

Abstract
Pathogen replication and transmission in Wolbachia infected insects are currently studied using three Wolbachia infection systems: naturally infected Wolbachia hosts, hosts transinfected with Wolbachia (stably maintained and inherited infections) and hosts transiently infected with Wolbachia. All three systems have been used to test the effect of Wolbachia on mosquito transmitted pathogens such as dengue virus (DENV), West Nile virus (WNV) and Plasmodium. From these studies it is becoming increasingly clear that the interaction between a particular pathogen and Wolbachia is heavily influenced by the host-Wolbachia interaction and the model of infection. In particular, there is some evidence that under very specific conditions, Wolbachia can enhance pathogen infection in some hosts. In this study, we compared the effect of Wolbachia in two infection models (stable transinfected and transiently infected) on the replication, infection- and transmission rates of two flaviviruses, DENV and WNV (Kunjin strain). Our results indicate that Wolbachia had similar blocking effects in both stable and transient models of infection, however, the magnitude of the blocking effect was significantly lower in mosquitoes transiently infected with Wolbachia. More importantly, no evidence was found for any enhancement of either DENV or WNV (Kunjin strain) infection in Ae. aegypti infected with Wolbachia, supporting a role for Wolbachia as an effective and safe means for restricting transmission of these viruses.
Author Summary
Wolbachia is a naturally occurring endosymbiotic bacterium that, when introduced into a naïve mosquito host, has been shown to effectively reduce the replication and transmission of pathogens such as dengue virus, West Nile virus, Chikungunya virus, yellow fever virus and Plasmodium. However, a recent study has indicated that, under certain conditions, transiently infected Wolbachia can enhance West Nile virus infection in Culex tarsalis mosquitoes. We wanted to investigate whether this enhancement effect could also be observed in Aedes aegypti mosquitoes and if so, whether it is specific to the nature of the Wolbachia infection model under study (transient vs stable). We compared the replication and transmission of dengue virus and WNV (Kunjin strain) in Aedes aegypti mosquitoes transiently infected with Wolbachia and mosquitoes stably infected with the identical Wolbachia strain. Contrary to the previous study, our results show no enhancement of replication or transmission for either dengue virus or WNV (Kunjin strain) in mosquitoes transiently or stably infected with Wolbachia.
Introduction
The potential for Wolbachia as a natural control method for mosquito-borne pathogens such as dengue virus (DENV), Chikungunya virus (CHIKV), West Nile virus (WNV), yellow fever virus (YFV) and malaria has been the focus of intense study in recent years [1–8]. The majority of these studies have focussed on DENV replication and have shown conclusively that Wolbachia effectively reduces DENV replication and transmission when introduced as a stable infection in the naturally uninfected host Aedes aegypti [8–11].
To date, only a few mosquito species have been successfully transinfected with Wolbachia. These include Ae. aegypti (transinfected with the Wolbachia strains wMel, wMelPop, wAlbB, and superinfection with wMelwAlbB [6, 8–10]), Ae. albopictus (cured of its natural Wolbachia infection and transinfected with the wMel Wolbachia strain [12]), and Ae. polynesienses and Anopheles stephensi (both transinfected with the wAlbB Wolbachia strain [13, 14]). These transinfected strains have shown excellent potential for the biocontrol of several important mosquito-transmitted diseases (for recent reviews see [15–17]). However, several disease transmitting mosquito species remain recalcitrant to Wolbachia transinfection, hampering efforts to better understand the interaction between Wolbachia, it’s host and disease causing pathogens [18].
Natural Wolbachia infection models have therefore also been examined to provide insight into Wolbachia-host-pathogen interactions. In this model, the naturally occurring Wolbachia infection is first cured from the host and pathogen replication is subsequently compared in cured and naturally infected hosts [19, 20].
Using this model Baton et al. found that wFlu infection in its natural host Ae. fluviatilis, enhanced oocyst infection with the avian malaria parasite P. gallinaceum [19, 20]. Zele et al. also showed that in the natural mosquito-Wolbachia-Plasmodium combination, Wolbachia increased the susceptibility of Culex pipiens mosquitoes to P. relictum [20]. Furthermore, a study by Mousson et al. using this model, found that Ae. albopictus naturally superinfected with the two Wolbachia strains (wAlbA and wAlbB) infection limited the transmission, but not replication of DENV. Here, both the naturally occurring Wolbachia strains were cured and the vector competence for DENV of the resulting uninfected line was compared to the superinfected line [21].
In addition to natural infection systems, transient infection systems have been used to investigate the effect of Wolbachia on Plasmodium and WNV infection. Here, Wolbachia is injected into an uninfected mosquito host and allowed to establish a transient somatic infection [22]. Using this model, the effect of the Wolbachia strains wAlbB and wMelPop on the malaria parasite P. berghei in A. gambiae was investigated [23]. Contrary to the results of [14], in this experimental setup, wAlbB was found to enhance P. berghei infection, whilst wMelPop only had a moderate blocking effect [23]. A more recent study utilised the same infection model to investigate the effect of the wAlbB Wolbachia strain on WNV infection in Culex tarsalis [24]. Contrary to previous studies that found wAlbB inhibited WNV infection in Ae. aegypti [5], C. tarsalis transiently infected with wAlbB enhanced WNV infection rates at 7 days post infection [24].
Together these results suggest that the degree of pathogen modulation from different host-Wolbachia combinations can differ considerably depending on the mode of infection, the host and the pathogen. Consequently, it is important not to base predictions of pathogen modulation in a particular host-Wolbachia strain combination on results obtained from divergent infection modes and host species. In this study we have compared the effect of wAlbB on replication and transmission of DENV and WNV (Kunjin strain) in Ae. aegypti infected through both transient somatic infection and stable transinfection. Our results showed significantly lower Wolbachia infection densities in transiently infected Ae. aegypti when compared to the stable infected line. More importantly, both Wolbachia infection models displayed similar effects, blocking replication and transmission of both DENV and WNV (Kunjin strain). These results conclusively show that neither DENV nor WNV (Kunjin strain) infection is enhanced in Ae. aegypti either transiently or stably infected with wAlbB.
