

Abstract
The endoplasmic reticulum (ER) is the principal organelle responsible for several specific cellular functions including synthesis and folding of secretory or membrane proteins, lipid metabolism, and Ca2+ storage. Different physiological as well as pathological stress conditions can, however, perturb ER homeostasis, giving rise to an accumulation of unfolded or misfolded proteins in the ER lumen, a condition termed ER stress. To deal with an increased folding demand, cells activate the unfolded protein response (UPR), which is initially protective but can become detrimental if ER stress is severe and prolonged. Accumulating evidence demonstrates a link between the UPR and ovarian development and function, including follicular growth and maturation, follicular atresia, and corpus luteum biogenesis. Additionally, ER stress and the UPR may also play an important role in the ovary under pathological conditions. Understanding the molecular mechanisms related to the dual role of unfolded protein response in the ovarian physiology and pathology may reveal the pathogenesis of some reproductive endocrine diseases and provide a new guidance to improve the assisted reproductive technology. Here we review the current literature and discuss concepts and progress in understanding the UPR, and we also analyze the role of ER stress and the UPR in the ovary.
Keywords: endoplasmic reticulum stress, obesity, ovary, follicle, ovarian diseases
Introduction
The endoplasmic reticulum (ER) is a major compartment within eukaryotic cells responsible for synthesis and folding of proteins destined for the secretory pathway or insertion into the membrane, trafficking and metabolism of lipids and sterols, and cellular Ca2+ storage (Duan et al., 2015; Wang et al., 2015; Daniele and Schiaffino 2016). Some physiological or pathological conditions that perturb ER function—such as glucose deprivation (Marjon et al., 2004), hypoxia (Gao et al., 2016), aberrant Ca2+ regulation(Zhou et al., 2015), and elevated free fatty acid levels (Cui et al., 2013)—lead to accumulation of misfolded proteins within the ER, resulting in the induction of ER stress (Fig. 1).
Figure 1.
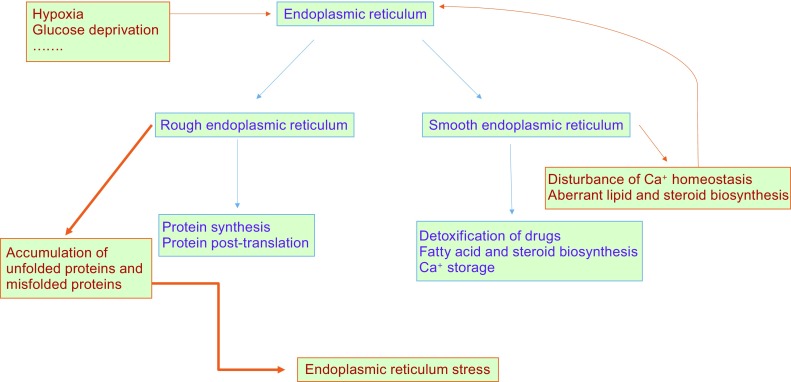
Normal ER function and ER stress. The ER has two main forms, namely the rough and smooth ER (RER and SER) according to their different structures and functions. RER is studded with abundant ribosomes and is mainly responsible for protein folding and post-translational modification. SER does not have attached ribosomes and is a primary site for drug detoxification, fatty acid and steroid biosynthesis, and Ca+ storage. Some physiological or pathological conditions, such as hypoxia and glucose deprivation, can perturb ER function, leading to accumulation of unfolded and misfolded proteins within the ER and resulting in induction of ER stress. In addition, perturbation of Ca2+ homeostasis and aberrant lipid and steroid biosynthesis may further impair ER function
To cope with ER stress, cells activate a series of protective intracellular signaling pathways, collectively termed the unfolded protein response (UPR). These include transient attenuation of translation to reduce the protein synthesis and folding load, activation of transcription of chaperone proteins and folding catalysts to expand the folding capacity of the ER, induction of ER-associated degradation (ERAD) to remove terminally misfolded proteins (i.e., that fail to fold into the correct native structure or assemble into proper protein complexes), and induction of apoptosis (Ron and Walter 2007; Olzmann et al., 2013). The UPR pathway elicits paradoxical outputs, inducing cytoprotective functions that reestablish homeostasis and cytodestructive functions that promote apoptosis, which is initially an effective means to eliminate a minority of cells that cannot recover from ER stress and protect the organism from damage induced by ER stress (Tabas and Ron, 2011; Iurlaro and Munoz-Pinedo 2016). Whether the UPR promotes cell survival or cell death depends on the duration and severity of the ER stress (Shore et al., 2011). The UPR can promote survival under conditions of transient and mild ER stress, or conversely promote cell death by activating downstream apoptosis signaling molecules if ER stress is prolonged and severe. Although cell death under severe ER stress may guard organisms from exposure to improperly folded proteins, many prevalent human diseases—such as diabetes mellitus (Back et al., 2012; Brozzi and Eizirik 2016), acute lung injury (Hu et al., 2015; Li et al., 2015), and retinopathies (Rana et al., 2014; Cai et al., 2015)—may be caused by excessive ER stress–induced cell death (Fig. 2).
Figure 2.
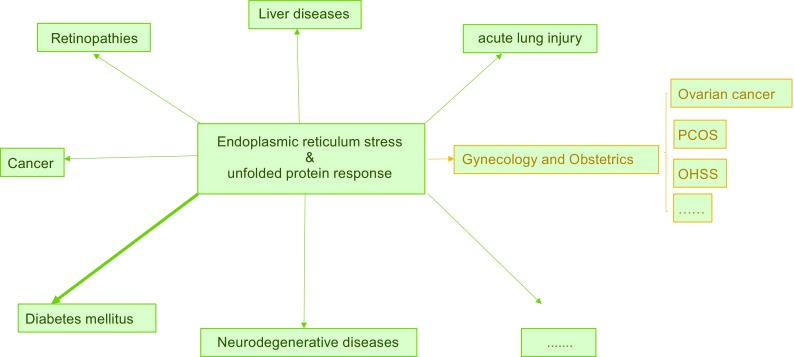
UPR and related diseases. Increasing evidence demonstrates the relationship between the unfolded protein response and diseases that involve different human organs. PCOS, polycystic ovarian syndrome; OHSS, ovarian hyperstimulation syndrome
The ovary has dual roles, i.e., reproductive function (responsible for the differentiation and release of mature oocytes for fertilization) and endocrine function (in charge of the synthesis and secretion of sex hormones, such as estrogen, progesterone and androgen). Follicles are the fundamental functional units of the mammalian ovary and consist of an innermost single oocyte, the surrounding granulosa cells (GCs) and the outer layers of theca interna cells, and theca externa cells. Beginning with the recruitment of primordial follicles from a reserve pool into the growing pathway, follicles develop through four major stages: primary, secondary, antral, and preovulatory (McGee and Hsueh 2000; Monniaux et al., 2014). Follicular growth is achieved through an increase in oocyte size accompanied by proliferation of the surrounding GCs. After the secondary follicular stage, the rate of oocyte growth and GC proliferation can increase progressively, which may result in a hypoxic condition (Monniaux et al., 1997; Monniaux et al., 2016). Such local conditions may contribute to ER dysfunction, leading to accumulation of unfolded and misfolded proteins that induce ER stress and the UPR (Harada et al., 2015). Under endocrine and paracrine control, one follicle is selected among a cohort of simultaneously growing follicles, and the other follicles enter follicular atresia (Scaramuzzi et al., 2011), which may be induced in part by ER stress (Lin et al., 2012). The corpus luteum develops from the follicle cells surrounding the ovulatory follicle and functions as a major site for the synthesis and secretion of progesterone. Some studies have indicated a link between the UPR and progesterone synthesis (Park et al., 2013a). In addition to their functions in ovarian physiology, ER stress and the UPR may also play a role in ovarian pathology.
