

Abstract
Background
In preparation for migration from freshwater to marine habitats, Atlantic salmon (Salmo salar L.) undergo smoltification, a transformation that includes the acquisition of hyposmoregulatory capacity. The growth hormone (Gh)/insulin-like growth-factor (Igf) axis promotes the development of branchial ionoregulatory functions that underlie ion secretion. Igfs interact with a suite of Igf binding proteins (Igfbps) that modulate hormone activity. In Atlantic salmon smolts, igfbp4,−5a,−5b1,−5b2,−6b1 and−6b2 transcripts are highly expressed in gill. We measured mRNA levels of branchial and hepatic igfbps during smoltification (March, April, and May), desmoltification (July) and following seawater (SW) exposure in March and May. We also characterized parallel changes in a broad suite of osmoregulatory (branchial Na+/K+-ATPase (Nka) activity, Na + /K + /2Cl − cotransporter 1 (nkcc1) and cystic fibrosis transmembrane regulator 1 (cftr1) transcription) and endocrine (plasma Gh and Igf1) parameters.
Results
Indicative of smoltification, we observed increased branchial Nka activity, nkcc1 and cftr1 transcription in May. Branchial igfbp6b1 and -6b2 expression increased coincidentally with smoltification. Following a SW challenge in March, igfbp6b1 showed increased expression while igfbp6b2 exhibited diminished expression. igfbp5a,−5b1 and−5b2 mRNA levels did not change during smolting, but each had lower levels following a SW exposure in March.
Conclusions
Salmonids express an especially large suite of igfbps. Our data suggest that dynamic expression of particular igfbps accompanies smoltification and SW challenges; thus, transcriptional control of igfbps may provide a mechanism for the local modulation of Igf activity in salmon gill.
Keywords: Insulin-like growth-factor, Binding proteins, Growth hormone, Atlantic salmon, Smoltification, Osmoregulation, Gill, Liver, Salinity
Background
Anadromous fishes such as Atlantic salmon (Salmo salar L.) exhibit a life history strategy that includes an initial phase in fresh water (FW) followed by migration to marine environments [1]. The transformation of stream-dwelling ‘parr’ to seawater (SW) tolerant ‘smolts’ entails the orchestrated development of physiological, morphological and behavioral traits that support migration to, and subsequent survival in, pelagic marine environments. While dependent upon reaching a necessary size, the timing of this transformation in Atlantic salmon is initiated by environmental cues such as photoperiod and temperature [2, 3]. Depending upon latitude, this transformation typically occurs at 1–4 years of age in wild Atlantic salmon [4, 5]. The months preceding migration are termed ‘smoltification’ and this stage remains incomplete prior to downstream migration; animals of this stage are termed ‘pre-smolts’. At the peak of smoltification, salmon smolts will move downstream into estuaries and then quickly enter marine environments. Upon reaching sexual maturity in the ocean, adults use olfactory cues to return to their natal FW streams to spawn [6]. Smolts that do not gain entry into marine environments will reverse some of the acquired phenotypes such as salinity tolerance, and revert to pre-smolt phenotypes that are better suited for FW environments.
In order to sustain hydromineral balance upon entry into marine environments the parr-smolt transformation is inextricably linked with the acquisition of SW tolerance. As is the paradigm for strictly marine teleosts, the ability of smolts to inhabit SW is sustained by an array of solute- and water-transporting activities in the gill, gut, kidney and urinary bladder [7]. Inasmuch as the gill is the primary tissue for the active transport of monovalent ions, the recruitment of branchial ionocytes (SW-type ionocytes) that extrude Na+ and Cl− is essential for gaining SW tolerance. SW-type ionocytes employ ion pumps, cotransporters and channels such as Na+/K+-ATPase (Nka), Na+/K+/2Cl− cotransporter 1 (Nkcc1) and cystic fibrosis transmembrane regulator 1 (Cftr1) [8, 9]. Accordingly, branchial Nka activity peaks concomitantly with nkcc1 and cftr1 mRNA levels when salmon achieve maximal SW tolerance [10–12]. Thus, the seasonal patterns of these three parameters reliably predict whether juvenile salmon will be able to sustain hydromineral balance upon exposure to SW [11, 12].
In Atlantic salmon, several endocrine systems synchronize the ontogeny of osmoregulatory systems with downstream migration [1, 12]. In particular, the growth hormone (Gh)/insulin-like growth factor (Igf) axis exhibits enhanced activity at the peak of smoltification [1, 13]. From the perspective of whole-organism performance, a connection between the somatotropic axis and salmonid osmoregulation is supported by numerous findings of improved salinity tolerance following exogenous Gh and/or Igf1 treatment [13–15]. The hyposmoregulatory actions of Gh are seemingly mediated by multiple molecular pathways, including: 1) branchial Gh receptors (Ghr1), 2) the synthesis and secretion of Igf1 from the liver, 3) the local production of Igf1 in the gill, and/or 4) enhanced responsiveness to cortisol [16–21]. Irrespective of the route of action, Gh and Igf1 promote salinity tolerance by regulating branchial Nka activity [22], the gene and protein expression of ionoregulatory factors [8, 23] and ionocyte density [24, 25].
Igfs interact with cognate binding proteins termed Igf binding proteins (Igfbps). The coordinated production, both spatially and temporally, of Igfbps permits modulation of Igf bioavailability in both positive and negative fashions [26]. Igfbps may also exert ligand-independent activities [27]. Studies on teleost Igfbps have primarily focused upon how they mediate growth responses to stressors such as food restriction, temperature, hypoxia, and handling [28], while relatively few studies have investigated Igfbp responses to ionoregulatory challenges [29–32]. Attributed to multiple whole-genome duplication events, Atlantic salmon express an expansive set of 19 igfbp genes [33]. Among these igfbps, igfbp4,−5a,−5b1,−5b2,−6b1 and−6b2 are highly expressed in the gill [33]. There is currently no understating of how branchial igfbp expression is modulated in preparation for ionoregulatory challenges faced by developing salmon.
Given that parr-smolt transformation encompasses numerous physiological preparations underlying marine survival, and consequently recruitment, knowledge of its physiological control informs efforts aimed at restoring endangered populations [34, 35]. Thus, the physiology of Atlantic salmon smoltification presents an important physiological context for how Igfbps underlie Gh/Igf-mediated life-history transitions. In turn, our first objective was to assess whether igfbp mRNA levels change during smoltification. We further investigated whether igfbps respond to abrupt transfer to SW, and whether such responses varied with the degree of SW tolerance. Because the gill is a key tissue underlying the development of SW adaptability, we primarily focused on igfbp transcripts that exhibit significant branchial expression.
Methods
Animals
Atlantic salmon (Salmo salar) parr were obtained in October of 2013 from the Kensington National Fish Hatchery, Kensington, CT, and held at the Conte Anadromous Fish Research Center, Turners Falls, MA. Individuals from this cohort were expected to smolt in the spring of 2014 on the basis of their size (>12 cm fork length) in early February [36]. Fish were held in a 1.5 m diameter fiberglass tank supplied with dechlorinated tap water under natural photoperiod. Water temperature was maintained at 9 °C until late June; water was then maintained at 10.5 °C until the conclusion of the experiment. Fish were fed to satiation twice daily with commercial feed (Bio-Oregon, Longview, WA). All experiments were carried out in accordance with US Geological Survey institutional guidelines and an approved IACUC review (SP 9065).
Experimental design
To sample juvenile Atlantic salmon before, during, and after smolting, fish (n = 8) of mixed sex were sacrificed on March 3, April 8, May 1 and July 10 at 09:00 h (Eastern Standard Time), with food withheld for 24 h prior to sampling. In addition, SW challenges were conducted on March 3 and May 1 at 09:00 h. Sixteen smolts were transferred to a tank with recirculating SW (35 ppt) at 9 °C with particle and charcoal filtration and continuous aeration. Food was withheld for the duration of the challenge. Fish were sampled (n = 8) at 09:00 h at 24 and 48 h after transfer to SW.
Sampling
At the time of sampling, fish were netted and anesthetized in buffered MS-222 (100 mg/l; pH 7.0; Sigma, St. Louis, MO). Blood was collected from the caudal vasculature by a needle and syringe treated with ammonium heparin. Blood samples were collected within 5 min from the initial netting. Blood was separated by centrifugation at 4 °C and plasma stored at−80 °C until analyses. Body mass and fork length were measured for calculation of condition factor: (body mass, g)/(fork length, cm)3 × 100. Gill and liver tissues were collected and immediately frozen on dry ice and stored at−80 °C. Four to six additional gill filaments were placed in ice-cold SEI buffer (150 mM sucrose, 10 mM EDTA, 50 mM imidazole, pH 7.3) and stored at−80 °C.