Results and Discussion
Wolbachia density and distribution in transiently infected Ae. aegypti mosquitoes
Wolbachia density and distribution was analysed in female Ae. aegypti mosquitoes transiently infected with the wAlbB Wolbachia strain and compared to the stable infected wAlbB line. Wolbachia density was determined using qPCR and primers specific to the Wolbachia surface protein (wsp) in conjunction with the Ae. aegypti actin gene for normalisation. In our experiments, even when Wolbachia was injected at very high densities (~1011 bacteria/mL), there were significantly lower (Mann-Whitney test, p = 0.007) Wolbachia densities at 7 days post injection (dpi) in the transiently infected mosquitoes than densities observed in the stable wAlbB infected line (Fig 1A).
Fig 1. Wolbachia density and localisation in transiently infected Aedes aegypti 7 days post injection (dpi).
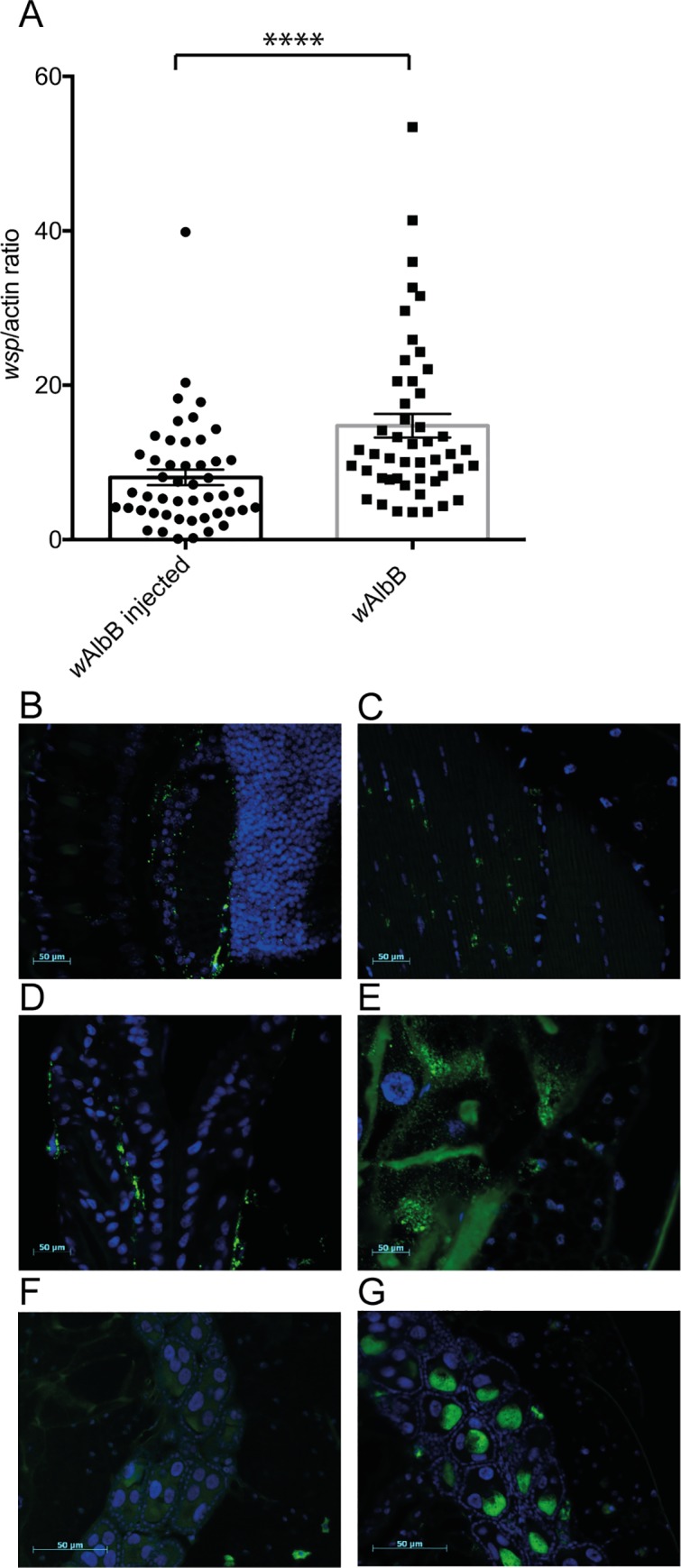
A) The wsp/actin ratio in transiently infected Ae. aegypti compared to the stable transinfected strain. The combined results from two independent experiments are shown. Wolbachia density was determined using qPCR as described and the mean and error of the mean are indicated. Statistical significance was determined using a Mann-Whitney test (****, p < 0.0001). B-F) FISH staining of wAlbB (green) in transiently infected female mosquitoes. B) brain tissue, C) muscle tissue, D) midgut tissue, E) fat body, F) ovaries. G) wAlbB localisation in ovaries of the stable, transinfected line.
Wolbachia in transiently infected mosquitoes were predominantly located in the brain (Fig 1B), muscle tissue (Fig 1C), the midgut (Fig 1D) and the fat body (Fig 1E). In stark contrast to the stable wAlbB infected line, however, very little to no Wolbachia could be detected in the ovaries of transiently infected lines (Fig 1F & 1G). In the stable wAlbB infected line, the vast majority of Wolbachia are found in the ovaries and the lack of Wolbachia found in the ovaries of transiently infected mosquitoes could explain the significant differences found in Wolbachia density between stable and transiently infected mosquitoes in our qPCR results (Fig 1A). These results are also consistent with previous studies that showed only limited Wolbachia localisation in the ovaries of transiently infected Culex tarsalis [24] and comparatively low levels of Wolbachia in the ovaries of transiently infected Anopheles gambiae compared to the rest of the body [25].
