

Abstract
The chromatin remodeling activity of the Swi/Snf complex is essential for the expression of several yeast genes. Previous studies have suggested that recruitment of Swi/Snf requires the action of transcriptional activators. However, reports in metazoans and in yeast have provided evidence of interactions between Swi/Snf and the RNA polymerase II holoenzyme/Mediator complex. Here we show that recruitment of Swi/Snf to the galactose-inducible gene GAL1 cannot be fully achieved without the integrity of the Mediator complex, TAFIIs, and RNA polymerase II. Moreover, artificial recruitment of Mediator is sufficient to tether both Swi/Snf and SAGA to the GAL1 UASG. We further demonstrate that Swi/Snf recruitment at GAL1 does not require acetylation of chromatin by Gcn5 nor the presence of SAGA. Based on these results, we conclude that interactions between the Gal4 activator and Swi/Snf are not sufficient to recruit the latter to the GAL1 UASG, since interactions with the Mediator, TAFIIs, and RNA polymerase II are also important.
Keywords: GAL1, Mediator, RNA polymerase II, Swi/Snf, TAFIIs
Introduction
Current views on mechanisms of gene transcription suggest that chromatin structure presents an important barrier to this fundamental cellular process. It is generally accepted that chromatin must be remodeled in order to allow transcription initiation to occur. Several mechanisms have been proposed to account for the process of chromatin remodeling. The first protein machinery to be exemplified as such, the Swi/Snf complex, hydrolyzes ATP as a source of energy to increase the accessibility of the transcriptional machinery to nucleosomal DNA (Vignali et al, 2000; Havas et al, 2001; Becker and Hörz, 2002). The Swi/Snf complex is composed of 11 tightly associated subunits, one of which is the DNA-dependent ATPase Swi2/Snf2 (Becker and Hörz, 2002). Swi/Snf has been shown to bind DNA and nucleosomes but without any known sequence specificity (Quinn et al, 1996; Côté et al, 1998). A second way designed to overcome the chromatin barrier is to covalently modify histone tails by machines such as the SAGA complex (Roth et al, 2001). SAGA possesses histone acetyltransferase (HAT) activity catalyzed by the Gcn5 subunit that preferentially targets histone H3 (Hampsey, 1997; Grant et al, 1998; Brown et al, 2000). The complex also performs other transcription-related activities, like facilitation of TATA-binding protein (TBP) binding (Dudley et al, 1999). The Ada1, Spt7, and Spt20 subunits are essential for SAGA integrity, whereas the other components are not (Grant et al, 1997; Sterner et al, 1999). A third way that has been suggested to account for chromatin remodeling is histone variants. In this case, incorporation of such variants into chromatin would either directly or indirectly make it more permissive to transcription. For example, in differentiated cells, the Drosophila histone variant H3.3 gradually replaces H3 in a replication-independent fashion upon exiting the cell cycle (Ahmad and Henikoff, 2002). This replacement forces the remodeling of nucleosomes in order to rapidly induce genes that were previously repressed (Ahmad and Henikoff, 2002). H2A.Z is another histone variant that could potentially create such specialized chromatin. H2A.Z is required for the proper expression of a subset of genes in yeast (Santisteban et al, 2000; Adam et al, 2001; Larochelle and Gaudreau, 2003), and has been shown to antagonize the spread of heterochromatin silencing (Meneghini et al, 2003).
How chromatin remodeling complexes are recruited to genes is a topic of intense research. Different mechanisms have been proposed to explain how Swi/Snf is targeted to specific genes. First, interactions with the general transcription machinery have been suggested to be involved in targeting Swi/Snf to promoters, as yeast and human Swi/Snf were shown to be associated with forms of the RNA polymerase II (polII) holoenzyme (Wilson et al, 1996; Cho et al, 1998; Neish et al, 1998), and artificial recruitment of the RNA polII holoenzyme, thus the Mediator complex, is sufficient to remodel chromatin at the PHO5 promoter in vivo (Gaudreau et al, 1997). Two recent reports propose a pathway in which the general transcription machinery mediates association of Swi/Snf with promoters (Sharma et al, 2003; Yoon et al, 2003). Second, it has been reported that ATP-dependent chromatin remodeling complexes and HAT complexes are targeted to promoters through interactions with gene-specific transcriptional activators. In the case of Swi/Snf, various studies have shown that it directly interacts with the acidic activation regions of Gcn4, Hap4, Gal4, Pho4, Swi5, and VP16 (Natarajan et al, 1999; Neely et al, 1999, 2002; Yudkovsky et al, 1999; Prochasson et al, 2003). Furthermore, an activating region is required for in vitro transcriptional stimulation of chromatin templates by Swi/Snf (Neely et al, 1999). Another study has demonstrated, by an in vitro assay, that the activating region of activators is required for the recruitment of Swi/Snf to a target gene (Yudkovsky et al, 1999). Two partially redundant activator-interaction domains have been identified in Swi/Snf that are important for targeting of the complex in vivo (Prochasson et al, 2003). Targeting has also been proposed to be achieved through a bromodomain that functions as an acetyl-lysine-binding domain. A bromodomain is found in the Swi2/Snf2 subunit of Swi/Snf (Haynes et al, 1992) and in nearly all nuclear HATs (Jeanmougin et al, 1997). Interestingly, several in vitro and in vivo studies suggest that histone H3 acetylation by Gcn5 is a prerequisite for recruitment and maintenance of the Swi/Snf complex at a promoter (Syntichaki et al, 2000; Hassan et al, 2001).
Various genetic studies in yeast suggest that the Swi/Snf chromatin remodeling complex functions together with the Gcn5 HAT to counteract the repressive effects of chromatin. Importantly, yeast genes regulated by Swi/Snf largely overlap those controlled by Gcn5 (Holstege et al, 1998). Moreover, combination of swi/snf and ada/gcn5 mutations has synthetic phenotypes suggesting that these two complexes can perform overlapping functions (Pollard and Peterson, 1997; Roberts and Winston, 1997). Recruitment of different types of chromatin remodeling complexes has been shown to be highly ordered at some promoters. For example, PHO8 induction first requires histone acetylation by Gcn5 and then ATP-dependent chromatin remodeling by Swi/Snf (Gregory et al, 1999). In addition, acetylation also precedes Swi/Snf action for the activation of the interferon-β (IFN-β) gene and activation by the retinoic acid receptor/retinoid X receptor (RAR/RXR) in human cells (Agalioti et al, 2000; Dilworth et al, 2000). On the other hand, in yeast, expression of the HO gene and expression of many genes in late mitosis require remodeling by Swi/Snf for the subsequent recruitment of SAGA (Cosma et al, 1999; Krebs et al, 1999).
The expression of the GAL1 gene in asynchronous Saccharomyces cerevisiae cells is generally independent of Swi/Snf (Burns and Peterson, 1997; Gaudreau et al, 1997; Krebs et al, 2000). However, certain conditions, such as a weakened Gal4 target promoter (Burns and Peterson, 1997; Gaudreau et al, 1997) or a more condensed chromatin structure during mitosis (Krebs et al, 2000), impose a requirement for Swi/Snf to fully induce the GAL1 gene. A conceptually similar result has been obtained by Santisteban et al (2000), which shows that either Swi/Snf or Gcn5 become essential at GAL1 and PHO5 when HTZ1, the H2A.Z-encoding gene, is deleted. These results raise the possibility that Swi/Snf and Gcn5, and perhaps other chromatin remodeling complexes, perform redundant functions at certain promoters that become essential only under conditions where chromatin is less permissive to transcription.
In order to investigate Swi/Snf targeting mechanisms, we performed chromatin immunoprecipitation (ChIP) at the GAL1 and GAL7 promoters. Our results show that Swi/Snf is recruited to the GAL1 and GAL7 UASG elements in asynchronous yeast cells even though the complex is not required for transcription of these genes. Moreover, Swi/Snf and SAGA are recruited with similar kinetics upon induction of the GAL genes and have a similar distribution over the GAL1-10 locus, being largely enriched at the GAL1 UASG region. Our results also show that neither histone acetylation by Gcn5 nor SAGA subunits are required for Swi/Snf recruitment at the GAL1 and GAL7 UASG elements. Furthermore, recruitment of the Swi/Snf and SAGA complexes can be achieved by artificially tethering the Mediator to GAL1 using a Gal4(DBD)-Gal11 fusion. Importantly, disruption of either the Mediator complex, TAFII subunits, or RNA polymerase II dramatically reduces Swi/Snf recruitment to the GAL1 UASG.
