

Abstract
Ecdysteroids are widely used as inducers for gene-switch systems based on insect ecdysteroid receptors and genes of interest placed under the control of ecdysteroid-response elements. We review here these systems, which are currently mainly used in vitro with cultured cells in order to analyse the role of a wide array of genes, but which are expected to represent the basis for future gene therapy strategies. Such developments raise several questions, which are addressed in detail.
First, the metabolic fate of ecdysteroids in mammals, including humans, is only poorly known, and the rapid catabolism of ecdysteroids may impede their use as in vivo inducers.
A second set of questions arose in fact much earlier with the pioneering “heterophylic” studies of Burdette in the early sixties on the pharmacological effects of ecdysteroids on mammals. These and subsequent studies showed a wide range of effects, most of them being beneficial for the organism (e.g. hypoglycaemic, hypocholesterolaemic, anabolic). These effects are reviewed and critically analysed, and some hypotheses are proposed to explain the putative mechanisms involved.
All of these pharmacological effects have led to the development of a wide array of ecdysteroid-containing preparations, which are primarily used for their anabolic and/or “adaptogenic” properties on humans (or horses or dogs). In the same way, increasing numbers of patents have been deposited concerning various beneficial effects of ecdysteroids in many medical or cosmetic domains, which make ecdysteroids very attractive candidates for several practical uses.
It may be questioned whether all these pharmacological actions are compatible with the development of ecdysteroid-inducible gene switches for gene therapy, and also if ecdysteroids should be classified among doping substances.
| Abbreviation: | |
|---|---|
| 20E | 20-hydroxyecdysone |
| 2d20E | 2-deoxy-20-hydroxyecdysone |
| 2dE | 2-deoxyecdysone |
| BAH | bisacylhydrazine |
| BmEcR | Bombyx mori EcR |
| CfEcR | Choristoneura fumiferana EcR |
| CfUSP | Choristoneura fumiferana USP |
| CHO | Chinese hamster ovary |
| CMV | cytomegalovirus |
| DBD | DNA-binding domain |
| DmEcR | Drosophila melanogaster EcR |
| AbbE | ecdysone |
| EcR | ecdysteroid receptor |
| EcRE | ecdysteroid response element |
| EHT | effective half-time |
| ERE | oestrogen response element |
| GR | glucocorticoid receptor |
| GRE | glucocorticoid response element |
| HEK | human embryonic kidney |
| HvEcR | Heliothis virescens EcR |
| LBD | ligand binding domain |
| murA | muristerone A |
| PKA | protein kinase A |
| polB | polypodine B |
| ponA | ponasterone A |
| PPAR | peroxisome proliferator-activated receptor |
| RAR | retinoic acid receptor |
| RXR | retinoid X receptor |
| TR | thyroid receptor |
| USP | ultraspiracle |
| VDR | vitamin D receptor |
| VEGF | vascular endothelial growth factor |
Introduction
Ecdysteroids (zooecdysteroids) are steroid hormones that control moulting and reproduction of arthropods. Whether they fulfil hormonal functions in other invertebrate groups is still a matter of debate. In 1966, the discovery of the same molecules (phytoecdysteroids) in several plant species made them easily available in large amounts, and this allowed pharmacological studies to be initiated on mammals. Such studies were at first undertaken in the hope of developing safer and more specific insecticides, and it was quickly shown that these molecules were not toxic to mammals. On the other hand, they displayed a wide array of rather beneficial pharmacological effects (e.g. against diabetes or asthenia), thus providing a plausible explanation for the properties of several plant species widely used in traditional medicine. Although they have been detected in ca. 6% of plant species analysed so far (Dinan, 2001), phytoecdysteroids are not so frequent in plant species used as human food (with the noticeable exception of spinach; Bathory et al., 1982; Grebenok et al., 1991). More than 300 different ecdysteroids have been isolated from animal and plant sources (all their structures can be found in the Ecdybase, http://ecdybase.org).
Ecdysteroids are structurally quite different from mammalian steroids, and they are not expected to bind to vertebrate steroid receptors. Soon after the isolation and cloning of Drosophila melanogaster ecdysteroid receptor proteins, it appeared very attractive to use them for designing inducible gene systems in mammalian cells. Such a system has been commercially developed by Invitrogen® and the potential use of ecdysteroid receptors for gene therapy is being investigated. The different ecdysteroid-based gene-switch systems will be reviewed in the first part of this article.
The in vivo use of ecdysteroids as inducers taken orally raises questions about their uptake, metabolism and half-life in mammals including humans, a topic which has not been extensively investigated up to now (Sláma and Lafont, 1995), and this question will be addressed in the second part of this review.
The development of ecdysteroid-regulated gene switches seems, however, to have neglected much of the previous pharmacological studies which showed the interference of ecdysteroids with many physiological processes in mammals and humans. All these effects will be summarised in the third part, paying special attention to the protocols used and the significance/limitations of the results obtained. In the light of recent data, we will present in the fourth section some working hypotheses, which could explain how ecdysteroids might act on mammalian cells.
The reported effects (mainly the anabolic effects) led initially to a (doping ?) use for high-performance sportsmen in the Eastern Bloc Countries, but nowadays a large number of ecdysteroid-based preparations are freely available on the market. Most of them are proposed as legal and non-toxic muscle-promoting substances for bodybuilders, but an extensive search on the web has led to more surprising findings (e.g. recommended use for golfers or for domestic animals). So, whether ecdysteroids should be considered as doping substances and whether their use should be controlled will be finally discussed.
Ecdysone-inducible gene expression systems
Basic requirements
Spatial and temporal control of heterologous gene expression is an area of considerable and growing interest with relevance to basic and applied biological and medical research, including gene therapy and functional genomics. However, these heterologous regulatory systems should interfere minimally with the complex endogenous regulatory networks. Ideally, heterologous modification of gene expression in host cells should give rapid, robust, precise and reversible induction (or suppression) of the target gene(s). The necessary criteria are thus (Saez et al., 1997; Bohl and Heard, 1998; DeMayo and Tsai, 2001; Fussenegger, 2001; Graham, 2002):
Specificity: the system should not interfere with endogenous regulatory networks and should be activated exclusively by exogenous nontoxic compounds.
Inducibility: the system should possess a low baseline expression and a high induction ratio.
Bioavailability of the inducer: control should be effected by a drug that readily penetrates tissue.
Reversibility: the elicitor should possess high pharmacokinetic turnover to enable reversal and permit repeated cycles of induction.
Low immunogenicity: the components of the system should not elicit immune responses in the host.
Flexibility: it should be possible to modify the system to take account of different tissue applications and to optimise the system for each of these.
Dose-dependence: the extent of the response should be dependent on the dose of elicitor applied.
Ecdysteroid receptors in arthropods
Ecdysteroid receptors are members of the nuclear receptor superfamily (Laudet, 1997), which are characterised by a domain structure. The N-terminal A/B-domain is highly variable and is associated with transcriptional activation. The C-domain is highly conserved and is involved in binding the receptor complex to specific response elements in the DNA. The D-domain is variable and represents a hinge region between the DNA-binding domain and the ligand-binding domain (E-domain). The E-domain is not only responsible for ligand binding, but also has been implicated in receptor dimerisation and interactions with other transcriptional activators. There may also be a C-terminal F-domain, which, if present, is highly variable between even closely related nuclear receptors (Kumar and Thompson, 1999). Nuclear receptors regulate gene expression as dimers, either as homodimers or as heterodimers with another member of the nuclear receptor superfamily. One of the most promiscuous heterodimeric partners for vertebrate nuclear receptors is RXR, of which the equivalent in insects is Ultraspiracle (USP; Oro et al., 1990). In the case of ecdysteroid receptors, only the EcR:USP (or EcR:RXR) (Yao et al., 1993) complex is able to bind the ecdysteroid ligand with high affinity and the presence of ecdysteroid promotes complex formation. The ecdysteroid binds to the EcR protein. No definitive ligand for USP has been identified, but it has been suggested that juvenile hormones (or methyl farnesoate in Crustacea) may bind to this receptor component and modify the transactivation capacity of the complex (Jones and Jones, 2000).
The most extensively studied ecdysteroid receptor system in arthropods is that of Drosophila melanogaster, where three isoforms (A, B1 and B2) of EcR occur (Koelle et al., 1991; Talbot et al., 1993). These isoforms arise through alternative promoter usage and differential splicing, resulting in different A/B-domains, but they all possess common DNA- and ligand-binding domains. The EcR isoforms show tissue- and stage-specificity. Although there is only one form of USP in D. melanogaster, two or more isoforms have been found in other arthropods. USP isoforms also show tissue- and stage-specificity (Kapitskaya et al., 1996).
EcR and USP gene homologues have now been characterised from a variety of arthropod species: Aedes aegypti (Cho et al., 1995; Kapitskaya et al., 1996), Amblyomma americana (Palmer et al., 1999), Bombyx mori (Swevers et al., 1996), Ceratitis capitata (Verras et al., 1999), Chironomus tentans (Imhof et al., 1993; Vögtli et al., 1999), Choristoneura fumiferana (Kothapalli et al., 1995; Perera et al., 1999), Heliothis virescens (Martinez et al., 1999c), Locusta migratoria (Saleh et al., 1998; Hayward et al., 1999), Lucilia cuprina (Hannan and Hill, 1997; 2001), Manduca sexta (Fujiwara et al., 1995; Jindra et al., 1997), Ostrinia nubilalis (Albertsen et al., 2000), Sarcophaga crassipalpis (Rinehart et al., 2001), Tenebrio molitor (Mouillet et al., 1997; Nicolai et al., 2000), Uca pugilator (Durica et al., 2002).
The biochemical characterisation of ecdysteroid receptor complexes lags well behind that of vertebrate steroid hormone receptors and has been in a period of quiescence for the past decade, as emphasis has been placed on the characterisation and expression of the genes. The generally accepted ligand for ecdysteroid receptors in arthropods is 20E, but this does not preclude the other ecdysteroids being significant at particular stages of development or in certain tissues (Wang et al., 2000). In fact, ecdysteroid receptor complexes recognise a wide range of ecdysteroid structural analogues and sophisticated structure-activity and molecular modelling studies are now beginning to be performed (Dinan et al., 1999a; Wurtz et al., 2000; Ravi et al., 2001; Kumar et al., 2002). Owing to the importance of ecdysteroid receptors in the regulation of arthropod development, they are seen as an appropriate target for the development of new pest control agents. In the context of this review, the identification of bisacylhydrazines as non-steroidal ecdysteroid agonists (Wing, 1988; Dhadialla et al., 1998) is particularly worth mentioning as, in addition to several of these molecules being commercialised as insecticides, other analogues appear appropriate as gene switching elicitors. Antagonists for ecdysteroid receptors are also being identified (Dinan et al., 1999b).
Ecdysteroid-responsive expression systems
Mammalian systems:
Ecdysteroids are apparently not endogenously generated components of mammalian systems. However, they are normal components of the diets of many animals. The low mammalian toxicity of these compounds (Sláma and Lafont, 1995), together with the specificity of the ecdysteroid receptor complex (EcR and USP proteins), indicate that a successful gene-switching system might be developed from this system (Fig. 2). With regard to plant systems (see below), there are a significant number (ca. 6% of higher terrestrial species) of plants which accumulate phytoecdysteroids (Dinan, 2001). This may restrict the use of steroidal and non-steroidal ecdysteroid analogues as elicitors in plant systems.
Figure 2.
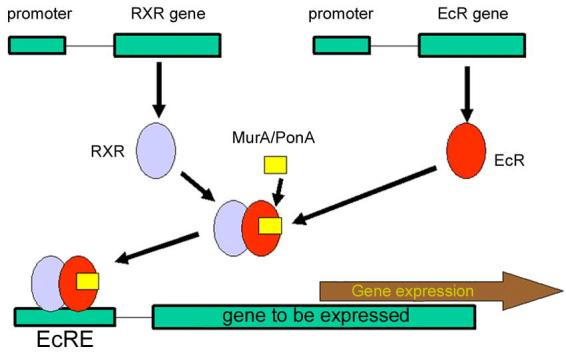
General scheme for ecdysteroid-based gene switches.
