

Abstract
Vibriosis is a major epizootic disease that impacts free-living and farmed fish species worldwide. Use of probiotics is a promising approach for prevention of Vibrio infections in aquaculture. A probiotic anti-Vibrio strain, Bacillus pumilus H2, was characterized, and the mechanism of its effect was investigated. All 29 Vibrio strains tested were growth-inhibited by H2. The anti-Vibrio substance present in cell-free supernatant of H2 was purified and characterized by reversed-phase HPLC. Minimum inhibitory concentrations of the purified substance, determined in liquid media for various Vibrio strains, ranged from 0.5 to 64 µg/ml. Addition of the purified substance to Vibrio vulnificus culture inhibited cell growth (estimated by OD600). Confocal microscopy and scanning electron microscopy analyses showed that surface structure of V. vulnificus cells was damaged by the purified substance, as reflected by presence of membrane holes, disappearance of cellular contents, and formation of cell cavities. The major mechanism of this anti-Vibrio activity appeared to involve disruption of cell membranes, and consequent cell lysis. The purified anti-Vibrio substance was shown to be structurally identical to amicoumacin A by MS and NMR analysis. Our findings indicate that B. pumilus H2 has strong potential for prevention or treatment of fish vibriosis in the aquaculture industry.
Electronic supplementary material
The online version of this article (doi:10.1186/s13568-017-0323-3) contains supplementary material, which is available to authorized users.
Keywords: Anti-Vibrio, Bacillus pumilus H2, Mechanism, Amicoumacin A, Vibriosis control
Introduction
During the course of aquaculture development, major production problems have been caused by a number of bacterial diseases (Paillard et al. 2004; Stentiford et al. 2012; Toranzo et al. 2005). These disease-related problems are the largest single cause of economic losses in aquaculture (Stentiford et al. 2012; Zhou et al.2009). A small number of opportunistic bacterial pathogens are responsible for the majority of such losses worldwide (Austin and Austin 2007). The Gram-negative genus Vibrio is one of the most important groups of bacterial pathogens, and a major source of mortality (Colwell and Griems 1984; Egidius 1987; Li and Woo 2003). Vibrio species are widespread and ubiquitous in aquatic environments worldwide, occupy a variety of habitats in marine, freshwater, and estuarine ecosystems, and are frequently found in aquaculture facilities (Heidelberg et al. 2002; Tall et al. 2013; Thompson et al. 2004).
Vibriosis, a collective Vibrio infection (Egidius 1987), is a widespread epizootic disease that affects most free-living and farmed fish species worldwide, and is currently the major limiting factor in development of intensive mariculture industry (Egidius 1987). In association with the rapid expansion of intensive mariculture and consequent deterioration of culture conditions, a steadily increasing number of Vibrio species are recognized as pathogens in vibriosis outbreaks (Austin and Zhang 2006; Cui et al. 2014; Hou et al. 2016). A limited number of antibiotics have been successfully applied, and resistance to these antibiotics may reduce the success of treatment programs (Al-Othrubi et al. 2014; Elmahdi et al. 2016).
The term “probiotic” was introduced by Parker in 1974, referring to “organisms and substances that have a beneficial effect on the host animal by contributing to its intestinal microbial balance” (Parker 1974). Many groups have investigated the benefits of using probiotic strains in aquaculture (Balcázar et al. 2006; Desriac et al. 2010; Moriarty 1998; Newaj-Fyzul et al. 2014; Verschuere et al. 2000). Species and strains of Bacillus, a genus of Gram-positive, rod-shaped bacteria, exert antagonistic or inhibitory activities against a variety of bacterial and fungal pathogens, and have been utilized frequently as probiotics for treatment and/or prevention of infectious processes in many plants and animals (Mongkolthanaruk 2012; Mondol et al. 2013; Patel et al. 2009; Wulff et al. 2002).
In previous study of our lab, probiotic effect of Bacillus pumilus H2 to juvenile shrimp was carried out in aquaculture tanks (Fu et al. 2009). Juvenile shrimp were exposed to B. pumilus H2 at 0 (as control), 103 and 104 CFU/ml for 14 days before a challenge with Vibrio natriegens at 104 CFU/ml for 1 day infection. The final mortality of the shrimp group treated with 104 CFU/ml B. pumilus H2 was only 12.5%, much lower than the group treated with 103 CFU/ml B. pumilus H2 (28.3%) and the control group (30.8%, P < 0.05); and the average weight and length of the shrimp group treated with 104 CFU/ml B. pumilus H2 were also higher than those of the control group (Fu et al. 2009). And results showed that H2 might have good application prospects and significance.
In the present study, we: (1) further screened Bacillus strains that displayed sufficient anti-Vibrio activity to be considered as biocontrol agents, (2) measured in vitro antagonistic activity of probiotic strain B. pumilus H2 against Vibrio species, and (3) extracted and purified antimicrobial compounds from H2, and made preliminary studies of their inhibitory mechanisms. The major of anti-Vibrio mechanism of H2 appeared to be disruption of the cell membrane, and the active anti-Vibrio compound was structurally identified as amicoumacin A. Our findings indicate that H2 has strong potential application in prevention or control of fish vibriosis.
Materials and methods
Bacterial strains and culture conditions
Bacterial strains used in this study included 29 Vibrio species, four Bacillus species, and two Aeromonas species (Table 1). All strains were confirmed by sequencing of their 16S rRNA gene. All Vibrio and Aeromonas species were used as target strains (indicator strains). Strains were recovered from a lyophilized ampoule or frozen stocks for 36 h aerobic incubation in liquid LB medium before use, and they were grown in LB medium or on LB plates at 30 °C under aerobic conditions.
Table 1.