DENV replication and transmission is reduced in female Ae. aegypti transiently infected with wAlbB
We next investigated whether female Ae. aegypti mosquitoes transiently infected with wAlbB displayed the same DENV blocking phenotype as the stable infected wAlbB line [24]. Townsville wild type (W.T.), W.T. transiently infected with wAlbB and stable wAlbB infected females were provided with a DENV infected blood meal 7 dpi. The mosquitoes were incubated for a further 7 days as described in materials and methods and subsequently analysed for DENV replication (Fig 2A), DENV infection rate (Fig 2B), DENV transmission rate (Fig 2C), as well as wAlbB density (Fig 2D).
Fig 2. DENV infection, replication and transmission in wild type (W.T.), transiently infected (wAlbB injected) and the stable transinfected (wAlbB) lines 7 days post an infectious blood meal.

The combined results of two independent experiments are shown (the results of each individual repeat are available as supplemental data in S2 Fig and S4 Fig). Statistical significance was determined using a Mann-Whitney test (A and D) or a Fisher exact test (B and C). In A and D the mean and error of the mean is indicated. In B and C, the error bars represent 95% confidence levels. A) DENV genome copies in whole mosquito bodies (***, p < 0.001; **, p = 0.004; Mann-Whitney). B) DENV infection rate as determined by the percentage of individuals infected 7 days post an infectious blood meal, n = 28 (***, p = 0.001; **, p = 0.004; *, p = 0.04; Fisher exact test). C) DENV transmission rate as determined by the percentage of infectious saliva expectorated 7 days post an infectious blood meal, n = 28 (***, p < 0.0004; *, p = 0.03; Fisher exact test). D) Wolbachia density 7 days post an infectious blood meal in transiently infected and the stable transinfected line (****, p < 0.0001; Mann-Whitney).
DENV copy number (as determined by positive strand genome copy number) in the bodies of transiently infected females was significantly reduced (Mann-Whitney, p = 0.0002) by ~1.5 logs when compared to DENV replication in W.T. mosquitoes. Ae. aegypti stably infected with wAlbB showed the greatest reduction in + strand DENV genome copies with a ~ 3 log reduction compared to W.T. (Mann-Whitney, p = 0.001) and ~ 2 log reduction compared to transiently infected females (Mann-Whitney, p = 0.004). Similarly, the DENV infection rate was significantly reduced (~2 fold, Fisher exact test, p = 0.004) in Ae. aegypti females transiently infected with wAlbB when compared with W.T. females (Fig 2B). Ae. aegypti stably infected with wAlbB again showed the greatest reduction in DENV transmission rates with an ~8 fold reduction compared to W.T. females (Fisher exact test, p = 0.0001) and ~4 fold reduction compared to Ae. aegypti females transiently infected with wAlbB (Fisher exact test, p = 0.04).
DENV transmission in transiently infected wAlbB mosquitoes was significantly reduced compared to W.T. mosquitoes. Saliva was collected 7 days post feeding from females fed with an infected blood meal and then injected into DENV-naïve W.T. females according to [26]. The mosquitoes were incubated for an additional 7 days before analysing DENV infection status by qRT-PCR (Fig 2C). No DENV infectious saliva was detected from female Ae. aegypti mosquitoes stably infected with wAlbB. In contrast, 22% of female Ae. aegypti mosquitoes transiently infected with wAlbB expectorated DENV infectious saliva.
Finally, we analysed the Wolbachia density in the same transiently and stably infected mosquitoes analysed for DENV replication and transmission. Our results indicate much lower Wolbachia densities in the transiently infected females compared to the stable wAlbB infected line (Fig 2D). Wolbachia density has been correlated with the degree of pathogen blocking in Wolbachia infected hosts [27] and the lower densities in transiently infected mosquitoes observed here provides a plausible explanation for the reduced DENV blocking phenotype we observed in the transiently infected mosquitoes compared to the stable infected line.
WNV (Kunjin strain) replication is reduced in female Ae. aegypti transiently infected with wAlbB
We repeated the infection experiments using WNV (Kunjin strain). W.T. and wAlbB infected females were provided with a virus infected blood meal at 7 dpi. The mosquitoes were incubated for a further 7 days as described and virus titres were determined in whole bodies (Fig 3A) and saliva (Fig 3B). We also compared the virus infection (Fig 3C) and transmission rates (Fig 3D) between Wolbachia infected and uninfected mosquitoes.
Fig 3. WNV (Kunjin strain) infection, replication and transmission in wild type (W.T.), transiently infected (wAlbB injected) and the stable transinfected (wAlbB) lines 7 days post an infectious blood meal.

The combined results of two independent experiments are shown (the results of each individual repeat are available as supplemental data in S3 Fig and S5 Fig). Statistical significance was determined using a Mann-Whitney test (A, B and E) or a Fisher exact test (C and D). In A, B and E, the mean and error of the mean is indicated. In C and D, the error bars represent 95% confidence levels. A) WNV (Kunjin strain) PFU per ml in whole mosquito bodies, (****, p < 0.0001; ***, p = 0.0003, **, p = 0.003; Mann-Whitney). B) WNV (Kunjin strain) PFU per ml in saliva (****, p < 0.0001; **, p = 0.001; *, p = 0.01; Mann-Whitney). C) WNV (Kunjin strain) infection rate as determined by the percentage of individuals infected 7 days post an infectious blood meal, n = 32 (***, p = 0.001; Fisher exact test). D) WNV (Kunjin strain) transmission rate as determined by the percentage of infectious saliva expectorated infected 7 days post an infectious blood meal, n = 32 (*, p = 0.03; Fisher exact test). E) Wolbachia density 7 days post an infectious blood meal in transiently infected and the stable transinfected lines (****, p < 0.0001; Mann-Whitney).