Plasma and gill analyses
Plasma Gh levels were measured by a radioimmunoassay (RIA) validated for Atlantic salmon by Björnsson et al. [37]. Plasma Igf1 levels were measured by a RIA validated for salmonids [38]. Plasma chloride was analyzed by the silver titration method using a Buchler-Cotlove digital chloridometer (Labconco, Kansas City, MO) and external standards. Branchial Nka activity was determined as described by McCormick [39]. Protein concentration of the gill homogenate was determined using a BCA protein assay (Thermo Fisher Scientific, Rockford, IL).
RNA extraction, cDNA synthesis and quantitative real-time PCR (qRT-PCR)
Total RNA was extracted from tissues by the TRI Reagent procedure (MRC, Cincinnati, OH) according to the manufacturer’s protocols. RNA concentration and purity were assessed by spectrophotometric absorbance (Nanodrop 1000, Thermo Scientific, Wilmington, DE). First strand cDNA was synthesized with a High Capacity cDNA Reverse Transcription Kit (Life Technologies, Carlsbad, CA). Relative mRNA levels were determined by qRT-PCR using the StepOnePlus real-time PCR system (Life Technologies). We employed previously described primer pairs for ghr1 [23], igf1, igf2, igf receptor 1a (igfr1a) and elongation factor 1α (ef1α) [40], igfbp1a1,−1b1,−1b2,−2a,−2b1,−2b2,−4,−5a,−5b1,−5b2,−6b1 and−6b2 [33], and nkcc1 and cftr1 [11]. qRT-PCR reactions were setup in a 15 μl final reaction volume with 400 nM of each primer, 1 μl cDNA and 7.5 μl of 2× SYBR Green PCR Master Mix (Life Technologies). The following cycling parameters were employed: 10 min at 95 °C followed by 40 cycles at 95 °C for 15 s, 60 °C for 30 s and 72 °C for 30 s. After verification that levels did not vary across groups, ef1α levels were used to normalize target genes. Reference and target gene levels were calculated by the relative quantification method with PCR efficiency correction [41]. Standard curves were prepared from serial dilutions of gill or liver cDNA and included on each plate to calculate the PCR efficiencies for target and normalization genes (>90%). Relative mRNA levels are reported as a fold-change from the March 3 group (Figs. 1–3; Table 1) or 0 h groups (Figs. 4–6; Table 2).
Fig. 1.
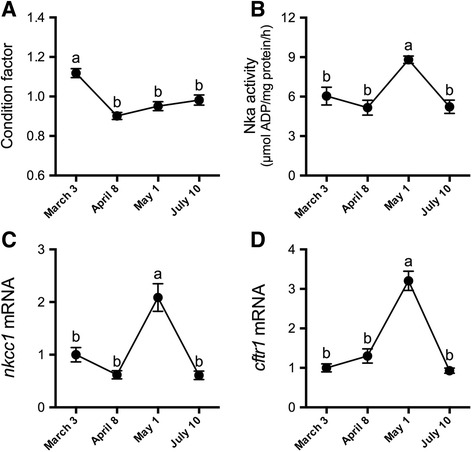
Seasonal dynamics of condition factor and ionoregulatory parameters. Condition factor (a) and branchial Nka activity (b), nkcc1 (c) and cftr1 (d) mRNA levels in Atlantic salmon maintained in FW from March 3 through July 10. Means ± S.E.M. (n = 8). mRNA levels are presented as a fold-change from the March 3 group. Means not sharing the same letter are significantly different (one-way ANOVA, Tukey’s HSD test, P < 0.05)
Fig. 3.
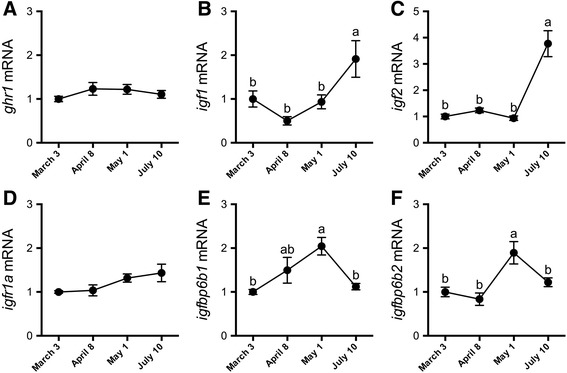
Seasonal dynamics of branchial gene expression. Branchial ghr1 (a), igf1 (b), igf2 (c), igfr1a (d), igfbp6b1 (e) and -6b2 (f) mRNA levels in Atlantic salmon maintained in FW from March 3 through July 10. Means ± S.E.M. (n = 8). mRNA levels are presented as a fold-change from the March 3 group. Means not sharing the same letter are significantly different (one-way ANOVA, Tukey’s HSD test, P < 0.05)
Table 1.
Branchial and hepatic mRNA levels in Atlantic salmon maintained in FW from March 3 through July 10
| Target Gene | March 3 | April 8 | May 1 | July 10 |
|---|---|---|---|---|
| Gill | ||||
| igfbp4 | 1.00 ± 0.08b | 1.06 ± 0.20b | 1.49 ± 0.15a,b | 1.61 ± 0.10a |
| igfbp5a | 1.00 ± 0.15a,b | 0.79 ± 0.14b | 1.51 ± 0.18a | 0.73 ± 0.05b |
| igfbp5b1 | 1.00 ± 0.06a,b | 0.47 ± 0.06c | 1.17 ± 0.14a | 0.70 ± 0.09b,c |
| igfbp5b2 | 1.00 ± 0.12a,b | 0.57 ± 0.07c | 1.13 ± 0.06a | 0.76 ± 0.05b,c |
| Liver | ||||
| ghr1 | 1.00 ± 0.30b | 2.75 ± 0.33a | 1.09 ± 0.16b | 0.38 ± 0.08b |
| igf1 | 1.00 ± 0.21b | 3.04 ± 0.71a | 3.49 ± 0.15a | 3.25 ± 0.35a |
| igf2 | 1.00 ± 0.27 | 0.91 ± 0.31 | 0.56 ± 0.11 | 0.46 ± 0.07 |
| igfbp1a1 | 1.00 ± 0.45 | 0.69 ± 0.37 | 0.10 ± 0.04 | 0.02 ± 0.004 |
| igfbp1b1 | 1.00 ± 0.21b | 2.50 ± 0.47a | 1.05 ± 0.14b | 0.36 ± 0.02b |
| igfbp1b2 | 1.00 ± 0.16b | 5.75 ± 1.26a | 2.01 ± 0.27b | 0.21 ± 0.04b |
| igfbp2a | 1.00 ± 0.10 | 0.83 ± 0.07 | 0.74 ± 0.06 | 0.73 ± 0.07 |
| igfbp2b1 | 1.00 ± 0.10a | 0.51 ± 0.06b | 0.44 ± 0.05b | 0.44 ± 0.03b |
| igfbp2b2 | 1.00 ± 0.12 | 1.29 ± 0.15 | 0.97 ± 0.09 | 0.90 ± 0.02 |
Expression levels are presented as a fold-change from the March 3 group. Means ± S.E.M. (n = 8). For a given transcript, means not sharing the same superscript letter are significantly different (one-way ANOVA, Tukey’s HSD test, P < 0.05)
Fig. 4.
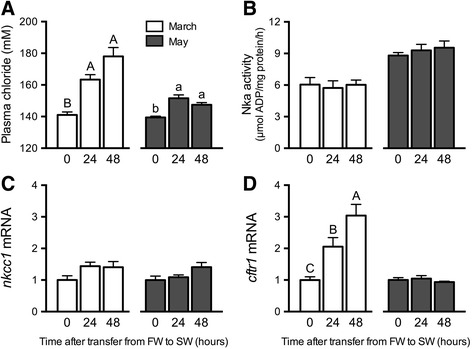
Effects of SW exposure on ionoregulatory parameters. Plasma chloride (a) and branchial Nka activity (b), nkcc1 (c) and cftr1 (d) mRNA levels in Atlantic salmon subjected to 24 and 48 h SW exposures in March (open bars) and May (shaded bars). Means ± S.E.M. (n = 8). mRNA levels are presented as a fold-change from the 0 h groups. Within a given experiment, denoted by uppercase or lowercase letters, means not sharing the same letter are significantly different (one-way ANOVA, Tukey’s HSD test, P < 0.05)
Fig. 6.