Bacterial strains used in this study
| Genus | Species and strain | Source(s) | Date of collection |
|---|---|---|---|
| Vibrio | V. vulnificus CZ-A2, V. diazotrophic CZ-G1, V. ponticus CZ-L7, V. neptunius CZ-D1, V. rotiferianus CZ-F1, V. sinaloensis PE7, V. communis J7, V. azureus D3, V. scophthalmi E3, V. chagasii T3, V. campbellii AF5, Vibrio ponticus B8 | Biofilters, fish ponds of marine aquaculture recirculating system | Collected by our lab in 2011 |
| V. algoinfesta QBST8, V. alfacsensis QBST3, V. alginolyticus LM3-1, V. sinaloensis QBSM3, V. cyclitrophicus DFWB3, V. fortis QBLM3, V. owensii QBST1, V. ponticus W6-3, V. harveyi LM2, V. rotiferianus W5-3 | Skin, liver, and spleen of diseased marine aquaculture animals | Collected by our lab in 2013 | |
| V. alginolyticus CGMCC 1.1607, V. parahaemolyticus CGMCC 1.2164, V. fischeri CGMCC 1.1613 | China General Microbiological Culture Collection Center (CGMCC) | Bought from CGMCC in 2013 | |
| Bacillus | B. pumilus H2 (CGMCC No. 1004), B. safensis H2-2 (CGMCC No. 1006) | Marine sediment | Collected by our lab in 2005 |
| B. velezensis V4 (CGMCC No. 10149) | Marine aquaculture pond | Collected by our lab in 2011 | |
| B. methylotrophicus L7 | Stored in lab | Collected by our lab in 2011 | |
| Aeromonas | A. hydrophila CGMCC 1.0927 | CGMCC | Bought from CGMCC in 2013 |
| A. salmonicida E11I4 | Diseased marine fish | Collected by our lab in 2014 |
Preparation of cell suspension of indicator strains
Indicator strains were inoculated in LB broth, incubated 24 h at 30 °C with shaking (150 rpm), and optical density at 600 nm (OD600) was determined. Cell suspensions of indicator strains were obtained by adjusting OD600 to 0.8 using sterile LB broth.
Preparation of cell-free supernatant (CFS) of Bacillus strains
Bacillus strains were inoculated and incubated as above, and cells were removed by centrifugation (8000×g) for 10 min at 4 °C. Supernatants were passed through sterile syringe filters to obtain CFS.
Screening and characterization of anti-Vibrio strains
Two approaches were used for screening of Bacillus strains having anti-Vibrio activity: (1) A given Bacillus strain was inoculated as a spot (diameter ~2–3 mm) on the surface of a LB agar plate spread with cell suspension of a given indicator strain. Cells were incubated 48 h at 30 °C, and antagonistic activity was evaluated based on the presence of a growth inhibition zone around the spot. (2) 10 µl CFS was dropped onto a 6-mm paper disk on an agar plate spread with cell suspension of a given indicator strain, and incubated 24 h at 30 °C. Anti-Vibrio activity was assessed as diameter (mm) of the inhibition zone between the disk and the bacterial lawn.
Aeromonas salmonicida E11I4 and A. hydrophila CGMCC 1.0927 were tested as reference strains
Extraction and purification of antimicrobial compounds
Strain H2 was inoculated on three 200 ml LB broth for 24 h using shaking flasks (150 r/min) at 30 °C. 600 ml CFS in total was lyophilized (Heto PowerDry PL6000, Thermo Scientific, USA), lyophilized material was extracted with methanol, and dried methanol extract was dissolved in 20 mM Tris–HCl (pH 7.0) and applied to a solid-phase extraction (SPE) column (Bond Elut C18, Varian, USA) to remove impurities. Fractions (each 10 ml) were eluted from the SPE column by acetonitrile concentration gradient (0, 10, 20, 30, 40% acetonitrile in H2O), and anti-Vibrio activity of each fraction was tested using V. vulnificus as indicator. Active fractions were pooled, lyophilized, and further purified by reversed-phase high performance liquid chromatography (RP-HPLC). A C18 semi-preparative column (Zorbax SB-C18, 5 µm, 9.4 × 150 mm, Agilent) was developed with gradient 20% acetonitrile/0.1% trifluoroacetic acid (TFA) in H2O to 40% acetonitrile/0.1% TFA in H2O, from 5 to 42 min, at flow rate 2 ml/min. Anti-Vibrio activity of each collected peak was assessed using V. vulnificus as indicator. The active peak was identified at 16 min, which corresponds to acetonitrile concentration 34%. Purified anti-Vibrio substance was obtained by lyophilization of this fraction.
Determination of minimum inhibitory concentration (MIC)
MICs of purified antimicrobial substances from various Vibrio strains were determined by broth microdilution assays in 96-well microwell plates. 100 µl of twofold serial dilutions of purified anti-Vibrio substance was mixed with an equal volume of 1:100-diluted overnight Vibrio cultures in sterile LB. Negative control wells (without purified anti-Vibrio substance) and wells containing only LB were included in the assay. Plates were incubated 24 h at 30 °C. Concentration of colony-forming units (CFUs) in the bacterial inoculum was ~105 CFU/ml. MIC was defined as the lowest concentration of antimicrobial substance that completely inhibited bacterial growth.
Confocal microscopy
Vibrio vulnificus cells grown for 24 h were incubated 24 h with anti-Vibrio substance (final concentration 0.5, 5, or 10 µg/ml), or with PBS alone. SYTOX green (SG) (Molecular Probes, Invitrogen, USA) was then added (final concentration 0.8 µM), and samples were incubated 15 min in the dark. Cells were washed, resuspended in phosphate buffer saline (PBS), prepared as confocal slides, and visualized by confocal microscopy. Fluorescence was photographed with a fluorescence microscope (model CTR 5000, Leica, Germany), with filters set at excitation wavelength 488 nm/emission wavelength 538 nm, for SG detection.