Virus titres were significantly reduced in the bodies of both the transient and stable infected mosquito lines compared to W.T. females. Transient Wolbachia infection resulted in an approximate one log reduction (Mann-Whitney, p > 0.0001) in virus PFU (Fig 3A) in whole mosquito bodies. A small but significant (Mann-Whitney, p = 0.001), 0.28 log reduction in virus PFU was observed in saliva from these mosquitoes compared to W.T. (Fig 3B). The greatest reduction in virus PFU was observed in the stable infected mosquito line with a more than 1.5 log reduction in wAlbB-infected mosquito’s bodies (Fig 3A). No infectious virus was detected in saliva from these mosquitoes (Fig 3B).
We observed a small (less than two-fold), non-significant (Fisher exact test, p = 0.13) reduction of virus infection rates between transiently infected and W.T. mosquitoes (Fig 3C). Similarly, a small (less than two-fold), non-significant (Fisher exact test, p = 0.1) reduction in the percentage infectious saliva was found between transiently infected and W.T. mosquitoes (Fig 3D). A significant reduction (Fisher exact test, p = 0.001) in infection rates was observed between the stable wAlbB infected mosquito line and W.T. mosquitoes (Fig 3C).
As with the DENV infected mosquitoes, we analysed the Wolbachia density in the same transiently and stably infected mosquitoes analysed for WNV (Kunjin strain) replication and transmission. Similar to the DENV infected mosquitoes, our results indicate much lower Wolbachia densities in the transiently infected females compared to the stable wAlbB infected line (Fig 3E).
Conclusions
Wolbachia, when stably transinfected into mosquito hosts, has been shown to inhibit a range of pathogens, in particular DENV, CHIKV, WNV, YFV and Plasmodium [1–8]. There are however, a few studies that have demonstrated infection with Wolbachia can lead to enhanced pathogen replication [23, 24, 28, 29]. In particular, a study by Dodson et al. showed that when wAlbB transiently infects C. tarsalis, WNV infection rates can be enhanced [24]. These results are in contrast to a previous study that showed two stable transinfected Ae. aegypti lines (wMel and wMelPop) both inhibited WNV transmission [5].
This would suggest that the interaction between Wolbachia and a particular pathogen is highly dependent on either the infection model, the Wolbachia strain or the host background. To determine whether the results obtained by Dodson et al. [24] could be a result of the Wolbachia infection model, we compared DENV and WNV (Kunjin strain) infection in both Ae. aegypti transiently infected with wAlbB as well as Ae. aegypti stably transinfected with wAlbB. In our experimental setup, DENV and WNV (Kunjin strain) replication was significantly reduced in both Wolbachia infection models. In addition, DENV infection rate and transmission rate was significantly reduced in both models. We also observed a small, but not significant reduction in WNV (Kunjin strain) infection and transmission rates in transiently infected mosquitoes.
These observations differ markedly from those described by Dodson et al. [24] and suggest that in Ae. aegypti mosquitoes, transient and stable Wolbachia infections have similar pathogen modulation effects. In Ae. aegypti, unlike the observations in C. tarsalis, transient infection with wAlbB led to lower virus transmission rates in transiently infected mosquitoes compared to Wolbachia naive wild type mosquitoes. We also observed decreased pathogen blocking in transient Wolbachia infections compared to stable Wolbachia infections. Results generated through the use of transient Wolbachia infection models should therefore be interpreted with caution, as they could potentially underestimate the degree of pathogen blocking compared to the stably infected systems typically used for field disease control programs. Most importantly, our results conclusively show no enhancement of either DENV or WNV (Kunjin strain) infection in Wolbachia infected Ae. aegypti.
Materials and Methods
Ethics statement
Blood feeding by volunteers (Monash University human ethics permit no CF11/0766-2011000387) for this study was approved by the Monash University Human Research Ethics Committee (MUHREC). All adult volunteers provided informed written consent; no child participants were involved in the study.
Experimental design
The experimental design for this study is summarised in S1 Fig. We compared the replication and transmission of DENV and WNV (Kunjin strain) in Townsville wild type (W.T.) mosquitoes, W.T. mosquitoes injected with wAlbB and the stable wAlbB line described in [9]. To generate transient Wolbachia infections in female W.T. mosquitoes, Wolbachia was isolated from 200 wAlbB-infected ovaries and injected into 100 W.T. mosquitoes. For the W.T. and stable wAlbB controls, an extraction was done from 200 W.T. ovaries in the same fashion as the Wolbachia extraction.
This extract was used to inject 50 W.T. females and 50 wAlbB stable infected females. The injected females were incubated for 7 days as described and subsequently allowed to feed on virus infected blood. Fed females were separated from unfed females 24 h post feeding. Females that showed no evidence of feeding were used to analyse the Wolbachia infection 7 days post injection, using qPCR and FISH. Engorged females were incubated for a further 7 days. Seven days post feeding, saliva and carcasses (legs and wings were removed) were collected from all fed mosquitoes and assayed for Wolbachia density, DENV and WNV (Kunjin strain).
Mosquito colonies and lines
Wolbachia-uninfected Ae. aegypti eggs were collected from Townsville (Queensland, Australia) in 2015. The Wolbachia-infected wAlbB mosquito line has been described previously [9] and was a gift from Prof Zhiyong Xi (Michigan State University). All Ae. aegypti mosquitoes were reared and maintained as described in [6] with the following modification. For hatching, eggs were placed in hatching water (distilled H2O, boiled and supplemented with 50 mg/L fish food [Tetramin]) and allowed to hatch for 24 h. Larvae were subsequently reared at a set density of ~150 in 3 L of distilled water as described in [6]. wAlbB infected females were backcrossed for 2 generations with W.T. males prior to infection experiments.