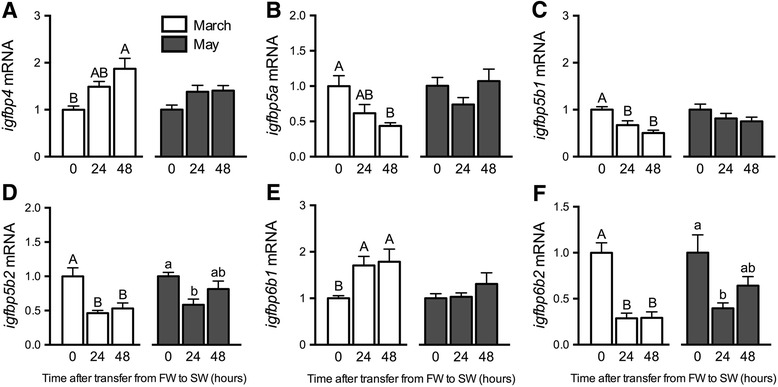
Effects of SW exposure on igfbp gene expression. Branchial igfbp4 (a),−5a (b),−5b1 (c),−5b2 (d),−6b1 (e) and−6b2 (f) mRNA levels in Atlantic salmon exposed to 24 and 48 h SW exposures in March (open bars) and May (shaded bars). Means ± S.E.M. (n = 8). Within a given experiment, denoted by uppercase or lowercase letters, means not sharing the same letter are significantly different (one-way ANOVA, Tukey’s HSD test, P < 0.05)
Table 2.
Hepatic mRNA levels in Atlantic salmon exposed to 24 and 48 h SW exposures in two separate experiments (March and May)
| Target Gene | Experiment | 0 h | 24 h | 48 h |
|---|---|---|---|---|
| ghr1 | March | 1.00 ± 0.30a,b | 0.36 ± 0.04b | 1.95 ± 0.55a |
| May | 1.00 ± 0.14 | 5.38 ± 4.00 | 1.79 ± 0.25 | |
| igf1 | March | 1.00 ± 0.21 | 0.67 ± 0.22 | 0.84 ± 0.30 |
| May | 1.00 ± 0.04b | 0.61 ± 0.17b | 1.55 ± 0.15a | |
| igf2 | March | 1.00 ± 0.27 | 0.63 ± 0.14 | 0.72 ± 0.20 |
| May | 1.00 ± 0.19 | 4.39 ± 3.35 | 0.95 ± 0.27 | |
| igfbp1a1 | March | 1.00 ± 0.44b | 6.06 ± 2.81a,b | 20.15 ± 8.67a |
| May | 1.00 ± 0.39 | 20.26 ± 10.23 | 144.97 ± 88.44 | |
| igfbp1b1 | March | 1.00 ± 0.21b | 4.45 ± 0.39a | 4.55 ± 1.15a |
| May | 1.00 ± 0.13 | 0.79 ± 0.09 | 1.38 ± 0.47 | |
| igfbp1b2 | March | 1.00 ± 0.16 | 2.41 ± 0.31 | 2.79 ± 0.89 |
| May | 1.00 ± 0.14a | 0.52 ± 0.04b | 1.24 ± 0.21a | |
| igfbp2a | March | 1.00 ± 0.10 | 0.98 ± 0.22 | 0.84 ± 0.17 |
| May | 1.00 ± 0.09 | 0.85 ± 0.11 | 1.27 ± 0.23 | |
| igfbp2b1 | March | 1.00 ± 0.10 | 0.91 ± 0.06 | 0.65 ± 0.12 |
| May | 1.00 ± 0.11 | 1.02 ± 0.14 | 1.53 ± 0.32 | |
| igfbp2b2 | March | 1.00 ± 0.12 | 0.87 ± 0.06 | 0.97 ± 0.13 |
| May | 1.00 ± 0.09 | 0.82 ± 0.10 | 0.91 ± 0.19 |
Expression levels are presented as a fold-change from the 0 h groups. Means ± S.E.M. (n = 8). Within an experiment, means not sharing the same superscript letter are significantly different (one-way ANOVA, Tukey’s HSD test, P < 0.05)
Statistics
Group comparisons were performed by one-way ANOVA followed by Tukey’s HSD test. Significance for all tests was set at P < 0.05. All statistical analyses were performed using GraphPad Prism 6 (San Diego, CA).
Results
Developmental/seasonal patterns
We verified that parr-smolt transformation and subsequent loss of salinity tolerance occurred by profiling multiple morphological and ionoregulatory parameters. Condition factor was significantly reduced on April 8, May 1 and July 10 compared with March 3 (Fig. 1a). We observed progressive silvering of the body and darkening of the fin margins leading to the May 1 sampling (data not shown). Branchial Nka activity was elevated in May compared with pre-smolts (March 3 and April 8) and post-smolts (July 10) (Fig. 1b). Branchial nkcc1 and cftr1 levels were elevated in May compared with all other sampled time points (Fig. 1c, d).
There was a significant effect of season on plasma Gh (one-way ANOVA; P < 0.001), but no significant differences between sampling points were detected by post hoc analysis (Fig. 2a). Plasma Igf1 was elevated in July when compared with April levels (Fig. 2b).
Fig. 2.
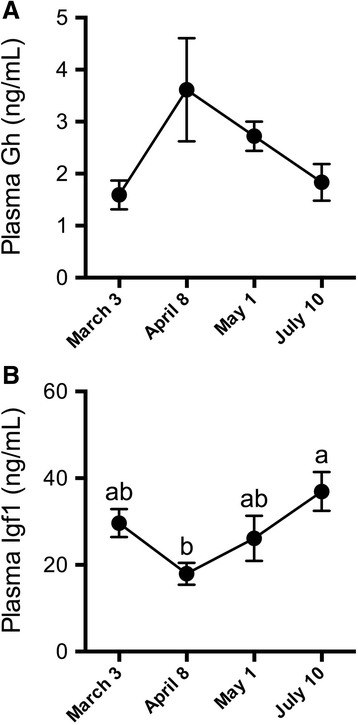
Seasonal dynamics of plasma hormones. Plasma Gh (a) and Igf1 (b) levels in Atlantic salmon maintained in FW from March 3 through July 10. Means ± S.E.M. (n = 8). Means not sharing the same letter are significantly different (one-way ANOVA, Tukey’s HSD test, P < 0.05)
Parr-smolt transformation did not coincide with any differences in branchial ghr1 or igfr1a expression (Fig. 3a, d). Both igf1 and−2 were elevated in post-smolts (July 10) compared with all preceding time points (Fig. 3b, c). Branchial igfbp6b1 and−6b2 (Fig. 3e, f) were significantly elevated near the peak of smoltification (May 1) while there were no clear seasonal effects on igfbp4,−5a, 5b1 and−5b2 (Table 1). In liver, ghr1 expression was elevated in April above all other time points, while igf1 expression was elevated above March 3 levels at all subsequent samplings. There were no clear seasonal effects on hepatic igf2. Hepatic igfbp1b1 and−1b2 were elevated in April above all other time points; igfbp2b1 was reduced from March 3 levels at all subsequent samplings (Table 1).
Seawater exposures in March and May
In both March and May, SW exposures elicited increases in plasma chloride at 24 and 48 h (Fig. 4a). The increase in plasma chloride after SW exposure was substantially greater in March than in May. There were no significant increases in branchial Nka activity or nkcc1 after SW exposure in March or May (Fig. 4b, c). SW exposure induced branchial cftr1 expression in March, but not in May (Fig. 4d).
Plasma Gh levels were elevated by 48 h after SW exposure in March; Gh levels did not respond to SW exposure in May (Fig. 5a). SW exposures did not elicit any changes in plasma Igf1 (Fig. 5b). SW induced branchial ghr1 levels by 48 and 24 h in March and May, respectively (Fig. 5c). Branchial igf1 was not responsive to SW in March or May (Fig. 5d) whereas SW induced igf2 in both March and May (Fig. 5e). In March, igfr1a showed modest increases in response to SW (Fig. 5 f).
Fig. 5.