Canonical UPR: a bifunctional response causing cell survival or cell death
As a homeostatic signaling pathway, the UPR affords unfolded or misfolded proteins an additional opportunity to fold into their native state triggered by the interactions of three ER stress signaling sensors: protein kinase RNA (PKR)-like ER kinase (PERK), inositol-requiring enzyme 1α (IRE1α), and activating transcription factor 6α (ATF6α). In the absence of ER stress, these sensors bind to the ER-resident glucose-regulated protein 78 (GRP78) chaperones in their intraluminal domains (N-terminal portion of IRE1α and PERK and C-terminal portion of ATF6) and thereby render the sensors inactive (Bertolotti et al., 2000; Shen et al., 2005). Upon stress, accumulated unfolded or misfolded proteins are sensed by the luminal domain of transmembrane proteins, resulting in the dissociation of stable GRP78-sensor complexes (consisting of ATF6, IRE1α, PERK) and inducing oligomerization of PERK and IRE1α as well as transport of ATF6 from the ER to the Golgi (Bertolotti et al., 2000; Shen et al., 2005). The role of GRP78 as a central regulator of the UPR makes it a master marker for detecting the induction of ER stress (Lee, 2005). In addition, some researchers have found that PERK and IRE1α can directly sense unfolded proteins through their core ER luminal domain to activate downstream pathways (Gardner and Walter 2011) (Fig. 3).
Figure 3.
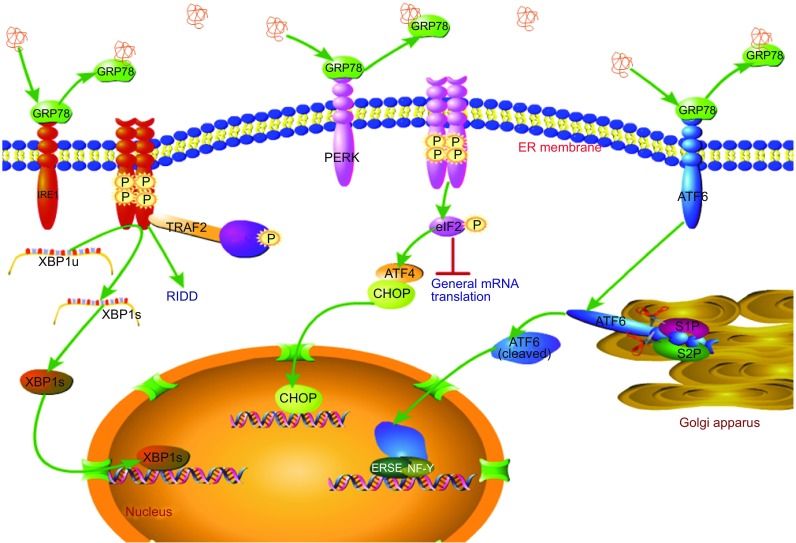
Canonical UPR pathway. The UPR is triggered by three ER stress sensors—PERK, IRE1α, and ATF6—which are rendered inactive by binding to GRP78 under non-stressed conditions. Progressive accumulation of misfolded proteins during ER stress results in dissociation of GRP78 from the GRP78-sensor complex, inducing dimerization of PERK and IRE1α as well as transport of ATF6 from the ER to the Golgi. IRE1α homodimerization and autophosphorylation initiates its kinase and endoribonuclease activities to generate an important transcription factor XBP1 s, which enters the nucleus to regulate the expression of a wide array of genes involved in ER function. Severe and chronic ER stress causes excess activation of the IRE1α kinase and RNase activities, leading to regulated IRE1α-dependent decay and phosphorylation of c-Jun terminal kinase (JNK). Similarly, PERK homodimerization and autophosphorylation induces phosphorylation of the α subunit of translation initiation factor 2 (eIF2α), which blocks formation of ribosomal preinitiation complexes and suppresses translation initiation, thereby attenuating general mRNA translation. PERK activation paradoxically promotes transcription initiation of the gene encoding ATF4 even when eIF2α is substantially phosphorylated, leading to upregulation of CHOP Once activated by ER stress, ATF6 moves to the Golgi, where it is cleaved by S1P and S2P proteases to generate an active form that mediates transcriptional induction of ER-localized molecular chaperones and folding enzymes
PERK pathway
PERK is a single-pass ER transmembrane kinase that is synthesized as a type I transmembrane glycoprotein (Haze et al., 1999). Upon activation by the accumulation of unfolded proteins in the ER lumen, PERK phosphorylates the Ser51 residue of the α subunit of translation initiation factor 2 (eIF2α) (Harding et al., 2000; Scheuner et al., 2001), which blocks formation of ribosomal preinitiation complexes and suppresses tion of translation, thereby decreasing the number of proteins translocated into the ER (Marciniak et al., 2006).
Although translation of most mRNAs is attenuated by limiting eIF2α, a subset of mRNAs is preferentially translated even with substantial eIF2α phosphorylation. In this way, PERK activation paradoxically promotes the transcription initiation of ATF4 (Harding et al., 2000) and induces ATF4 binding to the C/EBP-ATF composite site in the CHOP promoter to cause the transcription of the gene encoding CHOP (Ma et al., 2002). CHOP is a member of the C/EBP family of transcription factors and can induce apoptosis via several pathways (McCullough et al., 2001; Novoa et al., 2001; Marciniak et al., 2004; Yamaguchi and Wang, 2004; Lu et al., 2014).Some researchers have found that activation of TLR–TRIF signaling (e.g., by low dose of lipopolysaccharide) can selectively suppress ATF4-CHOP expression under prolonged ER stress, which may provide a promising therapeutic strategy for some degenerative diseases related to activation of CHOP (Woo et al., 2009).
IRE1α pathway
IRE1α, the most ancient of the three signaling sensors, consists of an N-terminal luminal domain, a single-pass transmembrane-spanning segment, and a cytosolic region subdivided into a Ser/Thr protein kinase domain and a C-terminal endoribonuclease (RNase) domain (Calfon et al., 2002). It is synthesized as a type I transmembrane glycoprotein. (Haze et al., 1999). Accumulation of unfolded and misfolded proteins activates IRE1α and causes oligomerization of the N-terminal luminal domain (Credle et al., 2005). Serving as a bifunctional enzyme possessing both a protein kinase and a site-specific RNase, activation of IRE1α promotes autophosphorylation of its cytoplasmic kinase domain, which leads to homo-oligomerization of the kinase/RNase domains (Korennykh et al., 2008). This gives the RNase the ability to site-specifically splice XBP1 mRNA, removing a 26-base intron and generating a form of the mRNA that encodes a transcriptional activator protein to enhance ER folding capacity and quality control (Yoshida et al., 2001; Calfon et al., 2002; Ghosh et al., 2014).
Under acute ER stress, IRE1α induces an adaptive response that promotes cell survival. However, chronic ER stress causes increased phosphorylation of the IRE1α kinase, proportionally increasing the oligomeric state of the kinase/RNase subunits past a critical threshold, thereby driving RNase activity into a hyperactive state and relaxing its substrate specificity to endonucleolytically cleave many other mRNAs. Consequently, high RNase activity leads to endonucleolytic decay of ER-localized mRNAs (Han et al., 2009) and activation of thioredoxin-interacting protein (Ghosh et al., 2014). Furthermore, increased oligomerization and activation of kinase/RNase subunits are sufficient for initiating the phosphorylation/activation of c-Jun terminal kinase (JNK) via clustering of the N-terminal effector domain of TRAF2, an adaptor protein that is bound to the C-terminal cytoplasmic protein IRE1 and couples the plasma membrane receptor to JNK activation (Urano et al., 2000).
The UPR can activate the mitochondrial apoptotic pathway, which is critically regulated] by several members of the Bcl-2 family. This family can be functionally classified as anti-apoptotic (i.e., Bcl-2, Bcl-XL, Bcl-w) and pro-apoptotic (i.e., Bad, Bid, Bik, Bim, Bad) (Martinou and Youle, 2011). Some studies have demonstrated that ER stress induces conformational changes in Bax and Bak, changing them from inactive to active forms and setting the apoptotic pathway in motion (Zong et al., 2003). The ATF4-CHOP pathway can downregulate the expression of Bcl-2 (McCullough et al., 2001), and the IRE1α-JNK pathway can lead to phosphorylation and subsequent inactivation of Bcl-2 and Bcl-XL (Fan et al., 2000). Taken together, these results suggest that the Bcl-2 family plays a crucial role in regulating ER stress–induced apoptosis.