Results
Swi/Snf is recruited to the GAL1 and GAL7 UASG elements upon galactose induction
Generally independent of Swi/Snf action, GAL1 gene expression can become strongly Swi/Snf-dependent either by changing promoter strength or chromatin architecture (Burns and Peterson, 1997; Gaudreau et al, 1997; Krebs et al, 2000). This raises the question of how and under which conditions Swi/Snf is targeted to the GAL1 promoter. Either the Swi/Snf complex is only recruited when transcription of GAL1 undergoes conditions adverse to the achievement of full gene induction (e.g. during mitosis) or Swi/Snf is always recruited upon GAL1 induction independently of the requirement for its activity. In order to distinguish between these two possibilities, we measured the binding of Swi/Snf to the GAL1 promoter under conditions not requiring Swi/Snf. Two representative subunits of the complex, Swi1 and Snf5, were Myc tagged and used in ChIP experiments, followed by quantitative PCR analyses. Cells were grown in the presence of raffinose, and galactose was then added to induce GAL gene expression. Aliquots for primer extension and ChIP assays were taken before, and 15, 30, 60, and 120 min following addition of galactose. A schematic representation of the GAL7 and GAL1-10 loci and the regions analyzed by PCR in the ChIP assays are depicted in the upper panel of Figure 1A. The bottom part of Figure 1A shows a representative titration of the input and immunoprecipitated material used for PCR to demonstrate linearity of the reactions. PCR amplifications corresponding to the ARN1 promoter region were also carried out in most of our ChIP assays as an internal control to normalize the assays.
Figure 1.
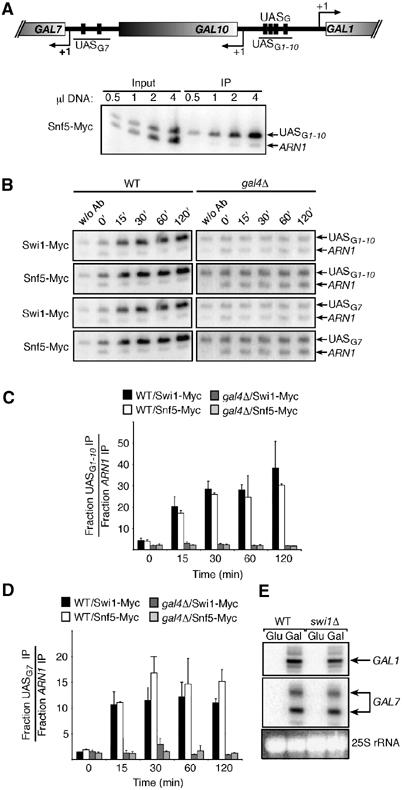
In vivo binding of Swi/Snf to the GAL1 and GAL7 UASG elements. (A) Top: Representation of the GAL7 and GAL1-10 loci. Transcriptional initiation sites (arrows with +1), Gal4 binding sites (UASG) (black crossbars), and open reading frames (open shaded rectangles) are represented. Regions amplified by PCR in the ChIP experiments are identified by a line. Bottom: PCR titration of input and immunoprecipitated material for Snf5-Myc at the GAL1 UASG. (B) ChIP analysis of the binding of Swi/Snf to the GAL1 and GAL7 promoters. Binding of Swi1-Myc and Snf5-Myc over time after galactose induction is shown for both the WT (KLY012 and KLY014) and gal4Δ strains (KLY015 and KLY016). (C, D) Binding of Swi/Snf to the GAL1 and GAL7 promoters, respectively. (E) Ability of swi1Δ cells to induce the GAL1 and GAL7 genes. GAL1 primer extension analyses from WT (CY258) and swi1Δ cells (CY340) grown either in the presence of glucose (Glu) or induced by galactose for 3 h (Gal).
Figure 1B first shows that Swi/Snf subunits, Swi1-Myc and Snf5-Myc, are efficiently recruited to the GAL1 UASG. However, recruitment of both subunits—and thus the Swi/Snf complex—is abolished when the ChIP assay is carried out in a strain lacking the Gal4 activator. Since Gal4 regulates expression of the GAL7 gene, association of Swi1-Myc and Snf5-Myc to the GAL7 UASG was therefore analyzed. Swi/Snf was also efficiently crosslinked to the GAL7 UASG in a wild-type (WT) strain but not in the gal4Δ strain (Figure 1B and D). Measurement of Swi/Snf binding at different time points following galactose induction demonstrates that the Swi/Snf complex is present and maintained at the GAL1 and GAL7 UASG elements throughout induction. In addition, Swi/Snf is rapidly recruited, reaching highest levels after 30 min of induction, and is then maintained at a constant level (Figure 1C and D). This is consistent with previous studies suggesting that the Swi/Snf complex is required for maintenance of activated transcription in vivo (Biggar and Crabtree, 1999; Sudarsanam et al, 1999). Figure 1E shows that, as previously reported (Burns and Peterson, 1997; Gaudreau et al, 1997), the GAL1 and GAL7 genes can be fully induced in a strain lacking SWI1. Taken together, these results indicate that the Swi/Snf complex is efficiently recruited to the GAL1 and GAL7 genes upon addition of galactose, and that recruitment is dependent on Gal4. Furthermore, Swi/Snf is recruited and maintained at the GAL1 promoter under activated conditions independently of its requirement for full gene activity.
Swi/Snf and SAGA are recruited to the same region at the GAL1-10 locus
Previous studies have demonstrated that the SAGA complex is recruited to the GAL1 UASG by the Gal4 activator (Bhaumik and Green, 2001; Larschan and Winston, 2001; Bhaumik et al, 2004). In order to compare the recruitment kinetics of SAGA with that of Swi/Snf, we measured binding of the Myc-tagged Gcn5 HAT subunit to both GAL1 and GAL7 UASG elements at different time points following induction. Gcn5-Myc is efficiently and rapidly recruited to both the GAL1 and GAL7 UASG elements upon galactose induction (Figure 2A and B). A quantification of these results is presented in Figure 2B. The results further show that the Gcn5 subunit of SAGA is recruited to the GAL1 and GAL7 UASG elements with binding kinetics similar to those of the Swi/Snf complex (compare Figures 1C, D and 2B).
Figure 2.
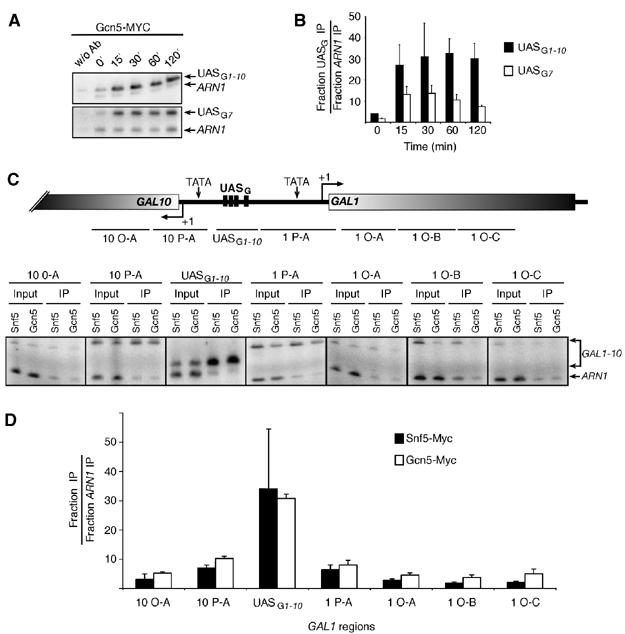
Distribution of Swi/Snf and SAGA at the GAL1-10 locus. (A) ChIP analysis of the binding of Gcn5-Myc (KLY031) to the GAL1 and GAL7 UASG during galactose induction. (B) Quantification of experiments as illustrated in (A). (C) Top: Representation of the GAL1-10 locus. Regions amplified in the GAL10 open reading frame (10 O-A), the GAL1-10 promoter region (10 P-A, UASG1-10, and 1 P-A) and in the GAL1 open reading frame (1 O-A to 1 O-C) are represented. Bottom: Distribution of Snf5-Myc and Gcn5-Myc over the GAL1-10 locus. ChIP analysis of the binding of Snf5-Myc and Gcn5-Myc to the GAL1-10 locus after 30 min addition of galactose. Data of input and immunoprecipitated material are shown for each region investigated. (D) Quantification of experiments as illustrated in (C).
We next investigated the distribution of the Swi/Snf and SAGA complexes over the GAL1-10 locus in order to compare their distribution patterns. Figure 2C first depicts a cartoon of the locus as well as the regions analyzed by PCR. A ChIP assay was carried out using yeast strains bearing Myc-tagged versions of Gcn5 or Snf5, and the GAL1-10 genes were induced by the addition of galactose. The bottom part of the figure shows a representative ChIP experiment, and Figure 2D shows a quantification of these experiments. The distribution of Swi/Snf is identical to that of SAGA, and both are enriched only over the UASG region.
