

Abstract
With the discovery of translesion synthesis DNA polymerases, great strides have been made in the last two decades in understanding the mode of replication of various DNA lesions in prokaryotes and eukaryotes. A database search indicated that approximately 2000 articles on this topic have been published in this period. This includes research involving genetic and structural studies as well as in vitro experiments using purified DNA polymerases and accessory proteins. It is a daunting task to comprehend this exciting and rapidly emerging area of research. Even so, as the majority of DNA damage occurs at 2′-deoxyguanosine residues, this perspective attempts to summarize a subset of this field, focusing on the most relevant eukaryotic DNA polymerases responsible for their bypass.
1. Introduction
A large fraction of all DNA damages are formed at 2′-deoxyguanosines (dGs).1 Of the four common nucleosides in DNA, oxidation takes place most easily at dG residues, giving rise to a variety of products including 7,8-dihydro-8-oxo-2′-deoxyguanosine (8-oxo-dG) (Figure 1).2,3 8-Oxo-dG is more susceptible to oxidation than dG, and it generates a number of secondary oxidation products.4,5 The pathway leading to 8-oxo-dG is believed to involve a C8-hydroxyl radical, which also forms Fapy·dG (Figure 1).6
Figure 1.

Structures of small and common ring-opened dG lesions.
Methylating and ethylating agents preferentially react at N7, but they also alkylate O6 of dG, and the fraction of alkylation at O6 increases with “harder” electrophiles.7,8 The N7-Methyl-dG (N7-Me-dG) adduct is unstable, which either depurinates to form an abasic site or undergoes ring opening to generate MeFapy·dG (Figure 1). Interestingly, a vast majority of the bulky adducts are formed at either N7 or the exocyclic N2 position of dG. The unstable dG-N7 adducts formed by the metabolically activated aromatic amines and nitro compounds rearrange to stable dG-C8 adducts,9 while minor adducts at the N2 position of dG have also been isolated.10,11 In contrast, a majority of the metabolically activated epoxides of polycyclic aromatic hydrocarbons (PAHs) form the dG-N2 adducts as the major products.12 Metabolically activated aflatoxin B1, however, forms the primary dG-N7 adduct, which undergoes ring opening to a stable formamidopyrimidine (Fapy) derivative.13 Like the PAH epoxides, the antitumor agent mitomycin C (MC) containing an aziridine ring preferentially forms the dG-N2 adducts.14 The genotoxicity and mutagenicity of many of these adducts have been investigated in prokaryotic and eukaryotic cells for the last three decades. Replication of these DNA lesions do not follow a unifying mechanism, and each lesion exhibits a characteristic mutational spectrum. However, increasingly it became clear that the mutational signature of a DNA lesion is directly related to the identity of the DNA polymerase(s) that bypass it and the mechanism of its nucleotide insertion and extension, though additional factors such as DNA sequence context play a role as well.
A human cell contains at least 17 different DNA polymerases (pols) to perform different functions of the cell, which include DNA replication of undamaged and damaged DNA, replication as part of various DNA repair pathways, recombination, telomere maintenance, and other tasks.15,16 On the basis of sequence homology, pols have been divided into seven families (A, B, C, D, X, Y, and RT), of which C family pols were only found in prokaryotes. In eukaryotes, the B-family enzymes are important since pol ε and pol δ of this family carry out a large fraction of nuclear DNA replication, whereas pol α is involved in initiation and priming. These three pols are essential for DNA replication in eukaryotes. In the current model of DNA replication, pol ε carries out a majority of leading strand DNA replication of the undamaged genome, whereas pol δ primarily replicates the lagging strand. However, this model has recently been challenged, and data supporting the involvement of pol δ in both leading and lagging strand replication have been presented.17−19
The discovery of translesion synthesis (TLS) DNA pols in the 1990s invigorated the area of replication of DNA lesions, and since then, numerous articles have been published on the catalytic and noncatalytic roles of these pols in the context of damaged DNA replication.20 Lesion bypass is carried out principally by the Y-family pols, although X- and B-family pols are also frequently involved. Like the replicative pols, these pols possess right-handed topology with the active site located in the “palm” domain, except that the active site is much larger in order to accommodate the DNA lesions. Unlike the replicative pols, in which the finger and thumb domains ensure correct pairing with the incoming nucleotide, they are shorter and make little interaction with the template and the incoming dNTP, thereby reducing the pol’s ability to discriminate the accuracy of nucleotide insertion. A little finger domain assists to stabilize the Y-family pol on DNA. An important aspect of the Y-family pols and pol ζ of the B-family is that they lack the 3′–5′ proofreading function, making them error-prone but letting them carry out TLS.
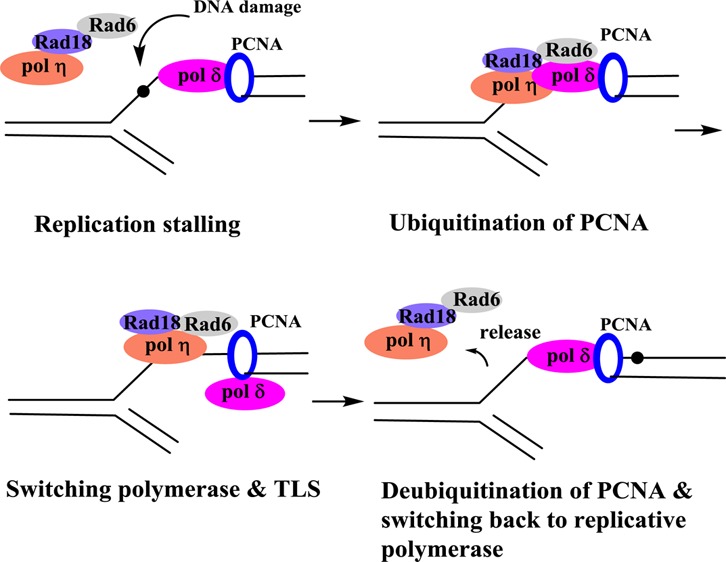
From the perspective of TLS, DNA lesions can be broadly divided between weak and strong replication blocks. Small DNA lesions such as O6-Me-dG and 8-oxo-dG stall but do not completely stop DNA synthesis, whereas most bulky DNA lesions, such as the adducts formed by the PAHs and aromatic amines, are much stronger replication blocks and require the assistance of TLS pols to bypass. The current paradigm on TLS is as follows. When a processive DNA pol encounters a blocking lesion, the pol dissociates, and a TLS pol binds to the DNA and incorporates a dNTP opposite the lesion. In many cases, the same pol continues elongation for a few more bases before dissociating, while in other occasions this TLS pol is replaced by another TLS pol for the elongation steps. TLS pols exhibit higher rate of errors on unmodified templates and are also highly error-prone when bypassing most DNA lesions. Soon after bypassing the lesion, the processive pol returns to continue DNA synthesis. However, the actual process of pol switching is still speculative, and many related questions remain unanswered at the present time.21−23 During cellular replication, the fork utilizes many proteins, including DNA pol, helicase, and single strand binding proteins, to name a few. A prerequisite for TLS is the Rad6/Rad18-mediated monoubiquitination of proliferating cell nuclear antigen (PCNA) at the highly conserved lysine K164.24−26 Y-family pols contain ubiquitin-binding domains that confer affinity to monoubiquitinated PCNA.27−31 In mammalian cells, a Rad18 orthologue is involved in PCNA ubiquitination.32 In addition, two human Rad5-related proteins, SNF2 histone-linker PHD-finger RING-finger helicase (SHPRH) and helicase-like transcription factor (HLTF), transform monoubiquitinated PCNA into the polyubiquitinated form.30,33−35 Additional DNA damage response pathways, including SHPRH/HLTF-mediated template switching, also depend on PCNA ubiquitination. So, when replication by pol δ or pol ε is blocked by a DNA lesion, PCNA is monoubiquitinated by the Rad6-Rad18 protein complex and promotes the switch to a TLS pol at the damage site (Figure 2). Evidently, the activity of the TLS pols must be tightly regulated so that they only gain access to genomic DNA when there is DNA damage. Indeed, regulation of TLS also involves ubiquitination of the TLS pols. For example, monoubiquitination of pol η inhibits its interaction with PCNA, thereby preventing its activity on undamaged DNA, but monoubiquitination is downregulated by the DNA damaging agents.36,37 This mechanism allows optimal availability of nonubiquitinated and active pol η following DNA damage. Post-translational regulation of these proteins is an area where much emphasis has recently been placed.36−39 Despite the predominant role of these bypass pols in TLS, it is also worth noting that there is evidence that replicative pols (such as pol δ) may take part in some TLS events.40
Figure 2.
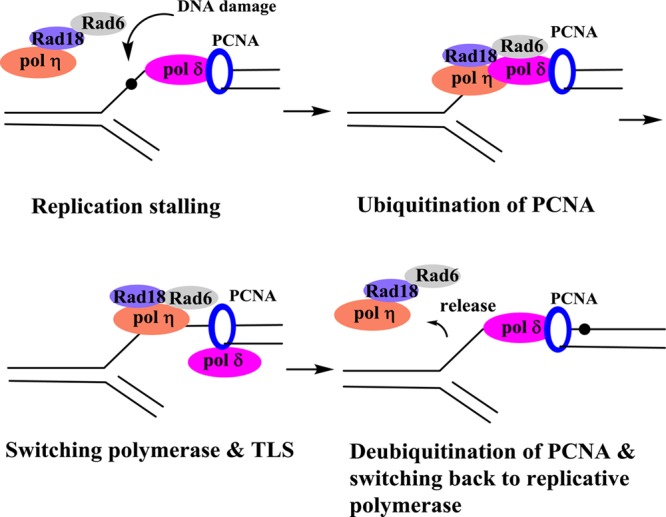
Abridged TLS scheme using pol η as an example of the TLS polymerase.