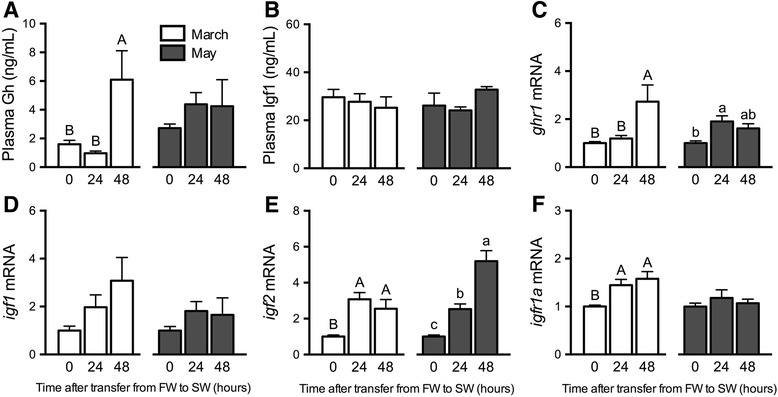
Effects of SW exposure on plasma hormones and branchial gene expression. Plasma Gh (a), Igf1 (b), and branchial ghr1 (c), igf1 (d), igf2 (e) and igfr1a (f) mRNA levels in Atlantic salmon exposed to 24 and 48 h SW exposures in March (open bars) and May (shaded bars). Means ± S.E.M. (n = 8). Within a given experiment, denoted by uppercase or lowercase letters, means not sharing the same letter are significantly different (one-way ANOVA, Tukey’s HSD test, P < 0.05)
Among the igfbps expressed in the gill, igfbp4 and−6b1 were induced by SW exposure in March (Fig. 6A, E), while igfbp5a,−5b1,−5b2 and−6b2 were diminished following SW exposure (Fig. 6B-D, F). As in March, igfbp6b2 was decreased following SW exposure in May (Fig. 6 f). In liver, there were no clear effects of SW exposures on ghr1 and igf2; however, SW induced igf1 in May. igfbp1a1 and−1b1 were similarly induced by SW exposure in March (Table 2).
Discussion
The progressive increase in the salt secretory capacity of the gill during smoltification entails developmentally cued patterns of ionocyte differentiation and proliferation, in addition to altered gene transcription within these ionocytes [8, 42, 43]. Knowing that the Gh/Igf axis directs the timing and nature of these cellular behaviors [13–15], we hypothesized that Igfbps contribute to smoltification and therefore would exhibit seasonal and SW-responsive patterns of gene expression. We report for the first time that increases in igfbp6b1 and−6b2 expression coincided with parr-smolt transformation and multiple igfbp4,−5 and−6 isoforms were modulated following SW exposures at different stages of smolt development.
Fish in this study underwent smoltification as indicated by multiple parameters. First, we observed the typical decrease in condition factor due to changes in body shape and the utilization of lipid and glycogen stores [36, 44–47]. In strong agreement with previous studies, branchial Nka activity and nkcc1 and cftr1 expression concomitantly peaked in May, a hallmark of SW-type ionocyte recruitment [10, 11, 20, 36, 47, 48]. The ability of juvenile salmon to maintain ionoregulatory balance upon direct transfer from FW to SW is readily employed as an operational definition of hyposmoregulatory capacity. In March (pre-smolts) we observed relatively large increases in plasma chloride after SW exposure, whereas in May (smolts), we observed modest increases in plasma chloride after SW exposure. It is interesting that in March, when fish had not yet developed SW tolerance, only cftr1, and not nkcc1, was activated in parallel with chloride perturbations. The branchial epithelium of pre-smolts harbors a population of SW-type ionocytes that presumably employ Cftr1 and Nkcc1 in the apical and basolateral cell membrane, respectively [8, 49, 50]. Within these cells, transcription of cftr1 may be rapidly activated by ionic/osmotic conditions (environmental or internal), a pattern reminiscent of how chloride secretion is activated in opercular epithelia of Fundulus heteroclitus [51]. In any event, the attendant changes in the gill with respect to ionocyte function are consistent with a developmental/seasonal increase in ion-secretion capacity.
Photoperiod-induced increases in plasma Gh levels coincide with Atlantic salmon smoltification [52] and we likewise observed a rise (albeit not significant following post hoc analysis) in plasma Gh levels in April. Nilsen et al. [53] observed increased plasma Gh levels in SW-challenged smolts in May, whereas we observed a Gh response in March. This Gh response was paralleled by increased branchial ghr1 expression. Kiilerich et al. [20] similarly observed increased ghr1 with SW transfer, albeit in smolts transferred to SW in April. While not specifically shown in salmonids yet, Gh release from the pituitary is induced by direct osmosensing in certain euryhaline species such as Mozambique tilapia (Oreochromis mossambicus) [54]. This mode of regulation is compatible with increased plasma Gh levels when blood plasma conditions, such as plasma chloride and presumably osmolality, were perturbed following SW transfer. The osmoregulatory actions of Gh are mediated by its capacity to increase circulating levels and local tissue production of Igfs [14]. Seasonal patterns of circulating Igf1 in Atlantic salmon smolts are variable. In some cases, increases [36, 55], decreases [53], or no well-defined changes [56] have been reported. While we did not observe increased plasma Igf1 in smolts, we did observe increased hepatic igf1 expression, perhaps mediated by increased sensitivity to Gh via upregulation of ghr1. On the other hand, local igf1 and−2 expression in the gill was not elevated in May smolts, patterns important to consider in light of the igfbp responses we subsequently observed.
This is the first time branchial igfbps have been assessed in a salmonid preparing for seaward migration; we assayed igfbp4,−5 and−6 isoforms that exhibit robust branchial expression [33]. igfbp4 exhibited a steady rise in expression throughout spring and summer, with increased expression following SW exposure in March. The function of Igfbp4, at least in mammals, highly depends on the physiological context surrounding its production, and may operate as either a stimulator or inhibitor of Igf1/2 signaling [57, 58]. The activities of a teleost Igfbp4 were first assessed in fugu (Takifugu rubripes) where overexpression delayed embryonic development [59]. Nonetheless, in Atlantic salmon and sea bream (Sparus aurata), igfbp4 expression is implicated in mediating enhanced post-prandial/fasting muscle growth [40, 60–62], thereby suggesting a stimulatory effect on Igf activity. The concomitant increases of igfbp4 along with igf2 and igfr1a following SW exposure may reflect a transcriptional program underlying enhanced paracrine signaling in response to ionoregulatory demands.
In contrast with igfbp4, igfbp5a,−5b1 and−5b2 were all reduced following SW exposure in March. Similar to Atlantic salmon igfbp5s, zebrafish (Danio rerio) igfbp5a and−5b are expressed in the gill [63]. igfbp5a is expressed in a sub-population of zebrafish ionocytes termed “NaR cells” specialized for Ca2+ uptake via Trpv5/6 channels. igfbp5a plays an essential role in Ca2+ homeostasis; igfbp5a expression is induced by low environmental [Ca2+] and igfbp5a knockdown inhibits compensatory increases in NaR cell proliferation following reductions in [Ca2+] [31]. While not yet established for Atlantic salmon, Ca2+ uptake across rainbow trout (Oncorhynchus mykiss) gill epithelia similarly employs a Trpv5/6 channel expressed in ionocytes and pavement cells [64]. If Igfbp5a is a conserved regulator of branchial Ca2+ uptake, then the SW-induced reductions in igfbp5a we observed in this study may reflect the increased [Ca2+] of SW compared with FW, and subsequent down regulation of Ca2+ uptake pathways. Interestingly, Dai et al. [63] showed that among the zebrafish Igfbp5 isoforms (−5a and−5b), only Igfbp5b exhibits ligand-independent transactivational activity. Thus, while igfbp5a,−5b1, and−5b2 showed similar responses to SW in the current study, it is likely that they are functionally distinct from one another, but such distinctions are entirely unresolved to date.
Wang et al. [65] described two teleost co-orthologs of human Igfbp6, denoted Igfbp6a and−6b. igfbp6a exhibits low expression in both zebrafish and Atlantic salmon gill, with igfbp6b2 being highly expressed in salmon gill [33, 65]. Among the igfbps we assayed, igfbp6b1 and−6b2 showed seasonal increases in expression with maximum levels in May smolts. Mammalian Igfbp6 exhibits a higher binding affinity for Igf2 versus Igf1, and inhibits Igf actions [66]. Similarly, zebrafish Igfbp6a and−6b attenuate Igf activities and embryonic growth and development [65]. There is currently no information on plasma Igf2 dynamics during smoltification; however, locally produced Igfbp6b1 and/or−6b2 may modulate Igf2 activity in the gill. Moreover, Igfbp6 modulates cell proliferation, migration, and apoptosis in mammalian systems [66, 67], and considering how cell turnover underlies branchial development during smoltification [42], Igfbp6s may similarly contribute to cell-cycle regulation in smolts. Adding further complexity is the disparate regulation of the two igfbp6 isoforms following SW exposures. Nonetheless, the seasonal patterns of both igfbp6s suggest future study of their role(s) in the gill is warranted.