Histone acetylation and SAGA are not required for Swi/Snf targeting and maintenance at GAL1 and GAL7
The experiments of Figure 2 demonstrate that Swi/Snf and SAGA are specifically recruited to the GAL1 and GAL7 UASG elements with similar kinetics. Since Swi2/Snf2 possesses a bromodomain, it is conceivable that histone acetylation by Gcn5 is involved in Swi/Snf recruitment and/or maintenance. To directly test this possibility, we measured the binding of Swi1-Myc at both UASG elements in a strain lacking the Gcn5 HAT. Figure 3A shows PCR analyses of the Swi1 ChIP at both the GAL1 and GAL7 UASG's in a gcn5Δ strain. Quantification of Swi1-Myc binding reveals that Swi/Snf recruitment in gcn5Δ cells is comparable to its recruitment in WT cells (Figure 3B). Similar results were obtained using Snf5-Myc (data not shown). These results suggest that an acetylation mark by Gcn5 is not essential to recruit and maintain the Swi/Snf complex at the GAL1 and GAL7 UASG elements.
Figure 3.
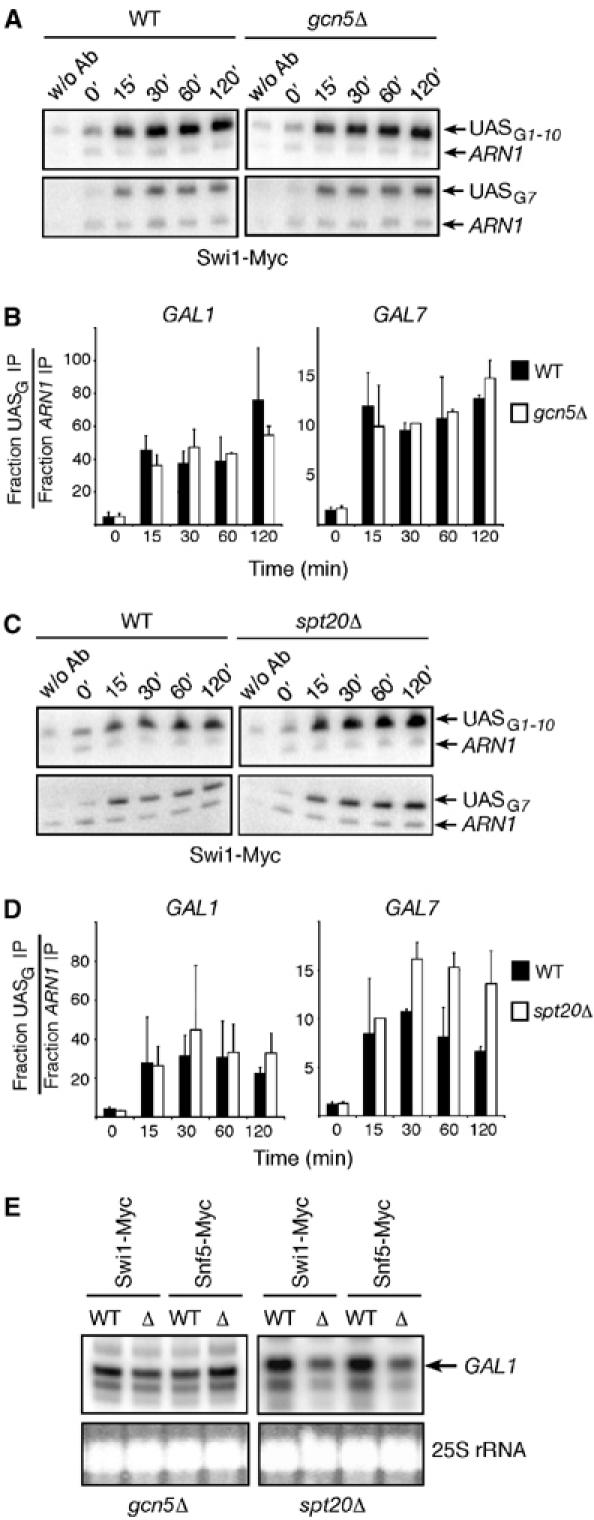
SAGA is not required for Swi/Snf recruitment at the GAL1 and GAL7 UASG elements. (A) ChIP analysis of Swi1-Myc binding to the GAL1 and GAL7 UASG in WT (KLY022) and gcn5Δ (KLY024) strains upon galactose induction. (B) Quantification of the experiment illustrated in (A). (C, D) Same experiment as in (A, B) with the exception that Swi1-Myc binding was analyzed in WT (KLY020) and spt20Δ (KLY028) cells. (E) Ability of Swi1-Myc- and Snf5-Myc-expressing yeast cells to induce the GAL1 gene. Primer extension assays were carried out with RNA purified from WT (KLY020, KLY021, KLY022, and KLY023), gcn5Δ (KLY024 and KLY025), and spt20Δ cells (KLY028 and KLY029) induced with galactose for 3 h.
We also tested whether Swi/Snf could be recruited to GAL genes in the absence of the entire SAGA complex, which is important for GAL1 gene induction (Marcus et al, 1994; Dudley et al, 1999; Bhaumik and Green, 2001). ChIP experiments were performed in spt20Δ cells (Figure 3C) since Spt20 is essential for the integrity of the SAGA complex (Grant et al, 1997; Sterner et al, 1999). Quantification of these results demonstrates that Swi1-Myc is efficiently recruited to the GAL1 and GAL7 UASG elements in the absence of SAGA (Figure 3D). Similar results were also obtained when testing the binding of Snf5-Myc (data not shown). We have also performed primer extension experiments to test the effect of either a GCN5 or SPT20 deletion on GAL1 induction in the Myc-tagged Swi1 or Snf5 yeast strains. As previously reported by others (Marcus et al, 1994; Dudley et al, 1999), GCN5 deletion did not affect GAL1 induction whereas SPT20 deletion significantly impaired GAL1 induction (Figure 3E). Taken together, these results show that neither histone acetylation by SAGA nor SAGA integrity is required for Swi/Snf recruitment at both the GAL1 and GAL7 UASG elements at least in the presence of the activator.
Swi/Snf and SAGA can be recruited to the GAL1 UASG by artificial recruitment of the Mediator complex
Since it was previously demonstrated that Swi/Snf copurifies with the Mediator (Wilson et al, 1996), and since recruitment of the Mediator is sufficient to remodel nucleosomes at the PHO5 promoter (Gaudreau et al, 1997), we wanted to test whether tethering the Mediator complex to the GAL1 UASG would be sufficient to recruit Swi/Snf in the absence of the Gal4 activator. In order to address this issue, we made use of a fusion protein bearing the Gal4 DNA-binding domain (DBD) fused to the C-terminal end of the Gal11 Mediator component. When appropriately targeted to a promoter region, this fusion protein directly recruits the transcriptional machinery to the promoter and thereby bypasses the requirement for a transcriptional activator to induce a target gene. Such bypass experiments have been extensively documented (Ptashne and Gann, 2002). Figure 4A shows that expression of the Gal4 DBD alone—in a strain deleted for GAL4 and bearing a Myc-tagged version of Snf5—does not induce GAL1 expression even when galactose is added. The figure further demonstrates that the Gal4-Gal11 fusion can efficiently induce GAL1 as previously reported (Farrell et al, 1996; Gaudreau et al, 1997). Moreover, the fusion protein is insensitive to galactose induction since it is not subjected to inhibition by Gal80 (Ma and Ptashne, 1987). PCR analyses of a ChIP assay using the same strains as in Figure 4A and quantification of the results are presented in Figure 4B and C. As expected, we observe no significant binding of Swi/Snf when the Gal4 DBD alone is expressed, whereas we observe efficient Swi/Snf recruitment when Gal4-Gal11 is expressed. Interestingly, we observe that Swi/Snf is constitutively recruited to the GAL1 UASG when using this fusion protein, a result consistent with that of Figure 4A. Similar results were also obtained when using other Mediator components fused to a DBD such as Gal4-Srb2, targeted to GAL1, or LexA-Srb2 and LexA-Srb6, targeted to a synthetic lacZ reporter gene construct (data not shown) (Farrell et al, 1996; Gaudreau et al, 1999).
Figure 4.