2. TLS of Small and Ring-Opened dG Lesions
2.1. 8-Oxo-dG
Oxidative stress generates many different DNA lesions, but 8-oxo-dG is the most widely studied DNA lesion formed by reactive oxygen species such as hydroxyl radicals (Figure 1).2 8-Oxo-dG does not strongly block DNA synthesis in eukaryotic cells, as reflected by the number of progeny derived from replication of singly adducted vectors.41,42 Crystallographic studies using a high fidelity pol indicated that 8-oxo-dG adopts syn conformation at the preinsertion stage and pairs preferentially with adenine via Hoogsteen base pairing in the pol active site.43 However, in vitro experiments using yeast pol δ showed that only about 10% TLS takes place in the absence of any accessory proteins.44 Even in the presence of PCNA, steady-state reactions of calf-thymus DNA pol δ were decreased by a factor of 12 for dATP and dCTP incorporation opposite 8-oxo-dG. The major DNA pols in mammalian cells, pol α, pol δ, and pol ε extend an 8-oxo-G:A pair more efficiently than the correct 8-oxo-G:C pair.44,45In vitro experiments showed that pol ζ is inefficient in nucleotide insertions opposite 8-oxo-dG, but it can efficiently extend from the nucleotides inserted opposite it by pol δ.46 Yet, in human cells TLS of 8-oxo-dG is largely error-free (mutation frequency (MF) ∼1% in duplex DNA and 4–20% in single stranded DNA).47−49 Several repair systems, including base excision repair and mismatch repair, excise 8-oxo-dG from duplex DNA, justifying low MF, but most repair systems are inefficient in 8-oxo-dG repair in single-stranded DNA.50−52 One might wonder why the TLS of 8-oxo-dG in single-stranded DNA is mostly error-free. The answer came from in vitro and cellular experiments, which determined a crucial role of pol λ, an X-family enzyme, in 8-oxo-dG bypass. The preference for dCTP incorporation over either dATP or dGTP incorporation opposite 8-oxo-dG is 12-fold by pol λ.45,53 However, it is remarkable that in the presence of the accessory proteins, human PCNA and replication protein A (RPA), correct incorporation of dCTP over other dNTPs opposite 8-oxo-dG increased to 1200-fold by pol λ.45,53 In a similar vein, PCNA and RPA increased the preference for dCTP over dATP or dGTP incorporation opposite 8-oxo-dG by pol η from 2.5-fold to 68-fold.45,53 On the basis of these results and additional data from mouse embryonic fibroblasts and human cell lines, it was suggested that the switch from pol δ involved pol λ and not pol β or pol η since mutations by 8-oxo-dG increased considerably in pol λ knockout or knockdown cells.53,54 In yeast chromosome, however, the switch to pol η, which replicates 8-oxo-dG with an accuracy of 94%, was reported.55 In the absence of pol η, accurate replication drops to 40%. DNA pol δ-interacting protein 2 (PolDIP2, also known as PDIP38) physically interacts with pol λ and increases the efficiency of elongation past 8-oxo-dG by pol λ, suggesting an important role of this protein in pol switch and elongation steps during TLS.56 If pol λ (in the presence of the accessory proteins) were the only pol that bypasses 8-oxo-dG, MF would have dropped to less than 1%. The 4–20% MF, which depends on the DNA sequence context and the type of assay, in single-stranded DNA indicates, however, that in addition to pol λ, other pols bypass the lesion. In human embryonic kidney (HEK) 293T cells, depending on the DNA sequence context, we observed 38–50% increase in mutations induced by 8-oxo-dG, upon knockdown of pol λ.57 It is interesting that G → T mutations were not significantly increased in pol λ knockdown cells. The increase in mutations was primarily due to an increase in dinucleotide deletions, involving the lesion and one of its neighboring bases. Others have also reported targeted one-base or small deletions in the absence of pol λ.53 It appears, therefore, that pol λ prevents these deletions induced by 8-oxo-dG. However, it is unclear which pol is causing the deletion mutations. In addition to the DNA pols, an additional factor is the participation of a homologue of MutY glycosylase. MutY human homologue (MUTYH) shares 41% and 79% of sequence homology to its E. coli counterpart MutY and mouse homologue mMYH, respectively.58 MutY removes adenine from the 8-oxo-G:A mispair, which allows another chance to incorporate C opposite 8-oxo-dG by a pol.59 In a study in human lymphoblastoid cells, replication of 8-oxo-dG generated ∼14% mutants, including 6% G → T and 2% targeted single-base deletions.60 Overexpression of MUTYH reduced the G → T mutations, but the deletions remained unaffected, which also suggests the role of an unidentified DNA pol in the 8-oxo-dG induced deletions. While the role of these deletions in human diseases is unknown, inherited variants of MUTYH in a family affected by colon cancers show a pattern of high G:C → T:A mutations implicating a role of unrepaired 8-oxo-dG lesions in human cancer.61
2.2. Fapy·dG and MeFapy·dG
Fapy·dG (Figure 1) is generated at comparable levels under many conditions to 8-oxo-dG, but only a limited number of biological studies have been conducted with this lesion.6 Bypass efficiency of purine-ring opened Fapy·dG is slower than 8-oxo-dG.62 Like 8-oxo-dG, Fapy·dG is mutagenic inducing predominantly G → T transeversions in mammalian cells.48,57 However, the MF is highly dependent on the DNA sequence context. For example, the MF of Fapy·dG in the TG*T sequence is significantly higher than when it is located in the TG*A sequence in both simian (COS-7) and human embryonic (293T) kidney cells.48,57 In human cells, in some sequence contexts Fapy·dG is more mutagenic than 8-oxo-dG, while in others the opposite is true. The major difference between the two lesions, however, is that knockdown of pol λ reduced the level of G → T mutations induced by Fapy·dG, in contrast to an increase in MF for 8-oxo-dG.57 This suggests that pol λ is involved in a significant fraction of Fapy·dG induced G → T mutations, whereas it carries out error-free bypass of 8-oxo-dG. It is interesting, however, that the level of small deletions increases upon replication of either 8-oxo-dG or Fapy·dG in human cells in which pol λ was knocked down. Unlike 8-oxo-dG, which adopts syn conformation to pair with adenine,43 a structural study of the carbocyclic analogue of Fapy·dG by a high fidelity pol (Bst pol I) showed that the lesion maintains its anti conformation of the glycosidic bond during both error-free and error-prone replication.63
Most biological assays indicate that N7-Me-dG is not mutagenic but that its ring-opened derivative MeFapy·dG (Figure 1) is mutagenic.7,64−68In vitro assays showed that the MeFapy·dG is a strong block to the high fidelity replicative DNA polymerases at both the insertion and the extension steps.67 However, hpol η and hpol κ as well as hRev1 and ypol ζ together can carry out facile TLS. With hpol η and hpol κ, the predominant replication product is the error-free extension product, whereas hRev1 and ypol ζ together accomplish entirely error-free TLS. Up to 29% mutagenic TLS, including each of the targeted base changes and one-nucleotide deletion products, were identified from replication products generated by hpol η and hpol κ. In COS-7 cells, MeFapy·dG induces G → T mutations and single and dinucleotide deletions as do 8-oxo-dG and Fapy·dG.68 However, cellular experiments in human cells analogous to Fapy·dG have not been performed with MeFapy·dG, and it would certainly be interesting to compare the replicative properties of Fapy·dG with MeFapy·dG using the same approach.
2.3. O6-Methyl-dG
O6-Methyl-dG (O6-Me-dG) is one of the first mutagenic DNA lesions identified as a result of DNA methylation (Figure 1).69 It is highly mutagenic but is quickly repaired in a cell by multiple repair systems.70−72 A great deal of circumstantial evidence indicates that it plays a role in the etiology of human cancer.73−76 Using an intrachromosomal probe, ∼19% G → A mutations were detected after replication of a site-specific O6-Me-dG in Chinese Hamster Ovary cells deficient in the repair enzyme O6-alkylguanine-DNA alkyltransferase, but in repair proficient cells, mutation frequency dropped to an ∼1% level.77 Like 8-oxo-dG, it allows partial bypass of several purified DNA polymerases, but pol δ is only slightly inhibited in vitro and inserts dCTP and dTTP equally well opposite O6-Me-dG.78 However, pol α is strongly blocked one base before O6-Me-dG.79O6-Me-dG also is a strong but not absolute block of human pol β, and even though hpol β inserts dTTP more efficiently than dCTP opposite the lesion, it preferentially extends the correct O6-Me-G:C pair.80 In the absence of accessory proteins, the human TLS pol ι and pol κ produce mainly one-base incorporation products opposite this lesion, but hpol η is much more efficient.78 Steady-state kinetic analysis showed similar efficiencies of insertion of dCTP and dTTP opposite O6-Me-dG by hpol η and hpol κ, whereas hpol ι showed a higher preference for dTTP insertion.78 Genetic studies in yeast implicate both pol δ and pol η in the TLS of O6-Me-dG, even though biochemical studies suggest that hpol η is more efficient than hpol δ.81 Similar to 8-oxo-dG, in yeast pol ζ is very inefficient at inserting nucleotides opposite O6-Me-dG, but it can efficiently extend from the nucleotides inserted opposite it by pol δ. As a result, the most efficient bypass can be accomplished in vitro when both pol δ and pol ζ were used for the TLS of templates containing O6-Me-dG.46
3. TLS of Bulky dG Lesions
Even though this perspective is focused on dG lesions, it may be pertinent to mention investigations that established a specialized role of pol η in efficient and error-free bypass of UV light-induced cis-syn cyclobutane pyrimidine dimers (CPDs). This is due to pol η’s unique ability to accommodate both pyrimidine residues of this bulky lesion in its active site and perform accurate and efficient TLS.82,83 Pol κ and pol ζ, on the other hand, provide an alternate, albeit highly error-prone, pathway of TLS of CPDs. In the absence of pol κ and pol ζ, TLS of CPDs carried out by pol η is error-free, and mutations decrease to the background level.84 Mutations in this gene (POLH) result in XPV, a variant type of the genetic disease, xeroderma pigmentosum, which is characterized by extreme sensitivity to UV light.85−87 No other DNA pol exhibits such a precise and dedicated role, but the main characteristics of the other bypass pols have been established. One example is the ability of pol κ in the error-free bypass of dG-N2 lesions (discussed later). The enlarged active site of pol η allows it to accommodate even the cisplatin-derived large intrastrand N7–Pt–N7 cross-linked two guanine residues and to bypass accurately.88,89 Crystal structure analysis showed that to allow the lesion to fit in its active site, pol η goes through a backbone rearrangement to stabilize the lesion and incorporate dCTP opposite the two guanines.90 However, it also shows that the rigid backbone of the ternary complex with pol η does not allow extension, which necessitates another TLS pol such as pol ζ to extend it.
3.1. Aflatoxin B1
The potent hepatocarcinogen aflatoxin B1 (AFB1) forms two major DNA adducts upon metabolic activation of AFB1 to AFB1-8,9-epoxide by the liver cytochrome P450 enzymes (Figure 3).91,92 The primary DNA adduct, AFB1–N7-dG, is formed at the N7 position of dG.13,93−95 This adduct is chemically unstable due to the positive charge at N7, which can undergo either spontaneous depurination to generate abasic sites or ring opening to form AFB1–Fapy-dG (Figure 3). Both these adducts are mutagenic in simian kidney (COS-7) cells when the adduct is located in a TTG*AA sequence, but AFB1–Fapy-dG induces 97% mutations compared to 45% mutations by the AFB1–N7-dG adduct.96,97 Both AFB1–N7-dG and AFB1–Fapy-dG predominantly induce G → T transversions. Interestingly, in vitro TLS assays showed that pol ζ bypasses AFB1–N7-dG in an error-free manner, whereas it is responsible for the erroneous bypass of AFB1–Fapy-dG. For AFB1–N7-dG, pol κ appears to be involved in the mutagenic bypass. Because of the importance of these adducts in human cancer, additional structural, genetic, and in vitro studies on the two DNA adducts in the future would certainly be of significant interest.
Figure 3.

Aflatoxin B1, its exo epoxide, and the major dG adducts.
3.2. Benzo[a]pyrene
PAHs are ubiquitous in our environment, and many of them, notably those with a “bay” or “fjord” region, are highly mutagenic and carcinogenic.12 The most extensively studied PAH is benzo[a]pyrene (BP), an extremely carcinogenic chemical, which upon metabolic activation binds to DNA, predominantly at the N2 position of dG (Figure 4). BP is metabolized by the mammalian monooxygenase enzymes to form the diastereomeric anti- and syn-benzo[a]pyrene 7,8-dihydrodiol-9,10-epoxide (BPDE). The metabolically activated (+)-anti BPDE is a potent mutagen and the most tumorigenic metabolite of BP. It is believed to be the ultimate carcinogenic form that leads to trans- and cis-dG-N2 adducts (Figure 4). The principal mutation in mammalian cells induced by the major dG adducts of BP is G:C → T:A transversion.98−100 BP adducts are strong blocks of replication by replicative pols, but the TLS pols can bypass them at varying efficiencies. In vitro studies using either hpol ι or hpol η showed that the BPDE dG adducts allow slow bypass, which results in a high frequency of nucleotide misincorporations.101,102 In yeast, however, (±)-anti-BPDE mutagenesis requires pol ζ and partially involves pol η, but pol η mainly contributes to deletions and insertions of 1–3 nucleotides.103 In contrast, pol κ performs accurate and reasonably efficient replication of the BPDE dG adducts. The extent of bypass drops, and mutagenesis increases significantly in human and murine cells lacking pol κ.104 Pol κ’s catalytic site, unlike that of pol η, can only accommodate one Watson–Crick base pair. However, it is capable of TLS of many dG-N2 adducts, including the DNA adducts formed by BP.104−106 Specifically, for the (+)-trans-anti-dG-N2-BPDE adduct, genetic, in vitro kinetics, and structural studies show that pol κ performs efficient and accurate TLS. For the mutagenic TLS, genetic evidence suggests that a non-Y family pol inserts a wrong nucleotide (dATP or dTTP) opposite the adduct but that extension is performed cooperatively by pol ζ and Rev1.107 It was postulated that Rev1 recruits pol ζ via interaction with Rev7.