Initial reports appeared in the early 1990s (Christopherson et al., 1992; Thomas et al., 1993; Yao et al., 1992; 1993). Christopherson et al. (1992) transfected a human embryonic kidney cell line (HEK293) with DmEcR and a reporter gene and assessed the ability of various ecdysteroids and vertebrate steroids (all at 1 µM) to induce reporter activity; E, 20E and polB and the vertebrate steroids were inactive, while ponA and murA were active. The domain structure of nuclear receptors allows the domains to operate autonomously (however, this should not be taken to mean that the domains operate exactly the same under all circumstances). This permitted the ligand binding domain of EcR to be fused with the DNA-binding and A/B-regions of the GR (GGEc) and the demonstration of the induction of a GRE-containing reporter gene by murA and with the same ecdysteroid specificity as for EcR in the same mammalian cells. MurA could also induce a reporter gene via a chimeric receptor recognising a consensus oestrogen response element (ERE). They also demonstrated that replacement of a portion of the GR N-terminal activation domain in GGEc with the activation domain of the Herpes simplex viral protein (VP16) resulted in 5-fold greater activity (Christopherson et al., 1992).
Yao et al. (1992) showed that USP could substitute for RXR as a heterodimeric partner for RAR, TR, VDR and PPAR and showed that, for many mammalian cells types, cotransfection of USP with EcR was necessary to make the cells ecdysteroid-responsive, demonstrating that USP is an essential part of the ecdysteroid receptor complex.
Thomas et al. (1993) found that certain mammalian cell lines (e.g. HeLa) could support ecdysteroid-responsive transactivation while others (e.g. CV-1) could not. They demonstrated that the factor responsible for this was RXR. RXR could not be replaced by RAR-α, TR-α or COUP-TF, but USP was an effective partner for EcR. MurA was effective at enhancing the DNA-binding activity (as assessed by gel-shift assays) of EcR:RXR, but not EcR:USP. Interestingly, the ligand of RXR 9-cis-retinoic acid, also enhanced the DNA-binding activity of EcR:RXR complexes.
The system has been further developed (No et al., 1996). The final form of this development (VgEcR) was more specific and gave a lower basal activity than tetracycline-responsive systems. The starting point for the developments of No et al. (1996) was the observation that mammalian cells cotransfected with EcR and USP only produce a 3-fold induction on treatment with murA (1 µM). To improve the induction ratio they carried out a number of modifications. Replacement of USP by RXR gave 34-fold induction. Creation of a fusion protein consisting of an N-terminal truncation of EcR attached to the VP16 activation domain (generating VpEcR) gave 212-fold induction. Inclusion of binding sites for the transcription factor Sp1 into the reporter vector between minimal promoter and the EcREs enhanced the induction by a further 5-fold.
Since the ecdysteroid response element might be weakly activated by endogenous farnesoid X receptors FXR, the EcRE (to give 2 different half-sites with a 1-nucleotide spacer, AGGTCA-AGAACA, generating E/GRE) and DBD of EcR (by mutating 3 amino acids in the P-box of the DNA-binding domain, generating VgEcR) have been modified to ensure that the response element will only bind the modified EcR. The transcription-regulatory potential of EcR has been enhanced by replacing the endogenous activation domain by the Herpes simplex virus VP16 activation domain (DeMayo and Tsai, 2001). The final system gives a 1200-fold induction with 1 µM murA, without interference from glucocorticoid or farnesoid.
No et al. (1996) also generated transgenic mice harbouring an ecdysteroid-inducible promoter or a T-cell-specific expression construct of VpEcR and RXR. Crossing of these two strains of mice gave double transgenic offspring, which were induced to generate the reporter gene transcript specifically in the thymus by injection of murA (10 mg/mouse). Mice expressing VpEcR and RXR were healthy, fertile and apparently normal.
Yang et al. (1995) produced a Chinese hamster ovary (CHO) cell line stably transfected with EcR isoform B1 and showed that the cells produce functional receptor of the correct Mr (105 kDa), which is recognised by specific antibodies, binds to EcRE in gel-shift assays and mediates reporter gene expression in a ligand-dependent manner (ponA; 4 – 100 µM). The authors suggest that CHO cells produce high levels of RXR, which can heterodimerise with EcR to generate functional receptor complexes.
A parallel system using the Bombyx mori receptor (Swevers et al., 1995; Swevers et al., 1996) has been developed (Suhr et al., 1998), who found that BmEcR, in conjunction with murA (1 µM) or RH5992 (tebufenozide; 1 µM) could effect high level transactivation of a reporter gene in the absence of exogenous heterodimeric partner in mammalian cells (HEK293 cells and African green monkey CV-1 cells). BmEcR is much shorter (616 amino acids; Swevers et al., 1995) and has less than 42% overall amino acid identity with the B1 isoform of DmEcR (878 amino acids; Suhr et al., 1998). It has been recognised that RXR (which is present to at least some extent in most, if not all, mammalian cells) is a reluctant heterodimerisation partner for DmEcR (Thomas et al., 1993; Yao et al., 1993), but it appears to be less reluctant for BmEcR. By creating chimaeric EcRs consisting of Dm and Bm domains in various combinations, Suhr et al. (1998) could demonstrate that the regions responsible for high affinity, ligand-dependent heterodimerisation in BmEcR were present in the hinge region (D) and the ligand-binding (E) domain. Only the D-region appears to be involved in heterodimerisation of BmEcR with USP.
Hoppe et al. (2000) created a hybrid Drosophila/Bombyx ecdysteroid receptor (DB-EcR), which is independent of recombinant RXR, and demonstrated its efficacy in vitro and in vivo.
The commercially available Invitrogen system (http://www.invitrogen.com: Fig. 3) has been used to regulate the expression of a wide range of transfected genes in mammalian cells (Sawicki et al., 1998; Chen et al., 2000; Lüers et al., 2000; Niikura et al., 2000; Rampazzo et al., 2000; Abeysinghe et al., 2001; Baba et al., 2001; Cole et al., 2001; Gill et al., 2001; Hennigan and Stambrook, 2001; Iwata et al., 2001; Jana et al., 2001; Kondo et al., 2001; Patrick et al., 2001; Schmidt and Fan, 2001; Shi et al., 2001; Sparacio et al., 2001; Stauffer et al., 2001; Stolarov et al., 2001; Wang et al., 2001; Xu et al., 2001; Yam et al., 2001; Yarovoi and Pederson, 2001; Zhu et al., 2001; Chen et al., 2002; Coulthard et al., 2002; Davis et al., 2002; Hashimoto et al., 2002; Kuate et al., 2002; Kudo et al., 2002; Meents et al., 2002; Mellon et al., 2002; Odero-Marah et al., 2002; Plows et al., 2002; Vickers and Sharrocks, 2002; Wang et al., 2002; Wolter et al., 2002; Xiao et al., 2003; Xu and Mellgren, 2002; Zhang et al., 2002) and in a tissue-specific manner in mice (No et al., 1996; Albanese et al., 2000;). The literature on the application of ecdysteroid-regulated transgenic systems is currently growing exponentially.
Figure 3.
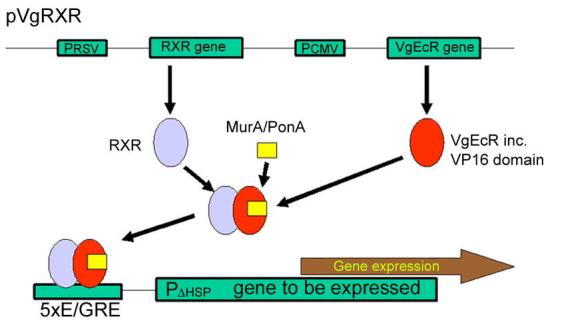
The Invitrogen system for mammalian cells; the elicitor is muristerone A (murA) or ponasterone A (ponA). See text for further details (www.invitrogen.com).
Wyborski et al. (2001) developed a bicistronic expression vector from which VgEcR and RXR can be co-expressed. They used the general cytomegalovirus (CMV) promoter, but this can be replaced with a cell-type specific promoter.
Albanese et al. (2000) developed a system for mammary gland-specific expression of an ecdysteroid-regulatable gene in mice and have examined the pharmacokinetics of injected ponA in the animals. Serum clearance was rapid (activity half-life = 48 min).
Karns et al. (2001) have developed an alternative to the Invitrogen system. The basic system (Figure 4) consists of the (i) plasmid pGAL4-EcR, encoding a fusion protein of the yeast GAL4 DNA-binding domain and the ligand-binding domain of the ecdysteroid receptor from Choristoneura fumiferana, (ii) plasmid pVP16-mRXR, encoding a fusion protein of the Herpes simplex transcriptional transactivator VP16 and mouse RXR protein, and which, in the presence of ecdysteroid-type ligands heterodimerises with GAL4-EcR, (iii) an indicator and selection plasmid, either pGAL4-EGFP-SV40-neo (consisting of 5 copies of the GAL4 response element, followed by the minimal promoter region of the major late promoter from adenovirus, the coding region of enhanced green fluorescent protein [EGFP], the SV40 promoter and the neomycin resistance locus) or pGAL4-SEAP (for stable transformation, containing 5 × the GAL4 response element and the coding region of the SEAP protein as reporter gene) iv) the BAH GS-E (1 – 15 µM). This system forms the basis of RHeoGene's RHeoswitch Technology. As RHeoGene have access to a large number of BAH analogues (RHeoChem Ligands) and EcR genes from a wide variety of insect species (RHeocept Receptors), they are able to identify BAH analogues which are specific for particular EcR LBDs and, thus, have the possibility to regulate multiple genes in a coordinated manner, and this is being developed under the title of RHeoPlex Systems (www.rheogene.com). Karns et al. (2001) also considered the suitablitiy of GS-E as an in vivo inducer in mice and obtained maximal induction of reporter protein in 6–12 hrs and return to basal expression levels by 12–24 hrs.
Figure 4.
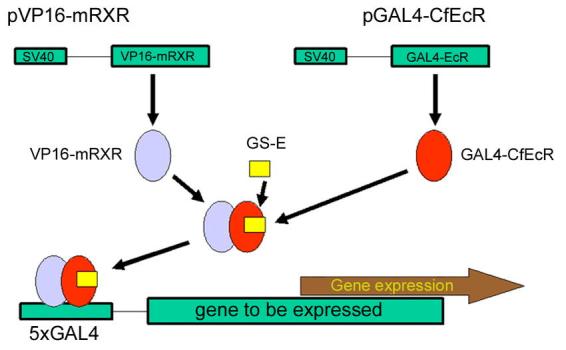
The RHeoGene system for mammalian cells; the elicitor depicted is the bisacylhydrazine GS-E (1-[3-methoxy-2-ethylbenzoyl]-2-[3,5-dimethylbenzoyl]-2-tert-butylhydrazine). See text for further details (Karns et al., 2001).