Wolbachia isolation for transient infection
Wolbachia was isolated from the ovaries of wAlbB-infected females according to [24] with the following modifications. Ovaries from 200 wAlbB-infected females were dissected on ice and suspended in 50 μL of ice-cold Schneiders media (Sigma-Aldrich) in a 1.5 mL eppendorf tube. The ovaries were crushed briefly using a small plastic pestle after which one 3 mm glass bead was added and the suspension vortexed for 2 min. One mL of ice-cold Schneiders media was added to the homogenised and the solution were centrifuged at 4°C for 5 min at 2000 x g. The supernatant was subsequently sequentially filtered through 5 μM and 1.2 μM syringe filters. The resulting filtrate was centrifuged for 4°C for 10 min at 12000 x g. The supernatant was discarded and the pellet resuspended in 50 μL of ice-cold Schneiders media until use. The extraction was repeated with ovaries from W.T. females for use in control injections. Total bacterial counts were estimated using the LIVE/DEAD staining kit (Thermofisher) and counting the live stained bacteria in a hemocytometer.
Generation of transiently Wolbachia-infected Ae. aegypti
Female mosquitoes were injected intrathoracically with ~ 1 μL of Wolbachia suspension solution (~1011 bacteria/ml in Schneiders media) using a pulled glass capillary and a handheld microinjector (Nanoject II, Drummond Sci.). Injected mosquitoes were incubated for 7 days (40 mosquitoes per cup) at 26°C with 65% relative humidity and a 12h light/dark cycle. All injection experiments were conducted in duplicate.
Wolbachia density and distribution
Wolbachia density and distribution in the transient infected mosquitoes were compared 7 and 14 days post injection (dpi) to the wAlbB line using qPCR and fluorescence in situ hybridisation (FISH). DNA was extracted from stable and transiently Wolbachia infected mosquitoes using the DNeasy 96 Blood & Tissue kit (Qiagen) according to the manufacturer’s specifications. Quantitative PCR to determine the total relative Wolbachia densities of infected lines was performed as described by [30] using primers specific to the gene coding for the Wolbachia surface protein (wsp) (forward primer 5’-GCATTTGGTTAYAAAATGGACGA-3’, reverse primer 5’- GGAGTGATAGGCATATCTTCAAT-3’), as well as the Ae. aegypti actin gene (forward primer 5’- GACTACCTGATGAAGATCCTGAC-3’, reverse primer: 5’- GCACAGCTTCTCCTTAATGTCAC-3’) [24]. Statistical differences were determined using a Mann-Whitney (Graphpad Prism version 6.0f).
Wolbachia was localized in sections of paraffin-embedded 5–7 day old female mosquitoes by FISH, as described in [31], except that only one probe against 16S rRNA was used and its concentration was increased 10-fold to improve the signal. wAlbB was detected using AlbBW5: 5’-CTTAGGCTTGCGCACCTTGCAA-3’, labelled with Alexa 488 dye (green). DAPI was used to stain total DNA.
Mosquito infection with DENV and the WNV (Kunjin strain)
The propagation and maintenance of dengue virus serotype 2 (DENV-2) ET300 [32] was carried out as previously described [33]. WNV (Kunjin strain) was obtained as a gift from Prof Jason Mackenzie (Melbourne University). WNV (Kunjin strain) was propagated on C6/36 cells in a fashion similar to DENV-2.
Mosquitoes were infected with either DENV-2 (ET300) or WNV (Kunjin strain) (virus strains were grown fresh for each infection) through an infectious blood meal. For feeding experiments with virus infected blood, Ae. aegypti female mosquitoes were placed in 500 mL plastic containers (40/container), starved for 24 hours and allowed to feed on a 50:50 mixture of defibrinated sheep blood and tissue culture supernatant containing 107 genome copies/mL of DENV-2 or 108 pfu/mL of WNV (Kunjin strain). Feeding was done through a piece of desalted porcine intestine stretched over a water-jacketed membrane feeding apparatus preheated to 37°C for approximately three hours. Fully engorged mosquitoes were placed in 500 mL containers and incubated for 7 days at 26°C with 65% relative humidity and a 12h light/dark cycle. All infection experiments were conducted in duplicate.
Virus isolation and quantification from infected mosquitoes
Saliva from infected mosquitoes was collected 7 days post feeding (dpf) as described by [26]. Following saliva collection, the bodies of infected mosquitoes were collected in 100 μL serum free RPMI media (Sigma-Aldrich) and stored at -80°C until processing. For DENV-2, the collected saliva was re-injected into 3-day-old W.T. female mosquitoes as described by [26]. Female mosquitoes injected with saliva were incubated for 7 days at 26°C with 65% relative humidity and a 12h light/dark cycle after which they were collected in RPMI media as above. DENV-2 genome copies were subsequently determined in the blood fed and saliva injected mosquitoes using qRT-PCR as described.
For WNV (Kunjin strain), the mosquitoes and saliva were collected as described above. After collection the mosquito bodies were homogenised in a bead beater at 30 beats/min for 3 min using one 3 mm sterile glass bead. The suspension was briefly centrifuged at 2000 x g and 10 μL of the supernatant was used in plaque assays as described by [24]. Collected saliva was used directly in plaque assays.
To quantify DENV-2 genomic copies, total RNA was isolated from DENV-2 injected mosquitoes using the Nucleospin 96 RNA kit (Macherey-Nagel). DENV-2 qPCR analysis was done using cDNA prepared from individual mosquitoes according to [31] using forward primer 5’-AAGGACTAGAGGTTAGAGGAGACCC-3’ and reverse primer 5’-CGTTCTGTGCCTGGAATGATG-3’. Infectious virus titre of WNV (Kunjin strain) was quantified using plaque assays as described by [24].
Supporting Information
Abbreviations used: BF–Blood fed; dpi–days post injection; dpf–days post feeding; W.T.–wild type Aedes aegypti females.