Scanning electron microscopy (SEM)
Vibrio vulnificus cells grown for two days in LB were incubated 24 h with anti-Vibrio substance (0.5 µg/ml), with sterile PBS (pH 7.2) as control. Cells were resuspended in 2.5% (v:v) glutaraldehyde solution in 0.1 M PBS, and fixed for 24 h. The glutaraldehyde was removed, and 1% osmium tetroxide solution (pH 7.2) was added. After 1.5 h, cells were washed three times with PBS. Cells were then (1) dehydrated by cold ethanol concentration gradient (10, 30, 50, 70, 90%; 10 min each), and (2) dehydrated twice in 100% ethanol at 10 min intervals. For SEM assay, cells were washed with 50, 70, 90, and 100% isoamyl acetate (each 3 min), critical point dried, coated with gold/palladium, and observed and photographed with a scanning electron microscope (model S-3400N, Hitachi Instruments, Japan).
Structure determination of antimicrobial compound
For mass spectrometer (MS) analysis, purified anti-Vibrio substance was dissolved in 30% acetonitrile in H2O and injected into an Orbitrap Fusion mass spectrometer (Thermo-Fisher, USA). For nuclear magnetic resonance (NMR) analysis, purified anti-Vibrio substance (5 mg) was dissolved in 200 μl dimethyl sulfoxide (DMSO), and samples were pipetted into a DMSO-matched NMR tube (Shigemi Co., Japan) for NMR analysis (model Avance III, 500 MHz, Bruker, USA).
Results
Screening of probiotic Bacillus strains
Four Bacillus strains (B. velezensis V4, B. methylotrophicus L7, B. pumilus H2, B. safensis H2-2) were screened for anti-Vibrio activity. Each of the four strains had growth-inhibiting effects on various Vibrio strains. B. pumilus H2 had the broadest anti-Vibrio activity spectrum; it inhibited all 29 Vibrio strains tested to varying degrees (Additional file 1: Table S1). When CFS of H2 was tested, the diameter of its inhibition zone for the Vibrio strains ranged from 7 to 18 mm. When supernatant was concentrated, the inhibition zone diameters became significantly larger (17 to 25 mm). We therefore selected H2 as the probiotic strain used in further studies of anti-Vibrio activity.
Growth curve and anti-Vibrio activity of B. pumilus H2
H2 accumulated anti-Vibrio substance in culture broth before 24 h. As incubation continued, anti-Vibrio activity declined, and was essentially gone after 3–4 days (Fig. 1).
Fig. 1.

Growth curve and anti-Vibrio activity of B. pumilus strain H2
Effects of enzymes, heat, pH, and chemicals on anti-Vibrio activity
We performed a series of stability assays to gather information on chemical structure of the anti-Vibrio substance, and reference data for its practical application as a probiotic.
The results (Additional file 1: Table S2) indicated that the anti-Vibrio substance presented in the CFS was very thermal stable, there was 69.73% activity remained after being treated at 121 °C for 15 min, in comparison with the control (−20 °C, 60 min). The anti- Vibrio substance also performed quite well in resistance to enzyme digestion, since none of the enzymes tested (proteinase K, trypsin, chymotrypsin, lysozyme) caused complete disappearance of activity. The results also showed that organic solvents only slightly affected the anti-Vibrio substance, most of the activity (80–90% relative activity) was remained after being treated with addition of equal volume of organic solvents at 37 °C for 1 h. Activity was maintained over a wide range of pH values, from 2 to 10. UV irradiation had little effect on activity, there was only a 12% reduction even after exposure to UV at a distance of 25 cm for 5 h (Additional file 1: Table S2).
Extraction and purification of anti-Vibrio substance
The anti-Vibrio substance present in B. pumilus H2 CFS was purified by SPE and RP-HPLC. The HPLC spectrum showed three peaks (Fig. 2). In anti-Vibrio activity assays using V. vulnificus CZ-A2 as target organism, activity was strong for peak 1 and very weak for peak 2, suggesting that the peak 1 substance was the major anti-Vibrio compound. In CFS cultured for 24 h, peak 1 was dominant, whereas in CFS cultured for 36 h peak 2 was dominant one and peak 1 declined greatly. This observation is consistent with the activity curve associated with H2 growth (Fig. 1).
Fig. 2.
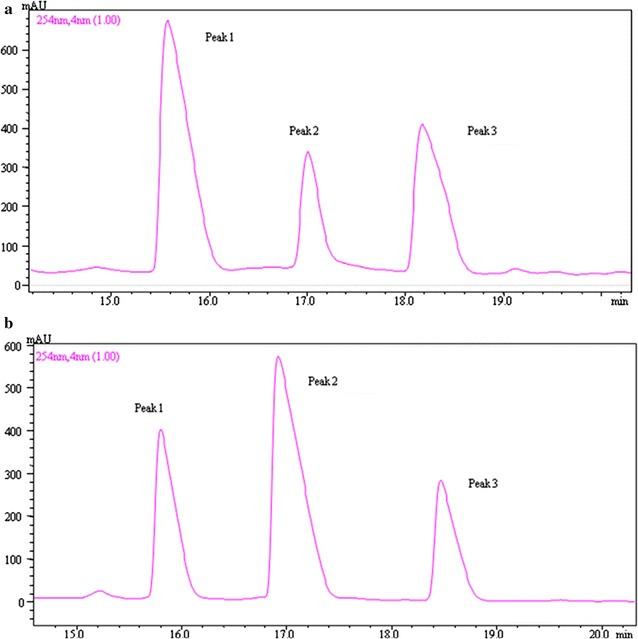
Spectra of crude extract after purification by SPE and RP-HPLC, from 24 h-CFS (a) and 36 h-CFS (b)
A total of 20 mg purified anti-Vibrio substance was obtained by semi-preparative RP-HPLC and used for subsequent experiments.