Figure 4.

Metabilic activation and the major DNA adducts formed by benzo[a]pyrene.
Crystal structure analyses of the (+)-trans-anti-dG-N2-BPDE adduct showed that the active site of pol κ is opened up at the minor groove side of the primer–template complex allowing accommodation of the bulky BPDE-dG adduct.108 The amino acid residues of the protein in the minor groove side of DNA stabilizes the hydrophobic BPDE ring and maintains Watson–Crick base pairing with an incoming dCTP for accurate replication.
Pol κ also bypasses many other dG-N2 adducts accurately and efficiently, which includes N2-(1-carboxyethyl)-dG and N2-furfuryl-dG as well as much bulkier adducts formed by IQ and mitomycin C (discussed in the next section).109−112
3.3. dG-N2 Adducts of IQ and Mitomycin C
We have recently studied the minor, albeit persistent, dG-N2 adduct (Figure 5) formed by the carcinogen 2-amino-3-methylimidazo[4,5-f]quinoline (IQ), a heterocyclic aromatic amine formed during high temperature cooking of meat,113−115 and two dG-N2adducts (Figure 6) formed by the antitumor agent, mitomycin C (MC), and its metabolite, 2–7-diaminomitosene (2,7-DAM).14,116,117
Figure 5.

Structure of IQ and its dG adducts.
Figure 6.

Structures of the dG-N2 adducts formed by mitomycin C and its metabolite 2,7-diaminomitosene.
The dG-N2-IQ adduct was studied in the three different guanines of the NarI restriction site (5′-G1G2CG3CC-3′). As shown in Table 1, in HEK293T cells MF increases upon knockdown of only pol κ, whereas knockdown of pol η, pol ι, pol ζ, or Rev1 results in a reduction in MF. The greatest reduction in MF occurred when pol η, pol ζ, and Rev1 were concurrently knocked down. This suggests that pol κ is involved in error-free bypass of the dG-N2 adduct formed by IQ, whereas pol η, pol ζ, and Rev1 cooperatively carried out mutagenic TLS.111 Similar results were obtained with the mitomycin C adducts (Table 1), indicating that they also follow analogous mechanisms.112 It was also established that with the increasing bulk of the dG-N2 adducts, the misincorporation frequency of dATP relative to dCTP increases significantly.118 Taken together, there seems to be a predictable pattern of error-free and error-prone TLS of dG-N2 adducts by the TLS pols.
Table 1. Mutation Frequency of dG-N2-IQ (in Three Different Guanines of the NarI Site), dG-N2-MC, and dG-N2-2,7-DAM and Their Change in Percentages upon Knockdown of Specific Pols.
| lesion | MF (%) | % change in MF in pol η-deficient cells | % change in MF in pol κ-deficient cells | % change in MF in pol ι-deficient cells | % change in MF in pol ζ-deficient cells | % change in MF in Rev1-deficient cells | % change in MF in (η, ζ, Rev1) deficient cells |
|---|---|---|---|---|---|---|---|
| dG1-N2-IQ | 22.7 | –21 | +23 | –12 | –16 | –20 | –84 |
| dG2-N2-IQ | 17 | –21 | +18 | –18 | –18 | –21 | –87 |
| dG3-N2-IQ | 11 | –27 | +5 | –15 | –18 | –22 | –90 |
| dG-N2-MC | 18 | –44 | +39 | NDa | –33 | ND | –78 |
| dG-N2-2,7-DAM | 10 | –20 | +50 | ND | –10 | ND | –81 |
ND, not determined.
Exceptions to this rule, however, are the minor groove adducts γ-hydroxy-1,N2-propano-dG and trans-4-hydroxy-2-nonenal-dG, in which case pol κ is inefficient in nucleotide insertion opposite the lesion, but it efficiently acts as an extender.119,120 In both these cases, pol ι can insert dCTP opposite the lesions but is inefficient in extending the G*:C pair. In contrast, pol κ is unable to insert a nucleotide opposite these lesions, but it can extend the G*:C pair. Thus, the sequential act of pol ι and pol κ promotes efficient and error-free TLS of these lesions. It is noteworthy that these are cyclic adducts with a covalent bond with N1 in addition to N2 of dG, suggesting that pol κ’s ability to insert a nucleotide is impaired for adducts with dual linkages.
3.4. N-Acetyl-2-aminofluorene
One of the most extensively studied DNA adduct is dG-C8-AAF (Figure 7), the dG-C8 adduct formed by N-acetyl-2-aminofluorene (AAF), which induces frameshift mutation in bacteria and human cells, but in simian kidney (COS-7) cells, when the adduct is placed in a single stranded plasmid, it causes largely G → T mutations.121−124 However, in a subsequent study, also in COS-7 cells but in duplex DNA, at the third guanine of 5′-GGG-3′ and 5′-GGCGCC-3′ (NarI site), −1 and −2 frameshift mutations, respectively, were detected.125 The frameshifts also occurred in human cell-free extracts. The frameshift mutations at the 5′-GGG-3′ sequence are dependent on pol η but not pol ι or pol ζ. Furthermore, this pol η-mediated erroneous pathway requires Rad18 and ubiquitination of PCNA. On the other hand, TLS is only partially dependent on pol η and Rad18 when the adduct is situated at the NarI site. This indicates that the same adduct may follow different mechanisms for mutagenesis in different sequence contexts.
Figure 7.

Structures of the dG-C8 adducts formed by N-acetyl-2-aminofluorene, 1-nitropyrene, and 3-nitrobenzanthrone.
The mechanism of both −1 and −2 frameshifts was suggested to follow a slipped frameshift intermediate,126,127 and while most pols are inefficient in extending such an intermediate, pol η can extend them, albeit slowly.128 In duplex DNA, dG-C8-AAF is known to rotate the guanine base to syn conformation, in contrast to an overwhelming anti conformation of an unmodified dG.129,130 Biophysical and computational studies indicate that syn conformation in a base-displaced intercalated structure of the dG adduct allows formation of stable slipped intermediates.126,130,131 Such intermediates, upon elongation, would cause frameshift mutations, the major types of mutations detected in bacteria11,122,132 and occasionally in mammalian cells (or cell-free extracts).125,133 The role of pol η in bypassing misaligned adducts has been explored, which showed that depending on the base sequence, a cytosine inserted opposite the dG-C8 lesion slips to generate a −1, −2, or −3 frameshift intermediate that pol η can continue to replicate, in spite of a bulge.128 In a crystal structure study, however, pol η was able to incorporate dCTP opposite the dG-C8-AAF adduct, in which TLS occured without rotation of the adduct into the anti conformation, and only one hydrogen bond was formed between the lesion and dCTP.134 This structural investigation recognized pol η’s ability to perform error-free replication of dG-C8-AAF, in addition to its propensity to carry out frameshifts.
3.5. dG-C8 Adducts Formed by IQ, 3-Nitrobenzanthrone, and 1-Nitropyrene
Like the dG-N2 adducts, the roles of TLS DNA pols in bypassing the C8-dG adduct (dG-C8-IQ) (Figure 5) formed by IQ were explored at the G1-, G2-, or G3-positions of the NarI recognition sequence after replication in HEK293T cells.135 MF was the highest (50%) when the adduct was placed at G3, compared to 18% and 24% MF when the adduct was located at G1 and G2, respectively, inducing mainly G → T transversions at each site. MF of dG-C8-IQ was reduced in varying degrees upon siRNA knockdown of pol κ, pol ι-, pol ζ-, or Rev1-knockdown cells (Table 2), indicating that these pols are involved in error-prone synthesis of this adduct. In contrast, MF was increased by 8–26% in pol η knockdown cells, suggesting that pol η bypasses the lesion accurately.
Table 2. Mutation Frequency of dG-C8-IQ (in Three Different Guanines of the NarI Site) and dG-C8-3-ABA and Their Change in Percentages upon Knockdown of Specific Pols.
| lesion | MF (%) | % change in MF in pol η-deficient cells | % change in MF in pol κ-deficient cells | % change in MF in pol ι-deficient cells | % change in MF in pol ζ-deficient cells | % change in MF in Rev1-deficient cells | % change in MF in (κ, ζ, and Rev1)-deficient cells |
|---|---|---|---|---|---|---|---|
| dG1-C8-IQ | 17.8 | +13 | –43 | –13 | –6 | –39 | –93 |
| dG2-C8-IQ | 24 | +8 | –68 | –31 | –50 | –58 | –99 |
| dG3-C8-IQ | 50 | +26 | –36 | –18 | –26 | –38 | –96 |
| dG-C8-3-ABA | 14 | –39 | +15 | –29 | +60 | –61 | a |
Largest % change was noted with pol η and pol κ simultaneous knockdown, which gave 70% reduction in MF.
Upon simultaneous knockdown of pol κ, pol ζ, and Rev 1, a synergy was observed in that MF was reduced by more than 90% in each case (Table 2). In vitro experiments using yeast pol ζ confirmed that it can extend the G3*:A pair more efficiently than the G3*:C pair, although it is inefficient at nucleotide incorporation opposite dG-C8-IQ. It is, therefore, conceivable that pol κ and pol ζ cooperatively carry out the majority of the error-prone TLS of dG-C8-IQ, whereas Rev1 may play a noncatalytic role in assembling the TLS pols. By contrast, pol η is involved mostly in its error-free bypass. Similar experiments have also been conducted with dG-C8–3-ABA,136 the major DNA adduct formed by the carcinogen 3-nitrobenzanthrone (3-NBA) (Figure 7).137,138 Like dG-C8-IQ, dG-C8–3-ABA induces G → T as the major type of mutations in human cells.136 However, the polymerase knockdown results are different. Pol η and pol κ were found to be the major contributors of the mutagenic TLS of dG-C8–3-ABA since MF dropped by 70%, when these pols were simultaneously knocked down, although MF actually increased upon knockdown of pol κ alone. In contrast, pol ζ is involved in the error-free bypass of the lesion since MF increased by 60% in pol ζ knockdown cells. A recent in vitro presteady state kinetic investigation showed that hpol η and hpol κ efficiently bypassed a site-specifically placed dG-C8–3-ABA, whereas hpol ι and hRev1 were severely stalled by the lesion.139 Crystal structure analysis of dG-C8–3-ABA at the insertion stage of hpol η showed that the adduct is wedged at the hydrophobic cleft in the active site in anti conformation stabilized by a hydrogen bond between the C8 amino group and the phosphate, while the 2′-deoxyribose adopts C3′-endo pucker.140 This structure provides a model for an accurate but slow bypass of the adduct by pol η. The structure of an erroneous bypass of dG-C8–3-ABA by a pol is yet to be solved. We postulate that both pol κ and pol ζ conduct error-free TLS of dG-C8–3-ABA. However, pol κ also extends mispairs generated by incorporation of dATP by pol η opposite the adduct. It is noteworthy that single-nucleotide incorporation opposite a dG-C8–3-ABA lesion catalyzed by hpol η in vitro showed that at short reaction time frames incorporation of dCTP is greater than dATP but that with longer time incorporation of these two nucleotides becomes comparable.140 Rev1 likewise is important for mutagenesis, as reflected by 60% reduction in MF upon Rev1 knockdown, but as with dG-C8-IQ, it probably plays a noncatalytic role by physically interacting with the other two Y-family pols. The noncatalytic role of Rev1 was indicated by its inability to bypass the lesion in vitro. Therefore, the C8-dG adducts dG-C8-IQ and dG-C8–3-ABA do not behave the same way with different polymerases.