ATF6α pathway
ATF6 is synthesized as a type II transmembrane glycoprotein and is embedded in the ER membrane (Haze et al., 1999). It has a luminal domain, a single transmembrane domain, and a cytoplasmic domain consisting of an N-terminal bZIP domain and a transcriptional activation domain (TAD). When unfolded proteins accumulate in the ER, the CD1 region of the luminal domain senses this ER stress and provides a positive signal for ATF6 translocation from the ER to the Golgi (Chen et al., 2002; Sato et al., 2011), in which reside the site 1 and site 2 proteases (S1P and S2P, the enzymes that process SREBPs in response to cholesterol deprivation) (Ye et al., 2000). ATF6 is cleaved in a two-step process in the Golgi, first by S1P in the luminal domain (near amino acid 418) and then by S2P near the junction between the cytoplasmic and transmembrane domains, allowing dissociation of the N-terminal bZIP and TAD (Ye et al., 2000). The TAD then moves to the nucleus, where it activates transcription of target genes by recognizing and directly binding the ER stress element (ERSE) in cooperation with the transcription factor NF-Y, which is also bound to the ERSE (Yoshida et al., 1998; Yoshida et al., 2000). ATF6-mediated transcriptional induction of ER-localized molecular chaperones and folding enzymes can improve the ER folding capacity, contributing to the maintenance of ER homeostasis in mammals (Yoshida et al., 1998). In addition to promoting expression of chaperones, cleaved ATF6 also transactivates CHOP and XBP1 by binding to the ERSE together with NF-Y, which initialized a pathway that converges with the PERK/IRE1α pathway (Yoshida et al., 2000; Ma et al., 2002; Guo et al., 2014).
Activation of the UPR in the ovarian cycle
As mentioned above, the follicle is the fundamental functional unit of the ovary. The ovarian cycle describes the normal changes that occur in the follicles and can be divided into three phases: follicular, ovulation, and luteal.
Follicular growth and maturation
Recent evidence suggests the involvement of the UPR in the physiology of follicular growth and maturation. XBP1 s and HSPA5 (also known as GRP78) are expressed in GCs of late-stage follicles (i.e., at larger secondary, antral and pre-ovulatory stages), and this expression is accompanied by activation of IRE1 and PERK (Harada et al., 2015). During follicular growth after the secondary follicle stage, more protein must be synthesized to support proliferation of GCs. Over-loaded proteins] in the ER can induce ER stress and subsequently induce UPR pathways Some findings have demonstrated that the levels of XBP1 s mRNA in CC(cumulus cells) enclosingoocytes that achieve fertilization were higher than those in CC enclosing oocytes without the capacity for fertilization, and this may be related to the pro-angiogenic role of XBP1 (Calfon et al., 2002; Zeng et al., 2013; Harada et al., 2015).
Follicle-stimulating hormone (FSH) is the major stimulator of follicle growth and development in the final and preantral stages (Oktay et al., 1997; Hsueh et al., 2015; Babayev et al., 2016). FSH attenuates ER stress in mouse GCs in vivo and in vitro (Babayev et al., 2016). Conversely, induction of severe ER stress inhibits the FSH response and negates the effects of FSH on GCs expressed ER stress-associated gene (Babayev et al., 2016).
Follicular atresia
In each reproductive cycle, only a limited number of follicles in the mammalian ovary undergo maturation and ovulation, whereas most follicles undergo a degenerative process known as atresia. Many studies have demonstrated that follicular atresia is predominantly regulated by GC apoptosis (Asselin et al., 2000). Severe and persistent ER stress will overwhelm adaption mechanisms, causing the UPR to initiate apoptosis. Thus, it is possible that ER stress plays an important role in regulating GC apoptosis. The protein GRP78 is present in GCs of non-atretic and atretic follicles of goat ovaries, but levels are higher in atretic follicles than in non-atretic ones. In contrast, CHOP is detected in GCs of atretic follicles but not in those of non-atretic follicles. Furthermore, the localization of GRP78 and DDIT3 on the antral side of the granulosa layer of atresia follicles (Lin et al., 2012) is similar to the localization of apoptotic cells in atretic follicles of goat ovaries (Bhardwaj and Sharma, 2011). ATF6 and ATF4 mRNAs are also increased during GC apoptosis (Lin et al., 2012). In addition, spontaneous apoptosis is also observed in vitro in GCs exposed to tunicamycin and serum withdrawal treatment through the activation of ER stress (Lin et al., 2012), which further demonstrates the crucial role of the UPR in the regulation of GC apoptosis. The low level of GRP78 found in healthy follicles indicates that GRP78 may play an important role in GC proliferation (Luo et al., 2006) and follicular development, and the high level of GRP78 and DDIT3 found in atretic follicles demonstrates that ER stress-induced apoptosis may act in the regulation of selective GC apoptosis in goat ovaries (McCullough et al., 2001; Novoa et al., 2001; Marciniak et al., 2004; Yamaguchi and Wang 2004; Woo et al., 2009).
Death ligand-receptor and mitochondria-mediated apoptotic systems have been shown to be active in GCs of mammalian ovaries (Manabe et al., 2008; Zhang et al., 2015a). However, several signaling molecules involved in these systems (including JNK, Bcl-2, Bax and TRAF) are also present in the UPR and induce apoptosis (Urano et al., 2000). Cross-talk between these two pathways and the UPR indicates that the ER stress system may play an indirect role through interaction with death ligand-receptor and mitochondria-mediated molecular signaling pathways in the regulation of GC apoptosis.
Corpus luteum biogenesis
The corpus luteum is a transient endocrine organ derived from ovulated follicles (Devoto et al., 2009). The main function of the corpus luteum is to synthesize and secrete progesterone, which is indispensable for maintenance of pregnancy and regulation of the estrous cycle (Stouffer 2003; Devoto et al., 2009). The luteal phase can be divided into three main stages, namely development, maintenance, and regression, according to the macroscopic appearance of corpus luteum tissues (Stouffer et al., 2013). The pathway for progesterone synthesis mainly involves three steroidogenic enzymes, namely steroidogenic acute regulatory protein (known as StAR), p450 cholesterol side-chain cleavage enzyme (p450), and 3β-hydroxysteroid dehydrogenase, and two cellular organelles (mitochondria and ER) (Niswender 2002; Rekawiecki et al., 2008). As mentioned previously, the ER is important not only for protein synthesis and lipid metabolism but also for steroid hormone synthesis. Therefore, ER stress and the UPR may play a significant role in regulating the production of progesterone and the development of the corpus luteum. All three UPR signaling pathways, including adaptive or apoptotic signaling molecules (Grp78, p-eIF2α, ATF4, CHOP, ATF6, p-IRE1α, XBP1 s and p-JNK), are activated during luteal-phase progression during the estrous cycle of bovines] (Park et al., 2013a) and mice (Park et al., 2014). Pro-apoptotic signaling molecules such as cleaved caspase3, JNK, and CHOP were detected, and JNK activation and CHOP expression via ER stress–mediated pro-apoptotic signaling cascades occurred prior to caspase3 activation during the regression stage of the corpus luteum (Urano et al., 2000; McCullough et al., 2001; Novoa et al., 2001; Marciniak et al., 2004; Yamaguchi and Wang 2004; Woo et al., 2009; Park et al., 2013a; Park et al., 2014). Furthermore, the dynamic change of these molecules seems to have some connection with the steroidogenic enzymes, which suggests that the UPR may influence the expression of steroidogenic enzymes (Park et al., 2013a; Park et al., 2013b). Molecules involved in UPR signaling, as activated during different stages of the luteal phase, may be related to progression of the luteal phase.
In conclusion, ER stress and the short-term UPR as an adaptive response are beneficial and necessary in the physiology of follicular growth and maturation.
Obesity and aberrant activation of the UPR in the ovary
Obesity has become a worldwide public health concern, as it is a major factor leading to development of insulin resistance, type 2 diabetes, fatty liver disease, and some cancers (Grundy 2015; Font-Burgada et al., 2016; Baidal and Lavine 2016). Obesity impairs oocyte function and causes alterations in the follicle and oocyte, which induces various types of reproduction dysfunction, including reduced conception, infertility, and early pregnancy loss (Hou et al., 2016; Sessions-Bresnahan et al., 2016). It is well established that obesity is closely connected with ER stress and the UPR.