MICs of purified anti-Vibrio substance for various Vibrio strains
MICs of purified anti-Vibrio substance for various Vibrio strains, determined in liquid media, ranged from 0.25 to 64 μg/ml (Table 2). The purified substance showed high inhibitory activity against V. natriegens FS-1, V. vulnificus CZ-A2, V. harveyi PH4, V. sinaloensis PE7, and V. ponticus B8, but less activity against V. diazotrophicus CZ-G1, V. alginolyticus CGMCC 1.1607, and V. parahaemolyticus CGMCC 1.2164.
Table 2.
MICs of purified anti-Vibrio substance for various Vibrio strains
| MIC (µg/ml) | Strain |
|---|---|
| 0.25 | Vibrio natriegens FS-1 |
| 0.5 | V. vulnificus CZ-A2, V. harveyi PH4, V. sinaloensis PE7, V. ponticus B8 |
| 2 | V. alfacsensis QBST3, V. communis J7 |
| 4 | V. azureus D3, V. algoinfesta QBST8, V. fortis QBLM4 |
| 8 | V. alginolyticus LM3-1, V. cyclitrophicus DFWB3, V. scophthalmi E3, V. fortis QBLM3, V. owensii QBST1, V. ponticus W6-3, V. rotiferianus CZ-F1, V. rotiferianus W5-3 |
| 16 | V. neptunius CZ-D1, V. sinaloensis QBSM3, V. campbellii AF5, V. harveyi LM2, V. fischeri CGMCC 1.1613 |
| 32 | V. ponticus CZ-L7, V. chagasii T3, V. anguillarum XP |
| 64 | V. diazotrophic CZ-G1, V. alginolyticus CGMCC 1.1607, V. parahaemolyticus CGMCC 1.2164 |
Effect of purified anti-Vibrio substance on growth of V. vulnificus
Vibrio vulnificus was selected as a model target strain for experiments on the mode of interaction of purified anti-Vibrio substance with Vibrio strains. V. vulnificus was inoculated into LB broth (100 ml) to OD600 ~0.25, and purified anti-Vibrio substance was added (control = sterile distilled water). Growth was monitored by measuring OD600 of the culture at predetermined intervals. Growth of the target strain was clearly inhibited by addition of anti-Vibrio substance; i.e., OD600 did not increase as it did in control culture (Fig. 3). OD600 declined continuously after 8 h incubation, suggesting that cell lysis was occurring. In the control group, the target strain showed exponential growth immediately after inoculation (Fig. 3).
Fig. 3.

Effect of purified anti-Vibrio substance on growth of V. vulnificus
Effect of purified anti-Vibrio substance on cell surface structure of V. vulnificus
Membrane integrity of V. vulnificus (target strain) following treatment with purified anti-Vibrio substance was evaluated by confocal microscopy and an assay based on uptake of the fluorescent dye SG. SG, a high-affinity nucleic acid stain, is often used to assess integrity of plasma membranes, because it easily penetrates cells with compromised membranes, but does not pass through membranes of non-compromised cells.
Nontreated V. vulnificus cells (control) showed no appreciable fluorescent signal (Fig. 4). Treatment with 0.5 μg/ml purified substance resulted in detection of only a very weak fluorescent signal. Intensity of the fluorescent signal increased steadily as substance concentration increased. At substance concentration 10 µg/ml, the fluorescent signal was distinct, clear, and strong, indicating that the cell membrane was completely permeabilized.
Fig. 4.
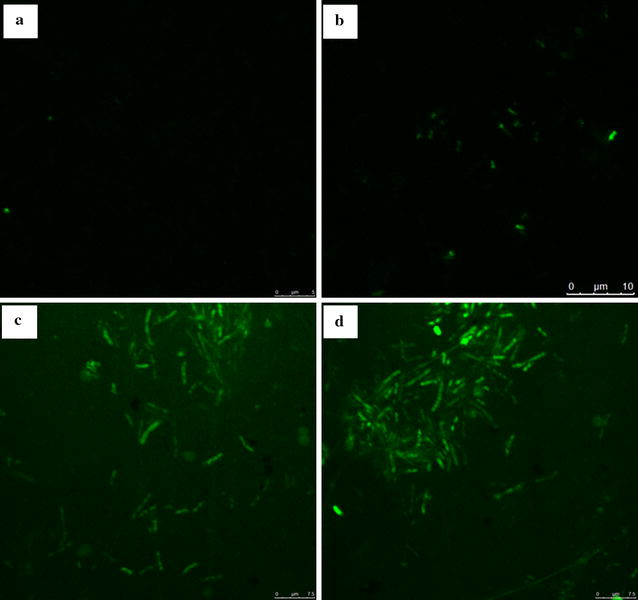
Confocal microscopic images of V. vulnificus cells treated with purified anti-Vibrio substance at 0 μg/ml (a), 0.5 μg/ml (b), 5 μg/ml (c), and 10 μg/ml (d)
Changes in surface structure of V. vulnificus cells resulting from treatment with purified anti-Vibrio substance were analyzed by SEM. Nontreated cells were intact, smooth, and displayed fine structure (Fig. 5a). In contrast, cells treated with the substance showed clear surface structure damage, including appearance of membrane holes, disappearance of cellular contents, and formation of cell cavities (Fig. 5b–d). The size of these cavities (292 × 732 nm in Fig. 5c; 361 × 559 nm in Fig. 5d) indicated that cell lysis had occurred.