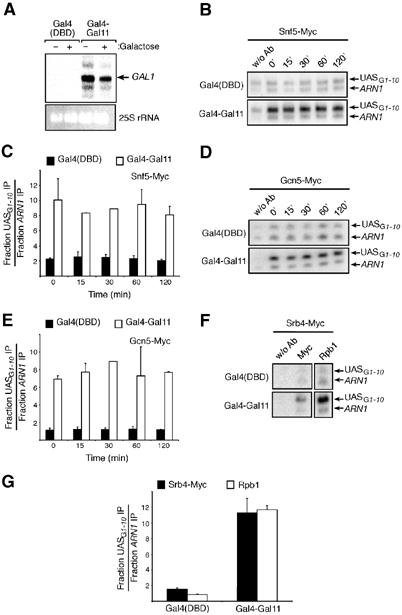
Swi/Snf and SAGA are recruited to the GAL1 UASG by artificial recruitment of the Mediator. (A) Artificial recruitment of the Mediator bypasses the requirement for a transcriptional activator. A strain deleted for GAL4 and bearing a Myc-tagged version of Snf5 was transformed by a plasmid expressing the Gal4 DBD alone (KLY019) or fused to the C-terminal end of Gal11 (KLY018). Primer extension analyses were carried out with purified RNA from both strains. Yeast cells were grown in the presence of raffinose (−) and then galactose (+) was added in order to induce GAL1 gene expression. (B) ChIP analysis of the binding of Swi/Snf to the GAL1 UASG in yeast cells expressing Gal4-Gal11. Binding of Snf5-Myc in the strains used in (A). (C) Quantification of experiments as illustrated in (B) is shown. (D, E) Same experiment as in (B, D) with the exception that the binding of Gcn5-Myc (KLY044 and KLY043) was assessed. (F) ChIP analysis of the binding of Srb4-Myc and Rpb1 to the GAL1 UASG in yeast cells expressing a Gal4-Gal11 fusion (KLY091 and KLY092). (G) Quantification of experiments as illustrated in (F).
We next tested if artificial recruitment of the Mediator would also be able to recruit SAGA to the GAL1 UASG. The assay presented above was repeated with Myc-tagged Gcn5 instead of Myc-tagged Snf5 and, surprisingly, similar results were obtained. Expression of the Gal4-DBD alone did not allow binding of Gcn5-Myc, while significant recruitment of Gcn5-Myc was achieved when expressing the Gal4-Gal11 fusion (Figure 4D). Quantification of the results shows that Gcn5-Myc is also efficiently recruited to the GAL1 UASG in a constitutive fashion (Figure 4E). Thus, even though a direct interaction between the Gal4 activator and the Tra1 subunit of SAGA has been demonstrated in vivo (Bhaumik et al, 2004), direct or indirect interactions with the Mediator are sufficient to target SAGA, as well as Swi/Snf, to the GAL1 UASG.
We next wanted to confirm that Gal4-Gal11 would indeed be able to recruit the Mediator as well as RNA polII at GAL1. To test this, we used a Myc-tagged Srb4 strain expressing either Gal4 (DBD) or Gal4-Gal11. Figure 4F thus shows a ChIP assay using anti-Myc antibodies to immunoprecipitate Srb4-containing complexes or anti-Rpb1 antibodies to immunoprecipitate RNA polII-containing complexes. The results of the figure show that both Srb4 and Rpb1 can be efficiently recruited to the GAL1 UASG by the Gal4-Gal11 fusion but not by Gal4(DBD). Quantification of these results is shown in Figure 4G.
The Mediator is required for Swi/Snf recruitment at the GAL1 UASG
Efficient recruitment of Swi/Snf to GAL1 by the Gal4-Gal11 fusion protein suggested that integrity of the Mediator might be important for the process. In order to address this issue, we performed ChIP experiments with a yeast strain bearing a thermosensitive (ts) allele of Srb4 (Thompson and Young, 1995), as well as Myc-tagged Swi1. When shifting these cells to a nonpermissive temperature, Srb4 is quickly inactivated and the integrity of the entire Mediator complex is compromised (Koh et al, 1998; Kang et al, 2001). Figure 5A shows that Swi1-Myc is efficiently recruited to GAL1 upon galactose induction when cells are grown at permissive temperature (28°C), whereas Swi1-Myc recruitment is significantly reduced when the same cells are shifted to nonpermissive temperature (37°C) (Figure 5B). We next wanted to verify if shifting the srb4-138 strain to 37°C would indeed compromise recruitment of the Mediator to GAL1. We thus Myc-tagged Gal11 in the srb4-138 strain background and carried out a ChIP experiment. Figure 5C shows that Gal11-Myc is efficiently recruited at the GAL1 UASG upon galactose induction when cells are grown at 28°C. However, when cells are grown at 37°C, Gal11-Myc recruitment at GAL1 is severely affected, a result demonstrating that the integrity of the Mediator complex is severely compromised, although not completely, under our experimental conditions. Quantification of these results is shown in Figure 5D. It is interesting to note that the extent of Swi/Snf recruitment at GAL1 when cells are shifted to 37°C is comparable to Mediator recruitment. This thus underscores the importance of Mediator integrity for the recruitment of Swi/Snf to GAL1. Taken together, the results presented here and in the previous figure suggest that the Swi/Snf complex must interact, directly or indirectly, with the Mediator in order to be recruited to the GAL1 gene. A similar result was also obtained at the GAL7 gene thereby extending our results to another GAL-inducible gene (data not shown).
Figure 5.
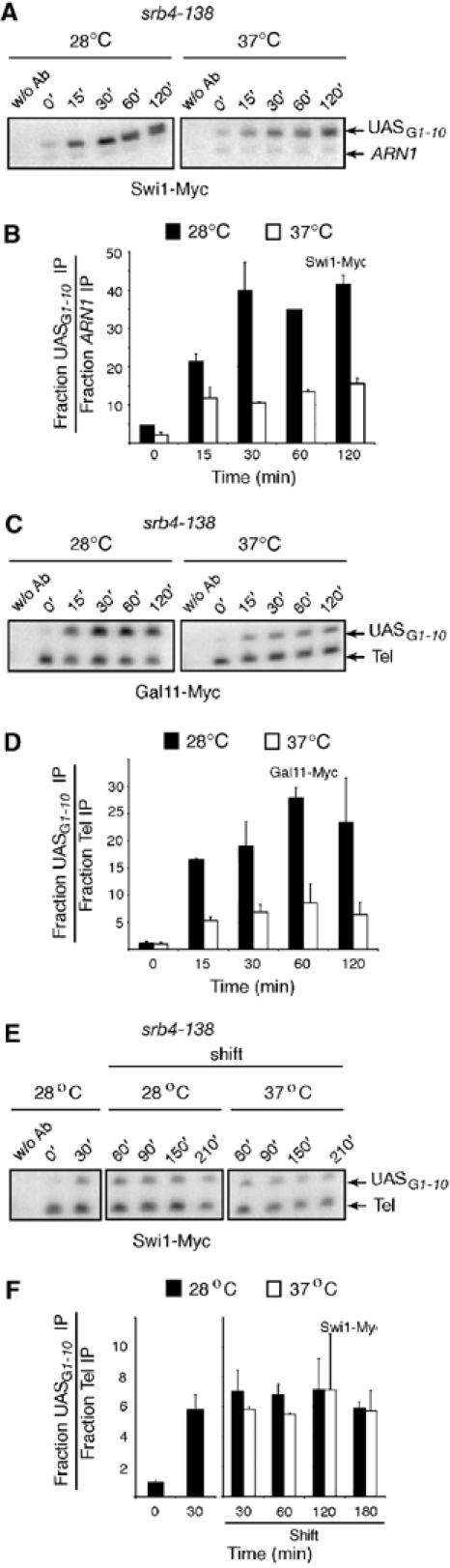
The Mediator is required for Swi/Snf recruitment to the GAL1 UASG. (A) ChIP analysis of the binding of Swi1-Myc to the GAL1 UASG in the absence of the Mediator. Temperature-sensitive srb4-138 mutant (KLY042) containing Swi1-Myc was grown at 28°C and either shifted to 37°C for 45 min to inactivate srb4 (37°C) or kept at the permissive temperature (28°C). Galactose was added to media as in previous experiments GAL gene expression. (B) Quantification of experiments as shown in (A). (C, D) Similar experiment as in (A, B) except that binding of Gal11-Myc was analyzed (KLY090). PCRs contain chromosome II telomere primers used here as an internal control to normalize signals for each lane. (E) Persistence of the Swi/Snf complex in the absence of the Mediator. KLY042 cells were grown at the permissive temperature and galactose was added to the media for 30 min to induced GAL gene expression. Cells were then shifted to inducing media at permissive (28°C) or nonpermissive temperature (37°C). Aliquots were taken 30, 60, 120, and 180 min after a shift to the nonpermissive temperature. PCRs contain chromosome II telomere primers as an internal control. (F) Quantification of experiments as illustrated in (E).