Plant systems:
Most of the published research in this area has been conducted by the industrial research labs at Zeneca Agrochemicals (now Syngenta) and has been based on the ecdysteroid receptor protein from Heliothis virescens (HvEcR), which was cloned and characterised (Martinez et al., 1999c). This protein has most similarity to EcRs from other lepidopteran species and is closely related to the B1 isoform from D. melanogaster (DmEcRB1). Transfection of mammalian HEK293 cells (RXR-containing) with HvEcR and a reporter gene resulted in induction of the reporter gene by murA (50% response at ca. 5 µM), but not by 20E (Martinez et al., 1999a). For the development of the plant system (Fig. 5), a chimeric receptor consisting of the hinge and ligand-binding domains of HvEcR was fused to the transactivation domain of the Herpes simplex VP16 protein and the DNA-binding domain of the glucocorticoid receptor and transfected into tobacco protoplasts. The use of the GR DNA binding domain circumvents the need to incorporate USP/RXR into the system, since glucocorticoid receptors bind to their response elements as homodimers. The second component of the gene regulatory system consisted of 6 copies of the glucocorticoid response element fused to the minimal 35S cauliflower mosaic virus (35SCaMV) promoter (conferring expression in all tissues and throughout development) and a β-glucuronidase gene. Although induction was observed with murA (100 µM), its steroidal nature precludes its use under field conditions. Consequently, the non-steroidal BAH RH5992 (1 – 10 µM) was used as an elicitor. In addition to being ecdysteroid agonists, these compounds are not phytotoxic. Incorporating the regulatory and reporter components via Agrobacterium tumefaciens transformation generated transgenic lines of tobacco plants. Germination of the transformed seeds in the presence of murA or RH5992 resulted in induction of the reporter gene activity (up to 420-fold). RH5992 is 100-fold more potent than murA in this system, giving maximal activation at 12.5 µM and 50% activation at ca. 1 µM (Martinez et al., 1999a). Parallel studies using maize protoplasts compared chimeric receptors involving ligand and hinge regions of either the D. melanogaster or H. virescens EcR fused to the A/B/C-domains of the glucocorticoid receptor, showed that RH5992 activates in the presence of GRH, but not GRD (Martinez et al., 1999b). On the other hand, murA (100 µM) activates in the presence of GRD, but not GRH. The preferential activation of GRH by RH5992 is in accord with the higher affinity of this BAH for lepidopteran EcR/USP complexes than for dipteran complexes (Dhadialla et al., 1998), but the lack of activation of GRH by murA is not readily explained and seems to indicate that the conformation of the LBD of the chimeric receptor is significantly altered.
Figure 5.
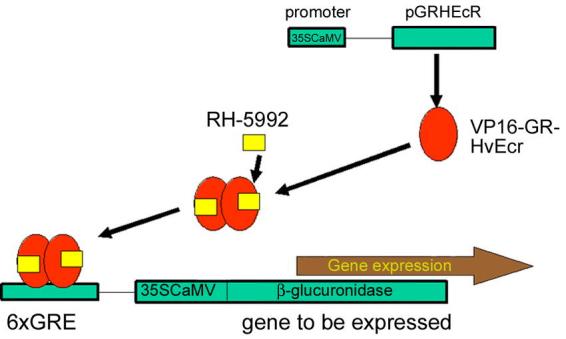
The Syngenta system for plant cells. See text for further details (Martinez et al., 1999a&b).
Unger et al. (2002) have developed a BAH-regulated system for the control of male fertility in maize. Ms45 is a nuclear male fertility gene, which is expressed in anthers. Homozygous recessive mutants are male sterile. The aim was to create a Ms45 construct which would allow male fertility to be restored after application of an elicitor. The hinge and ligand-binding domains (domains D–F) of the Ostrinia nubilalis EcR gene were linked to the VP16-GAL4 or C1-GAL4 transcription activators, under the regulation of the Ubiquitin 1 promoter, which gave constitutive expression of the receptor construct. The Ms45 promoter region was replaced by 5 copies of the yeast 17 bp UASG. Regulatory proteins containing the GAL4 DNA-binding domain bind to UASG. The authors demonstrate that treatment of transformed maize callus with methoxyfenozide (10 µM) induces Ms45 expression. Further, when incorporated into plants, the plants were male sterile in the absence of methoxyfenozide, but fertility was restored by treating plants with methoxyfenozide.
Padidam et al. (2002) have recently developed an ecdysteroid receptor-based gene expression system based on a modified Choristoneura fumiferana EcR and the BAH methoxyfenozide and demonstrated its effectiveness in transgenic Arabidopsis thaliana and Nicotiana tabacum.
In addition to ecdysteroid/BAH controllable systems, other chemically inducible gene expression systems for plants are also being investigated (reviewed in Gatz and Lenk, 1998; Jepson et al., 1998). These systems have been recently compared and reviewed (Padidam, 2003).
Fungi:
When transfected into Saccharomyces cerevisiae, DmEcR is able to transactivate a reporter gene in the absence of USP/RXR or ecdysteroid (ponA or murA) (Dela Cruz and Mak, 1997). Activation is EcRE-dependent, but, unexpectedly, ecdysteroid- and heterodimerisation partner-independent. Interestingly, high affinity specific binding of [3H]ponA (Kd = 1.8 nM) by yeast extracts was dependent on coexpression of EcR and USP (or RXR). Radiolabelled hormone displacement assays for yeast-expressed EcR/USP with ponA, murA, 20E and RH5849 (Dela Cruz and Mak, 1997) indicate similar specificity and affinity to D. melanogaster ecdysteroid receptor complexes in insect systems (Bidmon and Sliter, 1990). Thus, the situation prevailing when ecdysteroid receptors are expressed in yeast cells is apparently very different to that for mammalian or plant cells.
Using the ecdysteroid receptor genes from Choristoneura fumiferana (CfEcR and CfUSP) coexpressed in yeast with a reporter gene containing EcREs, Tran et al. (2001) showed that EcR and USP together (but not individually) induced reporter gene expression in the absence of ligand, with RH5992 (10 µM) only providing a small enhancement in reporter gene expression. Deletion of the A/B-regions of CfEcR, in conjunction with CfUSP, still gave ligand-independent transactivation with some enhancement on addition of RH5992. However, deletion of the A/B-regions of CfUSP (generating Cf▵USP) abolished reporter gene expression, regardless of whether co-expression was with CfEcR or Cf▵EcR and in the presence or absence of RH5992. Together, these data showed that EcR:USP is not suitable for a ligand-dependent transactivation assay in yeast. Replacement of USP with RXRα, RXRβ or RXRγ, when co-expressed with EcR, resulted in no induction of the reporter gene in the presence or absence of RH5992. However, co-expression of GRIP1 (a member of the p160 family of coactivators) and Cf▵EcR:RXR or Cf▵EcR:Cf▵USP resulted in significant ligand-dependent transactivation of the reporter gene activity. The system with RXRβ appeared to have a low sensitivity to RH5992 and other BAHs and was not pursued further. Comparison of three yeast systems (▵EcR:▵USP:GRIP1, ▵EcR:RXRα:GRIP1 and ▵EcR:RXRγ:GRIP1) with an insect cell CfEcR:USP-containing system (L57; DmEcR-negative Kc cells transfected with CfEcR and β-galactosidase reporter controlled by 6 × EcRE and a minimal promoter) using a range of BAHs and murA and ponA showed induction in all systems by active compounds, but i) the degree of induction was far lower in the yeast systems and ii) the ecdysteroids were very much poorer inducers in the yeast ▵EcR:▵USP system than in the insect cells and did not induce the ▵EcR:RXR(α or γ) systems. Further, 9-cis-retinoic acid (a natural ligand of RXR receptors) induced the ▵EcR:RXRα:GRIP1 and ▵EcR:RXRγ:GRIP1 systems, complicating interpretation of results from these systems if they were used in screening processes. In part these problems may derive from poor access of test compounds through the thick yeast cell wall or rapid export from the cells. Tran et al. (2001) provide evidence that use of yeast strains with mutations in certain ABC transporter pathway loci results in improved sensitivity (100-fold for RH5992). Tran et al. (2001) propose their transactivation assay as a screen to identify potential insecticides with ecdysteroid agonist activity.
Commercially available systems:
The system devised by No et al. (1996) has been developed and commercialised by Invitrogen (http://order.invitrogen.com/). The company provides kits consisting of mammalian cells (CV-1, HEK293 and CHO) stably expressing a functional ecdysteroid receptor from pVgRXR, the inducing agent (now ponA) and a vector by which the gene of choice can be introduced into the cells, after introduction of the gene into the vector by simple recombination using Cre recombinase. The components are also available individually. The pVgRXR expression vector includes VgEcR, RXR and a gene for Zeocin resistance, which allows for selection of stable cells expressing the heterodimeric receptor (VgEcR:RXR).
Stable ecdysteroid-inducible mammalian cell lines can be difficult to establish because of either high basal expression of the target gene or poor induction of gene expression, because of low expression of the receptors (VgEcR and RXR) or the transgene. Ideally, stable cell lines expressing the receptors should be established first and the cells should be screened by transient expression of an ecdysteroid-regulatable transgene to identify those expressing the receptor proteins effectively. An improvement on Invitrogen's pIND/lacZ reporter system (which generates β-galactosidase activity has been reported (Wakita et al., 2001), which uses a firefly luciferase reporter system. This considerably reduces analysis time (15 s, rather than 2 h) and obviates background interference from endogenous β-galactosidase activity. A similar advance has been suggested by Lüers et al. (2000) who prepared an expression plasmid for green fluorescent protein (EGFP) and the reporter protein of interest. The co-inducible production of EGFP permits the visual verification of target gene expression and the selection of expressing cells by flow-cytometry.
As described above (Section 2.3.1), RHeoGene LLC are commercialising their ecdysteroid receptor-based gene switching system (www.rheogene.com) and are identifying specific receptors/ligand pairs which allow the simultaneous, but independent, regulation of transfected genes (Kumar et al., 2002). Further, hybrid receptors are being optimised to give very low basal activity and high induction on addition of ligand (Palli et al., 2003). A two-hybrid format switch where the GAL4 DNA-binding domain was fused to CfEcR domains D, E and F and the VP16 activation domain was fused to mouse RXR domains E and F, transactivating a reporter gene under the control of GAL4 response elements and a synthetic TATAA promoter was found to give the best combination with rapid turn-on and turn-off responses on the addition and removal of RG-102240 (GS-E), respectively.
Ecdysteroid systems vs. other systems
Fussenegger (2001) provides a comprehensive description of the heterologous molecular switching systems currently under consideration. Each has its own advantages, but none fulfils all the desirable criteria perfectly. From the time of early studies, transgenic ecdysteroid-inducible gene expression systems in mammalian cells have appeared to possess lower basal activity and higher inducibility than tetracycline-based systems (No et al., 1996). Senner et al. (2001) compared 3 inducer systems (tetracycline, dimerizer and ecdysteroid) in one system (rat C6 glioma cells) under identical conditions. Each system required transient transfection with two plasmids (a regulator plasmid and a reporter plasmid) and treatment with an elicitor (inducers for the ecdysteroid and dimerizer systems and a repressor for the tetracycline system). The ecdysteroid system provided the highest induced activity, but the authors conclude that each of the systems may be beneficial, depending on what the experimental goals are. Van Craenenbroeck et al. (2001) compared the tetracycline and ecdysteroid systems to regulate the expression of neurotransmitter receptors in mouse fibrosarcoma L929sA and HEK293 cells. The tetracycline system resulted in higher levels of the neurotransmitter receptors being expressed, but the ecdysteroid system gave more tightly regulated expression. Moreover, this study underlined the importance of the genetic background of the cells being used.
Morgan et al. (1999) compare several exogenously regulatable promoter systems for their suitability for the study of the functions of genes implicated in aging.
Ecdysteroid specificity
Gene expression systems in mammalian and plants cells possess markedly different ecdysteroid specificities to ecdysteroid receptors in insect systems. Both the affinity and specificity seem to be affected. Thus, the generally accepted endogenous hormone in insects, 20E, is inactive in transgenic systems. Two phytoecdysteroids, murA and ponA, are normally used to activate the transgenic systems, but even these are required at least 100-fold higher concentrations than in insect systems; e.g. EC50 values for murA and ponA in the Drosophila melanogaster BII bioassay are 2.2 × 10−8M and 3.1 × 10−10M, respectively (Dinan et al., 1999a), while concentrations of 1 – 10 µM are required to induce transgenic expression. The basis of this altered affinity/specificity is not clear and it could derive from: (i) altered metabolism, (ii) use of RXR, rather than USP, (iii) altered transportation into cells, (iv) fusion of the EcR ligand-binding domain to the GR DNA-binding domain and/or VP16 to form VgEcR, (v) the different properties of mammalian transcription factors, enhancers, repressors etc.