Fig. 5.
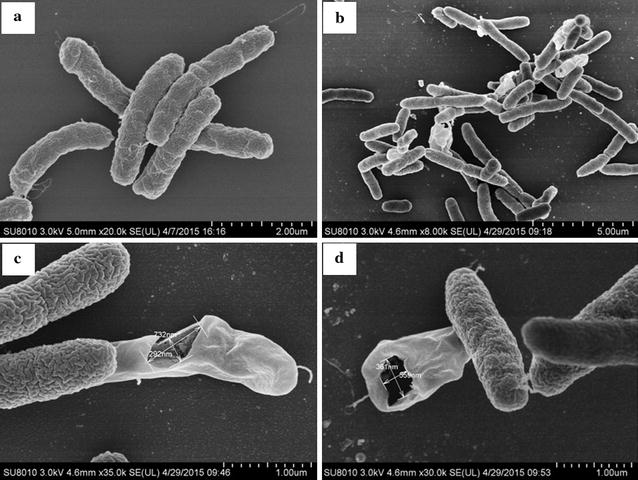
SEM images of V. vulnificus CZ-A2 cells treated with purified anti-Vibrio substance at 0 μg/ml (a, 20,000×; control) and 0.5 μg/ml (b, 8000×; c, 35000×; d, 30,000×), showing formation of membrane holes
Structural characterization of purified anti-Vibrio substance
The molecular mass of the purified anti-Vibrio substance (peak 1 in Fig. 2) was 423.2076 Da, as determined by Orbitrap Fusion MS. The MS analysis also revealed a plausible chemical formula (C20H30N3O7) for the substance (Fig. 6). Chemical shifts and coupling constants were assigned to protons in the molecule through NMR analysis. The 13C-NMR spectra of the purified anti-Vibrio substance (peak 1 in Fig. 2) showed twenty signals at 21.7 (q), 23.6 (q), 25.2 (d), 30.0 (t), 32.3 (t), 39.1 (t), 50.2 (d), 51.2 (d), 71.2 (d), 73.2 (d), 82.1 (d), 108.6 (s), 170.5 (s), 173.8 (s) and 175.1 (s) ppm, respectively. Comparisons of molecular mass, chemical formula, and NMR data with those of compounds previously described in the literature indicated that the purified anti-Vibrio substance is identical to amicoumacin A (Itoh et al. 1981). Two structurally related substances (peaks 2 and 3 in Fig. 2) were also characterized. Based on MS analysis, peak 2 ((M + H)+ ion at m/z 425 Da) was identified as amicoumacin B (C20H28N2O8), and peak 3 ((M + H)+ ion at m/z 407 Da) was identified as amicoumacin C (C20H26N2O7) (data not shown).
Fig. 6.
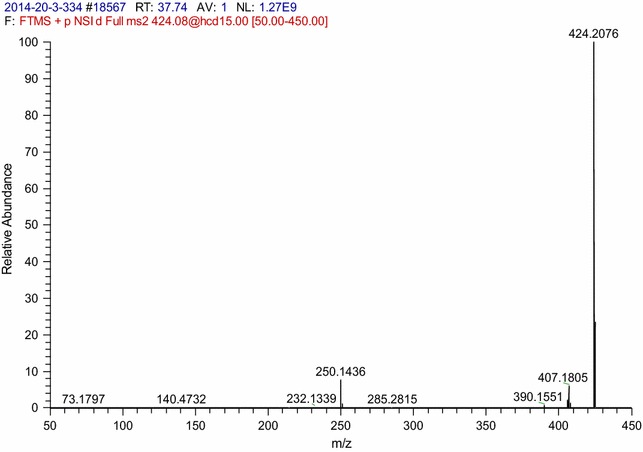
MS analysis of purified anti-Vibrio substance from HPLC peak 1 in Fig. 2
Discussion
As the aquaculture industry expands worldwide, and the variety of fish species involved increases, many unknown fish-pathogenic Vibrio species are reported (Li and Woo 2003; Thompson et al. 2004; Austin and Zhang 2006; Cui et al. 2014). In the past, vibriosis was controlled (prevented or treated) almost exclusively through application of antibiotics or chemotherapeutic agents, either as feed additives or in immersion baths. However, extensive use of this approach over time has resulted in increased resistance of pathogenic Vibrio strains to the commonly used antibiotics: ampicillin, amikacin, kanamycin, penicillin G, streptomycin, and tetracycline (Austin and Austin 2007; Li et al. 1999; Elmahdi et al. 2016).
In present study, we screened Bacillus strains for anti-Vibrio activity. Bacillus strains are good candidates as biological control agents for prevention or treatment of plant and animal infections for several reasons (Wulff et al. 2002; Mongkolthanaruk 2012; Mondol et al. 2013). (1) They produce antibiotics having well-documented antagonistic activity against a variety of fungal and bacterial pathogens. (2) They form spores that can be easily formulated, and have high viability in comparison with vegetative cells. (3) The robustness of the spores enables them to cross the gastric barrier. A certain proportion of spores is thus able to germinate in and colonize (albeit briefly) the intestinal tract (Mazza 1994). (4) Bacillus species are abundant in a wide variety of environments and habitats.
Bacillus pumilus strain H2 has notable anti-Vibrio effects. It inhibited 29 Vibrio strains to varying degrees (Table 2). No anti-Vibrio probiotic has been previously reported to inhibit such a large number of Vibrio strains. V. vulnificus CZ-A2, V. natriegens FS-1, V. harveyi PH4, V. sinaloensis PE7, V. ponticus B8, V. alfacsensis QBST3, and V. communis J7 were highly sensitive to purified anti-Vibrio substance. V. anguillarum is the most well-studied and widespread fish-pathogenic Vibrio species, and is responsible for the majority of fish loss in aquaculture worldwide (Austin and Austin 2007). We measured MIC of purified anti-Vibrio substance from H2 against V. anguillarum XP as 32 µg/ml, indicating its potential application for control of this major pathogen.