We wanted to determine whether Mediator was also necessary for Swi/Snf maintenance at the GAL1 UASG after it has been fully recruited to this gene. To investigate this, we again made use of the Swi1-Myc srb4-138 strain as described in Figure 5A and B. Cells were grown at 28°C, and Swi1-Myc binding was monitored after 30 min of GAL1 induction. In this strain, we had observed that maximal Swi/Snf binding at GAL1 could be achieved within 30 min (see Figure 5B). We next shifted the same cell culture to 37°C and monitored Swi1-Myc binding after 30, 60, 120, and 180 min after inactivation of the Mediator. The results show that Swi1-Myc binding is not significantly affected under these conditions, suggesting that the Mediator is not required to maintain the Swi/Snf complex at GAL1 after it has been recruited.
TAFIIs and RNA polII are required for Swi/Snf recruitment at the GAL1 UASG
A previous report had demonstrated that Swi/Snf recruitment to a stress-inducible gene, RNR3, required TAFIIs, the general transcription machinery, as well as the Mediator (Sharma et al, 2003). We imagined that GAL1, much like RNR3, could also require TAFIIs and the general transcription machinery in order to recruit Swi/Snf. However, in the case of RNR3, no activator seems to be required for Swi/Snf recruitment since this gene is predominantly regulated by a repression mechanism (Sharma et al, 2003). It is thus conceivable that the requirement of RNR3 for TAFIIs and the general transcription machinery would be unique and not applicable for genes that are induced predominantly by the action of an activator. Thus, in order to test if recruitment of Swi/Snf at GAL1 also required TAFIIs and the general transcription machinery, we made use of three yeast strains, each bearing a thermosensitive allele of either RPB1 (rpb1-1), which encodes the large subunit of RNA polII, TAF1 (taf1-2), and TAF12 (taf12-9) (see Sharma et al, 2003). All three yeast strains bear a Myc-tagged version of Swi2/Snf2. ChIP experiments were carried out with all three yeast strains either at the permissive temperature (28°C) or at the nonpermissive temperature (37°C). Figure 6A, C, and E shows ChIP data for the rpb1-1, taf1-2, and taf12-9 strains, respectively, whereas Figure 6B, D, and F shows quantifications of these data. The results show that in all three strains, as expected, Swi2-Myc recruitment to the GAL1 UASG is efficient when cells are grown at the permissive temperature, whereas this binding is almost abolished when cells are grown at the nonpermissive temperature. Taken together, these results show that TAFIIs and RNA polII are also essential for Swi/Snf targeting to the GAL1 UASG.
Figure 6.
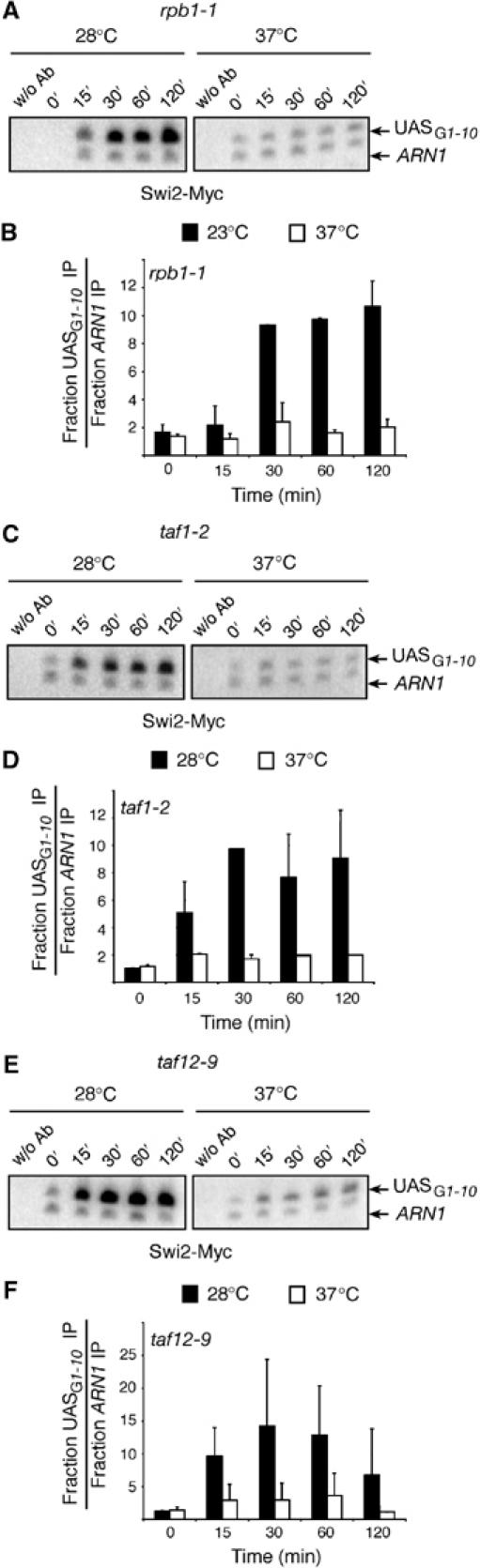
RNA polII and TAFIIs are required for Swi/Snf recruitment to the GAL1 UASG. (A) ChIP analysis of the binding of Swi2-Myc (YJR598) to the GAL1 UASG in an rpb1-1 yeast strain. The experiment was performed as described in Figure 5A. (B) Quantification of experiments as illustrated in (A). (C, D) Similar experiments as in (A, B) except that binding of Swi2-Myc was assessed in a taf1-2 mutant (YJR595). (E, F) Analysis of the binding of Swi2-Myc in a taf12-9 mutant (YJR592).
Discussion
In this study, we have shown that the Swi/Snf chromatin remodeling complex is recruited to both the GAL1 and GAL7 UASG elements in asynchronous cells, a context that does not require the function of Swi/Snf for maximal gene activity. Furthermore, recruitment of Swi/Snf does not require action of SAGA. Importantly, targeting of both complexes to the GAL1 UASG can be achieved by artificial recruitment of the Mediator complex, a condition that bypasses the requirement for a transcriptional activator. In agreement with these findings, Swi/Snf recruitment to the GAL1 UASG in the absence of either the Mediator, TAFIIs, or RNA polII is strongly affected.
Since Swi/Snf is present at as few as 200 copies per yeast cell (Côté et al, 1994; Cairns et al, 1996), it may be assumed that it is recruited to target genes only if there is a strict requirement for the activity of the complex. Interestingly, we have shown by ChIP assays that Swi/Snf is recruited to the GAL1 and GAL7 UASG elements regardless of the level of requirement for that chromatin remodeling complex. Why then is Swi/Snf recruited to GAL1? It is conceivable that Swi/Snf facilitates chromatin remodeling in asynchronous yeast cells as well as during mitosis. However, in unsynchronized cells, chromatin remodeling by Swi/Snf is dispensable since Swi/Snf and SAGA can perform redundant functions (Biggar and Crabtree, 1999; Sudarsanam et al, 1999), but when gene expression conditions become more subject to chromatin-mediated repression, such as when yeast cells enter mitosis, Swi/Snf might have a rate determinant role (Krebs et al, 1999).
Different orders of recruitment schemes for the Swi/Snf and SAGA complexes have been described for different promoters (Cosma et al, 1999; Gregory et al, 1999; Krebs et al, 1999; Agalioti et al, 2000; Dilworth et al, 2000). Our results show that both complexes are recruited with similar kinetics upon galactose induction, suggesting that they are simultaneously recruited to the GAL1 UASG. Moreover, the presence of Swi/Snf or SAGA is not a prerequisite for the recruitment of the other complex since the amount of Swi/Snf immunoprecipitated in gcn5Δ and spt20Δ mutant cells is similar to a WT strain. In addition, the Swi/Snf independence of GAL1 in unsynchronized cells makes it unlikely to be important for the subsequent recruitment of the required SAGA complex (Marcus et al, 1994). Another argument in favor of the suggestion that Swi/Snf and SAGA recruitment are not interdependent is that histone H3 acetylation levels at the GAL1 promoter are maintained when Swi/Snf is absent (Krebs et al, 2000). Based on these results, we propose that no special order of recruitment is necessary in promoter targeting for Swi/Snf and SAGA at GAL1 and GAL7. However, there could be an order of action for these cellular regulators in cases where the chromatin architecture of the promoter is altered. This idea is supported by the fact that Swi/Snf is a prerequisite for acetylation by Gcn5 in late mitosis (Krebs et al, 2000).