(TIF)
Statistical significance was determined using a Mann-Whitney test (A and D) or a Fisher exact test (B and C). In A and D the mean and error of the mean is indicated. In B and C, the error bars represent 95% confidence levels. A) DENV genome copies in whole mosquito bodies (****, p < 0.0001; **, p = 0.004; Mann-Whitney). B) DENV infection rate as determined by the percentage of individuals infected 7 days post an infectious blood meal (****, p = 0.001; *, p < 0.05; Fisher exact test). C) DENV transmission rate as determined by the percentage of infectious saliva expectorated 7 days post an infectious blood meal, (***, p = 0.0002; *, p = 0.03; Fisher exact test). D) Wolbachia density 7 days post an infectious blood meal in transiently infected and the stable transinfected line (****, p < 0.0001; Mann-Whitney).
(TIF)
Statistical significance was determined using a Mann-Whitney test (A, B and E) or a Fisher exact test (C and D). In A, B and E, the mean and error of the mean is indicated. In C and D, the error bars represent 95% confidence levels. A) WNV (Kunjin strain) PFU per ml in whole mosquito bodies, (**, p < 0.01; Mann-Whitney). B) WNV (Kunjin strain) PFU per ml in saliva (**, p = 0.002; Mann-Whitney). C) WNV (Kunjin strain) infection rate as determined by the percentage of individuals infected 7 days post an infectious blood meal, (*, p = 0.02; Fisher exact test). D) WNV (Kunjin strain) transmission rate as determined by the percentage of infectious saliva expectorated infected 7 days post an infectious blood meal (**, p = 0.002, Mann-Whitney). E) Wolbachia density 7 days post an infectious blood meal in transiently infected and the stable transinfected lines (****, p < 0.0001; Mann-Whitney).
(TIF)
Statistical significance was determined using a Mann-Whitney test (A and D) or a Fisher exact test (B and C). In A and D, the mean and error of the mean is indicated. In B and C, the error bars represent 95% confidence levels. A) DENV genome copies in whole mosquito bodies (***, p ≤ 0.001; **, p = 0.004; Mann-Whitney). B) DENV infection rate as determined by the percentage of individuals infected 7 days post an infectious blood meal, (****, p = 0.0003; Fisher exact test). C) DENV transmission rate as determined by the percentage of infectious saliva expectorated 7 days post an infectious blood meal, (****, p = 0.0002; *, p = 0.03; Fisher exact test). D) Wolbachia density 7 days post an infectious blood meal in transiently infected and the stable transinfected line (****, p < 0.0001; Mann-Whitney).
(TIF)
Statistical significance was determined using a Mann-Whitney test (A, B and E) or a Fisher exact test (C and D). In A, B and E, the mean and error of the mean is indicated. In C and D, the error bars represent 95% confidence levels. A) WNV (Kunjin strain) PFU per ml in whole mosquito bodies, (**, p < 0.006; *, p = 0.04; Mann-Whitney). B) WNV (Kunjin strain) PFU per ml in saliva (****, p < 0.0001, **, p < 0.009; Mann-Whitney). C) WNV (Kunjin strain) infection rate as determined by the percentage of individuals infected 7 days post an infectious blood meal, (**, p = 0.002; Fisher exact test). D) WNV (Kunjin strain) transmission rate as determined by the percentage of infectious saliva expectorated infected 7 days post an infectious blood meal (**, p = 0.007; Mann-Whitney). E) Wolbachia density 7 days post an infectious blood meal in transiently infected and the stable transinfected lines (****, p < 0.0001; Mann-Whitney).
(TIF)
Acknowledgments
The authors would like to acknowledge Rhiannon Burns for excellent technical assistance, Prof Jason Mackenzie for supplying the WNV (Kunjin strain) isolate used in the study and Prof Zhiyong Xi for supplying the wAlbB Ae. aegypti line.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was support by grants to SLO from the Foundation for the National Institutes of Health through the Grand Challenges in Global Health Initiative of the Bill and Melinda Gates Foundation (http://www.gatesfoundation.org/) and the National Health & Medical Research Council of Australia (https://www.nhmrc.gov.au/) programme grant 1037003. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Bourtzis K, Dobson SL, Xi Z, Rasgon JL, Calvitti M, Moreira LA, et al. Harnessing mosquito-Wolbachia symbiosis for vector and disease control. Acta tropica. 2014;132 Suppl:S150–63. [DOI] [PubMed] [Google Scholar]
- 2.Bull JJ, Turelli M. Wolbachia versus dengue: Evolutionary forecasts. Evolution, medicine, and public health. 2013;2013(1):197–207. PubMed Central PMCID: PMC3847891. 10.1093/emph/eot018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Hughes GL, Koga R, Xue P, Fukatsu T, Rasgon JL. Wolbachia infections are virulent and inhibit the human malaria parasite Plasmodium falciparum in Anopheles gambiae. PLoS pathogens. 2011;7(5):e1002043 PubMed Central PMCID: PMC3098226. 10.1371/journal.ppat.1002043 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Hughes H, Britton NF. Modelling the use of Wolbachia to control dengue fever transmission. Bulletin of mathematical biology. 2013;75(5):796–818. 10.1007/s11538-013-9835-4 [DOI] [PubMed] [Google Scholar]
- 5.Hussain M, Lu G, Torres S, Edmonds JH, Kay BH, Khromykh AA, et al. Effect of Wolbachia on replication of West Nile virus in a mosquito cell line and adult mosquitoes. Journal of virology. 2013;87(2):851–8. PubMed Central PMCID: PMC3554047. 10.1128/JVI.01837-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.McMeniman CJ, Lane RV, Cass BN, Fong AW, Sidhu M, Wang YF, et al. Stable introduction of a life-shortening Wolbachia infection into the mosquito Aedes aegypti. Science. 2009;323(5910):141–4. 10.1126/science.1165326 [DOI] [PubMed] [Google Scholar]