A limited investigation of the ecdysteroid specificity of VgEcR/RXR in CV-1 cells has been performed (Saez et al., 2000). MurA, ponA and 14-deoxymuristerone A were almost equivalently active (EC50 = ca. 5 × 10−7M), with ponasterone C being moderately active (EC50 = 2 × 10−5M), polB being weakly active and 20E, inokosterone, makisterone A, E, 2-deoxyecdysone, 20E 22-acetate and 2-deoxy-20-hydroxyecdysone being inactive or only very weakly active at 10−4M. This study also showed that the presence of a natural (9-cis-retinoic acid) or synthetic (LG268 or LG1069) RXR ligands, while inactive in itself, potentiated the activity of ponA by 3- to 5-fold.
Availability of ligands
Ecdysteroids:
MurA has only been isolated once in large amounts (Canonica et al., 1972), and then from a Himalayan plant (Ipomoea calonyction). Consequently, world supplies of this phytoecdysteroid became very restricted and did not suffice for full-scale trials of ecdysteroid-induced transgenic systems. However, Sequoia Sciences (http://www.sequoiasciences.com) report on their website that they have recently re-isolated murA. PonA, which has been isolated from several named plant species and which can be chemically generated from 20E (Heinrich, 1970), is also active. Examination of the ecdysteroid specificity in more detail could result in the identification of more active inducers. While in vitro work may not suffer unduely from the poor activity of currently used ecdysteroid inducers, other than having to use much larger amounts of expensive chemicals, the in vivo prospects for transgenic systems using ecdysteroids would be enormously enhanced if they were as active as in insect systems.
Bisacylhydrazines (BAHs):
Bisacylhydrazines were identified as non-steroidal agonists of ecdysteroid receptors in 1988 (Wing, 1988). Their chemical simplicity, low mammalian toxicity and selectivity for certain Orders of insects has led to several being developed as insecticides (Dhadialla et al., 1998). They could also be used for the induction of transgenic systems (Carlson et al., 2001) and, as is apparent above, RH-5992 has found application in this context. Further analogues (e.g. GS-E; Fig. 1) have been identified which appear to be more potent for use with mammalian systems (Carlson, 2000). However, the very limited water solubility of these compounds may limit their application in vivo.
Figure 1.
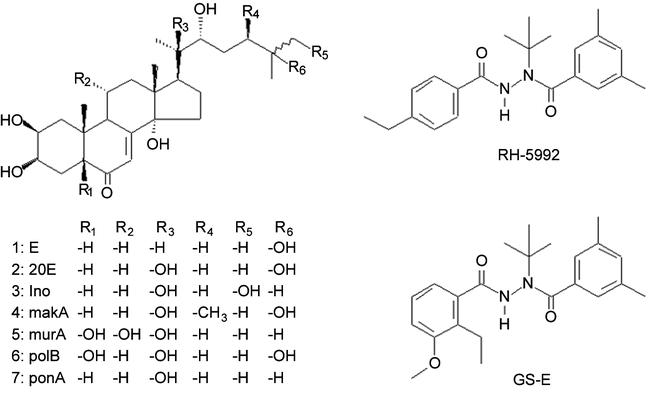
Structures of ligands used for ecdysteroid-inducible gene expression systems in mammalian and plant cells.
Modified receptors:
A further approach to overcoming the current lack of really potent ligands for transgenic induction would be to modify the ligand-binding domain of the transgenic EcR to either enhance the affinity for a particular analogue, or to alter the specificity, so that a readily available analogue (e.g. 20E) or a non-dietary ecdysteroid is recognised. Cloning and sequence data for ecdysteroid receptor proteins (EcR and USP) from a range of arthropod species provide the basis for site-directed mutagenesis to modify specific amino-acid residues. Both this and the previous approach require a more thorough understanding of ecdysteroid receptor recognition, not only in D. melanogaster (Dinan et al., 1999a; Ravi et al., 2001), but also in other arthropod species and in transgenic systems. The ultimate goal of such studies is to engineer a range of EcR proteins, some of which respond to non-steroidal inducers, but not to ecdysteroids, while others respond to selected ecdysteroids, but not to other classes of agonists (Graham, 2002). Strategies are being developed for the synthesis of further non-steroidal ligands for selective activation of ecdysteroid receptors (Tice et al., 2003) and for the targeted modification of ligand specificity of ecdysteroid receptors (Kumar et al., 2002).
Registration problems
Development of ecdysteroid systems for human therapeutic use may be hampered by the steroidal nature of ecdysteroids and the insecticidal origin of BAHs, which may prejudice their use as elicitors, this being in spite of the fact that both ecdysteroids and BAHs have low mammalian toxicities and ecdysteroids are a normal (but small) component of the human diet. For plant systems, ecdysteroids per se cannot be considered because of penetration problems and BAHs may not be acceptable because of the enhanced risk of development of resistance to insecticidal analogues. However, use might be restricted to specified crops under conditions where exposure to sensitive insect species is minimal.
Biochemical problems
Although the systems developed to date are effective for use in in vitro expression systems, the requirements for an effective in vivo system are much more stringent. In this context, one can identify the following aspects of ecdysteroid-regulatable systems which would need to be improved in order to generate a medically viable system:
Integration of heterologous DNA into host cells is not site-specific and is unpredictable with regard to copy number.
The current systems are genetically complex, requiring both VgEcR and RXR.
The artificial transactivator is potentially immunogenic.
RXR is a reluctant dimerization partner for EcR and, therefore, very high cellular RXR concentrations are required. Overexpression of RXR may results in pleiotropic effects in mammals.
The maximal expression levels achieved are modest.
Most ecdysteroids are not very active. Only muristerone A and ponasterone A are effective.
Ecdysteroids are not orally available
Ecdysteroids or ecdysteroid analogues are not likely to get approval for human therapeutic use.
Prospects
There is little doubt that ecdysteroid-regulated transgenic systems have considerable potential for in vitro work. The applied potential is somewhat more questionable at present, owing to the following current limitations: i) genetic complexity, ii) altered affinity and selectivity of VgEcR for ligands and iii) potential problems in the registration of ecdysteroids and BAHs for human therapeutic uses or with plant transgenic systems. However, significant progress is being made in designing chimaeric receptors which would allow only one trans-acting factor to be transfected. It is only a matter of time until the reasons for the altered affinity and selectivity of ecdysteroid receptors in mammalian and plant cells are elucidated and more efficient systems are developed either by identifying more effective ligands or site-directed mutagenesis of EcR to enhance affinity for currently used ligands. Although ecdysteroids and BAHs are nontoxic to humans, general public resistance to steroids and insecticides may hamper their registration.
Ecdysteroid metabolism in mammals, including humans
Although the question of mammalian metabolism is certainly of importance for the practicability of the in vivo use of ecdysteroid-inducible gene expression systems (with the aim of using them for gene therapy), it is not well documented at the present time. Ecdysteroids have a very low toxicity in mammals: in the mouse, the LD50 of 20E is 6.4 g/kg (for intra-peritoneal injection) and it is >9 g/kg when given orally (Matsuda et al., 1970; Ogawa et al., 1974). Up to now, studies have concerned mice, rats, lambs and humans, and all have shown that these molecules are short-lived in mammals. Several strategies have been used to analyse the metabolic fate of ecdysteroids.
Ecdysteroids are rapidly eliminated
In the case of humans, two different studies have been performed. Simon and Koolman (1989) analysed the pharmacokinetics of E and 20E (given orally, 0.2 mg/kg b.w.) to a male volunteer, by monitoring with a radioimmunoassay the subsequent plasma and urine titres. This gave an effective half-time (EHT) of elimination of 4 hours for E and 9 hours for 20E. In lambs, EHT for 20E was shown to depend strongly on the mode of administration, with values of 0.4, 0.2 and 2 hours after oral, intravenous and intramuscular administration, respectively (Simon and Koolman, 1989). The method used did not allow the detection of metabolites, if present. The half-life seems shorter in smaller mammals, with reported values of 8.15 min for 20E in mice (Dzukharova et al., 1987). More recently, Albanese et al. (2000) found a plasma half-life of 48 min for ponA in mice after intra-peritoneal injection of 750 µg of this compound.
Both urinary and faecal routes seem to be used for the elimination of the administered molecules. In mice, the faecal route was found to be the major one by Hikino et al. (1972a&b) and Lafont et al. (1988), although Dzukharova et al. (1987) found that faecal and urinary routes were equally important. Such a question can be easily assessed only by the use of radiolabelled molecules, but no data are available for humans. Kinetic studies in mice showed that ecdysteroids were taken up by the liver and then excreted into the gut via the bile (Hikino et al.,1972a&b; Lafont et al., 1988).
Metabolic conversions
Another question concerns whether ecdysteroids undergo metabolic conversions in mammals. The presently available data are not fully consistent. In mice, Girault et al. (1988) analysed the faecal metabolites of injected E and isolated unchanged E, a major metabolite identified by MS and proton NMR as 14-deoxyecdysone together with molecules with a fully reduced B-ring and, additionally, epimerized in position 3 (Figure 6A). Such a metabolism is reminiscent of the hepatic reduction of the 4-en-3-one on ring-A of vertebrate steroid hormones, whereas dehydroxylation resembles that of bile acids and could result from the actions of anaerobic intestinal bacteria.
Figure 6.
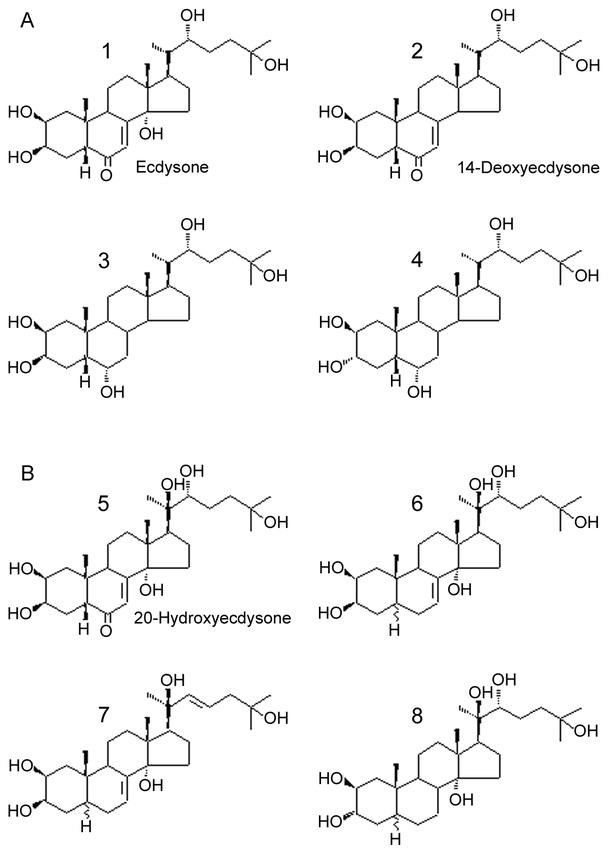
Major E and 20E metabolites in Mammals (see text for details). A: E metabolites (2–4) isolated from murine faeces (Girault et al., 1988); B: 20E metabolites (6–8) isolated from rat urine (Ramazanov et al., 1996).
More recent studies were performed on ingested 20E in rats (Ramazanov et al., 1996) and humans (Tsitsimpikou et al., 2001). In these cases only urine was analyzed. Ramazanov et al. administered 20E to 40 rats (50 mg/kg) directly in stomach with a special probe, and they collected urine (3.5 L) over the following 10 days. After several chromatographic steps, they isolated unchanged 20E and three new metabolites, which were analyzed by IR and mass spectrometry. The IR spectra showed the disappearance of the signal at 650 cm −1 (7-en-6-one) and the structures were deduced from MS data (Figure 6B).