Several models have been proposed to explain how chromatin remodeling complexes are recruited to their target genes, and among these, targeting by a transcriptional activator has been widely reported (Natarajan et al, 1999; Neely et al, 1999, 2002; Yudkovsky et al, 1999). In a recent study, Bhaumik et al (2004) identified a direct interaction between the Gal4 activator and the Tra1 subunit of SAGA in vivo. Our results are also consistent with the fact that the Gal4 transcriptional activator plays a determinant role in the recruitment of both the Swi/Snf and SAGA complexes to the GAL1 and GAL7 loci upon galactose induction. However, targeting of Swi/Snf and SAGA to GAL1 is unlikely to be the consequence of their sole interaction with the acidic activator Gal4 since artificial recruitment of the Mediator can also recruit both complexes to GAL1. These results suggest that both Swi/Snf and SAGA can be actively recruited to those GAL genes simply by direct or indirect interactions with a Gal11-bearing Mediator. Moreover and importantly, Swi/Snf recruitment to GAL1 is strongly affected in the absence of the Mediator, but the latter is not necessary to maintain Swi/Snf at GAL1 once it has been fully recruited. It is conceivable that once recruited, Swi/Snf is able to interact with either other components of the transcriptional machinery or with chromatin, thereby making interaction with the Mediator dispensable. In agreement with this last possibility, we find that TAFIIs and RNA polII are also required to recruit Swi/Snf to GAL1. While we anticipate functional interactions between Swi/Snf and the Mediator, it is likely that another important role of the Mediator in Swi/Snf recruitment would be to assemble a functional transcription initiation complex—as its recruitment to GAL1 precedes that of the general transcription machinery (Bryant and Ptashne, 2003)—including polII and TFIID, thus allowing Swi/Snf to interact with the transcriptional machinery. This possibility remains to be tested.
The fact that SAGA is not required for the recruitment of Swi/Snf to GAL1 could imply that it is neither required for the recruitment of the Mediator, a prediction that is in agreement with the study of Bryant and Ptashne (2003). However, this result was recently challenged by Bhaumik et al (2004). We have thus carried out ChIP experiments aimed at measuring Mediator recruitment in the absence of SAGA under our experimental conditions and find that while there is a notable delay in Mediator recruitment, SAGA is not strictly essential for this process (see Supplementary Figure 1). Interestingly, it was also shown that the Mediator was not necessary for the recruitment of SAGA at the GAL1 UASG (Bhaumik et al, 2004). Thus, it is likely that the interaction between Swi/Snf and the Mediator is more significant for its action than the interaction that exists between SAGA and the Mediator. In fact, Swi/Snf but not SAGA has previously been shown to copurify with RNA polII holoenzymes (Wilson et al, 1996). We surmise that even though the action of Gal4 alone may be sufficient to recruit Swi/Snf and SAGA to the GAL1 and GAL7 genes, the Mediator/transcriptional machinery can also independently fulfill this function. In contrast, the study of Yudkovsky et al (1999) has suggested that the Swi/Snf complex can be efficiently recruited to a chimeric HIS4-based promoter template from a yeast extract preparation bearing a deletion in the Mediator component SRB2. In a similar in vitro experiment, deletion of SRB2 was shown to prevent recruitment of the Mediator and RNA polII to a promoter (Ranish et al, 1999). It is conceivable that at some promoters, the Mediator might not elicit interactions with Swi/Snf, or alternatively the discrepancy existing between our results and theirs might be due to the use of different experimental systems. The involvement of other types of interactions, such as those elicited by the bromodomain of Swi2/Snf2 and acetylated chromatin, remains to be assessed at the GAL genes. However, we consider the contribution of the Swi2/Snf2 bromodomain to be nonsignificant at GAL1 since it was shown that Swi/Snf is still efficiently recruited in gcn5Δ cells to GAL1, and since Gcn5 is the primary H3 HAT at GAL1 (Krebs et al, 2000). Furthermore, H4 was shown to be deacetylated upon galactose induction at GAL1 (Deckert and Struhl, 2001). Thus, we imagine that activator and Mediator/transcriptional machinery-dependent interactions with certain chromatin remodeling/coactivator complexes could perform redundant recruitment functions that would make up a system designed to quickly and efficiently induce certain genes. This mechanism could be particularly useful for genes that require quick and robust induction, such as the GAL genes, in order for an organism to adapt to a rapidly changing environment.
Materials and methods
Yeast strains and genetic methods
A list of yeast strains used in this study is provided in Table I. The Myc-tagged strains used in our studies were generated by homologous recombination using a PCR-directed method with a 9Myc-URA3 cassette (Cosma et al, 1999).
Table 1.
Yeast strains used in this study
| Strain | Genotype | Source |
|---|---|---|
| BY4741 | MATa, his3Δ1, leu2Δ0, met1Δ50, ura3Δ0 | RESGEN |
| CY258 | MATα swi1Δ::LEU2, his3-Δ200, ura3-Δ99, leu2-Δ1, ade2-101, lys2-801 | C Peterson (Univ. Mass. Medical School) |
| CY340 | MATα, his3-Δ200, ura3-Δ99, leu2-Δ1, lys2− | C Peterson (Univ. Mass. Medical School) |
| FY250 | MATα, his3-Δ200, ura3-52, leu2-Δ1, trp1Δ63 | F Winston (Harvard Medical School) |
| JPY9 | MATα, his3-Δ200, ura3-52, leu2-Δ1, trp1Δ63, gal4Δ11 | Barberis et al (1995) |
| KLY012 | FY250; SWI1-9myc::URA3 | This study |
| KLY014 | FY250; SNF5-9myc::URA3 | This study |
| KLY015 | JPY9; SWI1-9myc::URA3 | This study |
| KLY016 | JPY9; SNF5-9myc::URA3 | This study |
| KLY018 | KLY016; pSO74::LEU2 | This study |
| KLY019 | KLY016; pLRG234::LEU2 | This study |
| KLY020 | BY4741; SWI1-9myc::URA3 | This study |
| KLY021 | BY4741; SNF5-9myc::URA3 | This study |
| KLY022 | PSY316; SWI1-9myc::URA3 | This study |
| KLY023 | PSY316; SNF5-9myc::URA3 | This study |
| KLY024 | PSY316 gcn5Δ; SWI1-9myc::URA3 | This study |
| KLY025 | PSY316 gcn5Δ; SNF5-9myc::URA3 | This study |
| KLY028 | 7390; SWI1-9myc::URA3 | This study |
| KLY029 | 7390; SNF5-9myc::URA3 | This study |
| KLY031 | FY250; GCN5-9myc::URA3 | This study |
| KLY033 | JPY9; GCN5-9myc::URA3 | This study |
| KLY042 | Z628; SWI1-9myc::URA3 | This study |
| KLY043 | KLY033; pSO74::LEU2 | This study |
| KLY044 | KLY033; pLRG234::LEU2 | This study |
| KLY053 | BY4741; GAL11-9myc::URA3 | This study |
| KLY054 | 7390; GAL11-9myc::URA3 | This study |
| KLY055 | BY4741; SRB4-9myc::URA3 | This study |
| KLY056 | 7390; SRB4-9myc::URA3 | This study |
| KLY079 | JPY9; SRB4-9myc::URA3, pSO74::LEU2 | This study |
| KLY080 | JPY9; SRB4-9myc::URA3, pLRG234::LEU2 | This study |
| KLY090 | Z628; GAL11-9myc::URA3 | This study |
| KLY091 | JPY9; SRB4-9myc::URA3, pSO74::LEU2 | This study |
| KLY092 | JPY9; SRB4-9myc::URA3, pLRG234::LEU2 | This study |
| MAY424 | W303; htz1Δ::hisg | Adam et al (2001) |
| PSY316 | MATα, ade2-101, his3-Δ200, leu2-3, 112, lys2, ura3-53 | Candau et al (1996) |
| PSY316 gcn5Δ | PSY316; gcn5Δ::hisg | Candau et al (1996) |
| JR592 | PH499; taf12-9::HIS3, SNF2-13-myc::KanMX | Sharma et al (2003) |
| JR595 | PH499; taf1-2::TRP1, SNF2-13-myc::KanMX | Sharma et al (2003) |
| JR598 | Z676; rpb1-1, SNF2-13myc::KanMX | Sharma et al (2003) |
| W303α | MATα, ura3-1, leu2-3, 112, ade2-1, his3-11, 15, trp1-1, can1-100 | Primig et al (2000) |
| Z628 | MATa, his3-Δ200, leu2-3, ura3-52, srb4Δ2::HIS3, pR42882::LEU2 | Thompson and Young (1995) |
| 7390 | BY4741; spt20Δ::kanMX4 | RESGEN |
Gene induction and primer extension assays
GAL genes expression was induced by growing cells to OD600 of 1.0 in yeast nitrogen base supplemented with the required amino acids and 2% raffinose. Galactose was then added to a final concentration of 5% for up to 60 min. In order to monitor GAL gene expression in an swi1Δ strain, cells were grown in yeast extract–peptone–2% dextrose to OD600 of 1.0, and then the cells were washed three times with sterile water and resuspended in yeast extract–peptone–2% galactose. Cells were further incubated for 3 h. For primer extension assays, 20 μg of RNA was used, and primer extension analyses were carried out essentially as previously described (Ma and Ptashne, 1987). The sequences for the oligonucleotide primers used were as follows: GAL1, CTCCTTGACGTTAAAGTATAGAGG; GAL7, GGATGGTAACGTCTATGGGAATGGC. For RNA loading controls, 10 μg of total RNA was loaded onto agarose–formaldehyde gels.