The lipid composition of the ER membrane is unique, as the cholesterol content is particularly low (Feng et al., 2003) and unsaturated phosphatidylcholine is the major phospholipid (Leamy et al., 2013). This special membrane composition allows the ER to maintain a high degree of fluidity that facilitates its function. Abnormal incorporation of free cholesterol or saturated phospholipid species can result in detrimental stiffening of cellular membranes and loss of function. A large amount of free cholesterol trafficking to the ER of macrophages increases cholesterol-induced lipid ordering of the ER membrane and perturbs membrane protein conformation and function, providing a plausible mechanism for the decrease in SERCA activity induced by loading of the membrane with free cholesterol (Li et al., 2004). In addition, a recent study showed that the increased phosphatidylcholine: phosphatidylethanolamine ratio found in the ER of obese liver (both genetically induced and diet-induced) significantly inhibits SERCA function (Fu et al., 2011). The ER lumen is the major intracellular site for Ca2+ storage and release and provides a unique environment with a high concentration of Ca2+-binding proteins. This directly influences the function of the ER, affecting its roles in protein and sterol synthesis, lipid metabolism, and signal transduction (Coe and Michalak, 2009; Krebs et al., 2015). Inhibition of SERCA causes depletion of ER Ca2+ stores and disruption of ER Ca2+ homeostasis, leading to induction of ER stress and the UPR to control damage and re-establish ER homeostasis (Guerrero-Hernandez et al., 2014).
Increased body mass index in women is associated with elevated triglycerides, insulin, and free fatty acids in ovarian follicular fluid. (Pantasri et al., 2015; Robker et al., 2009). Treatment of mouse oocytes with this lipid-rich follicular fluid impairs oocyte maturation and induces ER stress and the UPR (Robker et al., 2009; Yang et al., 2012; Pantasri et al., 2015). Another study directly demonstrated that a high-fat diet dramatically increases the lipid content in mouse oocytes, both before and after ovulation, and induces ER stress pathway genes, alters mitochondrial membrane potential, and increases the incidence of apoptosis in ovarian cells. The UPR marker ATF4 is specifically upregulated in GCs and COCs (cumulus-oocyte complex)isolated from pre-ovulatory follicles of mice given a high-fat diet (GRP78 is also upregulated in COCs). ATF4 is also upregulated in the GCs of obese women (Wu et al., 2010). Compared with mice on a control diet, mice given a high-fat diet have higher rates of anovulation and lower rates of fertilization (Wu et al., 2010).
Palmitic acid, a saturated fatty acid that is a physiological component in human follicular fluid, is a potential physiological inducer of ER stress (Danino et al., 2015; Haywood and Yammani, 2015). Exposure of COCs to a high dose of palmitic acid can induce ER stress and the UPR, accompanied with reduced secretion of PTX3 (a protein marker for oocyte development and fertilization; (Zhang et al., 2005; Baranova et al., 2014), mitochondrial dysfunction and impaired oocyte maturation, and fertilization. These results are similar to those observed in COCs treated with the classical ER stress inducer thapsigargin (Wu et al., 2012). Notably, each of these defects is restored with the ER stress inhibitor salubrinal, further demonstrating that ER stress is a key mechanism mediating fatty acid–induced defects in oocyte and developmental potential (Wu et al., 2012).
The UPR and ovarian diseases
As an intracellular defense mechanism to attenuate ER stress and maintain organism homeostasis, the UPR is involved in the pathogenesis] of numerous human diseases, including ovarian diseases. Recent studies have shown a positive correlation between spliced XBP1 (induced by ER stress) and ovarian hyperstimulation syndrome (OHSS), a major complication during infertility treatment that is mainly characterized by increased capillary permeability (Elchalal and Schenker, 1997; Takahashi et al., 2016). Compared with patients without OHSS, follicles from OHSS patients express higher levels of XBP1 s in cumulus cells, which partly mediates upregulation of VEGFA, a key molecule in angiogenesis in the ovary that is regarded as a major cause of OHSS induction (Kosaka et al., 2007; Takahashi et al., 2016).
In primary solid tumors, the UPR is activated as a result of aberrant regulation of protein synthesis in cancer cells and changes in the tumor microenvironment, such as hypoxia, nutrition deprivation, and low pH (Brown and Giaccia, 1998; Giampietri et al., 2015). Cancer cells have the unique ability to exploit the UPR and its capacity to promote survival and growth. The UPR-related proteins GRP78and ATF6 are highly expressed in cancer cells and are involved in promoting cancer cell proliferation and survival under extreme conditions (Fu et al., 2007; Ye et al., 2010). Spliced XBP1 is a crucial factor for stimulating cancer progression by promoting tumor cell survival and metastatic potential and driving dendritic cell dysfunction to further inhibit anti-cancer immunity (Chen et al., 2014; Cubillos-Ruiz et al., 2015). An increasing number of studies have investigated similar roles the UPR may play in ovarian cancer and explored the potential for developing a promising target for therapy (McLean et al., 2009; De Carolis et al., 2014; Cubillos-Ruiz et al., 2015; Zhang et al., 2015b).
Conclusions
The UPR, induced by ER stress, is an indispensable response for cells to restore cellular homeostasis; conversely, it may contribute to cell death when over-activated. The molecular pathways involved in the UPR have been clearly elucidated. Accumulating evidence suggests that ER stress and the UPR play a significant role in regulating ovarian structure and function, but our understanding of the molecular mechanisms involved remains incomplete. Future studies should further focus on the molecular mechanisms related to the dual role of the UPR in ovarian physiology and pathology to better understand changes in the ovarian cycle and provide new guidance to improve assisted reproductive technology.
Acknowledgments
This work was supported in part by the National Basic Research Program (973 Program) (No. 2014CB943203), “Reproductive health and major birth defects prevention and control research” Key Special Fund (No. 2016YFC1000601), the National Natural Science Foundation of China (Grant Nos.31371521 and 81571400), Beijing Nova Program (xxjh2015011), and Special Projects for Cultivation and Development of Science and Technology Innovation Base (No. Z151100001615023).
Abbreviations
ATF6α, activating transcription factor 6α; COC, cumulus-oocyte complex; ER, endoplasmic reticulum; ERAD, ER-associated degradation; FSH, follicle-stimulating hormone; GC, granulosa cell; GRP78, glucose-regulated protein 78; IRE1α, inositol-requiring enzyme 1α; JNK, c-Jun terminal kinase; PEAK, PKR-like ER kinase; PKR, protein kinase RNA; TAD, transcriptional activation domain; UPR, unfolded protein response.
Compliance with ethics guidelines
Ning Huang, Yang Yu and Jie Qiao declare that they have no conflict of interest.
This article does not contain any studies with human or animal subjects performed by the any of the authors.
Contributor Information
Yang Yu, Email: yuyang5012@hotmail.com.
Jie Qiao, Email: jie.qiao@263.net.