Tsitsimpikou et al. (2001) analysed the urine of a volunteer having ingested 20 mg of “Ecdysten™” (a commercial preparation containing 20E – see section 6); they collected urine over 5 days and analysed ecdysteroids by GC-MS after derivatization. They found, together with 20E, two less hydroxylated metabolites, which they tentatively identified as 2d20E and 2dE by comparison with available reference molecules.
Mass spectrometry does not provide sufficient information, and only NMR can allow an unambiguous determination of structures. Anyway, it seems reasonable to assume that modification of the B-ring and dehydroxylation are general features of ecdysteroid metabolism in mammals.
Conclusions/prospects
There is rapid catabolism/elimination of ecdysteroids, which means that large amounts would have to be used in order to maintain circulating levels above the concentration required for gene switches systems to be activated. Alternatively, slow-delivery systems like subcutaneous implants represent another way to maintain sustained ecdysteroid levels for several days (Albanese et al., 2000). Another remaining question concerns the metabolism in peripheral tissues. As we have seen with mice, the observed conversions are most probably performed by hepatocytes and intestinal bacteria. It would be of interest to determine whether other mammalian tissues are able to metabolise ecdysteroids, and the nature of the reactions they can perform.
Whether side-chain cleavage between C-20 and C-22 (and possibly also between C-17 and C-20) can take place is a very important question which remains to be investigated by using ecdysteroids labelled on the steroid nucleus, as labelling on the side-chain would be lost if such a reaction would occur. This question seems particularly important for several reasons: (1) cleavage between C-20 and C-22 would result in the formation of 21C steroids that would share some resemblance with vertebrate neurosteroids (Lafont and Sláma, 1995), and (2) in some pharmacological studies rubrosterone (2β,3β,14α-trihydroxy-5β-androst-7-ene-6,17-dione) was as active as 20E (Otaka et al., 1968).
Pharmacological effects of ecdysteroids on vertebrates
The pharmacological actions of ecdysteroids on vertebrates have been reviewed in several previous articles (Burdette, 1962, 1972; Ogawa et al., 1974; Syrov, 1984, 1994; Sláma and Lafont, 1995; Xu et al., 1997; Syrov, 2000; Kholodova, 2001; Báthori, 2002). We will therefore focus on some aspects only, especially on those where recent developments have occurred. The most important data are summarised in Table 1.
Table 1.
Pharmacological effects of ecdysteroids on mammals or humans (see also Sláma and Lafont, 1995 for additional references – in red : references to patents)

Ecdysteroids and growth (Table 2)
Table 2.
Effects of ingested or injected ecdysteroids on growth of various vertebrate species (in red, reference to patents).

The anabolic effects of several phytoecdysteroids (20E, cyasterone, turkesterone, viticosterone E – see structures on Ecdybase) on mice or rats were reported long ago (see e.g. Okui et al., 1968; Syrov and Kurmukov, 1975a&b; 1976a–c, Syrov et al., 1978, 1981a; Stopka et al., 1999). Growth-promoting effects have also been more recently reported for pigs (Kratky et al., 1997) and Japanese quails (Koudela et al., 1995; Sláma et al., 1996). In many instances however, these effects are not spectacular when considering the growth (weight) curves as they are observed during certain phases of growth or for one sex only and, in many cases, adequate statistical analyses are lacking. Nevertheless, even small effects (i.e. <5 % increase) on growth could be of economical interest for nutritionists, but their firm establishment requires the use of large numbers of animals, which is hardly feasible with large mammals. The addition of E to sheep food increases body growth rate and also wool growth (Purser and Baker, 1994). Surprisingly, these effects were obtained with minute amounts of ecdysone (0.02 µg/kg per day!), and were more evident when animals were fed on a poor quality diet, which indicates that E improves food utilization. In this case, it has been suggested that the effect results from the toxicity of E towards rumen protozoa, but this has not been fully established. In fact, through a stimulation of protein synthesis (and/or a reduction of protein catabolism), ecdysteroids would increase the lean body mass. In pigs, doses of 0.2–0.4 mg/kg/day resulted in better nitrogen retention and a body weight increase of 112–116% relative to controls, while food consumption was lowered by 11–17% (Kratky et al., 1997). Other experiments used diets supplemented with ecdysteroid-containing plants (e.g. Rhaponticum carthamoides) and reported similar growth-promoting effects on pigs over a 30-day period (Selepcova et al., 1993b). In quails, 20E in the diet promoted increased growth (115% of controls), but this was associated with a decreased index of food conversion (Sláma et al., 1996). From these data, it appears difficult to draw general conclusions.
Ecdysteroids and physical performance
20E is claimed to have tonic properties (Abubakirov et al., 1988). Indeed it stimulates muscle growth, provided that protein supply is adequate. Such anabolic effects result in increased physical performance without training (Chermnykh et al., 1988). This was for instance demonstrated using the forced swimming test with rats: animals given ecdysteroids for one week were able to swim for significantly longer times (Azizov and Seifulla, 1998). These effects are similar to those of anabolic steroids. 20E is also able to increase muscle ATP content in vitamin D-deprived rats (Kholodova et al., 1997).
Ecdysteroids: effects on cellular proliferation and differentiation
Wound-healing effects of ecdysteroids have been described (Syrov and Khushbatkova, 1996; Darmograi et al., 1998). 20E (applied at 0.1% w/w in liposomes) shortens the duration of skin repair after superficial wounding and 20E (2 × 10−4M) stimulates keratinocyte differentiation in vitro (Detmar et al., 1994), an effect measured by the increase of the activity of transglutaminase (an enzyme involved in protein connection through isopeptidic bond formation). Accordingly, ecdysteroids show psoriasis-inhibiting effects (Inaoka et al. 1997). These results have led to many patents concerning the use of ecdysteroids in cosmetics (Lin and Lin, 1989; Meybeck and Bonté, 1990, 1993; Meybeck et al., 1994; Tsuji et al., 1995a&b; Darmograi et al. 1998; Meybeck 1999a&b). In this context, the incorporation of 20E or its acyl ester (2,3,22-tripalmitate) into liposomes has been tested as a slow-release form (Politova et al., 2001).
20E administered orally to rats (5 mg/kg) accelerates the healing process after an experimental bone fracture (Syrov et al., 1986a), and the same molecule (10–100 ng/ml) can stimulate the in vitro proliferation of rat osteosarcoma UMR106 cells (osteoblasts) by 41% (Gao and Wang, 2000). Similarly, 20E stimulates proliferation of human umbilical vein endothelial cells (Lin et al., 1997; Wu et al., 1998b), and several phytoecdysteroids can stimulate erythropoiesis in rats (Syrov et al., 1997b).
The effects of ecdysteroids on tumorous cell proliferation are somewhat conflicting: Lagova and Valueva (1981) reported that 20E (0.1–300 mg/kg, subcutaneous injections for 5 days) was mainly ineffective on tumour growth in mice, but it stimulated the growth of mammary gland carcinomas in mice and rats. El-Mofti (1987, 1994) reported that E was able to induce neoplastic lesions in toads and mice; other authors reported inhibitory effects on tumor cell proliferation (Hirono et al., 1969; Burdette, 1974; Shibatani et al., 1996). More recently, Konovalova et al. (2002) showed that injected 20E had a synergistic effect with low doses of an antitumour drug (cis-platin). Most probably, the results may differ according with the cell types, the nature and concentration of ecdysteroids used, and this clearly requires more extensive studies. In addition, genoprotective effects of ecdysteroids have been reported (Gubskii et al., 1993; Levitskii et al., 1993a&b, 1996; Chabanny et al., 1994); ecdysteroids can prevent chromatin damages induced by various chemicals.
Ecdysteroids and protein synthesis
Stimulatory effects of ecdysone on protein synthesis were reported as early as 1963 (Burdette and Coda, 1963), and the discovery of phytoecdysteroids made these molecules available in large amounts for pharmacological assays. It was rapidly shown that ecdysteroids were able to stimulate protein synthesis in mouse liver (Okui et al., 1968; Otaka et al., 1968, 1969a&b). In fact, it was shown that 20E stimulates the incorporation of [14C]leucine in a cell-free translation system (rat liver polysomes), i.e. it increases the efficiency of the translational machinery (Syrov et al., 1978). Such conclusions have been confirmed and extended to other tissues, especially heart and muscles (Syrov et al., 1975a; Aizikov et al., 1978; Khimiko et al., 2000). Recent structure-activity studies (Syrov et al., 2001) as measured by a stimulation of [14C] aminoacid incorporation into proteins showed that among the compounds tested turkesterone was the most active, followed by cyasterone and 20E.
Ecdysteroids and glucose metabolism
It was shown early on (Table 3) that 20E given per os to rats reduces hyperglycaemia induced either by glucagon or by alloxan treatment (Matsuda et al., 1970; Uchiyama and Ogawa, 1970; Yoshida et al., 1971, Uchiyama and Yoshida, 1974). In fact, 20E stimulates the incorporation of glucose into glycogen and protein in mouse liver (Yoshida et al., 1971) and more generally it enhances glucose utilization by tissues (Syrov et al., 1997a). The mechanism involved seems to be an increase of tissue sensitivity to insulin (Kosovsky et al., 1989) and preparations containing phytoecdysteroids have been proposed as oral antidiabetics (Takahashi and Nishimoto, 1992; Yang et al., 2001). Depending on the extent of hyperglycaemia, phytoecdysteroid effects may be more or less pronounced that those of manilil, a widely used pharmacological molecule (Kutepova et al., 2001).
Table 3.
Effects of ingested or injected ecdysteroids on carbohydrate metabolism (in red, reference to patents).

Ecdysteroids and lipid metabolism
Ecdysteroids display hypocholesterolaemic effects (Lupien et al., 1969; Mironova et al., 1982; Syrov et al., 1983), through a reduction of cholesterol biosynthesis and an increase of its catabolism (Uchiyama and Yoshida, 1974). 20E (5 mg/kg per os) stimulates the conversion of cholesterol into bile acids in rats (Syrov et al., 1986b), and such an effect is reminiscent of some oxysterols (Schroepfer, 2000). In connection with these effects, ecdysteroids may also have antiatherosclerotic actions (Matsuda et al., 1974; Syrov et al., 1983). Intraperitoneally injected 20E (0.5 mg/kg in rats) also enhances [14C]acetate incorporation into liver triglycerides and reduces triglyceride lipase activity (Catalán et al., 1985).
Ecdysteroids: a “universal medicine“?
An impressive number of papers dealing with ecdysteroid effects are available in the literature. They concern almost every physiological function, and we will give below a brief insight of the published data. It must be noted, however, that in many instances that, in addition to the difficulties caused by language barriers, the experiments are not always described with all the desirable details.
Ecdysteroids improve nervous function: in early studies, it was shown that 20E induced glutamic decarboxylase (an enzyme involved in GABA biosynthesis) in rat brain (Chaudhary et al., 1969), and that E was able to induce acetylcholinesterase in rat brain too (Catalán et al., 1984). More recently, ecdysteroids were shown to represent neuron-protective agents; they reduce glutamate-induced cell death in cortex neurons of rat foetuses and they are proposed as a therapy against mental and behavioural disorders (Aikake et al., 1996). In addition, they may protect against amnesia induced by diazepam or alcohol (Xu et al., 1999). Similar neuroprotective effects have been described for progesterone and oestradiol mixtures in animal models of neurodegeneration (Vongher and Frye, 1999).
Ecdysteroids stimulate hepatic functions: 20E accelerates recovery after hepatitis induced by heliotrine treatment (Syrov et al., 1981b). 20E and other ecdysteroids (turkesterone, cyasterone) administered (10 mg/kg) to rats with hepatitis induced by subcutaneous injection of carbon tetrachloride prevent its hepatotoxic action (Syrov et al., 1992). Moreover, a pretreatment with 20E (5 mg/kg) for one week will reduce the effects of a subsequent heliotrine treatment (Badal'yants et al., 1996).