We selected V. vulnificus CZ-A2 as the target strain for screening of Bacillus strains having anti-Vibrio activity, and for follow-up studies of the mechanism of H2 activity, because CZ-A2 was highly sensitive to the substance present in H2 CFS. V. vulnificus is a widespread marine bacterium categorized into three biotypes. Strains of V. vulnificus include one of the most widely occurring fish pathogens, and another strain that can cause wound infections in humans, resulting in high mortality among susceptible individuals (Efimov et al. 2013; Ziolo et al. 2014).
The anti-Vibrio substance produced by B. pumilus H2 was found to be structurally identical to amicoumacin A, which was described in 1981 (Itoh et al. 1981). Amicoumacin A and related compounds display inhibitory activity against numerous pathogenic bacteria, including Helicobacter pylori and methicillin-resistant Staphylococcus aureus (MRSA) (Pinchuk et al. 2001; Lama et al. 2012). Anti-inflammatory and antitumor effects of amicoumacin A have also been reported (Itoh et al. 1981), but no study to date has addressed its effects against pathogenic Vibrio that cause economic losses in aquaculture. Amicoumacin B was isolated from B. pumilus and reported to display gastroprotective activity, but weak antibacterial and weak antiulcer activity (Shimojima et al. 1984; Han et al. 2013), consistent with the weak anti-Vibrio activity that we observed. Activity of amicoumacin C has not been studied. Although application of H2 preparation was used and showed no toxic to juvenile shrimp, further tests on toxic of purified Amicoumacin A to one or more farmed species need to be conducted in the future.
The mode of action of amicoumacin A remains unclear. Lama et al. reported amicoumacin A-induced alteration of transcription of genes that regulate various cellular processes, including cell envelope turnover, cross-membrane transport, virulence, metabolism, and general stress. The gene most highly induced by amicoumacin A was lrgA, which encodes an antiholin-like product (LrgA) that appears in cells undergoing collapse of Δψ, and modulates murein hydrolase activity (Lama et al. 2012). Taken together, the findings of Lama et al. suggest that amicoumacin A provokes perturbation of the cell membrane and consequent energy dissipation (Lama et al. 2012). Polikanov et al. proposed that amicoumacin A is a potent inhibitor of protein synthesis, but without direct experimental evidence (Polikanov et al. 2014). Our observations of reduced cell density (Fig. 3), formation of membrane holes, disappearance of cellular contents, and formation of cell cavities (Figs. 4, 5) indicates that the major mechanism of amicoumacin A activity against pathogens involves disruption of cell membranes, and consequent cell lysis.
Bacillus pumilus H2 was isolated from marine sediment, and therefore has priority and inherent advantages for use in marine aquaculture as a biocontrol agent or probiotic. Under the generally accepted definition of the term “probiotic”, we can consider two application approaches. In the first approach, H2 fermentation broth would be added, in proportion, directly to the aquaculture pond. Live H2 cells would then produce amicoumacin A continuously. In simulated gastroenteric environments designed to test spore robustness, H2 spores were able to cross the gastric barrier (data not shown), and may therefore have the ability to germinate and colonize in the intestinal tract. Probiotic effects in aquaculture are not limited to the intestinal tract, but may also improve the health of the host by inhibiting pathogens and improving water quality through modification of microbial community composition in the water and sediment (Perez-Sanchez et al. 2013).
In the second approach, extracted amicoumacin A would be added to aquaculture feeds. Amicoumacin A is potentially suitable for this purpose because it is heat stable, pH stable, UV stable, and not sensitive to various enzymes and organic solvents. On the other hand, a major disadvantage of the second approach is that amicoumacin A is difficult to extract and purify. The first approach appears more feasible.
In conclusion, probiotic B. pumilus strain H2 demonstrated notable antagonistic activity against 29 Vibrio strains tested. This activity was attributable to production of amicoumacin A, which has been reported previously to inhibit methicillin-resistant Staphylococcus aureus and Helicobacter pylori, and to display anti-inflammatory and antitumor effects. The major mechanism of amicoumacin A activity against pathogens involves disruption of cell membranes, and consequent cell lysis.
Authors’ contributions
XYG performed the experiments, analyzed the data as well as results and wrote the manuscript. EWL assisted NMR data analysis. TTH isolated vibrio strains used in this work. ZPL, YL and LLM conceived this study. ZPL supervised all the experiments and revised the manuscript. All authors read and approved the final manuscript.
Acknowledgements
The authors are grateful to Dr. S. Anderson for English editing of the manuscript.
Competing interests
The authors declare that they have no competing interests.
Funding
This study was supported by the Innovation Project of Shandong Province, China (2014ZZCX06204) and CAS Knowledge Innovation Project (KZCX2-EW-Q212).
Abbreviations
- CFS
cell-free supernatant
- SPE
solid-phase extraction
- RP-HPLC
reversed-phase high performance liquid chromatography
- MIC
minimum inhibitory concentration
- CFUs
colony-forming units
- SG
SYTOX green
- PBS
phosphate buffer saline
- DMSO
dimethyl sulfoxide
- MS
mass spectrometer
- NMR
nuclear magnetic resonance
- SEM
scanning electron microscope
- MRSA
methicillin-Resistant Staphylococcus aureus
Additional file
Contributor Information
Xi-Yan Gao, Email: gaoxiyan88@126.com.