We also assayed hepatic igfbp1 and−2 isoforms because their translated products modulate endocrine Igfs [26]. As in mammals, igfbp1 and−2 isoforms are highly expressed in teleost liver [33, 68–74]. Igfbp1 inhibits somatic growth, development, and glucose metabolism by restricting Igfs from binding Igf receptors [69, 75, 76]. The only report to date of plasma Igfbp dynamics in smolts (coho salmon; Oncorhychus kisutch) revealed an April peak in plasma Igfbp1 [77]. This elevated Igfbp1 coincided with a drop in condition factor. Here, we observed 2.5- and 5.6-fold increases in igfbp1b1 and -1b2 expression, respectively, in April compared with March. Recall that we also observed a decline in condition factor in early spring, a pattern that routinely occurs when Atlantic salmon smolts, but not parr, are allowed to feed ad libitum [36]. Reduced condition factor is due to both smoltification-related changes in body shape and the utilization of energy reserves such as lipid stores and liver glycogen [44, 45, 47]. Previous work has established that Gh is involved in the lipolysis that occurs during smolting, and likely interacts with cortisol to affect other catabolic changes [78]. When these patterns are further considered with the feeding ecology of migrating smolts [46, 79, 80], smoltification emerges as inherently catabolic. Thus, igfbp1b1 and−1b2 may be further modulating growth and metabolism as part of the metabolic demands of smolt development and in preparation for seaward migration. Interestingly, we detected no seasonal changes in igfbp1a1, an isoform that is sensitive to nutrient conditions [81]. Hepatic igfbp1a1 was, however, induced by SW exposure, a response also seen with a 32-kD Igfbp (putative Igfbp1) in rainbow trout plasma [30, 82]. Collectively, these patterns suggest that duplicated igfbp1s allow for multifactorial control of Igf signaling during development and in response to salinity change. Future work should investigate whether divergent igfbp1 responses are aligned with contrasting sensitivities to hormones such as cortisol, thyroid hormones and insulin, which display seasonal changes and/or mediate stress responses [83–85].
With a subset of hepatic and branchial igfbps whose expression patterns parallel parr-smolt transformation now revealed, future study is warranted to resolve how these dynamics relate to changes in circulating levels of actual Igfbp proteins. As the liver is regarded as a major source of circulating Igfbp1 [75, 86], we hypothesize that plasma Igfbp1b1 and−1b2 may be enhanced in early April provided that mRNA levels are suggestive of protein production and secretion. Moreover, with marked changes in branchial igfbp6b1 and−6b2 levels occurring in May, it should be resolved whether their translated products are retained (and act) locally, or whether they enter circulation as endocrine factors. In any event, the development of isoform-specific detection of Igfbps is the next step towards establishing how the complex expression patterns of Atlantic salmon igfbps across varied tissues [33] relate to local and endocrine protein levels.
Conclusions
Salmonids express an especially wide array of igfbps, and our data suggest that multiple Igfbps may contribute to the development of SW tolerance and associated metabolic changes that occur during the parr-smolt transformation. With igfbps such as igfbp6b1 and−6b2 showing increases coincident with smolt development, the challenge is to now identify the specific activities of these isoforms. By comparing the physiologies of anadromous and landlocked salmon populations, investigators have discerned how relaxed selection for SW adaptability affects both endocrine and ionoregulatory systems [11, 53, 87]. We propose that a similar approach, which compares igfbp expression patterns across Atlantic salmon populations, will aid our understanding how Igfbps operate within, and modulate, the hormonal mechanisms that drive smoltification.
Acknowledgements
We appreciate the laboratory assistance provided by Andrew Weinstock and Amy Regish during the course of this study. We also acknowledge Spencer Chicoine for assistance with plasma chloride measurements.
Funding
This work was supported by Skidmore College (Start-Up Funds; JPB). Any use of trade, product, or firm names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Authors’ contributions
JPB conceived and designed experiments, collected and analyzed qPCR data, and drafted the manuscript. CKF and SKPC collected and analyzed qPCR data. IEE collected and analyzed plasma hormone data. BThB analyzed plasma hormone data and coordinated the study. SDM conceived and designed experiments and coordinated the study. All authors contributed to revising manuscript drafts and approved the final version.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Ethics approval and consent to participate
All experiments were carried out in accordance with USGS institutional guidelines and approved by the USGS, Leetown Science Center Institutional Animal Care and Use Committee (Protocol SP9065).
Abbreviations
- Cftr
Cystic fibrosis transmembrane regulator
- Ef1α
Elongation factor 1α
- FW
Fresh water
- Gh
Growth hormone
- Ghr
Growth hormone receptor
- Igf
Insulin-like growth-factor
- Igfbp
Insulin-like growth-factor binding protein
- Igfr
Insulin-like growth-factor receptor
- Nka
Na+/K+-ATPase
- Nkcc
Na+/K+/2Cl− cotransporter
- qRT-PCR
Quantitative real-time PCR
- RIA
Radioimmunoassay
- SW
Seawater
Contributor Information
Jason P. Breves, Phone: (518)-580-5079, Email: jbreves@skidmore.edu
Chelsea K. Fujimoto, Email: cfkalila@gmail.com
Silas K. Phipps-Costin, Email: sphippsc@skidmore.edu
Ingibjörg E. Einarsdottir, Email: ingibjorg.einarsdottir@bioenv.gu.se
Björn Thrandur Björnsson, Email: thrandur.bjornsson@bioenv.gu.se.
Stephen D. McCormick, Email: mccormick@umext.umass.edu
References
- 1.Hoar WS. The physiology of smolting salmonids. In: Hoar WS, Randall DJ, editors. Fish Physiology. New York: Academic; 1988. pp. 275–343. [Google Scholar]
- 2.McCormick SD. Ontogeny and evolution of salinity tolerance in anadromous salmonids: hormones and heterochrony. Estuaries. 1994;17:26–33. doi: 10.2307/1352332. [DOI] [Google Scholar]
- 3.McCormick SD, Shrimpton JM, Moriyama S, Björnsson BT. Effects of an advanced temperature cycle on smolt development and endocrinology indicate that temperature is not a zeitgeber for smolting in Atlantic salmon. J Exp Biol. 2002;205:3553–60. doi: 10.1242/jeb.205.22.3553. [DOI] [PubMed] [Google Scholar]
- 4.Boeuf G. Salmonid smolting: a pre-adaptation to the oceanic environment. In: Rankin JC, Jensen FB, editors. Fish ecophysiology. London: Chapman and Hall; 1993. pp. 105–135. [Google Scholar]
- 5.McCormick SD, Hansen LP, Quinn TP, Saunders RL. Movement, migration, and smolting of Atlantic salmon (Salmo salar) Can J Fish Aquat Sci. 1998;55(Suppl 1):77–92. doi: 10.1139/d98-011. [DOI] [Google Scholar]
- 6.Johnstone KA, Lubieniecki KP, Koop BF, Davidson WS. Expression of olfactory receptors in different life stages and life histories of wild Atlantic salmon (Salmo salar) Mol Ecol. 2011;20:4059–69. doi: 10.1111/j.1365-294X.2011.05251.x. [DOI] [PubMed] [Google Scholar]