- 7.van den Hurk AF, Hall-Mendelin S, Pyke AT, Frentiu FD, McElroy K, Day A, et al. Impact of Wolbachia on infection with chikungunya and yellow fever viruses in the mosquito vector Aedes aegypti. PLoS neglected tropical diseases. 2012;6(11):e1892 PubMed Central PMCID: PMC3486898. 10.1371/journal.pntd.0001892 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Walker T, Johnson PH, Moreira LA, Iturbe-Ormaetxe I, Frentiu FD, McMeniman CJ, et al. The wMel Wolbachia strain blocks dengue and invades caged Aedes aegypti populations. Nature. 2011;476(7361):450–3. 10.1038/nature10355 [DOI] [PubMed] [Google Scholar]
- 9.Bian G, Xu Y, Lu P, Xie Y, Xi Z. The endosymbiotic bacterium Wolbachia induces resistance to dengue virus in Aedes aegypti. PLoS pathogens. 2010;6(4):e1000833 PubMed Central PMCID: PMC2848556. 10.1371/journal.ppat.1000833 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Joubert DA, Walker T, Carrington LB, De Bruyne JT, Kien DH, Hoang Nle T, et al. Establishment of a Wolbachia Superinfection in Aedes aegypti Mosquitoes as a Potential Approach for Future Resistance Management. PLoS pathogens. 2016;12(2):e1005434 10.1371/journal.ppat.1005434 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.McMeniman CJ, Hughes GL, O'Neill SL. A Wolbachia symbiont in Aedes aegypti disrupts mosquito egg development to a greater extent when mosquitoes feed on nonhuman versus human blood. Journal of medical entomology. 2011;48(1):76–84. [DOI] [PubMed] [Google Scholar]
- 12.Blagrove MS, Arias-Goeta C, Failloux AB, Sinkins SP. Wolbachia strain wMel induces cytoplasmic incompatibility and blocks dengue transmission in Aedes albopictus. Proceedings of the National Academy of Sciences of the United States of America. 2012;109(1):255–60. PubMed Central PMCID: PMC3252941. 10.1073/pnas.1112021108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Andrews ES, Crain PR, Fu Y, Howe DK, Dobson SL. Reactive oxygen species production and Brugia pahangi survivorship in Aedes polynesiensis with artificial Wolbachia infection types. PLoS pathogens. 2012;8(12):e1003075 PubMed Central PMCID: PMC3516568. 10.1371/journal.ppat.1003075 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Bian G, Joshi D, Dong Y, Lu P, Zhou G, Pan X, et al. Wolbachia invades Anopheles stephensi populations and induces refractoriness to Plasmodium infection. Science. 2013;340(6133):748–51. 10.1126/science.1236192 [DOI] [PubMed] [Google Scholar]
- 15.Caragata EP, Dutra HL, Moreira LA. Exploiting Intimate Relationships: Controlling Mosquito-Transmitted Disease with Wolbachia. Trends in parasitology. 2015. [DOI] [PubMed] [Google Scholar]
- 16.McGraw EA, O'Neill SL. Beyond insecticides: new thinking on an ancient problem. Nature reviews Microbiology. 2013;11(3):181–93. 10.1038/nrmicro2968 [DOI] [PubMed] [Google Scholar]
- 17.O'Neill S. The Dengue Stopper. Sci Am. 2015;312(6):72–7. [DOI] [PubMed] [Google Scholar]
- 18.Sinkins SP. Wolbachia and arbovirus inhibition in mosquitoes. Future microbiology. 2013;8(10):1249–56. 10.2217/fmb.13.95 [DOI] [PubMed] [Google Scholar]
- 19.Baton LA, Pacidonio EC, Goncalves DS, Moreira LA. wFlu: characterization and evaluation of a native Wolbachia from the mosquito Aedes fluviatilis as a potential vector control agent. PloS one. 2013;8(3):e59619 PubMed Central PMCID: PMC3608659. 10.1371/journal.pone.0059619 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Zele F, Nicot A, Berthomieu A, Weill M, Duron O, Rivero A. Wolbachia increases susceptibility to Plasmodium infection in a natural system. Proceedings Biological sciences / The Royal Society. 2014;281(1779):20132837 PubMed Central PMCID: PMC3924077. 10.1098/rspb.2013.2837 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Mousson L, Zouache K, Arias-Goeta C, Raquin V, Mavingui P, Failloux AB. The native Wolbachia symbionts limit transmission of dengue virus in Aedes albopictus. PLoS neglected tropical diseases. 2012;6(12):e1989 PubMed Central PMCID: PMCPMC3531523. 10.1371/journal.pntd.0001989 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Jin C, Ren X, Rasgon JL. The virulent Wolbachia strain wMelPop efficiently establishes somatic infections in the malaria vector Anopheles gambiae. Applied and environmental microbiology. 2009;75(10):3373–6. PubMed Central PMCID: PMC2681643. 10.1128/AEM.00207-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Hughes GL, Vega-Rodriguez J, Xue P, Rasgon JL. Wolbachia strain wAlbB enhances infection by the rodent malaria parasite Plasmodium berghei in Anopheles gambiae mosquitoes. Applied and environmental microbiology. 2012;78(5):1491–5. PubMed Central PMCID: PMC3294472. 10.1128/AEM.06751-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Dodson BL, Hughes GL, Paul O, Matacchiero AC, Kramer LD, Rasgon JL. Wolbachia Enhances West Nile Virus (WNV) Infection in the Mosquito Culex tarsalis. PLoS neglected tropical diseases. 2014;8(7):e2965 10.1371/journal.pntd.0002965 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Hughes GL, Dodson BL, Johnson RM, Murdock CC, Tsujimoto H, Suzuki Y, et al. Native microbiome impedes vertical transmission of Wolbachia in Anopheles mosquitoes. Proceedings of the National Academy of Sciences of the United States of America. 2014;111(34):12498–503. PubMed Central PMCID: PMC4151774. 10.1073/pnas.1408888111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Ferguson NM, Kien DT, Clapham H, Aguas R, Trung VT, Chau TN, et al. Modeling the impact on virus transmission of Wolbachia-mediated blocking of dengue virus infection of Aedes aegypti. Science translational medicine. 2015;7(279):279ra37 PubMed Central PMCID: PMC4390297. 