Ying Liu, Email: liuying@im.ac.cn.
Li-Li Miao, Email: miaoll@im.ac.cn.
Er-Wei Li, Email: liew@im.ac.cn.
Ting-Ting Hou, Email: zhuyeqingting@yeah.net.
Zhi-Pei Liu, Phone: +86-10-64806081, Email: liuzhp@im.ac.cn.
References
- Al-Othrubi SMY, Kqueen CY, Mirhosseini H, Hadi YA, Radu S. Antibiotic resistance of Vibrio parahaemolyticus isolated from cockles and shrimp sea food marketed in Selangor, Malaysia. Clin Microbiol. 2014;3:1–7. [Google Scholar]
- Austin B, Austin DA. Bacterial fish pathogens. 4. Chichester: Ellis Horwood Ltd.; 2007. [Google Scholar]
- Austin B, Zhang XH. Vibrio harveyi: a significant pathogen of marine vertebrates and invertebrates. Lett Appl Microbiol. 2006;43:119–124. doi: 10.1111/j.1472-765X.2006.01989.x. [DOI] [PubMed] [Google Scholar]
- Balcázar JL, Blas I, Ruiz-Zarzuela I, Cunningham D, Vendrell D, Múzquiz JL. The role of probiotics in aquaculture. Vet Microbiol. 2006;114:173–186. doi: 10.1016/j.vetmic.2006.01.009. [DOI] [PubMed] [Google Scholar]
- Colwell RR, Griems DJ. Vibrio diseases of marine fish populations. Helgol Mar Res. 1984;37:265–287. [Google Scholar]
- Cui J, Fan XT, Liu WZ, Li HY, Zhou YC, Wang SF, Xie ZY. Isolation and identification of vibriosis pathogens of marine cultured fishes in southern China. Nat Sci J HaiNan Univ. 2014;32:244–251. [Google Scholar]
- Desriac F, Defer D, Bourgougnon N, Brillet B, Chevalier PL, Fleury Y. Bacteriocin as weapons in the marine animal-associated bacteria warfare: inventory and potential applications as an aquaculture probiotic. Mar Drugs. 2010;8:1153–1177. doi: 10.3390/md8041153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Efimov V, Danin-Poleg Y, Raz N, Elgavish S, Linetsky A, Kashi Y. Insight into the evolution of Vibrio vulnificus biotype 3′s genome. Front Microbiol. 2013;4:393. doi: 10.3389/fmicb.2013.00393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Egidius E. Vibriosis: pathogenicity and pathology: a review. Aquaculture. 1987;67:15–28. doi: 10.1016/0044-8486(87)90004-4. [DOI] [Google Scholar]
- Elmahdi S, DaSilva LV, Parveen S. Antibiotic resistance of Vibrio parahaemolyticus and Vibrio vulnificus in various countries: a review. Food Microbiol. 2016;57:128–134. doi: 10.1016/j.fm.2016.02.008. [DOI] [PubMed] [Google Scholar]
- Fu SZ, Song BB, Liu Y, Liu ZP. Screening of probiotic strains and their protection against vibriosis in juvenile shrimp (Litopenaeus vannamei) China Environ Sci. 2009;29:867–872. [Google Scholar]
- Han XY, Liu SW, Wang FF, Liu JM, Chen C, Hu XX, Jiang ZK, You XF, Shang GD, Zhang YB, Sun CH. Research progress of amicoumacin group antibiotic. World Notes Antibiot. 2013;34:106–114. [Google Scholar]
- Heidelberg JF, Heidelberg KB, Colwell RR. Bacteria of the gamma-subclass proteobacteria associated with zooplankton in Chesapeake Bay. Appl Environ Microbiol. 2002;68:5498–5507. doi: 10.1128/AEM.68.11.5498-5507.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hou TT, Zhong ZP, Liu Y, Liu ZP. Bacterial community characterization of rearing water of marine recirculating aquaculture systems for yellow grouper (Epinephelus awoara) Acta Microbiol Sin. 2016;56:253–263. [PubMed] [Google Scholar]
- Itoh J, Omoto S, Shomura T, Nishizawa N, Miyado S, Yuda Y, Shibata U, Inouye S. Amicoumacin-A, a new antibiotic with strong anti-inflammatory and antiulcer activity. J Antibiot. 1981;34:611–613. doi: 10.7164/antibiotics.34.611. [DOI] [PubMed] [Google Scholar]
- Lama A, Pane-Farre J, Chon T, Wiersma AM, Sit CS, Vederas JC, Hecker M, Nakano MM. Response of Methicillin-resisitant Staphylococcus aureus to Amicoumacin A. PLoS ONE. 2012;7:e34037. doi: 10.1371/journal.pone.0034037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li J, Woo NYS. Pathogenicity of vibriosis in fish: an overview. J. Ocean Univ China. 2003;2:117–128. [Google Scholar]
- Li J, Yie J, Foo RWT, Ling JML, Xu H, Woo NYS. Antibiotic resistance and plasmid profiles of vibrio isolates from cultured silver sea bream, Sparus sarba. Mar Pollut Bull. 1999;39:245–249. doi: 10.1016/S0025-326X(99)00062-4. [DOI] [PubMed] [Google Scholar]
- Mazza P. The use of Bacillus subtilis as an antidiarrhoeal microorganism. Boll Chim Farm. 1994;133:3–18. [PubMed] [Google Scholar]
- Mondol MAM, Shin HJ, Islam MT. Diversity of secondary metabolites from marine Bacillus species: chemistry and biological activity. Mar Drugs. 2013;11:2846–2872. doi: 10.3390/md11082846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mongkolthanaruk W. Classification of Bacillus beneficial substances related to plants, humans and animals. J Microbiol Biotechnol. 2012;22:1597–1604. doi: 10.4014/jmb.1204.04013. [DOI] [PubMed] [Google Scholar]
