO, Vong K, Auclair K. Geminal dialkyl derivatives of N-substituted pantothenamides: synthesis and antibacterial activity. Bioorg Med Chem. 2011;19:2696–2706. doi: 10.1016/j.bmc.2011.02.053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Spry C, Macuamule C, Lin Z, Virga KG, Lee RE, Strauss E, Saliba KJ. Pantothenamides are potent, on-target inhibitors of Plasmodium falciparum growth when serum pantetheinase is inactivated. PLoS ONE. 2013;8 doi: 10.1371/journal.pone.0054974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Strauss E, Begley TP. The antibiotic activity of N-pentylpantothenamide results from its conversion to ethyldethia-coenzyme A, a coenzyme A antimetabolite. J Biol Chem. 2002;277:48205–48209. doi: 10.1074/jbc.M204560200. [DOI] [PubMed] [Google Scholar]
- 14.Yun M, Park CG, Kim JY, Rock CO, Jackowski S, Park HW. Structural basis for the feedback regulation of Escherichia coli pantothenate kinase by coenzyme A. J Biol Chem. 2000;275:28093–28099. doi: 10.1074/jbc.M003190200. [DOI] [PubMed] [Google Scholar]
- 15.Leonardi R, Chohnan S, Zhang YM, Virga KG, Lee RE, Rock CO, Jackowski S. A pantothenate kinase from Staphylococcus aureus refractory to feedback regulation by coenzyme A. J Biol Chem. 2005;280:3314–3322. doi: 10.1074/jbc.M411608200. [DOI] [PubMed] [Google Scholar]
- 16.Thomas J, Cronan JE. Antibacterial activity of N-pentylpantothenamide is due to inhibition of coenzyme A synthesis. Antimicrob Agents Chemother. 2010;54:1374–1377. doi: 10.1128/AAC.01473-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Worthington AS, Burkart MD. One-pot chemo-enzymatic synthesis of reporter-modified proteins. Org Biomol Chem. 2006;4:44–46. doi: 10.1039/b512735a. [DOI] [PubMed] [Google Scholar]
- 18.Meier JL, Mercer AC, Rivera H, Burkart MD. Synthesis and evaluation of bioorthogonal pantetheine analogues for in vivo protein modification. J Am Chem Soc. 2006;128:12174–12184. doi: 10.1021/ja063217n. [DOI] [PubMed] [Google Scholar]
- 19.Clarke KM, Mercer AC, La Clair JJ, Burkart MD. In vivo reporter labeling of proteins via metabolic delivery of coenzyme A analogues. J Am Chem Soc. 2005;127:11234–11235. doi: 10.1021/ja052911k. [DOI] [PubMed] [Google Scholar]
- 20.Vong K, Tam IS, Yan X, Auclair K. Inhibitors of aminoglycoside resistance activated in cells. ACS Chem Biol. 2012;7:470–475. doi: 10.1021/cb200366u. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Mandel AL, La Clair JJ, Burkart MD. Modular synthesis of pantetheine and phosphopantetheine. Org Lett. 2004;6:4801–4803. doi: 10.1021/ol048853+. [DOI] [PubMed] [Google Scholar]
- 22.Wu YQ, Hamilton SK, Wilkinson DE, Hamilton GS. Direct synthesis of guanidines using di(imidazole-1-yl)methanimine. J Org Chem. 2002;67:7553–7556. doi: 10.1021/jo0202381. [DOI] [PubMed] [Google Scholar]
- 23.Snell EE. A specific growth inhibition reversed by pantothenic acid. J Biol Chem. 1941;139:975–976. [Google Scholar]
- 24.Li B, Tempel W, Smil D, Bolshan Y, Schapira M, Park HW. Crystal structures of Klebsiella pneumoniae pantothenate kinase in complex with N-substituted pantothenamides. Proteins. 2013;81:1466–1472. doi: 10.1002/prot.24290. [DOI] [PubMed] [Google Scholar]
- 25.Patrone JD, Yao J, Scott NE, Dotson GD. Selective inhibitors of bacterial phosphopantothenoylcysteine synthetase. J Am Chem Soc. 2009;131:16340–16341. doi: 10.1021/ja906537f. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Gupta S, Babu P, Surolia A. Biphenyl ethers conjugated CdSe/ZnS core/shell quantum dots and interpretation of the mechanism of amyloid fibril disruption. Biomaterials. 2010;31:6809–6822. doi: 10.1016/j.biomaterials.2010.05.031. [DOI] [PubMed] [Google Scholar]
- 27.Fráter G, Müller U, Günther W. The stereoselective α-alkylation of chiral β-hydroxy esters and some applications thereof. Tetrahedron. 1984;40:1269–1277. [Google Scholar]
- 28.Ivey RA, Zhang YM, Virga KG, Hevener K, Lee RE, Rock CO, Jackowski S, Park HW. The structure of the pantothenate kinase·ADP·pantothenate ternary complex reveals the relationship between the binding sites for substrate, allosteric regulator, and antimetabolites. J Biol Chem. 2004;279:35622–35629. doi: 10.1074/jbc.M403152200. [DOI] [PubMed] [Google Scholar]
- 29.Bjorkelid C, Bergfors T, Raichurkar AKV, Mukherjee K, Malolanarasimhan K, Bandodkar B, Jones TA. Structural and biochemical characterization of compounds inhibiting mycobacterium tuberculosis pantothenate kinase. J Biol Chem. 2013;288:18260–18270. doi: 10.1074/jbc.M113.476473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Chetnani B, Kumar P, Surolia A, Vijayan M. M. tuberculosis pantothenate kinase: dual substrate specificity and unusual changes in ligand locations. J Mol Biol. 2010;400:171–185. doi: 10.1016/j.jmb.2010.04.064. [DOI] [PubMed] [Google Scholar]
- 31.Das S, Kumar P, Bhor V, Surolia A, Vijayan M. Invariance and variability in bacterial PanK: A study based on the crystal structure of Mycobacterium tuberculosis PanK. Acta Cryst D. 2006;62:628–638. doi: 10.1107/S0907444906012728. [DOI] [PubMed] [Google Scholar]
- 32.Li X. Click to Join Peptides/Proteins Together. Chem Asian J. 2011;6:2606–2616. doi: 10.1002/asia.201100329. [DOI] [PubMed] [Google Scholar]
- 33.Brik A, Alexandratos J, Lin YC, Elder JH, Olson AJ, Wlodawer A, Goodsell DS, Wong CH. 1,2,3-triazole as a peptide surrogate in the rapid synthesis of HIV-1 protease inhibitors. ChemBioChem. 2005;6:1167–1169. doi: 10.1002/cbic.200500101. [DOI] [PubMed] [Google Scholar]
- 34.Corbeil CR, Englebienne P, Moitessier N. Docking ligands into flexible and solvated macromolecules. 1. Development and validation of FITTED 1.0. J Chem Inf Model. 2007;47:435–449. doi: 10.1021/ci6002637. [DOI] [PubMed] [Google Scholar]
- 35.Nazi I, Koteva KP, Wright GD. One-pot chemoenzymatic preparation of coenzyme A analogues. Anal Biochem. 2004;324:100–105. doi: 10.1016/j.ab.2003.09.005. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.