References
- Asselin E, Xiao YF, Wang CW, Tsang BK. Mammalian follicular development and atresia—role of apoptosis. Biol Signals Recept. 2000;9:87–95. doi: 10.1159/000014627. [DOI] [PubMed] [Google Scholar]
- Babayev E, Lalioti MD, Favero F, Seli E. Cross-talk between FSH and endoplasmic reticulum stress: a mutually suppressive relationship. Reprod Sci. 2016;23:352–364. doi: 10.1177/1933719115602770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Back SH, Kang SW, Han J, Chung HT. Endoplasmic reticulum stress in the beta-cell pathogenesis of type 2 diabetes. Exp Diabetes Res. 2012;2012:618396. doi: 10.1155/2012/618396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baidal JAW, Lavine JE. The intersection of nonalcoholic fatty liver disease and obesity. Sci Transl Med. 2016;8(323):323rv1. doi: 10.1126/scitranslmed.aad8390. [DOI] [PubMed] [Google Scholar]
- Baranova NS, Inforzato A, Briggs DC, Tilakaratna V, Enghild JJ, Thakar D, Milner CM, Day AJ, Richter RP. Incorporation of pentraxin 3 into hyaluronan matrices is tightly regulated and promotes matrix cross-linking. J Biol Chem. 2014;289:30481–30498. doi: 10.1074/jbc.M114.568154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bertolotti A, Zhang Y, Hendershot LM, Harding HP, Ron D. Dynamic interaction of BiP and ER stress transducers in the unfolded-protein response. Nat Cell Biol. 2000;2:326–332. doi: 10.1038/35014014. [DOI] [PubMed] [Google Scholar]
- Bhardwaj JK, Sharma RK. Scanning electron microscopic changes in granulosa cells during follicular atresia in Caprine ovary. Scanning. 2011;33:21–24. doi: 10.1002/sca.20217. [DOI] [PubMed] [Google Scholar]
- Brown JM, Giaccia AJ. The unique physiology of solid tumors: opportunities (and problems) for cancer therapy. Cancer Res. 1998;58:1408–1416. [PubMed] [Google Scholar]
- Brozzi F, Eizirik DL. ER stress and the decline and fall of pancreatic beta cells in type 1 diabetes. Upsala J Med Sci. 2016;121:133–139. doi: 10.3109/03009734.2015.1135217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cai X, Chen L, McGinnis JF. Correlation of ER stress and retinal degeneration in tubby mice. Exp Eye Res. 2015;140:130–138. doi: 10.1016/j.exer.2015.08.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Calfon M, Zeng H, Urano F, Till JH, Hubbard SR, Harding HP, Clark SG, Ron D. IRE1 couples endoplasmic reticulum load to secretory capacity. Nature. 2002;415:92–96. doi: 10.1038/415092a. [DOI] [PubMed] [Google Scholar]
- Chen X, Shen J, Prywes R. The luminal domain of ATF6 senses endoplasmic reticulum (ER) stress and causes translocation of ATF6 from the ER to the Golgi. J Biol Chem. 2002;277:13045–13052. doi: 10.1074/jbc.M110636200. [DOI] [PubMed] [Google Scholar]
- Chen X, Iliopoulos D, Zhang Q, Tang Q, Greenblatt MB, Hatziapostolou M, Lim E, Tam WL, Ni M, Chen Y, Mai J, Shen H, Hu DZ, Adoro S, Hu B, Song M, Tan C, Landis MD, Ferrari M, Shin SJ, Brown M, Chang JC, Liu XS, Glimcher LH. XBP1 promotes triple-negative breast cancer by controlling the HIF1alpha pathway. Nature. 2014;508:103–107. doi: 10.1038/nature13119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coe H, Michalak M. Calcium binding chaperones of the endoplasmic reticulum. Gen Physiol Biophys. 2009;28:F96–F103. [PubMed] [Google Scholar]
- Credle JJ, Finer-Moore JS, Papa FR, Stroud RM, Walter P. On the mechanism of sensing unfolded protein in the endoplasmic reticulum. Proc Natl Acad Sci USA. 2005;102:18773–18784. doi: 10.1073/pnas.0509487102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cubillos-Ruiz JR, Silberman PC, Rutkowski MR, Chopra S, Perales-Puchalt A, Song M, Zhang S, Bettigole SE, Gupta D, Holcomb K, Ellenson LH, Caputo T, Lee AH, Conejo-Garcia JR, Glimcher LH. ER stress sensor XBP1 controls anti-tumor immunity by disrupting dendritic cell homeostasis. Cell. 2015;161:1527–1538. doi: 10.1016/j.cell.2015.05.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cui W, Ma J, Wang X, Yang W, Zhang J, Ji Q. Free fatty acid induces endoplasmic reticulum stress and apoptosis of beta-cells by Ca2+/calpain-2 pathways. PLoS One. 2013;8:e59921. doi: 10.1371/journal.pone.0059921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daniele T, Schiaffino MV. Lipid transfer and metabolism across the endolysosomal-mitochondrial boundary. Biochim Biophys Acta. 2016;1861:880. doi: 10.1016/j.bbalip.2016.02.001. [DOI] [PubMed] [Google Scholar]
- Danino H, Ben-Dror K, Birk R. Exocrine pancreas ER stress is differentially induced by different fatty acids. Exp Cell Res. 2015;339:397–406. doi: 10.1016/j.yexcr.2015.09.022. [DOI] [PubMed] [Google Scholar]
- De Carolis S, Salvi S, Botta A, Garofalo S, Garufi C, Ferrazzani S, De Carolis MP. The impact of primary Sjogren’s syndrome on pregnancy outcome: our series and review of the literature. Autoimmun Rev. 2014;13:103–107. doi: 10.1016/j.autrev.2013.09.003. [DOI] [PubMed] [Google Scholar]
- Devoto L, Fuentes A, Kohen P, Cespedes P, Palomino A, Pommer R, Munoz A, Strauss JF., 3rd The human corpus luteum: life cycle and function in natural cycles. Fertil Steril. 2009;92:1067–1079. doi: 10.1016/j.fertnstert.2008.07.1745. [DOI] [PubMed] [Google Scholar]
- Duan XQ, Li YH, Zhang XY, Zhao ZT, Wang Y, Wang H, Li GS, Jing L. Mechanisms of intracellular calcium homeostasis in MC3T3-E1 cells and bone tissues of sprague-dawley rats exposed to fluoride. Biol Trace Elem Res. 2015;170:331. doi: 10.1007/s12011-015-0465-6. [DOI] [PubMed] [Google Scholar]
- Elchalal U, Schenker JG. The pathophysiology of ovarian hyperstimulation syndrome—views and ideas. Hum Reprod. 1997;12:1129–1137. doi: 10.1093/humrep/12.6.1129. [DOI] [PubMed] [Google Scholar]
- Fan M, Goodwin M, Vu T, Brantley-Finley C, Gaarde WA, Chambers TC. Vinblastine-induced phosphorylation of Bcl-2 and Bcl-XL is mediated by JNK and occurs in parallel with inactivation of the Raf-1/MEK/ERK cascade. J Biol Chem. 2000;275:29980–29985. doi: 10.1074/jbc.M003776200. [DOI] [PubMed] [Google Scholar]
- Feng B, Yao PM, Li Y, Devlin CM, Zhang D, Harding HP, Sweeney M, Rong JX, Kuriakose G, Fisher EA, Marks AR, Ron D, Tabas I. The endoplasmic reticulum is the site of cholesterol-induced cytotoxicity in macrophages. Nat Cell Biol. 2003;5:781–792. doi: 10.1038/ncb1035. [DOI] [PubMed] [Google Scholar]
- Font-Burgada J, Sun B, Karin M. Obesity and cancer: the oil that feeds the flame. Cell Metab. 2016;23:48–62. doi: 10.1016/j.cmet.2015.12.015. [DOI] [PubMed] [Google Scholar]
- Fu Y, Li J, Lee AS. GRP78/BiP inhibits endoplasmic reticulum BIK and protects human breast cancer cells against estrogen starvation-induced apoptosis. Cancer Res. 2007;67:3734–3740. doi: 10.1158/0008-5472.CAN-06-4594. [DOI] [PubMed] [Google Scholar]
- Fu S, Yang L, Li P, Hofmann O, Dicker L, Hide W, Lin X, Watkins SM, Ivanov AR, Hotamisligil GS. Aberrant lipid metabolism disrupts calcium homeostasis causing liver endoplasmic reticulum stress in obesity. Nature. 2011;473:528–531. doi: 10.1038/nature09968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gao Y, Jia P, Shu W, Jia D. The protective effect of lycopene on hypoxia/reoxygenation-induced endoplasmic reticulum stress in H9C2 cardiomyocytes. Eur J Pharmacol. 2016;774:71. doi: 10.1016/j.ejphar.2016.02.005. [DOI] [PubMed] [Google Scholar]