- Moriarty DJW. Control of luminous Vibrio species in penaeid. Aquaculture. 1998;164:351–358. doi: 10.1016/S0044-8486(98)00199-9. [DOI] [Google Scholar]
- Newaj-Fyzul A, Al-Harbi AH, Austin B. Review: developments in the use of probiotics for disease control in aquaculture. Aquaculture. 2014;431:1–11. doi: 10.1016/j.aquaculture.2013.08.026. [DOI] [Google Scholar]
- Paillard C, Roux FL, Borrego JJ. Bacterial disease in marine bivalves, a review of recent studies: trends and evolution. Aquat Living Resour. 2004;17:477–498. doi: 10.1051/alr:2004054. [DOI] [Google Scholar]
- Parker RB. Probiotics, the other half of the antibiotic story. Anim Nutr Health. 1974;29:4–8. [Google Scholar]
- Patel AK, Deshattiwar MK, Chaudhari BL, Chincholkar SB. Production, purification and chemical characterization of the catecholate siderophore from potent probiotic strains of Bacillus spp. Bioresour Technol. 2009;100:368–373. doi: 10.1016/j.biortech.2008.05.008. [DOI] [PubMed] [Google Scholar]
- Perez-Sanchez T, Ruiz-Zarzuela I, Blas I, Balcazar JL. Probiotics in aquaculture: a current assessment. Rev in Aquac. 2013;5:1–14. doi: 10.1111/j.1753-5131.2012.01073.x. [DOI] [Google Scholar]
- Pinchuk IV, Bressollier P, Verneuil B, Fenet B, Sorokulova IB, MéGraud F, Urdaci MC. In vitro anti-Helicobacter pylori activity of the probiotic strain Bacillus subtilis 3 is due to secretion of antibiotics. Antimicrob Agents Chemother. 2001;45:3156–3161. doi: 10.1128/AAC.45.11.3156-3161.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Polikanov YS, Osterman IA, Szal T, Tashlitsky VN, Serebryakova MV, Kusochek P, Bulkley D, Malanicheva IA, Efimenko TA, Efremenkova OV, Konevega AL, Shaw KJ, Bogdanov AA, Rodnina MV, Dontsova OA, Mankin AS, Steitz TA, Sergiev PV. Amicoumacin A inhibits translation by stabilizing mRNA interaction with the ribosome. Mol Cell. 2014;56:531–540. doi: 10.1016/j.molcel.2014.09.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shimojima Y, Hayashi H, Ooka T, Shibukawa M. Studies on AI-77s, microbial products with gastroprotective activity. Structures and the chemical nature of AI-77s. Tetrahedron. 1984;40:2519–2527. doi: 10.1016/S0040-4020(01)83504-3. [DOI] [Google Scholar]
- Stentiford GD, Neil DM, Peeler EJ, Shields JD, Small HJ, Flegel TW, Vlak JM, Jones B, Morado F, Moss S, Lotz J, Bartholomay L, Behringer DC, Hauton C, Lightner DV. Disease will limit future food supply from the global crustacean fishery and aquaculture sectors. J Invertebr Pathol. 2012;110:141–157. doi: 10.1016/j.jip.2012.03.013. [DOI] [PubMed] [Google Scholar]
- Tall A, Hervio-Heath D, Teillon A, Boisset-Helbert C, Delesmont R, Bodilis J, Touron-Bodilis A. Diversity of Vibrio spp. isolated at ambient environmental temperature in the Eastern English Channel as determined by pyrH sequencing. J Appl Microbiol. 2013;114:1713–1724. doi: 10.1111/jam.12181. [DOI] [PubMed] [Google Scholar]
- Thompson JR, Randa MA, Marcelino LA, Tomita-Mitchell A, Lim E, Polz MF. Diversity and dynamics of a north Atlantic coastal Vibrio community. Appl Environ Microbiol. 2004;70:4103–4110. doi: 10.1128/AEM.70.7.4103-4110.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toranzo E, Magariños B, Romalde JL. A review of the main bacterial fish diseases in mariculture systems. Aquaculture. 2005;246:37–61. doi: 10.1016/j.aquaculture.2005.01.002. [DOI] [Google Scholar]
- Verschuere L, Rombaut G, Sorgeloos P, Verstraete W. Probiotic bacteria as biological control agents in aquaculture. Microbiol Mol Biol Rev. 2000;64:655–671. doi: 10.1128/MMBR.64.4.655-671.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wulff EG, Mguni CM, Mansfeld-Giese K, Fels J, Lübeck M, Hockenhull J. Biochemical and molecular characterization of Bacillus velezensis, B. subtilis and B. pumilus isolates with distinct antagonistic potential against Xanthomonas campestris pv. campestris. Plant Pathol. 2002;51:574–584. doi: 10.1046/j.1365-3059.2002.00753.x. [DOI] [Google Scholar]
- Zhou Q, Li K, Jun X, Bo L. Role and functions of beneficial microorganisms in sustainable aquaculture. Bioresour Technol. 2009;100:3780–3786. doi: 10.1016/j.biortech.2008.12.037. [DOI] [PubMed] [Google Scholar]
- Ziolo KJ, Jeong HG, Kwak JS, Yang S, Lavker RM, Satchell KJF. Vibrio vulnificus biotype 3 multifunctional autoprocessing RTX toxin is an adenylate cyclase toxin essential for virulence in mice. Infect Immun. 2014;82:2148–2157. doi: 10.1128/IAI.00017-14. [DOI] [PMC free article] [PubMed] [Google Scholar]