Ecdysteroids improve heart and lung function: 20E has been recommended for the prevention of myocardial ischaemia, arrhythmia and is described as enhancing VEGF expression (Wu, 2001). An antiarrhythmic effect of 20E was also reported by Kurmukov and Yermishina (1991) and Yang et al. (1996), and an extract of Leuzea carthamoides containing high amounts of 20E also showed a similar effect (Maimeskulova and Malslov, 2000). In rabbits experimentally rendered atherosclerotic (by a high cholesterol diet), 20E (10 mg/kg/day per os) given for 28 days was able to increase Na+/K+ ATPase in myocardium (Khushbaktova et al., 1987). Intravenous injection of 20E showed also a therapeutic effect after lung contusion (Wu et al., 1997, 1998a).
Ecdysteroids improve renal function: when rats are given a nephrotoxic mixture (uranyl acetate + glycerol), 20E (5 mg/kg) seems thereafter able to restore a normal glomerular filtration rate and to suppress albuminuria (Saatov et al., 1999; Syrov and Khushbaktova, 2001).
Ecdysteroids and the immune system: various immunomodulatory effects of ecdysteroids have been described. Single intraperitoneal injections of various ecdysteroids (20E, 2dE, 2d20E, polB, turkesterone, 1–5 mg/kg) increase the concentration of antibody-forming cells in the spleen of mice immunised with sheep red blood cells (Sakhibov et al., 1989). Low (7.5×10−12–7.5×10−8 M) concentrations of 20E induce the activation (E-rosette formation test) of human lymphocytes (Trenin et al., 1996; Trenin and Volodin, 1999). Low to moderate (10−12–10−5 M) concentrations of 20E or other ecdysteroids stimulate, whereas higher (10−4M) concentrations eventually inhibit, DNA synthesis in concanavalin A – activated lymphocytes (Kuzmitsky et al., 1990; Fomovska et al., 1992; Chiang et al., 1992).
20E (10–20 mg/kg/day per os) has antiinflammatory properties similar to cortisone acetate in rats and mice (Kurmukov and Syrov, 1988; Fomovska et al., 1992) and turkesterone improves lung defence mechanisms in diabetic rats (Najmutdinova and Saatov, 1999). 20E was shown to inhibit in a dose-dependent fashion (10−9–10−4 M) histamine release from rat peritoneal mast cells induced by anti-IgE or concanavailin A (Takei et al., 1991). Taniguchi et al. (1997), however, could not observe any antiinflammatory effect of 20E given orally to rats (5 mg/kg/day for 7 days).
Ecdysteroids have antioxidant properties: 20E has antioxidative and anti-free radical properties (Osynska et al., 1992) and it can thus reduce lipid peroxidation (Kuzmenko et al., 1997, 2001). Several models were used in these studies, as the chemiluminescence of blood serum induced by H2O2 using rats receiving a vitamin D-deficient diet eventually supplemented with 0.1 mg 20E/kg per day, or the uptake of oxygen by methyl linoleate micelles in the presence or absence of 20E.
Are ecdysteroids toxic to microorganisms?: there are a few reports about antimicrobial activity of ecdysteroids. However, Ahmad et al. (1996) reported antifungal and antibacterial activity of 20E at rather high concentrations (between 100 and 400 µg/ml, i.e. 2–8 × 10−4 M). An antimicrobial activity of 20E and its acetates was also observed by Volodin et al. (1999). Toxic effects on protozoa have also been reported; rabbits receiving 20E per os (5 mg/g/day for 3 months) showed a reduced infection with Lamblia duodenalis (Syrov et al., 1990), and the improvement of ruminant productivity by ecdysone was also interpreted by its toxicity towards rumen protozoa (Purser and Baker, 1994).
Ecdysteroids are not toxic to vertebrates: ecdysteroids have a very low toxicity (LD50 > 6g/kg), they are not hypertensive and, in spite of their anabolic action, they would have neither androgenic nor oestrogenic (or antioestrogenic) effects; they induce no virilisation and they do not induce significant changes in castrated animals (e.g. Prabhu and Nayar, 1974). All together this suggests that ecdysteroids are attractive compounds for a wide array of uses, which have been proposed, and of course it would be of particular interest to understand more precisely their mode(s) of action in mammals.
Genomic and/or non-genomic effects of ecdysteroids?
Do ecdysteroids have genomic effects on vertebrates?
In insects, ecdysteroids have well-known genomic effects which involve nuclear receptors (see Section 2). When considering the molecules in 3-dimensions, it is clear that they show striking differences to vertebrate sex or adrenal steroids, and their full cholesterol side-chain most probably prevents any binding to the receptors of these vertebrate hormones. Recently, however, it has been found that some previously “orphan” nuclear receptors (e.g. LXR, PXR) bind endogenously produced oxysterols (Janowski et al., 1999 ; Schroepffer, 2000) or have a broad specificity and may bind a wide array of xenobiotics including several steroids (Jones et al., 2000). Given this broad ligand specificity, these proteins might rather function as “endocrine sensors” rather than “true receptors” (Evans, 2002). So, until ecdysteroids are directly tested for binding to such receptors, it remains conceivable that they may have transcriptional effects through binding to some nuclear receptor(s); indeed early studies showed a rapid in vivo stimulation by 20E of the incorporation of [14C]orotic acid into RNA in mouse liver (Uchiyama and Otaka, 1974).
Ecdysteroids: membrane effects?
Membrane effects of steroids are nowadays well documented and they may proceed through three different pathways (Figure 7). According to Brann et al. (1995), these effects may either involve: (1) the dissolution of ecdysteroids in the membrane bilayer and a change in the environment of some membrane proteins (and hence of their activity), (2) their interaction with a specific membrane receptor, which will activate some transduction mechanism, or (3) their binding to a modulatory site of the receptor for another molecule. These different effects are not mutually exclusive. Very recently, a membrane progestin receptor involved in fish oocyte meiotic reinitiation was cloned and its physiological relevance was fully established (Zhu et al., 2003).
Figure 7.
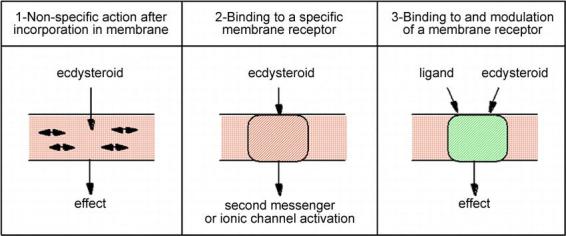
Three possible ways for a membrane effect of ecdysteroids (adapted from Brann et al., 1995).
Dissolution of edysteroids in the membrane lipid bilayer:
In order to test for the first hypothesis, Tuganova and Kotsyuruba (1996) developed experiments designed to analyse the dissolution of ecdysteroids in human erythrocyte membranes. They did not perform direct experiments, i.e. by measuring the incorporation of radiolabelled ecdysteroids. In a first set of experiments, erythrocytes were first incubated with various steroids (10−6 M) and then with [3H]cholesterol; both 2d20E and 20E pretreatments reduced the radioactivity associated with membrane fractions. In a second set of experiments, the authors first incubated erythrocytes with various concentrations of 20E (10−14 to 10−10 M) then with either radiolabelled cholesterol, cholecalciferol or calcitriol; 20E reduced mainly calcitriol incorporation. Such experiments support the idea that ecdysteroids can be incorporated into membrane bilayers, although they do not constitute an absolute proof. It is tempting to make a relation between these results and the rapid effect of 20E on Na+/K+ ATPase activity in D. melanogaster salivary gland cells (Schneider et al., 1996).
Rapid membrane effects:
A rapid increase of cGMP and a decrease of cAMP levels in mouse plasma, together with a decrease of PKA activity in liver were described 40 min after an intraperitoneal injection of 10 µg 20E (Catalán et al., 1979a&b, 1982). More recently, it was shown that 20E evokes rapid (1–2 min) and transitory effects on membranes (Kotsyuruba et al., 1995a–c, 1998a&b, 1999); 20E increases the pool of free arachidonic acid and the synthesis of leukotrienes and prostaglandins. Such responses were observed with different cell types (hepatocytes, erythocytes, lymphocytes, macrophages etc.). In many instances the effects of 20E resemble those evoked by calcitriol (1,25OH-D3), a molecule often used for comparison by those working on ecdysteroid pharmacology (Barsony and Marx, 1988). The same effects were also produced by 20E bound to magnetite nanoparticles (Mykhaylyk et al., 1999; 2001), a formulation which should prevent 20E diffusion into target cells, and thus restricting its possible action(s) to the plasma membrane level.
We should emphasise here that ecdysteroids can be recognised by membrane receptors in arthropods: ecdysteroids can be detected by taste cell receptors both in Crustacea (Tomaschko, 1999) and insects (Tanaka et al., 1994; Descoins and Marion-Poll, 1999), and in vertebrates too steroids can work as pheromones (see e.g. Sorensen et al., 1990). Thus, such a mode of action is conceivable.
Neuromodulatory actions:
Such effects are well documented for vertebrate neurosteroids, which may modulate the response of neurotransmitter receptors to their cognate ligands. The binding of the steroid alone has no apparent effect. Thus, the GABAA receptor possesses (in a domain separate for the neurotransmitter binding site) a binding site for steroids, which may therefore modify the response to GABA. In a similar way, 20-hydroxyecdysone showed a neuromodulatory effect on GABAA receptor of rat cortical neurons (Tsujiyama et al., 1995; Sasa et al., 1996), although this was observed for rather high concentrations (10–100 µM). In connection with this effect, 20E showed an antiepileptic activity in rats (Hanaya et al., 1997); when 20E was given orally (100–200 mg/kg) to spontaneous epileptic rats, it was able to reduce tonic convulsions.
Some recent data
Recently, Constantino et al. (2001) fortuitously made a crucial observation. Using the Invitrogen® ecdysteroid-inducible expression system to analyse the transduction mechanisms of interleukin-3 (IL-3) in a pro-B lymphocyte cell line, they found in control experiments that murA and ponA were able to potentiate the IL-3-dependent activation of PI 3-kinase/Akt pathway in non-transformed cells.
Given the central role of the Akt/PKB pathway in mammalian cell metabolism (e.g. Brazil and Hemmings, 2001; Whiteman et al., 2002), such results provide an interesting basis for explaining in a single way many effects of ecdysteroids on mammals, as concerns their hypoglycaemic, antiapoptotic and anabolic actions (Figure 8). The available data do not allow to decide whether edysteroids act on the IL-3 receptor itself or on a downstream step.
Figure 8.
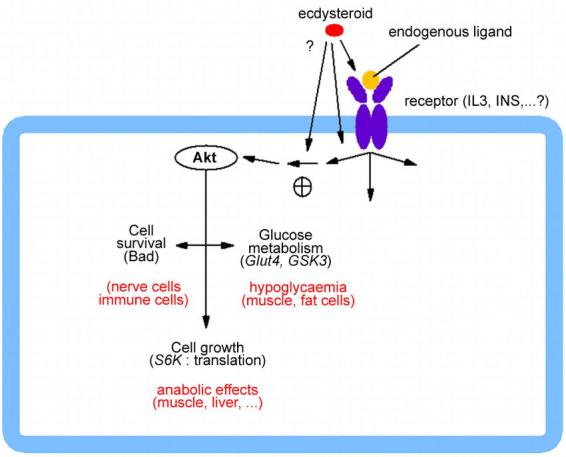
A working hypothesis for ecdysteroid action on mammalian cells: a stimulation of the Akt/PKB pathway would explain a large set of the described effects of 20E on mammals (see text for details). Bad: a proapoptotic factor, which is inhibited upon phosphorylation by Akt; Glut4: glucose transporter type 4; GSK3: glycogen synthase kinase-3; IL3: interleukin-3; INS: insulin; S6K: ribosomal protein S6-kinase.
Where is 20-hydroxyecdysone found?