10.1126/scitranslmed.3010370 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Osborne SE, Iturbe-Ormaetxe I, Brownlie JC, O'Neill SL, Johnson KN. Antiviral protection and the importance of Wolbachia density and tissue tropism in Drosophila simulans. Applied and environmental microbiology. 2012;78(19):6922–9. PubMed Central PMCID: PMC3457512. 10.1128/AEM.01727-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Hughes GL, Rivero A, Rasgon JL. Wolbachia can enhance Plasmodium infection in mosquitoes: implications for malaria control? PLoS pathogens. 2014;10(9):e1004182 PubMed Central PMCID: PMC4154766. 10.1371/journal.ppat.1004182 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Murdock CC, Blanford S, Hughes GL, Rasgon JL, Thomas MB. Temperature alters Plasmodium blocking by Wolbachia. Scientific reports. 2014;4:3932 PubMed Central PMCID: PMC3909897. 10.1038/srep03932 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Caragata EP, Rances E, Hedges LM, Gofton AW, Johnson KN, O'Neill SL, et al. Dietary cholesterol modulates pathogen blocking by Wolbachia. PLoS pathogens. 2013;9(6):e1003459 PubMed Central PMCID: PMC3694857. 10.1371/journal.ppat.1003459 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Moreira LA, Iturbe-Ormaetxe I, Jeffery JA, Lu G, Pyke AT, Hedges LM, et al. A Wolbachia symbiont in Aedes aegypti limits infection with dengue, Chikungunya, and Plasmodium. Cell. 2009;139(7):1268–78. 10.1016/j.cell.2009.11.042 [DOI] [PubMed] [Google Scholar]
- 32.Warrilow D, Northill JA, Pyke AT. Sources of dengue viruses imported into Queensland, australia, 2002–2010. Emerging infectious diseases. 2012;18(11):1850–7. PubMed Central PMCID: PMC3559152. 10.3201/eid1811.120014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Rances E, Ye YH, Woolfit M, McGraw EA, O'Neill SL. The relative importance of innate immune priming in Wolbachia-mediated dengue interference. PLoS pathogens. 2012;8(2):e1002548 PubMed Central PMCID: PMC3285598. 10.1371/journal.ppat.1002548 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Abbreviations used: BF–Blood fed; dpi–days post injection; dpf–days post feeding; W.T.–wild type Aedes aegypti females.
(TIF)
Statistical significance was determined using a Mann-Whitney test (A and D) or a Fisher exact test (B and C). In A and D the mean and error of the mean is indicated. In B and C, the error bars represent 95% confidence levels. A) DENV genome copies in whole mosquito bodies (****, p < 0.0001; **, p = 0.004; Mann-Whitney). B) DENV infection rate as determined by the percentage of individuals infected 7 days post an infectious blood meal (****, p = 0.001; *, p < 0.05; Fisher exact test). C) DENV transmission rate as determined by the percentage of infectious saliva expectorated 7 days post an infectious blood meal, (***, p = 0.0002; *, p = 0.03; Fisher exact test). D) Wolbachia density 7 days post an infectious blood meal in transiently infected and the stable transinfected line (****, p < 0.0001; Mann-Whitney).
(TIF)
Statistical significance was determined using a Mann-Whitney test (A, B and E) or a Fisher exact test (C and D). In A, B and E, the mean and error of the mean is indicated. In C and D, the error bars represent 95% confidence levels. A) WNV (Kunjin strain) PFU per ml in whole mosquito bodies, (**, p < 0.01; Mann-Whitney). B) WNV (Kunjin strain) PFU per ml in saliva (**, p = 0.002; Mann-Whitney). C) WNV (Kunjin strain) infection rate as determined by the percentage of individuals infected 7 days post an infectious blood meal, (*, p = 0.02; Fisher exact test). D) WNV (Kunjin strain) transmission rate as determined by the percentage of infectious saliva expectorated infected 7 days post an infectious blood meal (**, p = 0.002, Mann-Whitney). E) Wolbachia density 7 days post an infectious blood meal in transiently infected and the stable transinfected lines (****, p < 0.0001; Mann-Whitney).
(TIF)
Statistical significance was determined using a Mann-Whitney test (A and D) or a Fisher exact test (B and C). In A and D, the mean and error of the mean is indicated. In B and C, the error bars represent 95% confidence levels. A) DENV genome copies in whole mosquito bodies (***, p ≤ 0.001; **, p = 0.004; Mann-Whitney). B) DENV infection rate as determined by the percentage of individuals infected 7 days post an infectious blood meal, (****, p = 0.0003; Fisher exact test). C) DENV transmission rate as determined by the percentage of infectious saliva expectorated 7 days post an infectious blood meal, (****, p = 0.0002; *, p = 0.03; Fisher exact test). D) Wolbachia density 7 days post an infectious blood meal in transiently infected and the stable transinfected line (****, p < 0.0001; Mann-Whitney).
(TIF)
Statistical significance was determined using a Mann-Whitney test (A, B and E) or a Fisher exact test (C and D). In A, B and E, the mean and error of the mean is indicated. In C and D, the error bars represent 95% confidence levels. A) WNV (Kunjin strain) PFU per ml in whole mosquito bodies, (**, p < 0.006; *, p = 0.04; Mann-Whitney). B) WNV (Kunjin strain) PFU per ml in saliva (****, p < 0.0001, **, p < 0.009; Mann-Whitney). C) WNV (Kunjin strain) infection rate as determined by the percentage of individuals infected 7 days post an infectious blood meal, (**, p = 0.002; Fisher exact test). D) WNV (Kunjin strain) transmission rate as determined by the percentage of infectious saliva expectorated infected 7 days post an infectious blood meal (**, p = 0.007; Mann-Whitney). E) Wolbachia density 7 days post an infectious blood meal in transiently infected and the stable transinfected lines (****, p < 0.0001; Mann-Whitney).
(TIF)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.