As mentioned earlier on the mechanism of frameshift mutations induced by dG-C8-AAF, many bulky adducts formed at the C8 position of dG, such as dG-C8-IQ and dG-C8–3-ABA, rotate the base to syn conformation, which is believed to play a structural role in frameshift mutations observed in bacteria.141,142 More frequently in mammalian cells, however, these adducts induce base substitutions.143,144 Since these purine lesions rotate to syn conformation, one can anticipate a role of pol ι in bypassing them, as this enzyme uses Hoogsteen base pairing to select the incoming nucleotide.145 Pol ι can bypass only small dG-N2 adducts since N2 is oriented toward the major groove, and rotation to syn is inhibited for bulky dG-N2 adducts. In contrast, bulky dG-C8 adducts can be accommodated in the pol ι active site more efficiently. An example of pol ι’s potential involvement in dG-C8 adduct bypass is its interaction with dG-C8-AP, the major adduct formed by the environmental carcinogen, 1-nitropyrene (1-NP) (Figure 7). dG-C8-AP, like the other dG-C8 adduct mentioned earlier, induces predominantly G → T mutations in simian and human embryonic kidney cells.144 The adduct, as other bulky dG-C8 adducts, exists in syn conformation in a base-displaced intercalated solution structure.146,147 Replication of dG-C8-AP stalls when in vitro bypass is conducted by the TLS pols. Of the human TLS pols, hpol η is most proficient in bypassing it in vitro, but hpol κ and hpol ι can incorporate a nucleotide opposite the lesion.148,149 Crystal structure analyses showed that dCTP incorporation opposite dG-C8-AP forces the adduct to rotate to the anti conformation to avoid steric hindrance at the minor groove side.150 However, this structure inhibits further extension, due to a clash with the little finger domain of the enzyme. In contrast, the adduct can maintain syn conformation when dATP is inserted, in which the adenine is stacked above the pyrene ring intercalated in the helix. This structure allows further extension. Therefore, error-prone replication of dG-C8-AP potentially may occur by two TLS pols, with pol ι being involved in the insertion stage. Additional genetic evidence will be required to validate this pathway. Another dG-C8 adduct, (5′S)-8,5′-cyclo-dG, a cyclic DNA adduct containing a covalent bond between C8 of guanine and 5′ C of 2-deoxyribose, was investigated in human cells, which showed that pol η, pol ι, and pol ζ but not pol κ are involved in TLS.151
Unlike the dG-N2 adducts, therefore, a pattern for TLS of the dG-C8 adducts could not be determined. For example, pol ζ is involved in extension of the correct pair of the dG-C8–3-ABA, whereas it extends the wrong pair with dG-C8-IQ.135,136 Studies on additional dG-C8 adducts might give us a clue as to why they fail to follow a unifying mechanism of TLS.
4. Concluding Comments
TLS of various DNA damages have been conducted principally by three complementary approaches. Genetic studies in repair and replication competent cells provide data on the outcome of the damage, and a comparison of these in genetically altered cells (including knockout or knockdown of specific genes) has been employed to investigate the role of each TLS pol. In vitro experiments using purified pols and accessory proteins elucidate how each pol can deal with the DNA damage, whereas structural and computational studies give a more intimate snapshot of the lesion bypass. Each approach has its limitations, and consequently, combined approaches are essential to comprehend the mechanism of TLS of a DNA lesion. Mechanistic information on replication of the DNA lesions is critical to follow the underlying process for the development of cancer, aging, and various other diseases. These fundamental studies are now paving the way to application of the acquired knowledge toward therapeutic application, as inhibiting the activity of some of the TLS pols may enhance the effect of an antitumor agent. As yet, more TLS work has been done with the pols from prokaryotes and archaea than from eukaryotes. It is certain that this dynamic area of research is still in its early stage and will continue to enrich the field of toxicology with many novel findings.
Glossary
Abbreviations
- TLS
translesion synthesis
- PCNA
proliferating cell nuclear antigen
- RPA
replication protein A
- SHPRH
SNF2 histone-linker PHD-finger RING-finger helicase
- HLTF
helicase-like transcription factor
- MF
mutation frequency
- HEK
human embryonic kidney
- 8-oxo-dG
7,8-dihydro-8-oxo-2′-deoxyguanosine
- Fapy
formamidopyrimidine
- Fapy·dG and MeFapy·dG
N6-(2-deoxy-d-erythro-pentofuranosyl)-2,6-diamino-3,4 dihydro-4-oxo-5-formamidopyrimidine) and its 5N-methyl derivative, respectively
- N7-Me-dG
N7-methyl-2′-deoxyguanosine
- O6-Me-dG
O6-methyl-2′-deoxyguanosine
- CPD
cis-syn cyclobutane pyrimidine dimer
- AFB1
aflatoxin B1
- pol
DNA polymerase
- BP
benzo[a]pyrene
- BPDE
BP diol epoxide
- IQ
2-amino-3-methylimidazo[4,5-f]quinoline
- AAF
N-acetyl-2-aminofluorene
- 1-NP
1-nitropyrene
- AP
aminopyrene
- MC
mitomycin C
- 2,7-DAM
2,7-diaminomitosene
- 3-NBA
3-nitrobenzanthrone
- 3-ABA
3-aminobenzanthrone
Biographies
Ashis K. Basu is Professor of Chemistry at the University of Connecticut. He received his Ph.D. (1984) from Wayne State University in Bioorganic Chemistry under the supervision of Larry Marnett and did postdoctoral studies with John Essigmann at MIT. He launched his independent research career in the Chemistry Department at the University of Connecticut in 1990. His research focuses on the initiation phase in the etiology of cancer. Specifically, he investigates the biological consequences of DNA damages caused by nitroaromatic carcinogens, γ-radiation, and the antitumor drug, mitomycin C.
Paritosh Pande studied undergraduate chemistry and biochemistry at University of Mumbai from 1998 to 2003. Subsequently, he investigated the antidiabetic potential of natural products on Wistar rats at Nair Hospital in Mumbai. Fascinated by the chemistry–biology interface, he joined Professor Ashis Basu’s lab at the University of Connecticut in 2005, where he earned his Ph.D. by conducting research on the mutagenicity and repair of DNA lesions in mammalian cells. Later, as a university postdoctoral fellow at the Basu lab, he expanded his research horizons by investigating translesion synthesis of chemical carcinogen-derived DNA lesions and toxicity of nanomaterials in human cells.
Arindam Bose received his Master’s degree in chemistry from the University of Delhi. He then joined Professor Ashis Basu’s research group at the University of Connecticut and earned his Ph.D. in chemistry in 2015 working on DNA damage, mutagenesis, and DNA repair. Currently, he is a postdoctoral research associate at the University of Pittsburgh Cancer Institute, where his major area of research is to determine the mechanism of telomere loss and repair. He also investigates the role of telomere shortening in aging and cancer.
Research in the A.K.B. laboratory was supported by the NIEHS grants ES09127, ES021762, and ES023350.
The authors declare no competing financial interest.
References
- Singer B., and Grunberger D. (1983) Molecular Biology of Mutagens and Carcinogens, Plenum Press, New York. [Google Scholar]
- Evans M. D.; Dizdaroglu M.; Cooke M. S. (2004) Oxidative DNA damage and disease: induction, repair and significance. Mutat. Res. 567, 1–61. 10.1016/j.mrrev.2003.11.001. [DOI] [PubMed] [Google Scholar]
- van Loon B.; Markkanen E.; Hubscher U. (2010) Oxygen as a friend and enemy: How to combat the mutational potential of 8-oxo-guanine. DNA Repair 9, 604–616. 10.1016/j.dnarep.2010.03.004. [DOI] [PubMed] [Google Scholar]
- Candeias L. P.; Steenken S. (2000) Reaction of HO* with guanine derivatives in aqueous solution: formation of two different redox-active OH-adduct radicals and their unimolecular transformation reactions. Properties of G(-H)*. Chem. - Eur. J. 6, 475–484. . [DOI] [PubMed] [Google Scholar]
- Chatgilialoglu C.; D’Angelantonio M.; Kciuk G.; Bobrowski K. (2011) New insights into the reaction paths of hydroxyl radicals with 2′-deoxyguanosine. Chem. Res. Toxicol. 24, 2200–2206. 10.1021/tx2003245. [DOI] [PubMed] [Google Scholar]
- Greenberg M. M. (2012) The formamidopyrimidines: purine lesions formed in competition with 8-oxopurines from oxidative stress. Acc. Chem. Res. 45, 588–597. 10.1021/ar2002182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Basu A. K.; Essigmann J. M. (1990) Site-specifically alkylated oligodeoxynucleotides: probes for mutagenesis, DNA repair and the structural effects of DNA damage. Mutat. Res., Fundam. Mol. Mech. Mutagen. 233, 189–201. 10.1016/0027-5107(90)90162-W. [DOI] [PubMed] [Google Scholar]
- Loechler E. L. (1994) A violation of the Swain-Scott principle, and not SN1 versus SN2 reaction mechanisms, explains why carcinogenic alkylating agents can form different proportions of adducts at oxygen versus nitrogen in DNA. Chem. Res. Toxicol. 7, 277–280. 10.1021/tx00039a001. [DOI] [PubMed] [Google Scholar]
- Humphreys W. G.; Kadlubar F. F.; Guengerich F. P. (1992) Mechanism of C8 alkylation of guanine residues by activated arylamines: evidence for initial adduct formation at the N7 position. Proc. Natl. Acad. Sci. U. S. A. 89, 8278–8282. 10.1073/pnas.89.17.8278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kadlubar F. F. (1994) DNA adducts of carcinogenic aromatic amines. IARC Sci. Publ 199–216. [PubMed] [Google Scholar]
- Purohit V.; Basu A. K. (2000) Mutagenicity of nitroaromatic compounds. Chem. Res. Toxicol. 13, 673–692. 10.1021/tx000002x. [DOI] [PubMed] [Google Scholar]
- Harvey R. G. (2011) Historical Overview of Chemical Carcinogenesis. Current Cancer Res. 6, 1–26. 10.1007/978-1-61737-995-6_1. [DOI] [Google Scholar]