Chromatin immunoprecipitations
ChIP experiments were performed as described previously (Adam et al, 2001; Larochelle and Gaudreau, 2003) with some modifications. Briefly, cells were grown to an OD600 of 0.8 in yeast nitrogen base supplemented with the required amino acids and 2% raffinose. Galactose was then added to a final concentration of 5% to induce GAL genes, and 50 ml samples were collected 0, 15, 30, 60, and 120 min after induction. For temperature-shift experiments, cells were either grown in yeast nitrogen base supplemented with the required amino acids and 2% raffinose or in yeast extract–peptone–2% raffinose at 28°C until OD600 reached 0.8. Cells were then pelleted and resuspended in media prewarmed at 37°C. Following a 45-min preincubation period, galactose was added to induce GAL gene expression. To perform the Swi/Snf persistence experiment, cells were grown until OD600 reached 0.8. GAL gene expression was induced by the addition of galactose (5% final concentration). After 30 min induction, cells were pelleted and resuspended in inducing media at permissive (28°C) or nonpermissive temperature (37°C). Samples (50 ml) were collected 0 and 30 min after induction before the temperature shift, and 30, 60, 120, and 180 min after shifting cells to nonpermissive temperature (37°C). All immunoprecipitations were performed using 500 μl of whole-cell extract incubated with either anti-Myc (9E10, Santa Cruz) or anti-CTD (8WG16, BAbCo) antibodies coupled to magnetic beads (Dynal). In all, 1 or 3 μl of DNA was used for the PCRs. PCRs contained either ARN1 or chromosome II telomere primers (TEL) used as an internal control to normalize signals for each lane. All the ChIP data were quantified by phosphorImaging (Phosphor Imager, Molecular Dynamics). Reactions without antibodies were also carried out for all experiments. The sequence of the oligonucleotides used for all of the experiments is available upon request. Quantification of results represents mean values and standard deviations from at least two independent experiments.
Supplementary Material
Supplementary Figure 1
Acknowledgments
We thank Joëlle Brodeur, Alain Lavigueur, Benoît Leblanc, Annie Moisan, Sébastien Rodrigue, and Viktor Steimle for discussions and critical comments on the manuscript. We also thank François Robert for the 9Myc-URA3 tagging cassette, and especially Joseph Reese for providing valuable yeast strains. This work was supported by funds of the Canadian Institutes of Health Research to LG. LG holds a Canada Research Chair on mechanisms of gene transcription. KL is a recipient of a fellowship from the National Science and Engineering Research Council of Canada.
References
- Adam M, Robert F, Larochelle M, Gaudreau L (2001) H2A.Z is required for global chromatin integrity and for recruitment of RNA polymerase II under specific conditions. Mol Cell Biol 21: 6270–6279 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Agalioti T, Lomvardas S, Parekh B, Yie J, Maniatis T, Thanos D (2000) Ordered recruitment of chromatin modifying and general transcription factors to the IFN-beta promoter. Cell 103: 667–678 [DOI] [PubMed] [Google Scholar]
- Ahmad K, Henikoff S (2002) The histone variant H3.3 marks active chromatin by replication-independent nucleosome assembly. Mol Cell 9: 1191–1200 [DOI] [PubMed] [Google Scholar]
- Barberis A, Pearlberg J, Simkovich N, Farrell S, Reinagel P, Bamdad C, Sigal G, Ptashne M (1995) Contact with a component of the polymerase II holoenzyme suffices for gene activation. Cell 81: 359–368 [DOI] [PubMed] [Google Scholar]
- Becker PB, Hörz W (2002) ATP-dependent nucleosome remodeling. Annu Rev Biochem 71: 247–273 [DOI] [PubMed] [Google Scholar]
- Bhaumik SR, Green MR (2001) SAGA is an essential in vivo target of the yeast acidic activator Gal4p. Genes Dev 15: 1935–1945 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bhaumik SR, Raha T, Aiello DP, Green MR (2004) In vivo target of a transcriptional activator revealed by fluorescence resonance energy transfer. Genes Dev 18: 333–343 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Biggar SR, Crabtree GR (1999) Continuous and widespread roles for the Swi–Snf complex in transcription. EMBO J 18: 2254–2264 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown CE, Lechner T, Howe L, Workman JL (2000) The many HATs of transcription coactivators. Trends Biochem Sci 25: 15–19 [DOI] [PubMed] [Google Scholar]
- Bryant GO, Ptashne M (2003) Independent recruitment in vivo by Gal4 of two complexes required for transcription. Mol Cell 11: 1301–1309 [DOI] [PubMed] [Google Scholar]
- Burns LG, Peterson CL (1997) The yeast SWI–SNF complex facilitates binding of a transcriptional activator to nucleosomal sites in vivo. Mol Cell Biol 17: 4811–4819 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cairns BR, Lorch Y, Li Y, Zhang M, Lacomis L, Erdjument-Bromage H, Tempst P, Du J, Laurent B, Kornberg RD (1996) RSC, an essential, abundant chromatin-remodeling complex. Cell 87: 1249–1260 [DOI] [PubMed] [Google Scholar]
- Candau R, Moore PA, Wang L, Barlev N, Ying CY, Rosen CA, Berger SL (1996) Identification of human proteins functionally conserved with the yeast putative adaptors ADA2 and GCN5. Mol Cell Biol 16: 593–602 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cho H, Orphanides G, Sun X, Yang XJ, Ogryzko V, Lees E, Nakatani Y, Reinberg D (1998) A human RNA polymerase II complex containing factors that modify chromatin structure. Mol Cell Biol 18: 5355–5363 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cosma MP, Tanaka T, Nasmyth K (1999) Ordered recruitment of transcription and chromatin remodeling factors to a cell cycle- and developmentally regulated promoter. Cell 97: 299–311 [DOI] [PubMed] [Google Scholar]
- Côté J, Peterson CL, Workman JL (1998) Perturbation of nucleosome core structure by the SWI/SNF complex persists after its detachment, enhancing subsequent transcription factor binding. Proc Natl Acad Sci USA 95: 4947–4952 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Côt J, Quinn J, Workman JL, Peterson CL (1994) Stimulation of GAL4 derivative binding to nucleosomal DNA by the yeast SWI/SNF complex. Science 265: 53–60 [DOI] [PubMed] [Google Scholar]
- Deckert J, Struhl K (2001) Histone acetylation at promoters is differentially affected by specific activators and repressors. Mol Cell Biol 21: 2726–2735 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dilworth FJ, Fromental-Ramain C, Yamamoto K, Chambon P (2000) ATP-driven chromatin remodeling activity and histone acetyltransferases act sequentially during transactivation by RAR/RXR in vitro. Mol Cell 6: 1049–1058 [DOI] [PubMed] [Google Scholar]
- Dudley AM, Rougeulle C, Winston F (1999) The Spt components of SAGA facilitate TBP binding to a promoter at a post-activator-binding step in vivo. Genes Dev 13: 2940–2945 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Farrell S, Simkovich N, Wu Y, Barberis A, Ptashne M (1996) Gene activation by recruitment of the RNA polymerase II holoenzyme. Genes Dev 10: 2359–2367 [DOI] [PubMed] [Google Scholar]
- Gaudreau L, Keaveney M, Nevado J, Zaman Z, Bryant GO, Struhl K, Ptashne M (1999) Transcriptional activation by artificial recruitment in yeast is influenced by promoter architecture and downstream sequences. Proc Natl Acad Sci USA 96: 2668–2673 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gaudreau L, Schmid A, Blaschke D, Ptashne M, Hörz W (1997) RNA polymerase II holoenzyme recruitment is sufficient to remodel chromatin at the yeast PHO5 promoter. Cell 89: 55–62 [DOI] [PubMed] [Google Scholar]
- Grant PA, Duggan L, Côté J, Roberts SM, Brownell JE, Candau R, Ohba R, Owen-Hughes T, Allis CD, Winston F, Berger SL, Workman JL (1997) Yeast Gcn5 functions in two multisubunit complexes to acetylate nucleosomal histones: characterization of an Ada complex and the SAGA (Spt/Ada) complex. Genes Dev 11: 1640–1650 [DOI] [PubMed] [Google Scholar]