- 7.Marshall WS, Grosell M. Ion transport, osmoregulation, and acid–base balance. In: Evans DH, Claiborne JB, editors. The physiology of fishes. Boca Raton: CRC Press; 2006. pp. 177–230. [Google Scholar]
- 8.Pelis RM, McCormick SD. Effects of growth hormone and cortisol on Na+-K+-2Cl− cotransporter localization and abundance in the gills of Atlantic salmon. Gen Comp Endocrinol. 2001;124:134–43. doi: 10.1006/gcen.2001.7703. [DOI] [PubMed] [Google Scholar]
- 9.Singer TD, Clements KM, Semple JW, Schulte PM, Bystriansky JS, Finstad B, et al. Seawater tolerance and gene expression in two strains of Atlantic salmon smolts. Can J Fish Aquat Sci. 2002;59:125–35. doi: 10.1139/f01-205. [DOI] [Google Scholar]
- 10.Tipsmark CK, Madsen SS, Seidelin M, Christensen AS, Cutler CP, Cramb G. Dynamics of Na+, K+,2Cl − cotransporter and Na+, K + -ATPase expression in the branchial epithelium of brown trout (Salmo trutta) and Atlantic salmon (Salmo salar) J Exp Zool. 2002;293:106–18. doi: 10.1002/jez.10118. [DOI] [PubMed] [Google Scholar]
- 11.Nilsen TO, Ebbesson LOE, Madsen SS, McCormick SD, Andersson E, Björnsson BT, et al. Differential expression of gill Na+/K+-ATPase α- and β-subunits, Na+/K+/2Cl− cotransporter and CFTR anion channel in juvenile anadromous and landlocked Atlantic salmon Salmo salar. J Exp Biol. 2007;210:2885–96. doi: 10.1242/jeb.002873. [DOI] [PubMed] [Google Scholar]
- 12.McCormick SD, Regish AM, Christensen AK, Björnsson BT. Differential regulation of sodium-potassium pump isoforms during smolt development and seawater exposure of Atlantic salmon. J Exp Biol. 2013;216:1142–51. doi: 10.1242/jeb.080440. [DOI] [PubMed] [Google Scholar]
- 13.Björnsson BT. The biology of salmon growth hormone: from daylight to dominance. Fish Physiol Biochem. 1997;17:9–24. doi: 10.1023/A:1007712413908. [DOI] [Google Scholar]
- 14.Sakamoto T, McCormick SD, Hirano T. Osmoregulatory actions of growth hormone and its mode of action in salmonids: A review. Fish Physiol Biochem. 1993;11:155–64. doi: 10.1007/BF00004562. [DOI] [PubMed] [Google Scholar]
- 15.McCormick SD. Endocrine control of osmoregulation in teleost fish. Amer Zool. 2001;41:781–94. [Google Scholar]
- 16.McCormick SD, Dickhoff WW, Duston J, Nishioka RS, Bern HA. Developmental differences in the responsiveness of gill Na+, K + -ATPase to cortisol in salmonids. Gen Comp Endocrinol. 1991;84:308–17. doi: 10.1016/0016-6480(91)90054-A. [DOI] [PubMed] [Google Scholar]
- 17.Madsen SS, Bern HA. In-vitro effects of insulin-like growth factor-I on gill Na+,K+-ATPase in coho salmon, Oncorhynchus kisutch. J Endocrinol. 1993;138:23–30. doi: 10.1677/joe.0.1380023. [DOI] [PubMed] [Google Scholar]
- 18.Sakamoto T, Hirano T. Expression of insulin-like growth factor I gene in osmoregulatory organs during seawater adaptation of the salmonid fish: possible mode of osmoregulatory action of growth hormone. Proc Natl Acad Sci U S A. 1993;90:1912–16. doi: 10.1073/pnas.90.5.1912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Madsen SS, Jensen MK, Nhr J, Kristiansen K. Expression of Na+-K+-ATPase in the brown trout, Salmo trutta: in vivo modulation by hormones and seawater. Am J Physiol. 1995;269:R1339–45. doi: 10.1152/ajpregu.1995.269.6.R1339. [DOI] [PubMed] [Google Scholar]
- 20.Kiilerich P, Kristiansen K, Madsen SS. Hormone receptors in gills of smolting Atlantic salmon, Salmo salar: expression of growth hormone, prolactin, mineralocorticoid and glucocorticoid receptors and 11β-hydroxysteroid dehydrogenase type 2. Gen Comp Endocrinol. 2007;152:295–303. doi: 10.1016/j.ygcen.2006.12.018. [DOI] [PubMed] [Google Scholar]
- 21.Shrimpton JM, McCormick SD. Regulation of gill cytosolic corticosteroid receptors in juvenile Atlantic salmon: Interaction effects of growth hormone with prolactin and triiodothyronine. Gen Comp Endocrinol. 1998;112:262–74. doi: 10.1006/gcen.1998.7172. [DOI] [PubMed] [Google Scholar]
- 22.McCormick SD. Effects of growth hormone and insulin-like growth factor I on salinity tolerance and gill Na+/K+-ATPase in Atlantic salmon (Salmo salar): interaction with cortisol. Gen Comp Endocrinol. 1996;101:3–11. doi: 10.1006/gcen.1996.0002. [DOI] [PubMed] [Google Scholar]
- 23.Tipsmark CK, Madsen SS. Distinct hormonal regulation of Na+, K + -atpase genes in the gill of Atlantic salmon (Salmo salar L.) J Endocrinol. 2009;203:301–10. doi: 10.1677/JOE-09-0281. [DOI] [PubMed] [Google Scholar]
- 24.Madsen SS. The role of cortisol and growth hormone in seawater adaptation and development of hypoosmoregulatory mechanisms in sea trout parr (Salmo trutta trutta) Gen Comp Endocrinol. 1990;79:1–11. doi: 10.1016/0016-6480(90)90082-W. [DOI] [PubMed] [Google Scholar]
- 25.Prunet P, Pisam M, Claireaux JP, Boeuf G, Rambourg A. Effects of growth hormone on gill chloride cells in juvenile Atlantic salmon (Salmo salar) Am J Physiol. 1994;266:R850–7. doi: 10.1152/ajpregu.1994.266.3.R850. [DOI] [PubMed] [Google Scholar]
- 26.Duan C, Ren H, Gao S. Insulin-like growth factors (IGFs), IGF receptors, and IGF-binding proteins: roles in skeletal muscle growth and differentiation. Gen Comp Endocrinol. 2010;167:344–51. doi: 10.1016/j.ygcen.2010.04.009. [DOI] [PubMed] [Google Scholar]
- 27.Clemmons DR. Use of mutagenesis to probe IGF-binding protein structure/function relationships. Endocr Rev. 2001;22(6):800–17. doi: 10.1210/edrv.22.6.0449. [DOI] [PubMed] [Google Scholar]
- 28.Reindl KM, Sheridan MA. Peripheral regulation of the growth hormone-insulin-like growth factor system in fish and other vertebrates. Comp Biochem Physiol A. 2012;163:231–45. doi: 10.1016/j.cbpa.2012.08.003. [DOI] [PubMed] [Google Scholar]
- 29.Johnson J, Silverstein J, Wolters WR, Shimizu M, Dickhoff WW, Shepherd BS. Disparate regulation of insulin-like growth factor-binding proteins in a primitive, ictalurid, teleost (Ictalurus punctatus) Gen Comp Endocrinol. 2003;134(2):122–30. doi: 10.1016/S0016-6480(03)00244-2. [DOI] [PubMed] [Google Scholar]
- 30.Shepherd BS, Drennon K, Johnson J, Nichols JW, Playle RC, Singer TD, Vijayan MM. Salinity acclimation affects the somatotropic axis in rainbow trout. Am J Physiol Regul Integr Comp Physiol. 2005;288:R1385–95. doi: 10.1152/ajpregu.00443.2004. [DOI] [PubMed] [Google Scholar]
- 31.Dai W, Bai Y, Hebda L, Zhong X, Liu J, Kao J, Duan C. Calcium deficiency-induced and TRP channel-regulated IGF1R-PI3K-Akt signaling regulates abnormal epithelial cell proliferation. Cell Death Differ. 2014;21(4):568–81. doi: 10.1038/cdd.2013.177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Taniyama N, Kaneko N, Inatani Y, Miyakoshi Y, Shimizu M. Effects of seawater transfer and fasting on the endocrine and biochemical growth indices in juvenile chum salmon (Oncorhynchus keta) Gen Comp Endocrinol. 2016;236:146–56. doi: 10.1016/j.ygcen.2016.07.020. [DOI] [PubMed] [Google Scholar]
- 33.Macqueen DJ, García de la serrana D, Johnston IA. Evolution of ancient functions in the vertebrate insulin-like growth factor system uncovered by the study of duplicated salmonid fish genomes. Mol Biol Evol. 2013;30:1060–76. doi: 10.1093/molbev/mst017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Parrish DL, Behnke RJ, Gephard SR, McCormick SD, Reeves GH. Why aren’t there more Atlantic salmon (Salmo salar)? Can J Fish Aquat Sci. 1998;55(Suppl 1):281–87. doi: 10.1139/d98-012. [DOI] [Google Scholar]