- Hertzog P. J.; Smith J. R.; Garner R. C. (1982) Characterisation of the imidazole ring-opened forms of trans-8,9-dihydro-8,9-dihydro-8-(7-guanyl)9-hydroxy aflatoxin B1. Carcinogenesis 3, 723–725. 10.1093/carcin/3.6.723. [DOI] [PubMed] [Google Scholar]
- Tomasz M.; Chowdary D.; Lipman R.; Shimotakahara S.; Veiro D.; Walker V.; Verdine G. L. (1986) Reaction of DNA with chemically or enzymatically activated mitomycin C: isolation and structure of the major covalent adduct. Proc. Natl. Acad. Sci. U. S. A. 83, 6702–6706. 10.1073/pnas.83.18.6702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burgers P. M.; Koonin E. V.; Bruford E.; Blanco L.; Burtis K. C.; Christman M. F.; Copeland W. C.; Friedberg E. C.; Hanaoka F.; Hinkle D. C.; Lawrence C. W.; Nakanishi M.; Ohmori H.; Prakash L.; Prakash S.; Reynaud C. A.; Sugino A.; Todo T.; Wang Z.; Weill J. C.; Woodgate R. (2001) Eukaryotic DNA polymerases: proposal for a revised nomenclature. J. Biol. Chem. 276, 43487–43490. 10.1074/jbc.R100056200. [DOI] [PubMed] [Google Scholar]
- Ohmori H.; Friedberg E. C.; Fuchs R. P.; Goodman M. F.; Hanaoka F.; Hinkle D.; Kunkel T. A.; Lawrence C. W.; Livneh Z.; Nohmi T.; Prakash L.; Prakash S.; Todo T.; Walker G. C.; Wang Z.; Woodgate R. (2001) The Y-family of DNA polymerases. Mol. Cell 8, 7–8. 10.1016/S1097-2765(01)00278-7. [DOI] [PubMed] [Google Scholar]
- Johnson R. E.; Klassen R.; Prakash L.; Prakash S. (2016) Response to Burgers et al. Mol. Cell 61, 494–495. 10.1016/j.molcel.2016.01.018. [DOI] [PubMed] [Google Scholar]
- Johnson R. E.; Klassen R.; Prakash L.; Prakash S. (2015) A Major Role of DNA Polymerase delta in Replication of Both the Leading and Lagging DNA Strands. Mol. Cell 59, 163–175. 10.1016/j.molcel.2015.05.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burgers P. M.; Gordenin D.; Kunkel T. A. (2016) Who Is Leading the Replication Fork, Pol epsilon or Pol delta?. Mol. Cell 61, 492–493. 10.1016/j.molcel.2016.01.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang W.; Woodgate R. (2007) What a difference a decade makes: insights into translesion DNA synthesis. Proc. Natl. Acad. Sci. U. S. A. 104, 15591–15598. 10.1073/pnas.0704219104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plosky B. S.; Woodgate R. (2004) Switching from high-fidelity replicases to low-fidelity lesion-bypass polymerases. Curr. Opin. Genet. Dev. 14, 113–119. 10.1016/j.gde.2004.02.002. [DOI] [PubMed] [Google Scholar]
- Friedberg E. C.; Lehmann A. R.; Fuchs R. P. (2005) Trading places: how do DNA polymerases switch during translesion DNA synthesis?. Mol. Cell 18, 499–505. 10.1016/j.molcel.2005.03.032. [DOI] [PubMed] [Google Scholar]
- Chuang L. C.; Yew P. R. (2005) Proliferating cell nuclear antigen recruits cyclin-dependent kinase inhibitor Xic1 to DNA and couples its proteolysis to DNA polymerase switching. J. Biol. Chem. 280, 35299–35309. 10.1074/jbc.M506429200. [DOI] [PubMed] [Google Scholar]
- Hoege C.; Pfander B.; Moldovan G. L.; Pyrowolakis G.; Jentsch S. (2002) RAD6-dependent DNA repair is linked to modification of PCNA by ubiquitin and SUMO. Nature 419, 135–141. 10.1038/nature00991. [DOI] [PubMed] [Google Scholar]
- Stelter P.; Ulrich H. D. (2003) Control of spontaneous and damage-induced mutagenesis by SUMO and ubiquitin conjugation. Nature 425, 188–191. 10.1038/nature01965. [DOI] [PubMed] [Google Scholar]
- Haracska L.; Torres-Ramos C. A.; Johnson R. E.; Prakash S.; Prakash L. (2004) Opposing effects of ubiquitin conjugation and SUMO modification of PCNA on replicational bypass of DNA lesions in Saccharomyces cerevisiae. Mol. Cell. Biol. 24, 4267–4274. 10.1128/MCB.24.10.4267-4274.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Acharya N.; Brahma A.; Haracska L.; Prakash L.; Prakash S. (2007) Mutations in the ubiquitin binding UBZ motif of DNA polymerase eta do not impair its function in translesion synthesis during replication. Mol. Cell. Biol. 27, 7266–7272. 10.1128/MCB.01196-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Acharya N.; Yoon J. H.; Gali H.; Unk I.; Haracska L.; Johnson R. E.; Hurwitz J.; Prakash L.; Prakash S. (2008) Roles of PCNA-binding and ubiquitin-binding domains in human DNA polymerase eta in translesion DNA synthesis. Proc. Natl. Acad. Sci. U. S. A. 105, 17724–17729. 10.1073/pnas.0809844105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sood R.; Makalowska I.; Galdzicki M.; Hu P.; Eddings E.; Robbins C. M.; Moses T.; Namkoong J.; Chen S.; Trent J. M. (2003) Cloning and characterization of a novel gene, SHPRH, encoding a conserved putative protein with SNF2/helicase and PHD-finger domains from the 6q24 region. Genomics 82, 153–161. 10.1016/S0888-7543(03)00121-6. [DOI] [PubMed] [Google Scholar]
- Motegi A.; Sood R.; Moinova H.; Markowitz S. D.; Liu P. P.; Myung K. (2006) Human SHPRH suppresses genomic instability through proliferating cell nuclear antigen polyubiquitination. J. Cell Biol. 175, 703–708. 10.1083/jcb.200606145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moldovan G. L.; D’Andrea A. D. (2011) DNA damage discrimination at stalled replication forks by the Rad5 homologs HLTF and SHPRH. Mol. Cell 42, 141–143. 10.1016/j.molcel.2011.03.018. [DOI] [PubMed] [Google Scholar]
- Watanabe K.; Tateishi S.; Kawasuji M.; Tsurimoto T.; Inoue H.; Yamaizumi M. (2004) Rad18 guides poleta to replication stalling sites through physical interaction and PCNA monoubiquitination. EMBO J. 23, 3886–3896. 10.1038/sj.emboj.7600383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Motegi A.; Liaw H. J.; Lee K. Y.; Roest H. P.; Maas A.; Wu X.; Moinova H.; Markowitz S. D.; Ding H.; Hoeijmakers J. H.; Myung K. (2008) Polyubiquitination of proliferating cell nuclear antigen by HLTF and SHPRH prevents genomic instability from stalled replication forks. Proc. Natl. Acad. Sci. U. S. A. 105, 12411–12416. 10.1073/pnas.0805685105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Unk I.; Hajdu I.; Fatyol K.; Hurwitz J.; Yoon J. H.; Prakash L.; Prakash S.; Haracska L. (2008) Human HLTF functions as a ubiquitin ligase for proliferating cell nuclear antigen polyubiquitination. Proc. Natl. Acad. Sci. U. S. A. 105, 3768–3773. 10.1073/pnas.0800563105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Unk I.; Hajdu I.; Blastyak A.; Haracska L. (2010) Role of yeast Rad5 and its human orthologs, HLTF and SHPRH in DNA damage tolerance. DNA Repair 9, 257–267. 10.1016/j.dnarep.2009.12.013. [DOI] [PubMed] [Google Scholar]
- Bienko M.; Green C. M.; Crosetto N.; Rudolf F.; Zapart G.; Coull B.; Kannouche P.; Wider G.; Peter M.; Lehmann A. R.; Hofmann K.; Dikic I. (2005) Ubiquitin-binding domains in Y-family polymerases regulate translesion synthesis. Science 310, 1821–1824. 10.1126/science.1120615. [DOI] [PubMed] [Google Scholar]
- Bienko M.; Green C. M.; Sabbioneda S.; Crosetto N.; Matic I.; Hibbert R. G.; Begovic T.; Niimi A.; Mann M.; Lehmann A. R.; Dikic I. (2010) Regulation of translesion synthesis DNA polymerase eta by monoubiquitination. Mol. Cell 37, 396–407. 10.1016/j.molcel.2009.12.039. [DOI] [PubMed] [Google Scholar]
- Jung Y. S.; Liu G.; Chen X. (2010) Pirh2 E3 ubiquitin ligase targets DNA polymerase eta for 20S proteasomal degradation. Mol. Cell. Biol. 30, 1041–1048. 10.1128/MCB.01198-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wimmer U.; Ferrari E.; Hunziker P.; Hubscher U. (2008) Control of DNA polymerase lambda stability by phosphorylation and ubiquitination during the cell cycle. EMBO Rep. 9, 1027–1033. 10.1038/embor.2008.148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hirota K.; Tsuda M.; Mohiuddin; Tsurimoto T.; Cohen I. S.; Livneh Z.; Kobayashi K.; Narita T.; Nishihara K.; Murai J.; Iwai S.; Guilbaud G.; Sale J. E.; Takeda S. (2016) In vivo evidence for translesion synthesis by the replicative DNA polymerase delta. Nucleic Acids Res. gkw439. 10.1093/nar/gkw439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moriya M. (1993) Single-stranded shuttle phagemid for mutagenesis studies in mammalian cells: 8-oxoguanine in DNA induces targeted G.C-- > T.A transversions in simian kidney cells. Proc. Natl. Acad. Sci. U. S. A. 90, 1122–1126. 10.1073/pnas.90.3.1122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moriya M.; Ou C.; Bodepudi V.; Johnson F.; Takeshita M.; Grollman A. P. (1991) Site-specific mutagenesis using a gapped duplex vector: a study of translesion synthesis past 8-oxodeoxyguanosine in E. coli. Mutat. Res., DNA Repair 254, 281–288. 10.1016/0921-8777(91)90067-Y. [DOI] [PubMed] [Google Scholar]
- Hsu G. W.; Ober M.; Carell T.; Beese L. S. (2004) Error-prone replication of oxidatively damaged DNA by a high-fidelity DNA polymerase. Nature 431, 217–221. 10.1038/nature02908. [DOI] [PubMed] [Google Scholar]
- Einolf H. J.; Guengerich F. P. (2001) Fidelity of nucleotide insertion at 8-oxo-7,8-dihydroguanine by mammalian DNA polymerase delta. Steady-state and pre-steady-state kinetic analysis. J. Biol. Chem. 276, 3764–3771. 10.1074/jbc.M006696200. [DOI] [PubMed] [Google Scholar]
- Maga G.; Villani G.; Crespan E.; Wimmer U.; Ferrari E.; Bertocci B.; Hubscher U. (2007) 8-oxo-guanine bypass by human DNA polymerases in the presence of auxiliary proteins. Nature 447, 606–608. 10.1038/nature05843. [DOI] [PubMed] [Google Scholar]
- Haracska L.; Prakash S.; Prakash L. (2003) Yeast DNA polymerase zeta is an efficient extender of primer ends opposite from 7,8-dihydro-8-Oxoguanine and O6-methylguanine. Mol. Cell. Biol. 23, 1453–1459. 10.1128/MCB.23.4.1453-1459.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kamiya H. (2003) Mutagenic potentials of damaged nucleic acids produced by reactive oxygen/nitrogen species: approaches using synthetic oligonucleotides and nucleotides: survey and summary. Nucleic Acids Res. 31, 517–531. 10.1093/nar/gkg137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kalam M. A.; Haraguchi K.; Chandani S.; Loechler E. L.; Moriya M.; Greenberg M. M.; Basu A. K. (2006) Genetic effects of oxidative DNA damages: comparative mutagenesis of the imidazole ring-opened formamidopyrimidines (Fapy lesions) and 8-oxo-purines in simian kidney cells. Nucleic Acids Res. 34, 2305–2315. 10.1093/nar/gkl099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kalam M. A.; Basu A. K. (2005) Mutagenesis of 8-oxoguanine adjacent to an abasic site in simian kidney cells: tandem mutations and enhancement of G-- > T transversions. Chem. Res. Toxicol. 18, 1187–1192. 10.1021/tx050119r. [DOI] [PubMed] [Google Scholar]