- Gardner BM, Walter P. Unfolded proteins are Ire1-activating ligands that directly induce the unfolded protein response. Science. 2011;333:1891–1894. doi: 10.1126/science.1209126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghosh R, Wang L, Wang ES, Perera BG, Igbaria A, Morita S, Prado K, Thamsen M, Caswell D, Macias H, Weiberth KF, Gliedt MJ, Alavi MV, Hari SB, Mitra AK, Bhhatarai B, Schurer SC, Snapp EL, Gould DB, German MS, Backes BJ, Maly DJ, Oakes SA, Papa FR. Allosteric inhibition of the IRE1alpha RNase preserves cell viability and function during endoplasmic reticulum stress. Cell. 2014;158:534–548. doi: 10.1016/j.cell.2014.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giampietri C, Petrungaro S, Conti S, Facchiano A, Filippini A, Ziparo E. Cancer microenvironment and endoplasmic reticulum stress response. Mediat Inflamm. 2015;2015:417281. doi: 10.1155/2015/417281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grundy SM. Metabolic syndrome update. Trends Cardiovasc Med. 2015;26:364. doi: 10.1016/j.tcm.2015.10.004. [DOI] [PubMed] [Google Scholar]
- Guerrero-Hernandez A, Leon-Aparicio D, Chavez-Reyes J, Olivares-Reyes JA, DeJesus S. Endoplasmic reticulum stress in insulin resistance and diabetes. Cell Calcium. 2014;56:311–322. doi: 10.1016/j.ceca.2014.08.006. [DOI] [PubMed] [Google Scholar]
- Guo FJ, Xiong Z, Lu X, Ye M, Han X, Jiang R. ATF6 upregulates XBP1S and inhibits ER stress-mediated apoptosis in osteoarthritis cartilage. Cell Signal. 2014;26:332–342. doi: 10.1016/j.cellsig.2013.11.018. [DOI] [PubMed] [Google Scholar]
- Han D, Lerner AG, Vande Walle L, Upton JP, Xu W, Hagen A, Backes BJ, Oakes SA, Papa FR. IRE1alpha kinase activation modes control alternate endoribonuclease outputs to determine divergent cell fates. Cell. 2009;138:562–575. doi: 10.1016/j.cell.2009.07.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harada M, Nose E, Takahashi N, Hirota Y, Hirata T, Yoshino O, Koga K, Fujii T, Osuga Y. Evidence of the activation of unfolded protein response in granulosa and cumulus cells during follicular growth and maturation. Gynecol Endocrinol. 2015;31:783–787. doi: 10.3109/09513590.2015.1062862. [DOI] [PubMed] [Google Scholar]
- Harding HP, Novoa I, Zhang Y, Zeng H, Wek R, Schapira M, Ron D. Regulated translation initiation controls stress-induced gene expression in mammalian cells. Mol Cell. 2000;6:1099–1108. doi: 10.1016/s1097-2765(00)00108-8. [DOI] [PubMed] [Google Scholar]
- Haywood J, Yammani RR. Free fatty acid palmitate activates unfolded protein response pathway and promotes apoptosis in meniscus cells. Osteoarthr Cartil. 2015;24:942. doi: 10.1016/j.joca.2015.11.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haze K, Yoshida H, Yanagi H, Yura T, Mori K. Mammalian transcription factor ATF6 is synthesized as a transmembrane protein and activated by proteolysis in response to endoplasmic reticulum stress. Mol Biol Cell. 1999;10:3787–3799. doi: 10.1091/mbc.10.11.3787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hou YJ, Zhu CC, Duan X, Liu HL, Wang Q, Sun SC. Both diet and gene mutation induced obesity affect oocyte quality in mice. Sci Rep. 2016;6:18858. doi: 10.1038/srep18858. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hsueh AJ, Kawamura K, Cheng Y, Fauser BC. Intraovarian control of early folliculogenesis. Endocr Rev. 2015;36:1–24. doi: 10.1210/er.2014-1020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu R, Chen ZF, Yan J, Li QF, Huang Y, Xu H, Zhang XP, Jiang H. Endoplasmic reticulum stress of neutrophils is required for ischemia/reperfusion-induced acute lung injury. J Immunol. 2015;195:4802–4809. doi: 10.4049/jimmunol.1500073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iurlaro R, Munoz-Pinedo C. Cell death induced by endoplasmic reticulum stress. Febs J. 2016;283:2640–2652. doi: 10.1111/febs.13598. [DOI] [PubMed] [Google Scholar]
- Korennykh AV, Egea PF, Korostelev AA, Finer-Moore J, Zhang C, Shokat KM, Stroud RM, Walter P. The unfolded protein response signals through high-order assembly of Ire1. Nature. 2008;457(7230):687–693. doi: 10.1038/nature07661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kosaka K, Fujiwara H, Yoshioka S, Fujii S. Vascular endothelial growth factor production by circulating immune cells is elevated in ovarian hyperstimulation syndrome. Hum Reprod. 2007;22:1647–1651. doi: 10.1093/humrep/dem042. [DOI] [PubMed] [Google Scholar]
- Krebs J, Agellon LB, Michalak M. Ca(2 +) homeostasis and endoplasmic reticulum (ER) stress: an integrated view of calcium signaling. Biochem Biophys Res Commun. 2015;460:114–121. doi: 10.1016/j.bbrc.2015.02.004. [DOI] [PubMed] [Google Scholar]
- Leamy AK, Egnatchik RA, Young JD. Molecular mechanisms and the role of saturated fatty acids in the progression of non-alcoholic fatty liver disease. Prog Lipid Res. 2013;52:165–174. doi: 10.1016/j.plipres.2012.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee AS. The ER chaperone and signaling regulator GRP78/BiP as a monitor of endoplasmic reticulum stress. Methods. 2005;35:373–381. doi: 10.1016/j.ymeth.2004.10.010. [DOI] [PubMed] [Google Scholar]
- Li Y, Ge M, Ciani L, Kuriakose G, Westover EJ, Dura M, Covey DF, Freed JH, Maxfield FR, Lytton J, Tabas I. Enrichment of endoplasmic reticulum with cholesterol inhibits sarcoplasmic-endoplasmic reticulum calcium ATPase-2b activity in parallel with increased order of membrane lipids: implications for depletion of endoplasmic reticulum calcium stores and apoptosis in cholesterol-loaded macrophages. J Biol Chem. 2004;279:37030–37039. doi: 10.1074/jbc.M405195200. [DOI] [PubMed] [Google Scholar]
- Li S, Guo L, Qian P, Zhao Y, Liu A, Ji F, Chen L, Wu X, Qian G. Lipopolysaccharide induces autophagic cell death through the perk-dependent branch of the unfolded protein response in human alveolar epithelial A549 cells. Cell Physiol Biochem. 2015;36:2403–2417. doi: 10.1159/000430202. [DOI] [PubMed] [Google Scholar]
- Lin P, Yang Y, Li X, Chen F, Cui C, Hu L, Li Q, Liu W, Jin Y. Endoplasmic reticulum stress is involved in granulosa cell apoptosis during follicular atresia in goat ovaries. Mol Reprod Dev. 2012;79:423–432. doi: 10.1002/mrd.22045. [DOI] [PubMed] [Google Scholar]
- Lu M, Lawrence DA, Marsters S, Acosta-Alvear D, Kimmig P, Mendez AS, Paton AW, Paton JC, Walter P, Ashkenazi A. Opposing unfolded-protein-response signals converge on death receptor 5 to control apoptosis. Science. 2014;345(6192):98–101. doi: 10.1126/science.1254312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luo S, Mao C, Lee B, Lee AS. GRP78/BiP is required for cell proliferation and protecting the inner cell mass from apoptosis during early mouse embryonic development. Mol Cell Biol. 2006;26:5688–5697. doi: 10.1128/MCB.00779-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma Y, Brewer JW, Diehl JA, Hendershot LM. Two distinct stress signaling pathways converge upon the CHOP promoter during the mammalian unfolded protein response. J Mol Biol. 2002;318:1351–1365. doi: 10.1016/s0022-2836(02)00234-6. [DOI] [PubMed] [Google Scholar]