Phytoecdysteroids are found in many plant species, where they can reach concentrations above 1–2% of the plant dry weight (e.g. Lafont, 1998, Dinan, 2001). Ecdysteroid-rich species are found among ferns and angiosperms and some of these species are either very common (e.g. the fern Polypodium vulgare) or they are cultivated on a large scale for their pharmacological properties (e.g. Leuzea, Pfaffia, Cyanotis). Given the still limited market at the moment, a few plant species only (Table 4) are currently used as a source of phytoecdysteroids: (1) Leuzea (= Rhaponticum) carthamoides (Asteraceae) from Eastern Europe countries, where it is cultivated as a remedy in traditional medicine, (2) Pfaffia (in fact a group of related species) = Brazilian ginseng (= Suma), again a plant used in traditional medicine and (3) Cyanotis vaga or C. arachnoides, a monocotyledonous plant, extracts of which are used on a large scale also for the synchronization of spinning in silkworm larvae (Guo, 1989; Chandrakala et al., 1998).
Table 4.
Plants currently used to obtain the ecdysteroids used for various preparations

Over 140 different preparations containing ecdysteroids for oral use can be found on the market (Table 5). We may distinguish several categories among them: (1) those containing crude or semi-purified plant extracts (plant powders, or alcoholic extracts - elixirs) and (2) those containing “pure” 20E or a defined ecdysteroid mixture. Most of them are proposed for use by bodybuilders, but some have been designed for more specific users (e.g. golfers), or for animals (dogs, horses). In addition, ecdysteroids are also present in at least two cosmetic preparations (Hydrastar and Phenomen A from Christian Dior).
Table 5.
Preparations based on purified ecdysteroids or on ecdysteroid-containing plant powders/extracts.

The impressive development of preparations containing ecdysteroids suggests that this class of molecule has indeed at least some of the claimed effects. The scientific justification for such commercial developments relies, however, on just a few references (ca. 10), often with the same ones being cited to support quite different effects.
Conclusion
Ecdysteroids are probably the most abundant steroids in nature because they are produced not only by arthropods, but also by many plant species. They seem to display a wide array of pharmacological effects on vertebrates, many of which are beneficial. However, these claims require more thorough validation and clinical testing. Ecdysteroids are used by an increasing number of humans as anabolic compounds, and it may well be that in the near future they will also be used on domesticated animals. This is the reason why new methods of detection and quantification have been recently proposed (Tsitsimpikou, 2001; Le Bizec, 2002) and further developments in this area are required. Whether ecdysteroid use will become controlled (e.g. for high-performance sportsmen or domestic animals [e.g. race horses]) is still open.
Ecdysteroids have also been successfully developed as effective inducers for gene switch control systems, several of which are presently in use. Ecdysteroids and/or bisacylhydrazines fulfil many of the required criteria, but not all. There are still problems which need to be overcome (e.g. the need for highly potent ligands for modified ecdysteroid receptors in transformed mammalian or plant cells). However, there is clearly great potential in this area. The future of ecdysteroid-regulated gene switches as an experimental tool is assured, but the prospects as in vivo systems is more debatable; the numerous pharmacological effects of ecdysteroids may preclude the development of their use in humans for gene therapy systems. This can only be resolved if more effort is invested into examining the biochemical fate and pharmacological consequences of ecdysteroids in mammals, especially humans.
Table 1.
Continued from previous page

Table 5.
Continued from previous page

Table 5.
Continued from previous page

Table 5.
Continued from previous page

Acknowledgments
The authors wish to thank Dr. Juraj Harmatha (Prague, Czech Republic) and Dr. Maria Báthori (Szeged, Hungary) for their help in collecting the data of Table 5.
References
- Abeysinghe RD, Greene BT, Haynes R, Willingham MC, Turner JL, Planalp RP, Brechbeil MW, Torti FM, Torti SV. p53-Independent apoptosis mediated by tachpyridine, an anti-cancer iron chelator. Carcinogenesis. 2001;22:1607–1614. doi: 10.1093/carcin/22.10.1607. [DOI] [PubMed] [Google Scholar]
- Abubakirov NK, Sultanov MB, Syrov VN, Kurmukov AG, Baltaev U, Novosel'skaya IL, Mamatkhanov AV, Gorovits MB, Shakirov TT, Shamsutdinov I, Yakubova MR, and Genkinoy GL. 1988 Tonic preparation containing the phytoecdysteroid (ecdystene). Application SU 1312774 (Chemical Abstracts 110:121377). [Google Scholar]
- Ahmad VU, Khaliq-Uz-Zaman SM, Ali MS, Perveen S, and Ahmed W. 1996 An antimicrobial ecdysone from Asparagus dumosus. Fitoterapia. LXVII. 1:88–91. [Google Scholar]
- Aikake A, Matsumoto T, and Yamaguchi Y. 1996 Cerebral neuron protective agents containing ecdysteroids. Application JP 94-195279/19940819 (Chemical Abstracts 125:1395). [Google Scholar]
- Aizikov MI, Kurmukov AG, and Syrov VN. 1978 Physiological activity and correlative changes in protein, carbohydrate, and fat metabolism under the effect of ecdysone and nerobol. Farmakologiya Prirodnykh Veschestv. 107–125.(Chemical Abstracts 90 : 180683). [Google Scholar]
- Albanese C, Reutens AT, Bouzahzah B, Fu M, D'Amico M, Link T, Nicholson R, Depinho RA, Pestell RG. Sustained mammary gland-directed, ponasterone-A inducible expression in transgenic mice. FASEB Journal. 2000;14:877–844. doi: 10.1096/fasebj.14.7.877. [DOI] [PubMed] [Google Scholar]
- Albertsen MC, Brooke CD, Garnaat CW, and Roth BA. 2000 Ecdysone receptors and methods for their use. International Patent Application WO 00/15791. [Google Scholar]
- Azizov AP, Seifulla RD. The effect of elton, leveton, fitoton and adapton on the work capacity of experimental animals. Eksperimental'naya i Klinicheskaya Farmakologiya. 1998;61:61–63. [PubMed] [Google Scholar]
- Baba M, Hirai S, Kawakami S, Kishida T, Sakai N, Kaneko S, Yao M, Shuin T, Kubota Y, Hosaka M, Ohno S. Tumor suppressor protein VHL is induced at high cell density and mediates contact inhibition of cell growth. Oncogene. 2001;20:2727–2736. doi: 10.1038/sj.onc.1204397. [DOI] [PubMed] [Google Scholar]
- Badal'yants KL, Nabiev AN, Khushbaktova ZA, Syrov VN. Mechanism of hepatoprotective action of ecdystene in acute heliotrine intoxication. Doklady Akademii Nauk Respubliki Uzbekistana. 1996;(10):46–48. [Google Scholar]
- Barsony J, Marx SJ. Receptor-mediated rapid action of 1α,25-dihydroxy-cholecalciferol: increase in intracellular cGMP in human skin fibroblasts. Proceedings of the National Academy of Sciences of the United States of America. 1988;85:1223–1226. doi: 10.1073/pnas.85.4.1223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Báthori M. Phytoecdysteroids effects on mammalians, isolation and analysis. Mini Reviews in Medicinal Chemistry. 2002;2:285–293. doi: 10.2174/1389557023406269. [DOI] [PubMed] [Google Scholar]
- Bathory M, Toth I, Szendrei K, Reisch J. Ecdysteroids in Spinacia oleracea and Chenopodium bonus-henricus. Phytochemistry. 1982;21:236–238. [Google Scholar]
- Bidmon HJ, Sliter TJ. The ecdysteroid receptor. Invertebrate Reproduction and Development. 1990;18:13–27. [Google Scholar]
- Bohl D, Heard JM. Transcriptional modulation of foreign gene expression in engineered somatic tissues. Cell Biology and Toxicology. 1998;14:83–94. doi: 10.1023/a:1007480419685. [DOI] [PubMed] [Google Scholar]
- Brann DW, Hendry LB, Mahesh VB. Emerging diversities in the mechanism of action of steroid hormones. Journal of Steroid Biochemistry and Molecular Biology. 1995;52:113–133. doi: 10.1016/0960-0760(94)00160-n. [DOI] [PubMed] [Google Scholar]
- Brazil DP, Hemmings BA. Ten years of protein kinase B signalling: a hard Akt to follow. Trends in Biochemical Sciences. 2001;26:657–664. doi: 10.1016/s0968-0004(01)01958-2. [DOI] [PubMed] [Google Scholar]
- Burdette WJ. Effect of invertebrate hormones on vertebrate tissues. Science. 1962;139:987. [Google Scholar]
- Burdette WJ. Hormonal heterophylly, invertebrate endocrinology, and phyto-hormones. Cancer Research. 1972;32:1088–1090. [PubMed] [Google Scholar]
- Burdette WJ. 1974 Invertebrate hormones and tumors. In: Burdette, WJ, editor. Invertebrate Endocrinology and Hormonal Heterophylly. 351–367.Springer-Verlag, Berlin. [Google Scholar]
- Burdette WJ, Coda RL. Effect of ecdysone on the incorporation of 14C-Leucine into hepatic protein in vitro. Proceedings of the Society for Experimental Biology and Medicine. 1963;112:216–217. doi: 10.3181/00379727-112-27996. [DOI] [PubMed] [Google Scholar]
- Canonica L, Danieli B, Weisz-Vincze I, Ferrari G. Structure of muristerone A, a new phytoecdysone. Journal of the Chemical Society, Chemical Communications. 1972:1060–1061. [Google Scholar]
- Carlson GR. 2000 The past, present, and future of EcR and ecdysteroid research in the agrobusiness. Presentation at the 14th Ecdysone Workshop, Rapperswil, Switzerland. [Google Scholar]
- Carlson GR, Cress DE, Dhadialla TS, Hormann RE, and Le DP. 2001 Ligands for modulating the expression of exogenous genes via and ecdysone receptor complex. U.S. Patent No. 6,258,603. [Google Scholar]
- Catalán RE, Aragones MD, Godoy JE, Martinez AM. Ecdysterone induces acetylcholinesterase in mammalian brain. Comparative Biochemistry and Physiology. 1984;78c:193–195. doi: 10.1016/0742-8413(84)90068-9. [DOI] [PubMed] [Google Scholar]
- Catalán RE, Aragones MD, Martinez AM. Effect of ecdysterone on cyclic AMP and cyclic GMP in mouse plasma. Biochemical and Biophysical Research Communications. 1979a;87:1018–1023. doi: 10.1016/s0006-291x(79)80009-1. [DOI] [PubMed] [Google Scholar]
- Catalán RE, Aragones MD, Martinez AM. Effect of ecdysterone on the cyclic AMP-protein kinase system in mouse liver. Biochemical and Biophysical Research Communications. 1979b;89:44–49. doi: 10.1016/0006-291x(79)90940-9. [DOI] [PubMed] [Google Scholar]
- Catalán RE, Martinez AM, Aragones MD. Heterophylic action of ecdysterone. Die Naturwissenschaften. 1980;67:520. doi: 10.1007/BF01047642. [DOI] [PubMed] [Google Scholar]
- Catalán RE, Martinez AM, Aragones MD. In vitro effect of ecdysterone on protein kinase activity. Comparative Biochemistry and Physiology. 1982;71B:301–303. [Google Scholar]