- Hazra T. K.; Izumi T.; Maidt L.; Floyd R. A.; Mitra S. (1998) The presence of two distinct 8-oxoguanine repair enzymes in human cells: their potential complementary roles in preventing mutation. Nucleic Acids Res. 26, 5116–5122. 10.1093/nar/26.22.5116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hazra T. K.; Hill J. W.; Izumi T.; Mitra S. (2001) Multiple DNA glycosylases for repair of 8-oxoguanine and their potential in vivo functions. Prog. Nucleic Acid Res. Mol. Biol. 68, 193–205. 10.1016/S0079-6603(01)68100-5. [DOI] [PubMed] [Google Scholar]
- Kundu S.; Brinkmeyer M. K.; Eigenheer R. A.; David S. S. (2010) Ser 524 is a phosphorylation site in MUTYH and Ser 524 mutations alter 8-oxoguanine (OG): a mismatch recognition. DNA Repair 9, 1026–1037. 10.1016/j.dnarep.2010.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maga G.; Crespan E.; Wimmer U.; van Loon B.; Amoroso A.; Mondello C.; Belgiovine C.; Ferrari E.; Locatelli G.; Villani G.; Hubscher U. (2008) Replication protein A and proliferating cell nuclear antigen coordinate DNA polymerase selection in 8-oxo-guanine repair. Proc. Natl. Acad. Sci. U. S. A. 105, 20689–20694. 10.1073/pnas.0811241106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Markkanen E.; Castrec B.; Villani G.; Hubscher U. (2012) A switch between DNA polymerases delta and lambda promotes error-free bypass of 8-oxo-G lesions. Proc. Natl. Acad. Sci. U. S. A. 109, 20401–20406. 10.1073/pnas.1211532109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodriguez G. P.; Song J. B.; Crouse G. F. (2013) In vivo bypass of 8-oxodG. PLoS Genet. 9, e1003682. 10.1371/journal.pgen.1003682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maga G.; Crespan E.; Markkanen E.; Imhof R.; Furrer A.; Villani G.; Hubscher U.; van Loon B. (2013) DNA polymerase delta-interacting protein 2 is a processivity factor for DNA polymerase lambda during 8-oxo-7,8-dihydroguanine bypass. Proc. Natl. Acad. Sci. U. S. A. 110, 18850–18855. 10.1073/pnas.1308760110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pande P.; Haraguchi K.; Jiang Y. L.; Greenberg M. M.; Basu A. K. (2015) Unlike catalyzing error-free bypass of 8-oxodGuo, DNA polymerase lambda is responsible for a significant part of Fapy.dG-induced G --> T mutations in human cells. Biochemistry 54, 1859–1862. 10.1021/acs.biochem.5b00119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takao M.; Zhang Q. M.; Yonei S.; Yasui A. (1999) Differential subcellular localization of human MutY homolog (hMYH) and the functional activity of adenine:8-oxoguanine DNA glycosylase. Nucleic Acids Res. 27, 3638–3644. 10.1093/nar/27.18.3638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Loon B.; Hubscher U. (2009) An 8-oxo-guanine repair pathway coordinated by MUTYH glycosylase and DNA polymerase lambda. Proc. Natl. Acad. Sci. U. S. A. 106, 18201–18206. 10.1073/pnas.0907280106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yasui M.; Kanemaru Y.; Kamoshita N.; Suzuki T.; Arakawa T.; Honma M. (2014) Tracing the fates of site-specifically introduced DNA adducts in the human genome. DNA Repair 15, 11–20. 10.1016/j.dnarep.2014.01.003. [DOI] [PubMed] [Google Scholar]
- Al-Tassan N.; Chmiel N. H.; Maynard J.; Fleming N.; Livingston A. L.; Williams G. T.; Hodges A. K.; Davies D. R.; David S. S.; Sampson J. R.; Cheadle J. P. (2002) Inherited variants of MYH associated with somatic G:C-- > T:A mutations in colorectal tumors. Nat. Genet. 30, 227–232. 10.1038/ng828. [DOI] [PubMed] [Google Scholar]
- Patro J. N.; Wiederholt C. J.; Jiang Y. L.; Delaney J. C.; Essigmann J. M.; Greenberg M. M. (2007) Studies on the replication of the ring opened formamidopyrimidine, Fapy.dG in Escherichia coli. Biochemistry 46, 10202–10212. 10.1021/bi700628c. [DOI] [PubMed] [Google Scholar]
- Gehrke T. H.; Lischke U.; Gasteiger K. L.; Schneider S.; Arnold S.; Muller H. C.; Stephenson D. S.; Zipse H.; Carell T. (2013) Unexpected non-Hoogsteen-based mutagenicity mechanism of FaPy-DNA lesions. Nat. Chem. Biol. 9, 455–461. 10.1038/nchembio.1254. [DOI] [PubMed] [Google Scholar]
- Gates K. S.; Nooner T.; Dutta S. (2004) Biologically relevant chemical reactions of N7-alkylguanine residues in DNA. Chem. Res. Toxicol. 17, 839–856. 10.1021/tx049965c. [DOI] [PubMed] [Google Scholar]
- Kadlubar F. F.; Beranek D. T.; Weis C. C.; Evans F. E.; Cox R.; Irving C. C. (1984) Characterization of the purine ring-opened 7-methylguanine and its persistence in rat bladder epithelial DNA after treatment with the carcinogen N-methylnitrosourea. Carcinogenesis 5, 587–592. 10.1093/carcin/5.5.587. [DOI] [PubMed] [Google Scholar]
- Christov P. P.; Angel K. C.; Guengerich F. P.; Rizzo C. J. (2009) Replication past the N5-methyl-formamidopyrimidine lesion of deoxyguanosine by DNA polymerases and an improved procedure for sequence analysis of in vitro bypass products by mass spectrometry. Chem. Res. Toxicol. 22, 1086–1095. 10.1021/tx900047c. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Christov P. P.; Yamanaka K.; Choi J. Y.; Takata K.; Wood R. D.; Guengerich F. P.; Lloyd R. S.; Rizzo C. J. (2012) Replication of the 2,6-diamino-4-hydroxy-N(5)-(methyl)-formamidopyrimidine (MeFapy-dGuo) adduct by eukaryotic DNA polymerases. Chem. Res. Toxicol. 25, 1652–1661. 10.1021/tx300113e. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Earley L. F.; Minko I. G.; Christov P. P.; Rizzo C. J.; Lloyd R. S. (2013) Mutagenic spectra arising from replication bypass of the 2,6-diamino-4-hydroxy-N(5)-methyl formamidopyrimidine adduct in primate cells. Chem. Res. Toxicol. 26, 1108–1114. 10.1021/tx4001495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brooks K. (1990) A brief history of the Journal. Miss. Dent. Assoc. J. 46, 27. [PubMed] [Google Scholar]
- Karran P.; Macpherson P.; Ceccotti S.; Dogliotti E.; Griffin S.; Bignami M. (1993) O6-methylguanine residues elicit DNA repair synthesis by human cell extracts. J. Biol. Chem. 268, 15878–15886. [PubMed] [Google Scholar]
- Esteller M.; Toyota M.; Sanchez-Cespedes M.; Capella G.; Peinado M. A.; Watkins D. N.; Issa J. P.; Sidransky D.; Baylin S. B.; Herman J. G. (2000) Inactivation of the DNA repair gene O6-methylguanine-DNA methyltransferase by promoter hypermethylation is associated with G to A mutations in K-ras in colorectal tumorigenesis. Cancer Res. 60, 2368–2371. [PubMed] [Google Scholar]
- Rye P. T.; Delaney J. C.; Netirojjanakul C.; Sun D. X.; Liu J. Z.; Essigmann J. M. (2008) Mismatch repair proteins collaborate with methyltransferases in the repair of O(6)-methylguanine. DNA Repair 7, 170–176. 10.1016/j.dnarep.2007.09.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nicoll J. W.; Swann P. F.; Pegg A. E. (1975) Effect of dimethylnitrosamine on persistence of methylated guanines in rat liver and kidney DNA. Nature 254, 261–262. 10.1038/254261a0. [DOI] [PubMed] [Google Scholar]
- Sukumar S.; Notario V.; Martin-Zanca D.; Barbacid M. (1983) Induction of mammary carcinomas in rats by nitroso-methylurea involves malignant activation of H-ras-1 locus by single point mutations. Nature 306, 658–661. 10.1038/306658a0. [DOI] [PubMed] [Google Scholar]
- Zarbl H.; Sukumar S.; Arthur A. V.; Martin-Zanca D.; Barbacid M. (1985) Direct mutagenesis of Ha-ras-1 oncogenes by N-nitroso-N-methylurea during initiation of mammary carcinogenesis in rats. Nature 315, 382–385. 10.1038/315382a0. [DOI] [PubMed] [Google Scholar]
- Belinsky S. A.; Devereux T. R.; Maronpot R. R.; Stoner G. D.; Anderson M. W. (1989) Relationship between the formation of promutagenic adducts and the activation of the K-ras protooncogene in lung tumors from A/J mice treated with nitrosamines. Cancer Res. 49, 5305–5311. [PubMed] [Google Scholar]
- Ellison K. S.; Dogliotti E.; Connors T. D.; Basu A. K.; Essigmann J. M. (1989) Site-specific mutagenesis by O6-alkylguanines located in the chromosomes of mammalian cells: influence of the mammalian O6-alkylguanine-DNA alkyltransferase. Proc. Natl. Acad. Sci. U. S. A. 86, 8620–8624. 10.1073/pnas.86.22.8620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choi J. Y.; Chowdhury G.; Zang H.; Angel K. C.; Vu C. C.; Peterson L. A.; Guengerich F. P. (2006) Translesion synthesis across O6-alkylguanine DNA adducts by recombinant human DNA polymerases. J. Biol. Chem. 281, 38244–38256. 10.1074/jbc.M608369200. [DOI] [PubMed] [Google Scholar]
- Voigt J. M.; Topal M. D. (1995) O6-methylguanine-induced replication blocks. Carcinogenesis 16, 1775–1782. 10.1093/carcin/16.8.1775. [DOI] [PubMed] [Google Scholar]
- Singh J.; Su L.; Snow E. T. (1996) Replication across O6-methylguanine by human DNA polymerase beta in vitro. Insights into the futile cytotoxic repair and mutagenesis of O6-methylguanine. J. Biol. Chem. 271, 28391–28398. 10.1074/jbc.271.45.28391. [DOI] [PubMed] [Google Scholar]
- Haracska L.; Prakash S.; Prakash L. (2000) Replication past O(6)-methylguanine by yeast and human DNA polymerase eta. Mol. Cell. Biol. 20, 8001–8007. 10.1128/MCB.20.21.8001-8007.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson R. E.; Prakash S.; Prakash L. (1999) Efficient bypass of a thymine-thymine dimer by yeast DNA polymerase, Pol eta. Science 283, 1001–1004. 10.1126/science.283.5404.1001. [DOI] [PubMed] [Google Scholar]