- Manabe N, Matsuda-Minehata F, Goto Y, Maeda A, Cheng Y, Nakagawa S, Inoue N, Wongpanit K, Jin H, Gonda H, Li J. Role of cell death ligand and receptor system on regulation of follicular atresia in pig ovaries. Reprod Domest Anim. 2008;43(Suppl 2):268–272. doi: 10.1111/j.1439-0531.2008.01172.x. [DOI] [PubMed] [Google Scholar]
- Marciniak SJ, Yun CY, Oyadomari S, Novoa I, Zhang Y, Jungreis R, Nagata K, Harding HP, Ron D. CHOP induces death by promoting protein synthesis and oxidation in the stressed endoplasmic reticulum. Genes Dev. 2004;18:3066–3077. doi: 10.1101/gad.1250704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marciniak SJ, Garcia-Bonilla L, Hu J, Harding HP, Ron D. ctivation-dependent substrate recruitment by the eukaryotic translation initiation factor 2 kinase PERK. J Cell Biol. 2006;172(2):201–209. doi: 10.1083/jcb.200508099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marjon PL, Bobrovnikova-Marjon EV, Abcouwer SF. Expression of the pro-angiogenic factors vascular endothelial growth factor and interleukin-8/CXCL8 by human breast carcinomas is responsive to nutrient deprivation and endoplasmic reticulum stress. Mol Cancer. 2004;3:4. doi: 10.1186/1476-4598-3-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martinou JC, Youle RJ. Mitochondria in apoptosis: Bcl-2 family members and mitochondrial dynamics. Dev Cell. 2011;21:92–101. doi: 10.1016/j.devcel.2011.06.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCullough KD, Martindale JL, Klotz LO, Aw TY, Holbrook NJ. Gadd153 sensitizes cells to endoplasmic reticulum stress by down-regulating Bcl2 and perturbing the cellular redox state. Mol Cell Biol. 2001;21:1249–1259. doi: 10.1128/MCB.21.4.1249-1259.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McGee EA, Hsueh AJ. Initial and cyclic recruitment of ovarian follicles. Endocr Rev. 2000;21:200–214. doi: 10.1210/edrv.21.2.0394. [DOI] [PubMed] [Google Scholar]
- McLean K, VanDeVen NA, Sorenson DR, Daudi S, Liu JR. The HIV protease inhibitor saquinavir induces endoplasmic reticulum stress, autophagy, and apoptosis in ovarian cancer cells. Gynecol Oncol. 2009;112:623–630. doi: 10.1016/j.ygyno.2008.11.028. [DOI] [PubMed] [Google Scholar]
- Monniaux D, Huet C, Besnard N, Clement F, Bosc M, Pisselet C, Monget P, Mariana JC. Follicular growth and ovarian dynamics in mammals. J Reprod Fertil Suppl. 1997;51:3–23. [PubMed] [Google Scholar]
- Monniaux D, Clement F, Dalbies-Tran R, Estienne A, Fabre S, Mansanet C, Monget P. The ovarian reserve of primordial follicles and the dynamic reserve of antral growing follicles: what is the link? Biol Reprod. 2014;90:85. doi: 10.1095/biolreprod.113.117077. [DOI] [PubMed] [Google Scholar]
- Monniaux D, Michel P, Postel M, Clement F. Multiscale modeling of ovarian follicular development: From follicular morphogenesis to selection for ovulation. Biol Cell. 2016;108:149. doi: 10.1111/boc.201500087. [DOI] [PubMed] [Google Scholar]
- Niswender GD. Molecular control of luteal secretion of progesterone. Reproduction. 2002;123:333–339. doi: 10.1530/rep.0.1230333. [DOI] [PubMed] [Google Scholar]
- Novoa I, Zeng H, Harding HP, Ron D. Feedback inhibition of the unfolded protein response by GADD34-mediated dephosphorylation of eIF2alpha. J Cell Biol. 2001;153:1011–1022. doi: 10.1083/jcb.153.5.1011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oktay K, Briggs D, Gosden RG. Ontogeny of follicle-stimulating hormone receptor gene expression in isolated human ovarian follicles. J Clin Endocrinol Metab. 1997;82:3748–3751. doi: 10.1210/jcem.82.11.4346. [DOI] [PubMed] [Google Scholar]
- Olzmann JA, Kopito RR, Christianson JC. The mammalian endoplasmic reticulum-associated degradation system. Cold Spring Harb Perspect Biol. 2013;5:a013185. doi: 10.1101/cshperspect.a013185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pantasri T, Wu LL, Hull ML, Sullivan TR, Barry M, Norman RJ, Robker RL. Distinct localisation of lipids in the ovarian follicular environment. Reprod Fertil Dev. 2015;27:593. doi: 10.1071/RD14321. [DOI] [PubMed] [Google Scholar]
- Park HJ, Park SJ, Koo DB, Kong IK, Kim MK, Kim JM, Choi MS, Park YH, Kim SU, Chang KT, Park CK, Chae JI, Lee DS. Unfolding protein response signaling is involved in development, maintenance, and regression of the corpus luteum during the bovine estrous cycle. Biochem Biophys Res Commun. 2013;441:344–350. doi: 10.1016/j.bbrc.2013.10.056. [DOI] [PubMed] [Google Scholar]
- Park SJ, Kim TS, Park CK, Lee SH, Kim JM, Lee KS, Lee IK, Park JW, Lawson MA, Lee DS. hCG-induced endoplasmic reticulum stress triggers apoptosis and reduces steroidogenic enzyme expression through activating transcription factor 6 in Leydig cells of the testis. J Mol Endocrinol. 2013;50:151–166. doi: 10.1530/JME-12-0195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park HJ, Park SJ, Koo DB, Lee SR, Kong IK, Ryoo JW, Park YI, Chang KT, Lee DS. Progesterone production is affected by unfolded protein response (UPR) signaling during the luteal phase in mice. Life Sci. 2014;113:60–67. doi: 10.1016/j.lfs.2014.07.033. [DOI] [PubMed] [Google Scholar]
- Rana T, Shinde VM, Starr CR, Kruglov AA, Boitet ER, Kotla P, Zolotukhin S, Gross AK, Gorbatyuk MS. An activated unfolded protein response promotes retinal degeneration and triggers an inflammatory response in the mouse retina. Cell Death Dis. 2014;5:e1578. doi: 10.1038/cddis.2014.539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rekawiecki R, Kowalik MK, Slonina D, Kotwica J. Regulation of progesterone synthesis and action in bovine corpus luteum. J Physiol Pharmacol. 2008;59:74–89. [PubMed] [Google Scholar]
- Robker RL, Akison LK, Bennett BD, Thrupp PN, Chura LR, Russell DL, Lane M, Norman RJ. Obese women exhibit differences in ovarian metabolites, hormones, and gene expression compared with moderate-weight women. J Clin Endocrinol Metab. 2009;94:1533–1540. doi: 10.1210/jc.2008-2648. [DOI] [PubMed] [Google Scholar]
- Ron D, Walter P. Signal integration in the endoplasmic reticulum unfolded protein response. Nat Rev Mol Cell Biol. 2007;8:519–529. doi: 10.1038/nrm2199. [DOI] [PubMed] [Google Scholar]
- Sato Y, Nadanaka S, Okada T, Okawa K, Mori K. Luminal domain of ATF6 alone is sufficient for sencing endoplasmic reticulum stress and subsequent transport to golgi apparatus. Cell Struct Funct. 2011;36:35–47. doi: 10.1247/csf.10010. [DOI] [PubMed] [Google Scholar]
- Scaramuzzi RJ, Baird DT, Campbell BK, Driancourt MA, Dupont J, Fortune JE, Gilchrist RB, Martin GB, McNatty KP, McNeilly AS, Monget P, Monniaux D, Vinoles C, Webb R. Regulation of folliculogenesis and the determination of ovulation rate in ruminants. Reprod Fertil Dev. 2011;23:444–467. doi: 10.1071/RD09161. [DOI] [PubMed] [Google Scholar]