- 35.McCormick SD, Lerner DT, Monette MY, Nieves-Puigdoller K, Kelly JT, Björnsson BT. Taking it with you when you go: how perturbations to the freshwater environment, including temperature, dams, and contaminants, affect marine survival of salmon. Am Fish Soc Symp. 2009;69:195–214. [Google Scholar]
- 36.McCormick SD, Shrimpton JM, Moriyama S, Björnsson BT. Differential hormonal responses of Atlantic salmon parr and smolt to increased daylength: A possible developmental basis for smolting. Aquaculture. 2007;273:337–44. doi: 10.1016/j.aquaculture.2007.10.015. [DOI] [Google Scholar]
- 37.Björnsson BT, Taranger GL, Hansen T, Stefansson SO, Haux C. The interrelation between photoperiod, growth hormone, and sexual maturation of adult Atlantic salmon (Salmo salar) Gen Comp Endocrinol. 1994;93:70–81. doi: 10.1006/gcen.1994.1009. [DOI] [PubMed] [Google Scholar]
- 38.Moriyama S, Swanson P, Nishii M, Takahashi A, Kawauchi H, Dickhoff WW, Plisetskaya EM. Development of homologous radioimmunoassay for coho salmon insulin-like growth factor-I. Gen Comp Endocrinol. 1994;96:149–61. doi: 10.1006/gcen.1994.1167. [DOI] [PubMed] [Google Scholar]
- 39.McCormick SD. Methods for nonlethal gill biopsy and measurement of Na+, K+,-ATPase activity. Can J Fish Aquat Sci. 1993;50:656–58. doi: 10.1139/f93-075. [DOI] [Google Scholar]
- 40.Bower NI, Li X, Taylor R, Johnston IA. Switching to fast growth: the insulin-like growth factor (IGF) system in skeletal muscle of Atlantic salmon. J Exp Biol. 2008;211:3859–70. doi: 10.1242/jeb.024117. [DOI] [PubMed] [Google Scholar]
- 41.Pfaffl MW. A new mathematical model for relative quantification in real-time RT-PCR. Nucl Acids Res. 2001;29(9):e45. doi: 10.1093/nar/29.9.e45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Laurent P, Dunel-Erb S, Chevalier C, Lignon J. Gill epithelial cells kinetics in a freshwater teleost, Oncorhynchus mykiss during adaptation to ion-poor water and hormonal treatments. Fish Physiol Biochem. 1994;13(5):353–70. doi: 10.1007/BF00003415. [DOI] [PubMed] [Google Scholar]
- 43.Robertson LS, McCormick SD. Transcriptional profiling of the parr-smolt transformation in Atlantic salmon. Comp Biochem Physiol D. 2012;7:351–60. doi: 10.1016/j.cbd.2012.07.003. [DOI] [PubMed] [Google Scholar]
- 44.Saunders RL, Henderson EB. Influence of photoperiod on smolt development and growth of Atlantic salmon (Salmo salar) J Fish Res Board Can. 1970;27:1295–1311. doi: 10.1139/f70-151. [DOI] [Google Scholar]
- 45.Sheridan MA. Alterations in lipid metabolism accompanying smoltification and seawater adaptation of salmonid fish. Aquaculture. 1989;82:191–203. doi: 10.1016/0044-8486(89)90408-0. [DOI] [Google Scholar]
- 46.Stefansson SO, Björnsson BT, Sundell L, Nyhammer G, McCormick SD. Physiological characteristics of wild Atlantic salmon post-smolts during estuarine and coastal migration. J Fish Biol. 2003;63:942–55. doi: 10.1046/j.1095-8649.2003.00201.x. [DOI] [Google Scholar]
- 47.Winans GA, Nishioka RS. A multivariate description of change in body shape of coho salmon (Oncorhynchus kisutch) during smoltification. Aquaculture. 1987;66:235–45. doi: 10.1016/0044-8486(87)90109-8. [DOI] [Google Scholar]
- 48.Mackie PM, Gharbi K, Ballantyne JS, McCormick SD, Wright PA. Na+/K+/2Cl− cotransporter and CFTR gill expression after seawater transfer in smolts (0+) of different Atlantic salmon (Salmo salar) families. Aquaculture. 2007;272:625–35. doi: 10.1016/j.aquaculture.2007.08.032. [DOI] [Google Scholar]
- 49.Marshall WS, Singer TD. Cystic fibrosis transmembrane conductance regulator in teleost fish. Biochim Biophys Acta. 2002;1566:16–27. doi: 10.1016/S0005-2736(02)00584-9. [DOI] [PubMed] [Google Scholar]
- 50.McCormick SD, Sundell K, Björnsson BT, Brown CL, Hiroi J. Influence of salinity on the localization of Na+/K+-ATPase, Na+/K+/2Cl− cotransporter (NKCC) and CFTR anion channel in chloride cells of the Hawaiian goby (Stenogobius hawaiiensis) J Exp Biol. 2003;206:4575–83. doi: 10.1242/jeb.00711. [DOI] [PubMed] [Google Scholar]
- 51.Zadunaisky JA, Cardona S, Au L, Roberts DM, Fisher E, Lowenstein B, et al. Chloride transport activation by plasma osmolarity during rapid adaptation to high salinity of Fundulus heteroclitus. J Membr Biol. 1995;143:207–17. doi: 10.1007/BF00233449. [DOI] [PubMed] [Google Scholar]
- 52.Björnsson BT, Stefansson SO, Hansen T. Photoperiod regulation of plasma growth hormone levels during parr-smolt transformation of Atlantic salmon: implications for hypoosmoregulatory ability and growth. Gen Comp Endocrinol. 1995;100:73–82. doi: 10.1006/gcen.1995.1135. [DOI] [PubMed] [Google Scholar]
- 53.Nilsen TO, Ebbesson LO, Kiilerich P, Björnsson BT, Madsen SS, McCormick SD, Stefansson SO. Endocrine systems in juvenile anadromous and landlocked Atlantic salmon (Salmo salar): seasonal development and seawater acclimation. Gen Comp Endocrinol. 2008;155:762–72. doi: 10.1016/j.ygcen.2007.08.006. [DOI] [PubMed] [Google Scholar]
- 54.Seale AP, Riley LG, Leedom TA, Kajimura S, Dores RM, Hirano T, Grau EG. Effects of environmental osmolality on release of prolactin, growth hormone and ACTH from the tilapia pituitary. Gen Comp Endocrinol. 2002;128:91–101. doi: 10.1016/S0016-6480(02)00027-8. [DOI] [PubMed] [Google Scholar]
- 55.Ágústsson T, Sundell K, Sakamoto T, Johansson V, Ando M, Björnsson BT. Growth hormone endocrinology of Atlantic salmon (Salmo salar): pituitary gene expression, hormone storage, secretion and plasma levels during parr-smolt transformation. J Endocrinol. 2001;170:227–34. doi: 10.1677/joe.0.1700227. [DOI] [PubMed] [Google Scholar]
- 56.McCormick SD, Sheehan T, Björnsson BT, Lipsky C, Kocik J, Regish AM, O’Dea MF. Physiological and endocrine changes in Atlantic salmon smolts during hatchery rearing, downstream migration and ocean entry. Can J Fish Aquat Sci. 2013;70:105–18. doi: 10.1139/cjfas-2012-0151. [DOI] [Google Scholar]
- 57.Duan C, Clemmons DR. Differential expression and biological effects of insulin-like growth factor-binding protein-4 and−5 in vascular smooth muscle cells. J Biol Chem. 1998;273(27):16836–42. doi: 10.1074/jbc.273.27.16836. [DOI] [PubMed] [Google Scholar]
- 58.Ning Y, Schuller AG, Conover CA, Pintar JE. Insulin-like growth factor (IGF) binding protein-4 is both a positive and negative regulator of IGF activity in vivo. Mol Endocrinol. 2008;22:1213–25. doi: 10.1210/me.2007-0536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Li M, Li Y, Lu L, Wang X, Gong Q, Duan C. Structural, gene expression, and functional analysis of the fugu (Takifugu rubripes) insulin-like growth factor binding protein-4 gene. Am J Physiol Regul Integr Comp Physiol. 2009;296(3):R558–66. doi: 10.1152/ajpregu.90439.2008. [DOI] [PubMed] [Google Scholar]
- 60.Bower NI, Johnston IA. Transcriptional regulation of the IGF-signaling pathway by amino acids and insulin-like growth factors during myogenesis in Atlantic salmon. PLoS One. 2010;5(6):e11100. doi: 10.1371/journal.pone.0011100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.García de la serrana D, Codina M, Jiménez-Amilburu V, Navarro I, Du SJ, et al. Characterisation and expression of myogenesis regulatory factors during in vitro myoblast development and in vivo fasting in the gilthead sea bream (Sparus aurata) Comp Biochem Physiol A. 2014;167:90–9. doi: 10.1016/j.cbpa.2013.10.020. [DOI] [PubMed] [Google Scholar]