- Washington M. T.; Prakash L.; Prakash S. (2003) Mechanism of nucleotide incorporation opposite a thymine-thymine dimer by yeast DNA polymerase eta. Proc. Natl. Acad. Sci. U. S. A. 100, 12093–12098. 10.1073/pnas.2134223100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoon J. H.; Prakash L.; Prakash S. (2009) Highly error-free role of DNA polymerase eta in the replicative bypass of UV-induced pyrimidine dimers in mouse and human cells. Proc. Natl. Acad. Sci. U. S. A. 106, 18219–18224. 10.1073/pnas.0910121106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson R. E.; Kondratick C. M.; Prakash S.; Prakash L. (1999) hRAD30 mutations in the variant form of xeroderma pigmentosum. Science 285, 263–265. 10.1126/science.285.5425.263. [DOI] [PubMed] [Google Scholar]
- Masutani C.; Araki M.; Yamada A.; Kusumoto R.; Nogimori T.; Maekawa T.; Iwai S.; Hanaoka F. (1999) Xeroderma pigmentosum variant (XP-V) correcting protein from HeLa cells has a thymine dimer bypass DNA polymerase activity. EMBO J. 18, 3491–3501. 10.1093/emboj/18.12.3491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Masutani C.; Kusumoto R.; Yamada A.; Dohmae N.; Yokoi M.; Yuasa M.; Araki M.; Iwai S.; Takio K.; Hanaoka F. (1999) The XPV (xeroderma pigmentosum variant) gene encodes human DNA polymerase eta. Nature 399, 700–704. 10.1038/21447. [DOI] [PubMed] [Google Scholar]
- Vaisman A.; Masutani C.; Hanaoka F.; Chaney S. G. (2000) Efficient translesion replication past oxaliplatin and cisplatin GpG adducts by human DNA polymerase eta. Biochemistry 39, 4575–4580. 10.1021/bi000130k. [DOI] [PubMed] [Google Scholar]
- Bassett E.; King N. M.; Bryant M. F.; Hector S.; Pendyala L.; Chaney S. G.; Cordeiro-Stone M. (2004) The role of DNA polymerase eta in translesion synthesis past platinum-DNA adducts in human fibroblasts. Cancer Res. 64, 6469–6475. 10.1158/0008-5472.CAN-04-1328. [DOI] [PubMed] [Google Scholar]
- Zhao Y.; Biertumpfel C.; Gregory M. T.; Hua Y. J.; Hanaoka F.; Yang W. (2012) Structural basis of human DNA polymerase eta-mediated chemoresistance to cisplatin. Proc. Natl. Acad. Sci. U. S. A. 109, 7269–7274. 10.1073/pnas.1202681109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wogan G. N. (1999) Aflatoxin as a human carcinogen. Hepatology 30, 573–575. 10.1002/hep.510300231. [DOI] [PubMed] [Google Scholar]
- Kensler T. W.; Roebuck B. D.; Wogan G. N.; Groopman J. D. (2011) Aflatoxin: a 50-year odyssey of mechanistic and translational toxicology. Toxicol. Sci. 120, S28–S48. 10.1093/toxsci/kfq283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Essigmann J. M.; Croy R. G.; Nadzan A. M.; Busby W. F. Jr.; Reinhold V. N.; Buchi G.; Wogan G. N. (1977) Structural identification of the major DNA adduct formed by aflatoxin B1 in vitro. Proc. Natl. Acad. Sci. U. S. A. 74, 1870–1874. 10.1073/pnas.74.5.1870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Croy R. G.; Essigmann J. M.; Reinhold V. N.; Wogan G. N. (1978) Identification of the principal aflatoxin B1-DNA adduct formed in vivo in rat liver. Proc. Natl. Acad. Sci. U. S. A. 75, 1745–1749. 10.1073/pnas.75.4.1745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Croy R. G.; Essigmann J. M.; Wogan G. N. (1983) Aflatoxin B1: correlations of patterns of metabolism and DNA modification with biological effects. Basic Life Sci. 24, 49–62. 10.1007/978-1-4684-4400-1_3. [DOI] [PubMed] [Google Scholar]
- Lin Y. C.; Li L.; Makarova A. V.; Burgers P. M.; Stone M. P.; Lloyd R. S. (2014) Error-prone replication bypass of the primary aflatoxin B1 DNA adduct, AFB1-N7-Gua. J. Biol. Chem. 289, 18497–18506. 10.1074/jbc.M114.561563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin Y. C.; Li L.; Makarova A. V.; Burgers P. M.; Stone M. P.; Lloyd R. S. (2014) Molecular basis of aflatoxin-induced mutagenesis-role of the aflatoxin B1-formamidopyrimidine adduct. Carcinogenesis 35, 1461–1468. 10.1093/carcin/bgu003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fernandes A.; Liu T.; Amin S.; Geacintov N. E.; Grollman A. P.; Moriya M. (1998) Mutagenic potential of stereoisomeric bay region (+)- and (−)-cis-anti-benzo[a]pyrene diol epoxide-N2-2′-deoxyguanosine adducts in Escherichia coli and simian kidney cells. Biochemistry 37, 10164–10172. 10.1021/bi980401f. [DOI] [PubMed] [Google Scholar]
- Dong H.; Bonala R. R.; Suzuki N.; Johnson F.; Grollman A. P.; Shibutani S. (2004) Mutagenic potential of benzo[a]pyrene-derived DNA adducts positioned in codon 273 of the human P53 gene. Biochemistry 43, 15922–15928. 10.1021/bi0482194. [DOI] [PubMed] [Google Scholar]
- Moriya M.; Spiegel S.; Fernandes A.; Amin S.; Liu T.; Geacintov N.; Grollman A. P. (1996) Fidelity of translesional synthesis past benzo[a]pyrene diol epoxide-2′-deoxyguanosine DNA adducts: marked effects of host cell, sequence context, and chirality. Biochemistry 35, 16646–16651. 10.1021/bi9608875. [DOI] [PubMed] [Google Scholar]
- Frank E. G.; Sayer J. M.; Kroth H.; Ohashi E.; Ohmori H.; Jerina D. M.; Woodgate R. (2002) Translesion replication of benzo[a]pyrene and benzo[c]phenanthrene diol epoxide adducts of deoxyadenosine and deoxyguanosine by human DNA polymerase iota. Nucleic Acids Res. 30, 5284–5292. 10.1093/nar/gkf643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chiapperino D.; Kroth H.; Kramarczuk I. H.; Sayer J. M.; Masutani C.; Hanaoka F.; Jerina D. M.; Cheh A. M. (2002) Preferential misincorporation of purine nucleotides by human DNA polymerase eta opposite benzo[a]pyrene 7,8-diol 9,10-epoxide deoxyguanosine adducts. J. Biol. Chem. 277, 11765–11771. 10.1074/jbc.M112139200. [DOI] [PubMed] [Google Scholar]
- Xie Z.; Braithwaite E.; Guo D.; Zhao B.; Geacintov N. E.; Wang Z. (2003) Mutagenesis of benzo[a]pyrene diol epoxide in yeast: requirement for DNA polymerase zeta and involvement of DNA polymerase eta. Biochemistry 42, 11253–11262. 10.1021/bi0346704. [DOI] [PubMed] [Google Scholar]
- Avkin S.; Goldsmith M.; Velasco-Miguel S.; Geacintov N.; Friedberg E. C.; Livneh Z. (2004) Quantitative analysis of translesion DNA synthesis across a benzo[a]pyrene-guanine adduct in mammalian cells: the role of DNA polymerase kappa. J. Biol. Chem. 279, 53298–53305. 10.1074/jbc.M409155200. [DOI] [PubMed] [Google Scholar]
- Liu Y.; Yang Y.; Tang T. S.; Zhang H.; Wang Z.; Friedberg E.; Yang W.; Guo C. (2014) Variants of mouse DNA polymerase kappa reveal a mechanism of efficient and accurate translesion synthesis past a benzo[a]pyrene dG adduct. Proc. Natl. Acad. Sci. U. S. A. 111, 1789–1794. 10.1073/pnas.1324168111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choi J. Y.; Angel K. C.; Guengerich F. P. (2006) Translesion synthesis across bulky N2-alkyl guanine DNA adducts by human DNA polymerase kappa. J. Biol. Chem. 281, 21062–21072. 10.1074/jbc.M602246200. [DOI] [PubMed] [Google Scholar]
- Hashimoto K.; Cho Y.; Yang I. Y.; Akagi J.; Ohashi E.; Tateishi S.; de Wind N.; Hanaoka F.; Ohmori H.; Moriya M. (2012) The vital role of polymerase zeta and REV1 in mutagenic, but not correct, DNA synthesis across benzo[a]pyrene-dG and recruitment of polymerase zeta by REV1 to replication-stalled site. J. Biol. Chem. 287, 9613–9622. 10.1074/jbc.M111.331728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jha V.; Bian C.; Xing G.; Ling H. (2016) Structure and mechanism of error-free replication past the major benzo[a]pyrene adduct by human DNA polymerase kappa. Nucleic Acids Res. 44, 4957–4967. 10.1093/nar/gkw204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jarosz D. F.; Godoy V. G.; Delaney J. C.; Essigmann J. M.; Walker G. C. (2006) A single amino acid governs enhanced activity of DinB DNA polymerases on damaged templates. Nature 439, 225–228. 10.1038/nature04318. [DOI] [PubMed] [Google Scholar]
- Yuan B.; Cao H.; Jiang Y.; Hong H.; Wang Y. (2008) Efficient and accurate bypass of N2-(1-carboxyethyl)-2′-deoxyguanosine by DinB DNA polymerase in vitro and in vivo. Proc. Natl. Acad. Sci. U. S. A. 105, 8679–8684. 10.1073/pnas.0711546105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bose A.; Millsap A. D.; DeLeon A.; Rizzo C. J.; Basu A. K. (2016) Translesion synthesis of the N2-2′-deoxyguanosine adduct of the dietary mutagen IQ in human cells: Error-free replication by DNA polymerase kappa and mutagenic bypass by DNA polymerases eta, zeta, and Rev1. Chem. Res. Toxicol. 29, 1549–1559. 10.1021/acs.chemrestox.6b00221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bose A.; Surugihalli C.; Pande P.; Champeil E.; Basu A. K. (2016) Comparative Error-Free and Error-Prone Translesion Synthesis of N(2)-2′-Deoxyguanosine Adducts Formed by Mitomycin C and Its Metabolite, 2,7-Diaminomitosene, in Human Cells. Chem. Res. Toxicol. 29, 933–939. 10.1021/acs.chemrestox.6b00087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagao M.; Wakabayashi K.; Ushijima T.; Toyota M.; Totsuka Y.; Sugimura T. (1996) Human exposure to carcinogenic heterocyclic amines and their mutational fingerprints in experimental animals. Environ. Health Perspect. 104 (Suppl 3), 497–501. 10.1289/ehp.96104s3497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sugimura T.; Wakabayashi K.; Nakagama H.; Nagao M. (2004) Heterocyclic amines: Mutagens/carcinogens produced during cooking of meat and fish. Cancer Sci. 95, 290–299. 10.1111/j.1349-7006.2004.tb03205.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sugimura T.; Nagao M.; Wakabayashi K. (1994) Heterocyclic amines in cooked foods: candidates for causation of common cancers. J. Natl. Cancer Inst. 86, 2–4. 10.1093/jnci/86.1.2. [DOI] [PubMed] [Google Scholar]
- Tomasz M. (1995) Mitomycin C: small, fast and deadly (but very selective). Chem. Biol. 2, 575–579. 10.1016/1074-5521(95)90120-5. [DOI] [PubMed] [Google Scholar]
- Paz M. M.; Pritsos C. A. (2012) The molecular toxicology of mitomycin C. Advances in Mol. Toxicol. 6, 243–299. 10.1016/B978-0-444-59389-4.00007-0. [DOI] [Google Scholar]