- Grant PA, Sterner DE, Duggan LJ, Workman JL, Berger SL (1998) The SAGA unfolds: convergence of transcription regulators in chromatin-modifying complexes. Trends Cell Biol 8: 193–197 [DOI] [PubMed] [Google Scholar]
- Gregory PD, Schmid A, Zavari M, Munsterkotter M, Hörz W (1999) Chromatin remodelling at the PHO8 promoter requires SWI–SNF and SAGA at a step subsequent to activator binding. EMBO J 18: 6407–6414 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hampsey M (1997) A SAGA of histone acetylation and gene expression. Trends Genet 13: 427–429 [DOI] [PubMed] [Google Scholar]
- Hassan AH, Neely KE, Workman JL (2001) Histone acetyltransferase complexes stabilize swi/snf binding to promoter nucleosomes. Cell 104: 817–827 [DOI] [PubMed] [Google Scholar]
- Havas K, Whitehouse I, Owen-Hughes T (2001) ATP-dependent chromatin remodeling activities. Cell Mol Life Sci 58: 673–682 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haynes SR, Dollard C, Winston F, Beck S, Trowsdale J, Dawid IB (1992) The bromodomain: a conserved sequence found in human, Drosophila and yeast proteins. Nucleic Acids Res 20: 2603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holstege FC, Jennings EG, Wyrick JJ, Lee TI, Hengartner CJ, Green MR, Golub TR, Lander ES, Young RA (1998) Dissecting the regulatory circuitry of a eukaryotic genome. Cell 95: 717–728 [DOI] [PubMed] [Google Scholar]
- Jeanmougin F, Wurtz JM, Le Douarin B, Chambon P, Losson R (1997) The bromodomain revisited. Trends Biochem Sci 22: 151–153 [DOI] [PubMed] [Google Scholar]
- Kang JS, Kim SH, Hwang MS, Han SJ, Lee YC, Kim YJ (2001) The structural and functional organization of the yeast mediator complex. J Biol Chem 276: 42003–42010 [DOI] [PubMed] [Google Scholar]
- Koh SS, Ansari AZ, Ptashne M, Young RA (1998) An activator target in the RNA polymerase II holoenzyme. Mol Cell 1: 895–904 [DOI] [PubMed] [Google Scholar]
- Krebs JE, Fry CJ, Samuels ML, Peterson CL (2000) Global role for chromatin remodeling enzymes in mitotic gene expression. Cell 102: 587–598 [DOI] [PubMed] [Google Scholar]
- Krebs JE, Kuo MH, Allis CD, Peterson CL (1999) Cell cycle-regulated histone acetylation required for expression of the yeast HO gene. Genes Dev 13: 1412–1421 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Larochelle M, Gaudreau L (2003) H2A.Z has a function reminiscent of an activator required for preferential binding to intergenic DNA. EMBO J 22: 4512–4522 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Larschan E, Winston F (2001) The S. cerevisiae SAGA complex functions in vivo as a coactivator for transcriptional activation by Gal4. Genes Dev 15: 1946–1956 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma J, Ptashne M (1987) The carboxy-terminal 30 amino acids of GAL4 are recognized by GAL80. Cell 50: 137–142 [DOI] [PubMed] [Google Scholar]
- Marcus GA, Silverman N, Berger SL, Horiuchi J, Guarente L (1994) Functional similarity and physical association between GCN5 and ADA2: putative transcriptional adaptors. EMBO J 13: 4807–4815 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meneghini MD, Wu M, Madhani HD (2003) Conserved histone variant H2A.Z protects euchromatin from the ectopic spread of silent heterochromatin. Cell 112: 725–736 [DOI] [PubMed] [Google Scholar]
- Natarajan K, Jackson BM, Zhou H, Winston F, Hinnebusch AG (1999) Transcriptional activation by Gcn4p involves independent interactions with the SWI/SNF complex and the SRB/mediator. Mol Cell 4: 657–664 [DOI] [PubMed] [Google Scholar]
- Neely KE, Hassan AH, Brown CE, Howe L, Workman JL (2002) Transcription activator interactions with multiple SWI/SNF subunits. Mol Cell Biol 22: 1615–1625 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neely KE, Hassan AH, Wallberg AE, Steger DJ, Cairns BR, Wright AP, Workman JL (1999) Activation domain-mediated targeting of the SWI/SNF complex to promoters stimulates transcription from nucleosome arrays. Mol Cell 4: 649–655 [DOI] [PubMed] [Google Scholar]
- Neish AS, Anderson SF, Schlegel BP, Wei W, Parvin JD (1998) Factors associated with the mammalian RNA polymerase II holoenzyme. Nucleic Acids Res 26: 847–853 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pollard KJ, Peterson CL (1997) Role for ADA/GCN5 products in antagonizing chromatin-mediated transcriptional repression. Mol Cell Biol 17: 6212–6222 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Primig M, Williams RM, Winzeler EA, Tevzadze GG, Conway AR, Hwang SY, Davis RW, Esposito RE (2000) The core meiotic transcriptome in budding yeasts. Nat Genet 26: 415–423 [DOI] [PubMed] [Google Scholar]
- Prochasson P, Neely KE, Hassan AH, Li B, Workman JL (2003) Targeting activity is required for SWI/SNF function in vivo and is accomplished through two partially redundant activator-interaction domains. Mol Cell 12: 983–990 [DOI] [PubMed] [Google Scholar]
- Ptashne M, Gann A. (2002) Genes & Signals. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press [Google Scholar]
- Quinn J, Fyrberg AM, Ganster RW, Schmidt MC, Peterson CL (1996) DNA-binding properties of the yeast SWI/SNF complex. Nature 379: 844–847 [DOI] [PubMed] [Google Scholar]
- Ranish JA, Yudkovsky N, Hahn S (1999) Intermediates in formation and activity of the RNA polymerase II preinitiation complex: holoenzyme recruitment and a postrecruitment role for the TATA box and TFIIB. Genes Dev 13: 49–63 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roberts SM, Winston F (1997) Essential functional interactions of SAGA, a Saccharomyces cerevisiae complex of Spt, Ada, and Gcn5 proteins, with the Snf/Swi and Srb/mediator complexes. Genetics 147: 451–465 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roth SY, Denu JM, Allis CD (2001) Histone acetyltransferases. Annu Rev Biochem 70: 81–120 [DOI] [PubMed] [Google Scholar]
- Santisteban MS, Kalashnikova T, Smith MM (2000) Histone H2A.Z regulates transcription and is partially redundant with nucleosome remodeling complexes. Cell 103: 411–422 [DOI] [PubMed] [Google Scholar]
- Sharma VM, Li B, Reese JC (2003) SWI/SNF-dependent chromatin remodeling of RNR3 requires TAF(II)s and the general transcription machinery. Genes Dev 17: 502–515 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sterner DE, Grant PA, Roberts SM, Duggan LJ, Belotserkovskaya R, Pacella LA, Winston F, Workman JL, Berger SL (1999) Functional organization of the yeast SAGA complex: distinct components involved in structural integrity, nucleosome acetylation, and TATA-binding protein interaction. Mol Cell Biol 19: 86–98 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sudarsanam P, Cao Y, Wu L, Laurent BC, Winston F (1999) The nucleosome remodeling complex, Snf/Swi, is required for the maintenance of transcription in vivo and is partially redundant with the histone acetyltransferase, Gcn5. EMBO J 18: 3101–3106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Syntichaki P, Topalidou I, Thireos G (2000) The Gcn5 bromodomain co-ordinates nucleosome remodelling. Nature 404: 414–417 [DOI] [PubMed] [Google Scholar]
- Thompson CM, Young RA (1995) General requirement for RNA polymerase II holoenzymes in vivo. Proc Natl Acad Sci USA 92: 4587–4590 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vignali M, Hassan AH, Neely KE, Workman JL (2000) ATP-dependent chromatin-remodeling complexes. Mol Cell Biol 20: 1899–1910 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilson CJ, Chao DM, Imbalzano AN, Schnitzler GR, Kingston RE, Young RA (1996) RNA polymerase II holoenzyme contains SWI/SNF regulators involved in chromatin remodeling. Cell 84: 235–244 [DOI] [PubMed] [Google Scholar]
- Yoon S, Qiu H, Swanson MJ, Hinnebusch AG (2003) Recruitment of SWI/SNF by Gcn4p does not require Snf2p or Gcn5p but depends strongly on SWI/SNF integrity, SRB mediator, and SAGA. Mol Cell Biol 23: 8829–8845 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yudkovsky N, Logie C, Hahn S, Peterson CL (1999) Recruitment of the SWI/SNF chromatin remodeling complex by transcriptional activators. Genes Dev 13: 2369–2374 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary Figure 1