- 62.Valente LMP, Bower NI, Johnston IA. Postprandial expression of growth related genes in Atlantic salmon (Salmo salar L.) juveniles fasted for 1 week and fed a single meal to satiation. Br J Nutr. 2012;2:1–10. doi: 10.1017/S0007114512000396. [DOI] [PubMed] [Google Scholar]
- 63.Dai W, Kamei H, Zhao Y, Ding J, Du Z, Duan C. Duplicated zebrafish insulin-like growth factor binding protein-5 genes with split functional domains: evidence for evolutionarily conserved IGF binding, nuclear localization, and transactivation activity. FASEB J. 2010;24(6):2020–9. doi: 10.1096/fj.09-149435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Shahsavarani A, McNeill B, Galvez F, Wood CM, Goss GG, Hwang PP, Perry SF. Characterization of a branchial epithelial calcium channel (ECaC) in freshwater rainbow trout (Oncorhynchus mykiss) J Exp Biol. 2006;209:1928–43. doi: 10.1242/jeb.02190. [DOI] [PubMed] [Google Scholar]
- 65.Wang X, Lu L, Li Y, Li M, Chen C, Feng Q, et al. Molecular and functional characterization of two distinct IGF binding protein-6 genes in zebrafish. Am J Physiol Regul Integr Comp Physiol. 2009;296:R1348–57. doi: 10.1152/ajpregu.90969.2008. [DOI] [PubMed] [Google Scholar]
- 66.Gallicchio MA, Kneen M, Hall C, Scott AM, Bach LA. Overexpression of insulin-like growth factor binding protein-6 inhibits rhabdomyosarcoma growth in vivo. Int J Cancer. 2001;94(5):645–51. doi: 10.1002/ijc.1519. [DOI] [PubMed] [Google Scholar]
- 67.Fu P, Thompson JA, Bach LA. Promotion of cancer cell migration: an insulin-like growth factor (IGF)-independent action of IGF-binding protein-6. J Biol Chem. 2007;282(31):22298–306. doi: 10.1074/jbc.M703066200. [DOI] [PubMed] [Google Scholar]
- 68.Funkenstein B, Tsai W, Maures T, Duan C. Ontogeny, tissue distribution, and hormonal regulation of insulin-like growth factor binding protein-2 (IGFBP-2) in a marine fish, Sparus aurata. Gen Comp Endocrinol. 2002;128:112–22. doi: 10.1016/S0016-6480(02)00059-X. [DOI] [PubMed] [Google Scholar]
- 69.Kamei H, Lu L, Jiao S, Li Y, Gyrup C, Laursen LS, et al. Duplication and diversification of the hypoxia-inducible IGFBP-1 gene in zebrafish. PLoS One. 2008;3(8):e3091. doi: 10.1371/journal.pone.0003091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Zhou J, Li W, Kamei H, Duan C. Duplication of the IGFBP-2 gene in the teleost fish: protein structure and functionality conservation and gene expression divergence. PLoS One. 2008;3(12):e3926. doi: 10.1371/journal.pone.0003926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Pedroso FL, Fukada H, Masumoto T. Molecular characterization, tissue distribution patterns and nutritional regulation of IGFBP-1,−2,−3 and−5 in yellowtail, Seriola quinqueradiata. Gen Comp Endocrinol. 2009;161:344–53. doi: 10.1016/j.ygcen.2009.01.010. [DOI] [PubMed] [Google Scholar]
- 72.Shimizu M, Kishimoto K, Yamaguchi T, Nakano Y, Hata A, Dickhoff WW. Circulating salmon 28- and 22-kDa insulin-like growth factor binding protein (IGFBPs) are co-orthologs of IGFBP-1. Gen Comp Endocrinol. 2011;174:97–106. doi: 10.1016/j.ygcen.2011.08.005. [DOI] [PubMed] [Google Scholar]
- 73.Shimizu M, Suzuki S, Horikoshi M, Hara A, Dickhoff WW. Circulating salmon 41-kDa insulin-like growth factor binding protein (IGFBP) is not IGFBP-3 but an IGFBP-2 subtype. Gen Comp Endocrinol. 2011;171:326–31. doi: 10.1016/j.ygcen.2011.02.013. [DOI] [PubMed] [Google Scholar]
- 74.Safian D, Fuentes EN, Valdés JA, Molina A. Dynamic transcriptional regulation of autocrine/paracrine igfbp1, 2, 3, 4, 5, and 6 in the skeletal muscle of the fine flounder during different nutritional statuses. J Endocrinol. 2012;214:95–108. doi: 10.1530/JOE-12-0057. [DOI] [PubMed] [Google Scholar]
- 75.Lee PD, Giudice LC, Conover CA, Powell DR. Insulin-like growth factor binding protein-1: recent findings and new directions. Proc Soc Exp Biol Med. 1997;216:319–57. doi: 10.3181/00379727-216-44182. [DOI] [PubMed] [Google Scholar]
- 76.Kajimura S, Aida K, Duan C. Insulin-like growth factor-binding protein-1 (IGFBP-1) mediates hypoxia-induced embryonic growth and developmental retardation. Proc Natl Acad Sci U S A. 2005;102:1240–45. doi: 10.1073/pnas.0407443102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Shimizu M, Beckman BR, Hara A, Dickhoff WW. Measurement of circulating salmon IGF binding protein-1: assay development, response to feeding ration and temperature, and relation to growth parameters. J Endocrinol. 2006;188:101–10. doi: 10.1677/joe.1.06475. [DOI] [PubMed] [Google Scholar]
- 78.Sheridan MA. Effects of thyroxine, cortisol, growth hormone, and prolactin on lipid metabolism of coho salmon, Oncorhynchus kisutch, during smoltification. Gen Comp Endocrinol. 1986;64:220–38. doi: 10.1016/0016-6480(86)90007-9. [DOI] [PubMed] [Google Scholar]
- 79.Andreassen PMR, Martinussen MB, Hvidsten NA, Stefansson SO. Feeding and prey-selection of wild Atlantic salmon post-smolts. J Fish Biol. 2001;58:1667–79. doi: 10.1111/j.1095-8649.2001.tb02321.x. [DOI] [Google Scholar]
- 80.Renkawitz MD, Sheehan TF. Feeding ecology of early marine phase Atlantic salmon Salmo salar post-smolts. J Fish Biol. 2011;79:356–73. doi: 10.1111/j.1095-8649.2011.03020.x. [DOI] [PubMed] [Google Scholar]
- 81.Breves JP, Phipps-Costin SK, Fujimoto CK, Einarsdottir IE, Regish AM, Björnsson BT, McCormick SD. Hepatic insulin-like growth-factor binding protein (igfbp) responses to food restriction in Atlantic salmon smolts. Gen Comp Endocrinol. 2016;233:79–87. doi: 10.1016/j.ygcen.2016.05.015. [DOI] [PubMed] [Google Scholar]
- 82.Bauchat JR, Busby WH, Garmong A, Swanson P, Moore J, Lin M, Duan C. Biochemical and functional analysis of a conserved IGF-binding protein isolated from rainbow trout (Oncorhynchus mykiss) hepatoma cells. J Endocrinol. 2001;170(3):619–28. doi: 10.1677/joe.0.1700619. [DOI] [PubMed] [Google Scholar]
- 83.Mommsen TP, Plisetskaya EM. Insulin in fishes and agnathans: history, structure, and metabolic regulation. Rev Aquat Sci. 1991;4:225–59. [Google Scholar]
- 84.Pierce AL, Shimizu M, Felli L, Swanson P, Dickhoff WW. Metabolic hormones regulate insulin-like growth factor binding protein-1 mRNA levels in primary cultured salmon hepatocytes; lack of inhibition by insulin. J Endocrinol. 2006;191:379–86. doi: 10.1677/joe.1.06986. [DOI] [PubMed] [Google Scholar]
- 85.McCormick SD, Regish AM, Christensen AK. Distinct freshwater and seawater isoforms of Na+/K+-ATPase in gill chloride cells of Atlantic salmon. J Exp Biol. 2009;212:3994–4001. doi: 10.1242/jeb.037275. [DOI] [PubMed] [Google Scholar]
- 86.Maures TJ, Duan C. Structure, developmental expression, and physiological regulation of zebrafish IGF binding protein-1. Endocrinology. 2002;143:2722–31. doi: 10.1210/endo.143.7.8905. [DOI] [PubMed] [Google Scholar]
- 87.Nakajima T, Shimura H, Yamazaki M, Fujioka Y, Ura K, Hara A, Shimizu M. Lack of hormonal stimulation prevents the landlocked Biwa salmon (Oncorhynchus masou subspecies) from adapting to seawater. Am J Physiol Regul Integr Comp Physiol. 2014;307(4):R414–25. doi: 10.1152/ajpregu.00474.2013. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.