- Choi J. Y.; Guengerich F. P. (2005) Adduct size limits efficient and error-free bypass across bulky N2-guanine DNA lesions by human DNA polymerase eta. J. Mol. Biol. 352, 72–90. 10.1016/j.jmb.2005.06.079. [DOI] [PubMed] [Google Scholar]
- Washington M. T.; Minko I. G.; Johnson R. E.; Wolfle W. T.; Harris T. M.; Lloyd R. S.; Prakash S.; Prakash L. (2004) Efficient and error-free replication past a minor-groove DNA adduct by the sequential action of human DNA polymerases iota and kappa. Mol. Cell. Biol. 24, 5687–5693. 10.1128/MCB.24.13.5687-5693.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wolfle W. T.; Johnson R. E.; Minko I. G.; Lloyd R. S.; Prakash S.; Prakash L. (2006) Replication past a trans-4-hydroxynonenal minor-groove adduct by the sequential action of human DNA polymerases iota and kappa. Mol. Cell. Biol. 26, 381–386. 10.1128/MCB.26.1.381-386.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koffel-Schwartz N.; Fuchs R. P. (1989) Genetic control of AAF-induced mutagenesis at alternating GC sequences: an additional role for RecA. Mol. Gen. Genet. 215, 306–311. 10.1007/BF00339733. [DOI] [PubMed] [Google Scholar]
- Lambert I. B.; Napolitano R. L.; Fuchs R. P. (1992) Carcinogen-induced frameshift mutagenesis in repetitive sequences. Proc. Natl. Acad. Sci. U. S. A. 89, 1310–1314. 10.1073/pnas.89.4.1310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomas D. C.; Veaute X.; Kunkel T. A.; Fuchs R. P. (1994) Mutagenic replication in human cell extracts of DNA containing site-specific N-2-acetylaminofluorene adducts. Proc. Natl. Acad. Sci. U. S. A. 91, 7752–7756. 10.1073/pnas.91.16.7752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan X.; Suzuki N.; Grollman A. P.; Shibutani S. (2002) Mutagenic events in Escherichia coli and mammalian cells generated in response to acetylaminofluorene-derived DNA adducts positioned in the Nar I restriction enzyme site. Biochemistry 41, 14255–14262. 10.1021/bi0202878. [DOI] [PubMed] [Google Scholar]
- Janel-Bintz R.; Wagner J.; Haracska L.; Mah-Becherel M. C.; Bichara M.; Fuchs R. P.; Cordonnier A. M. (2012) Evidence for a Rad18-independent frameshift mutagenesis pathway in human cell-free extracts. PLoS One 7, e36004. 10.1371/journal.pone.0036004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Broyde S.; Hingerty B. E. (1987) Visualization of an AAF induced frameshift mutation: molecular views of base displacement in B-DNA from minimized potential energy calculations. Nucleic Acids Res. 15, 6539–6552. 10.1093/nar/15.16.6539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roy D.; Hingerty B. E.; Shapiro R.; Broyde S. (1998) A slipped replication intermediate model is stabilized by the syn orientation of N-2-aminofluorene- and N-2-(acetyl)aminofluorene-modified guanine at a mutational hotspot. Chem. Res. Toxicol. 11, 1301–1311. 10.1021/tx980106w. [DOI] [PubMed] [Google Scholar]
- Schorr S.; Carell T. (2010) Mechanism of acetylaminofluorene-dG induced frameshifting by polymerase eta. ChemBioChem 11, 2534–2537. 10.1002/cbic.201000579. [DOI] [PubMed] [Google Scholar]
- O’Handley S. F.; Sanford D. G.; Xu R.; Lester C. C.; Hingerty B. E.; Broyde S.; Krugh T. R. (1993) Structural characterization of an N-acetyl-2-aminofluorene (AAF) modified DNA oligomer by NMR, energy minimization, and molecular dynamics. Biochemistry 32, 2481–2497. 10.1021/bi00061a005. [DOI] [PubMed] [Google Scholar]
- Patel D. J.; Mao B.; Gu Z.; Hingerty B. E.; Gorin A.; Basu A. K.; Broyde S. (1998) Nuclear magnetic resonance solution structures of covalent aromatic amine-DNA adducts and their mutagenic relevance. Chem. Res. Toxicol. 11, 391–407. 10.1021/tx9702143. [DOI] [PubMed] [Google Scholar]
- Broyde S.; Wang L.; Zhang L.; Rechkoblit O.; Geacintov N. E.; Patel D. J. (2008) DNA adduct structure-function relationships: comparing solution with polymerase structures. Chem. Res. Toxicol. 21, 45–52. 10.1021/tx700193x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Napolitano R. L.; Lambert I. B.; Fuchs R. P. (1994) DNA sequence determinants of carcinogen-induced frameshift mutagenesis. Biochemistry 33, 1311–1315. 10.1021/bi00172a004. [DOI] [PubMed] [Google Scholar]
- Nolan S. J.; McNulty J. M.; Krishnasamy R.; McGregor W. G.; Basu A. K. (1999) C8-guanine adduct-induced stabilization of a −1 frame shift intermediate in a nonrepetitive DNA sequence. Biochemistry 38, 14056–14062. 10.1021/bi991342o. [DOI] [PubMed] [Google Scholar]
- Schorr S.; Schneider S.; Lammens K.; Hopfner K. P.; Carell T. (2010) Mechanism of replication blocking and bypass of Y-family polymerase {eta} by bulky acetylaminofluorene DNA adducts. Proc. Natl. Acad. Sci. U. S. A. 107, 20720–20725. 10.1073/pnas.1008894107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bose A.; Pande P.; Jasti V. P.; Millsap A. D.; Hawkins E. K.; Rizzo C. J.; Basu A. K. (2015) DNA polymerases kappa and zeta cooperatively perform mutagenic translesion synthesis of the C8–2′-deoxyguanosine adduct of the dietary mutagen IQ in human cells. Nucleic Acids Res. 43, 8340–8351. 10.1093/nar/gkv750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pande P.; Malik C. K.; Bose A.; Jasti V. P.; Basu A. K. (2014) Mutational analysis of the C8-guanine adduct of the environmental carcinogen 3-nitrobenzanthrone in human cells: critical roles of DNA polymerases eta and kappa and Rev1 in error-prone translesion synthesis. Biochemistry 53, 5323–5331. 10.1021/bi5007805. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arlt V. M. (2005) 3-Nitrobenzanthrone, a potential human cancer hazard in diesel exhaust and urban air pollution: a review of the evidence. Mutagenesis 20, 399–410. 10.1093/mutage/gei057. [DOI] [PubMed] [Google Scholar]
- Bieler C. A.; Cornelius M. G.; Stiborova M.; Arlt V. M.; Wiessler M.; Phillips D. H.; Schmeiser H. H. (2007) Formation and persistence of DNA adducts formed by the carcinogenic air pollutant 3-nitrobenzanthrone in target and non-target organs after intratracheal instillation in rats. Carcinogenesis 28, 1117–1121. 10.1093/carcin/bgl219. [DOI] [PubMed] [Google Scholar]
- Tokarsky E. J.; Gadkari V. V.; Zahurancika W. J.; Malik C. K.; Basu A. K.; Suo Z. (2016) Pre-Steady-State Kinetic Investigation of Bypass of a Bulky Guanine Lesion by Human Y-family DNA Polymerases. DNA Repair 46, 20. 10.1016/j.dnarep.2016.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patra A.; Politica D. A.; Chatterjee A.; Tokarsky E. J.; Suo Z.; Basu A. K.; Stone M. P.; Egli M. (2016) Mechanism of error free bypass of the environmental carcinogen N-(2′-deoxyguanosin-8-yl)-3-aminobenzanthrone adduct by human DNA polymerase η. ChemBioChem 17, 1–6. 10.1002/cbic.201600420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang F.; DeMuro N. E.; Elmquist C. E.; Stover J. S.; Rizzo C. J.; Stone M. P. (2006) Base-displaced intercalated structure of the food mutagen 2-amino-3-methylimidazo[4,5-f]quinoline in the recognition sequence of the NarI restriction enzyme, a hotspot for −2 bp deletions. J. Am. Chem. Soc. 128, 10085–10095. 10.1021/ja062004v. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Politica D. A.; Malik C. K.; Basu A. K.; Stone M. P. (2015) Base-Displaced Intercalated Structure of the N-(2′-Deoxyguanosin-8-yl)-3-aminobenzanthrone DNA Adduct. Chem. Res. Toxicol. 28, 2253–2266. 10.1021/acs.chemrestox.5b00277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shibutani S.; Suzuki N.; Grollman A. P. (1998) Mutagenic specificity of (acetylamino)fluorene-derived DNA adducts in mammalian cells. Biochemistry 37, 12034–12041. 10.1021/bi981059+. [DOI] [PubMed] [Google Scholar]
- Watt D. L.; Utzat C. D.; Hilario P.; Basu A. K. (2007) Mutagenicity of the 1-nitropyrene-DNA adduct N-(deoxyguanosin-8-yl)-1-aminopyrene in mammalian cells. Chem. Res. Toxicol. 20, 1658–1664. 10.1021/tx700131e. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nair D. T.; Johnson R. E.; Prakash S.; Prakash L.; Aggarwal A. K. (2004) Replication by human DNA polymerase-iota occurs by Hoogsteen base-pairing. Nature 430, 377–380. 10.1038/nature02692. [DOI] [PubMed] [Google Scholar]
- Mao B.; Vyas R. R.; Hingerty B. E.; Broyde S.; Basu A. K.; Patel D. J. (1996) Solution conformation of the N-(deoxyguanosin-8-yl)-1-aminopyrene ([AP]dG) adduct opposite dC in a DNA duplex. Biochemistry 35, 12659–12670. 10.1021/bi961078o. [DOI] [PubMed] [Google Scholar]
- Gu Z.; Gorin A.; Krishnasamy R.; Hingerty B. E.; Basu A. K.; Broyde S.; Patel D. J. (1999) Solution structure of the N-(deoxyguanosin-8-yl)-1-aminopyrene ([AP]dG) adduct opposite dA in a DNA duplex. Biochemistry 38, 10843–10854. 10.1021/bi9912138. [DOI] [PubMed] [Google Scholar]
- Sherrer S. M.; Sanman L. E.; Xia C. X.; Bolin E. R.; Malik C. K.; Efthimiopoulos G.; Basu A. K.; Suo Z. (2012) Kinetic analysis of the bypass of a bulky DNA lesion catalyzed by human Y-family DNA polymerases. Chem. Res. Toxicol. 25, 730–740. 10.1021/tx200531y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sherrer S. M.; Taggart D. J.; Pack L. R.; Malik C. K.; Basu A. K.; Suo Z. (2012) Quantitative analysis of the mutagenic potential of 1-aminopyrene-DNA adduct bypass catalyzed by Y-family DNA polymerases. Mutat. Res., Fundam. Mol. Mech. Mutagen. 737, 25–33. 10.1016/j.mrfmmm.2012.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kirouac K. N.; Basu A. K.; Ling H. (2013) Replication of a carcinogenic nitropyrene DNA lesion by human Y-family DNA polymerase. Nucleic Acids Res. 41, 2060–2071. 10.1093/nar/gks1296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- You C.; Swanson A. L.; Dai X.; Yuan B.; Wang J.; Wang Y. (2013) Translesion synthesis of 8,5′-cyclopurine-2′-deoxynucleosides by DNA polymerases eta, iota, and zeta. J. Biol. Chem. 288, 28548–28556. 10.1074/jbc.M113.480459. [DOI] [PMC free article] [PubMed] [Google Scholar]