- Scheuner D, Song B, McEwen E, Liu C, Laybutt R, Gillespie P, Saunders T, Bonner-Weir S, Kaufman RJ. Translational control is required for the unfolded protein response and in vivo glucose homeostasis. Mol Cell. 2001;7:1165–1176. doi: 10.1016/s1097-2765(01)00265-9. [DOI] [PubMed] [Google Scholar]
- Sessions-Bresnahan DR, Schauer KL, Heuberger AL, Carnevale EM. Effect of obesity on the preovulatory follicle and lipid fingerprint of equine oocytes. Biol Reprod. 2016;94:15. doi: 10.1095/biolreprod.115.130187. [DOI] [PubMed] [Google Scholar]
- Shen J, Snapp EL, Lippincott-Schwartz J, Prywes R. Stable binding of ATF6 to BiP in the endoplasmic reticulum stress response. Mol Cell Biol. 2005;25:921–932. doi: 10.1128/MCB.25.3.921-932.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shore GC, Papa FR, Oakes SA. Signaling cell death from the endoplasmic reticulum stress response. Curr Opin Cell Biol. 2011;23:143–149. doi: 10.1016/j.ceb.2010.11.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stouffer RL. Progesterone as a mediator of gonadotrophin action in the corpus luteum: beyond steroidogenesis. Hum Reprod Update. 2003;9:99–117. doi: 10.1093/humupd/dmg016. [DOI] [PubMed] [Google Scholar]
- Stouffer RL, Bishop CV, Bogan RL, Xu F, Hennebold JD. Endocrine and local control of the primate corpus luteum. Reprod Biol. 2013;13:259–271. doi: 10.1016/j.repbio.2013.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tabas I, Ron D. Integrating the mechanisms of apoptosis induced by endoplasmic reticulum stress. Nat Cell Biol. 2011;13:184–190. doi: 10.1038/ncb0311-184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takahashi N, Harada M, Hirota Y, Zhao L, Yoshino O, Urata Y, Izumi G, Takamura M, Hirata T, Koga K, Wada-Hiraike O, Fujii T, Osuga Y. A potential role of endoplasmic reticulum stress in development of ovarian hyperstimulation syndrome. Mol Cell Endocrinol. 2016;428:161. doi: 10.1016/j.mce.2016.03.032. [DOI] [PubMed] [Google Scholar]
- Urano F, Wang X, Bertolotti A, Zhang Y, Chung P, Harding HP, Ron D. Coupling of stress in the ER to activation of JNK protein kinases by transmembrane protein kinase IRE1. Science. 2000;287:664–666. doi: 10.1126/science.287.5453.664. [DOI] [PubMed] [Google Scholar]
- Wang Q, Groenendyk J, Michalak M. Glycoprotein quality control and endoplasmic reticulum stress. Molecules. 2015;20:13689–13704. doi: 10.3390/molecules200813689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woo CW, Cui D, Arellano J, Dorweiler B, Harding H, Fitzgerald KA, Ron D, Tabas I. Adaptive suppression of the ATF4-CHOP branch of the unfolded protein response by toll-like receptor signalling. Nat Cell Biol. 2009;11:1473–1480. doi: 10.1038/ncb1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu LL, Dunning KR, Yang X, Russell DL, Lane M, Norman RJ, Robker RL. High-fat diet causes lipotoxicity responses in cumulus-oocyte complexes and decreased fertilization rates. Endocrinology. 2010;151:5438–5445. doi: 10.1210/en.2010-0551. [DOI] [PubMed] [Google Scholar]
- Wu LL, Russell DL, Norman RJ, Robker RL. Endoplasmic reticulum (ER) stress in cumulus-oocyte complexes impairs pentraxin-3 secretion, mitochondrial membrane potential (DeltaPsi m), and embryo development. Mol Endocrinol. 2012;26:562–573. doi: 10.1210/me.2011-1362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamaguchi H, Wang HG. CHOP is involved in endoplasmic reticulum stress-induced apoptosis by enhancing DR5 expression in human carcinoma cells. J Biol Chem. 2004;279:45495–45502. doi: 10.1074/jbc.M406933200. [DOI] [PubMed] [Google Scholar]
- Yang X, Wu LL, Chura LR, Liang X, Lane M, Norman RJ, Robker RL. Exposure to lipid-rich follicular fluid is associated with endoplasmic reticulum stress and impaired oocyte maturation in cumulus-oocyte complexes. Fertil Steril. 2012;97:1438–1443. doi: 10.1016/j.fertnstert.2012.02.034. [DOI] [PubMed] [Google Scholar]
- Ye J, Rawson RB, Komuro R, Chen X, Davé UP, Prywes R, Brown MS, Goldstein JL. ER stress induces cleavage of membrane-bound ATF6 by the same proteases that process SREBPs. Mol Cell. 2000;6:1355–1364. doi: 10.1016/s1097-2765(00)00133-7. [DOI] [PubMed] [Google Scholar]
- Ye J, Kumanova M, Hart LS, Sloane K, Zhang H, De Panis DN, Bobrovnikova-Marjon E, Diehl JA, Ron D, Koumenis C. The GCN2-ATF4 pathway is critical for tumour cell survival and proliferation in response to nutrient deprivation. EMBO J. 2010;29:2082–2096. doi: 10.1038/emboj.2010.81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshida H, Haze K, Yanagi H, Yura T, Mori K. Identification of the cis-acting endoplasmic reticulum stress response element responsible for transcriptional induction of mammalian glucose-regulated proteins. J Biol Chem. 1998;273:33741–33749. doi: 10.1074/jbc.273.50.33741. [DOI] [PubMed] [Google Scholar]
- Yoshida H, Okada T, Haze K, Yanagi H, Yura T, Negishi M, Mori K. ATF6 activated by proteolysis binds in the presence of NF-Y (CBF) directly to the cis-acting element responsible for the mammalian unfolded protein response. Mol Cell Biol. 2000;20:6755–6767. doi: 10.1128/mcb.20.18.6755-6767.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshida H, Matsui T, Yamamoto A, Okada T, Mori K. XBP1 mRNA is induced by ATF6 and spliced by IRE1 in response to ER stress to produce a highly active transcription factor. Cell. 2001;107:881–891. doi: 10.1016/s0092-8674(01)00611-0. [DOI] [PubMed] [Google Scholar]
- Zeng L, Xiao Q, Chen M, Margariti A, Martin D, Ivetic A, Xu H, Mason J, Wang W, Cockerill G, Mori K, Li JY, Chien S, Hu Y, Xu Q. Vascular endothelial cell growth-activated XBP1 splicing in endothelial cells is crucial for angiogenesis. Circulation. 2013;127:1712–1722. doi: 10.1161/CIRCULATIONAHA.112.001337. [DOI] [PubMed] [Google Scholar]
- Zhang X, Jafari N, Barnes RB, Confino E, Milad M, Kazer RR. Studies of gene expression in human cumulus cells indicate pentraxin 3 as a possible marker for oocyte quality. Fertil Steril. 2005;83(Suppl 1):1169–1179. doi: 10.1016/j.fertnstert.2004.11.030. [DOI] [PubMed] [Google Scholar]
- Zhang G, Wan Y, Zhang Y, Lan S, Jia R, Wang Z, Fan Y, Wang F. Expression of mitochondria-associated genes (PPARGC1A, NRF-1, BCL-2 and BAX) in follicular development and atresia of goat ovaries. Reprod Domest Anim. 2015;50:465–473. doi: 10.1111/rda.12514. [DOI] [PubMed] [Google Scholar]
- Zhang LY, Li PL, Xu A, Zhang XC. Involvement of GRP78 in the resistance of ovarian carcinoma cells to paclitaxel. Asian Pac J Cancer Prev. 2015;16:3517–3522. doi: 10.7314/apjcp.2015.16.8.3517. [DOI] [PubMed] [Google Scholar]
- Zhou Y, Sun P, Wang T, Chen K, Zhu W, Wang H. Inhibition of calcium influx reduces dysfunction and apoptosis in lipotoxic pancreatic beta-cells via regulation of endoplasmic reticulum stress. PLoS One. 2015;10:e0132411. doi: 10.1371/journal.pone.0132411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zong WX, Li C, Hatzivassiliou G, Lindsten T, Yu QC, Yuan J, Thompson CB. Bax and Bak can localize to the endoplasmic reticulum to initiate apoptosis. J Cell Biol. 2003;162:59–69. doi: 10.1083/jcb.200302084. [DOI] [PMC free article] [PubMed] [Google Scholar]