- Catalán RE, Martinez AM, Aragones MD, Miguel BG, Robles A, Godoy JE. Alterations in rat lipid metabolism following ecdysterone treatment. Comparative Biochemistry and Physiology. 1985;81B:771–775. doi: 10.1016/0305-0491(85)90403-1. [DOI] [PubMed] [Google Scholar]
- Chabanny VN, Levitsky EL, Gubsky YuI, Kholodova YuD. Gene-protective effect of the preparation based on ecdysteroids under rat poisoning with tetrachloromethane and chlorphos. Ukrainskii Biokhimicheskii Zhurnal. 1994;66(5):67–77. [PubMed] [Google Scholar]
- Chandrakala MV, Maribashetty VG, Jyothi HK. Application of phytoecdysteroids in sericulture. Current Science (India) 1998;74:341–346. [Google Scholar]
- Chaudhary KD, Lupien PJ, Hinse C. Effect of ecdysone on glutamic decarboxylase in rat brain. Experientia. 1969;25:250–251. doi: 10.1007/BF02034373. [DOI] [PubMed] [Google Scholar]
- Chen B, Athanasiou M, Gu Q, Blair DG. Drm/Gremlin transcriptionally activates p21Cip1 via a novel mechanism and inhibits neoplastic transformation. Biochemical and Biophysical Research Communications. 2002;295:1135–1141. doi: 10.1016/s0006-291x(02)00828-8. [DOI] [PubMed] [Google Scholar]
- Chen F, Zhang F, Rao J, Studzinski GP. Ectopic expression of truncated Sp1 transcription factor prolongs the S phase and reduces the growth rate. Anticancer Research. 2000;20(2A):661–667. [PubMed] [Google Scholar]
- Chermnykh NS, Shimanovsky NL, Shutko GV, Syrov VN. Effects of methandrostenolone and ecdysterone on physical endurance of animals and protein metabolism in the skeletal muscles. Farmakologiya i Toksikologiya. 1988;6:57–62. [PubMed] [Google Scholar]
- Chiang HC, Wang JJ, Wu RT. Immunomodulating effects of the hydrolysis products of formosamin C and β-ecdysone from Paris formosana Hayata. Anticancer Research. 1992;12:1475–1478. [PubMed] [Google Scholar]
- Cho WL, Kapitskaya MZ, Rhaikel AS. Mosquito ecdysteroid receptor: analysis of the cDNA and expression during vitellogenesis. Insect Biochemistry and Molecular Biology. 1995;25:19–27. doi: 10.1016/0965-1748(94)00045-j. [DOI] [PubMed] [Google Scholar]
- Christopherson KS, Mark MR, Bajaj V, Godowski PJ. Ecdysteroid-dependent regulation of genes in mammalian cells by a Drosophila ecdysone receptor and chimeric transactivators. Proceedings of the National Academy of Sciences, U.S.A. 1992;89:6314–6318. doi: 10.1073/pnas.89.14.6314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cole S.L, Schindler M, Sellers L.A, Humphrey P.P.A. Titrating the expression of a Gi protein-coupled receptor using an ecdysone-inducible system in CHO-K! cells. Receptors and Channels. 2001;7:289–302. [PubMed] [Google Scholar]
- Constantino S, Santo R, Gisselbrecht S, Gouilleux F. The ecdysteroid inducible gene expression system : unexpected effects of muristerone A and ponasterone A on cytokine signalling in mammalian cells. European Cytokine Network. 2001;12:365–367. [PubMed] [Google Scholar]
- Coulthard SA, Hogarth LA, Little M, Matheson EC, Redfern CPF, Minto L, Hall AG. The effect of thiopurine methyltransferase expression on sensitivity to thiopurine drugs. Molecular Pharmacology. 2002;62:102–109. doi: 10.1124/mol.62.1.102. [DOI] [PubMed] [Google Scholar]
- Darmograi VN, Potekhinskii SM, Ukhov YuI, Petrov VK, Potekhinskii SS, and Darmograi SV. 1998 Vitaderm containing phytoecdysteroids for treatment of burns and wounds. Application RU 96-96104062 / 19960229 (Chem. Abstr. 133:110028). [Google Scholar]
- Davis EM, Musch MW, Goldstein L. Transfection of an inducible trout anion exchanger (AE1) into HEK-EcR cells. Journal of Experimental Zoology. 2002;293:46–57. doi: 10.1002/jez.10111. [DOI] [PubMed] [Google Scholar]
- Dela Cruz F, Mak P. Drosophila ecdysone receptor functions as a constitutive activator in yeast. Journal of Steroid Biochemistry and Molecular Biology. 1997;62:353–359. doi: 10.1016/s0960-0760(97)00046-0. [DOI] [PubMed] [Google Scholar]
- DeMayo FJ, Tsai SY. Targeted gene regulation and gene ablation. Trends in Endocrinology & Metabolism. 2001;12:348–353. doi: 10.1016/s1043-2760(01)00462-3. [DOI] [PubMed] [Google Scholar]
- Descoins C Jr, Marion-Poll F. Electrophysiological responses of gustatory sensilla of Mamestra brassicae (Lepidoptera, Noctuidae) larvae to three ecdysteroids:ecdysone, 20-hydroxyecdysone and ponasterone A. Journal of Insect Physiology. 1999;45:871–876. doi: 10.1016/s0022-1910(99)00057-8. [DOI] [PubMed] [Google Scholar]
- Detmar M, Dumas M, Bonté F, Meybeck A, Orfanos CE. Effects of ecdysterone on the differentiation of normal human keratinocytes in vitro. European Journal of Dermatology. 1994;4:558–562. [Google Scholar]
- Dhadialla TS, Carlson GR, Le DP. New insecticides with ecdysteroidal and juvenile hormone activity. Annual Review of Entomology. 1998;43:545–569. doi: 10.1146/annurev.ento.43.1.545. [DOI] [PubMed] [Google Scholar]
- Dinan L. Phytoecdysteroids: biological aspects. Phytochemistry. 2001;57:325–339. doi: 10.1016/s0031-9422(01)00078-4. [DOI] [PubMed] [Google Scholar]
- Dinan L, Hormann RE, Fujimoto T. An extensive ecdysteroid CoMFA. Journal of Computer-aided Molecular Design. 1999a;13:185–207. doi: 10.1023/a:1008052320014. [DOI] [PubMed] [Google Scholar]
- Dinan L, Savchenko T, Whiting P, Sarker SD. Plant natural products as insect steroid receptor agonists and antagonists. Pesticide Science. 1999b;55:331–335. [Google Scholar]
- Durica DS, Wu X, Anilkumar G, Hopkins PM, Chung ACK. Characterization of crab EcR and RXR homologs and expression during limb regeneration and oocyte maturation. Molecular and Cellular Endocrinology. 2002;189:59–76. doi: 10.1016/s0303-7207(01)00740-7. [DOI] [PubMed] [Google Scholar]
- Dzukharova MKh, Sakhibov AD, Kasymov B, Syrov VN, Takanaev AA, Saatov Z. Pharmacokinetic experiments with ecdysterone. Khimiko-Farmatsevticheskii Zhurnal. 1987;21:1163–1167. [Google Scholar]
- El-Mofty M, Sadek I, Soliman A, Mohamed A, Sakre S. α-Ecdysone : a new bracken fern factor responsible for neoplasm induction in the Egyptian toad (Bufo regularis) Nutrition and Cancer. 1987;9:103–107. doi: 10.1080/01635588709513917. [DOI] [PubMed] [Google Scholar]
- El-Mofty MM, Sakre SA, Rizk AM, Moussa EA. Induction of breast and lung neoplastic lesions in mice by alpha-ecdysone. Oncology Reports. 1994;1:435–438. doi: 10.3892/or.1.2.435. [DOI] [PubMed] [Google Scholar]
- Evans R. 2002 Nuclear receptors: lipid physiology and the exotics of xenobiotics. Communication presented at the XVth International Ecdysone Workshop, Kolymbari, Crete, Greece (June 30 – July 6, 2002). [Google Scholar]
- Fomovska GN, Berdyshev AG, Kholodova YuD. Immunomodulatory effect of ecdysteroids. Ukrainskii Biokhimicheskii Zhurnal. 1992;64(2):56–61. [PubMed] [Google Scholar]
- Fujiwara H, Jindra M, Newitt R, Palli SR, Hiruma K, Riddiford LM. Cloning of an ecdysone receptor homolog from Manduca sexta and the developmental profile of its mRNA in wings. Insect Biochemistry and Molecular Biology. 1995;25:845–856. doi: 10.1016/0965-1748(95)00023-o. [DOI] [PubMed] [Google Scholar]
- Fussenegger M. The impact of mammalian gene regulation concepts on functional genomic research, metabolic engineering, and advanced gene therapies. Biotechnology Progress. 2001;17:1–51. doi: 10.1021/bp000129c. [DOI] [PubMed] [Google Scholar]
- Gao Z, Wang D, Li F. Determination of ecdysterone in Achyranthes bidentata BL. and its activity promoting proliferation of osteoblast-like cells. Acta Pharmaceutica Sinica. 2000;35:868–870. [PubMed] [Google Scholar]
- Gatz C, Lenk I. Promoters that respond to chemical inducers. Trends in Plant Science. 1998;3:352–358. [Google Scholar]
- Gill PK, Gescher A, Gant TW. Regulation of MDR1 promoter activity in human breast carcinoma cells by protein kinase C isozymes alpha and theta. European Journal of Biochemistry. 2001;268:4151–4157. doi: 10.1046/j.1432-1327.2001.02326.x. [DOI] [PubMed] [Google Scholar]
- Girault JP, Lafont R, Kerb U. Ecdysone catabolism in the white mouse. Drug Metabolism and Disposition. 1988;16:716–720. [PubMed] [Google Scholar]
- Gizatullina ZZ, Gagelgans AI, Syrov VN. Effects of ecdysterone, turkesterone and nerobol on thymocyte energy metabolism. Doklady Akademii Nauk Uzbeckoy SSR. 1994;(10):49–52. [Google Scholar]
- Graham LD. Ecdysone-controlled expression of transgenes. Expert Opinion on Biological Therapy. 2002;2:525–535. doi: 10.1517/14712598.2.5.525. [DOI] [PubMed] [Google Scholar]
- Grebenok RJ, Ripa PV, Adler JH. Occurrence and levels of ecdysteroids in spinach. Lipids. 1991;26:666–668. [Google Scholar]
- Gubskii YuI, Levitskii EL, Kholodova YuD, Goriushko AG, Primak RG, Vistunova IE, Sachenko LG. Mechanisms of genoprotective action of a phytoecdysteroid drug (BTK-8L) in chromatin damage by tetrachloromethane. Ukrainskii Biokhimicheskii Zhurnal. 1993;65(6):75–83. [PubMed] [Google Scholar]
- Guo F. 1989 Ecdysteroids in vertebrates : pharmalogical aspects. In Koolman J, editor. Ecdysone - from chemistry to mode of action. 442–446.Georg Thieme Verlag, Stuttgart. [Google Scholar]
- Hanaya R, Sasa M, Ishihara K, Akimitsu T, Iida K, Amano T, Serikawa T, Arita K, Kurisu K. Antiepileptic effects of 20-hydroxyecdysone on convulsive seizures in spontaneously epileptic rats. Japanese Journal of Pharmacology. 1997;74:331–335. doi: 10.1254/jjp.74.331. [DOI] [PubMed] [Google Scholar]
- Hannan GN, Hill RJ. Cloning and characterization of LcEcR: a functional ecdysone receptor from the sheep blowfly Lucilia cuprina. Insect Biochemistry and Molecular Biology. 1997;27:881–897. doi: 10.1016/s0965-1748(97)00019-2. [DOI] [PubMed] [Google Scholar]
- Hannan GN, Hill RJ. Lcusp, an ultraspiracle gene from the sheep blowfly, Lucilia cuprina: cDNA cloning, developmental expression of RNA and confirmation of function. Insect Biochemistry and Molecular Biology. 2001;31:771–781. doi: 10.1016/s0965-1748(00)00182-x. [DOI] [PubMed] [Google Scholar]
- Hashimoto Y, Ito Y, Arakawa E, Kita Y, Terashita K, Niikura T, Nishimoto I. Neurotoxic mechanism triggered by Alzheimer's disease-linked mutant M146L presenilin: involvement of NO synthase via a novel pertussis toxin target. Journal of Neurochemistry. 2002;80:426–437. doi: 10.1046/j.0022-3042.2001.00722.x. [DOI] [PubMed] [Google Scholar]
- Hayward DC, Bastiani MJ, Trueman MJ, Truman JW, Riddiford LM, Ball EE. The sequence of Locusta RXR, homologous to Drosophila Ultraspiracle and its evolutionary implications. Developmental Genes and Evolution. 1999;209:564–571. doi: 10.1007/s004270050290. [DOI] [PubMed] [Google Scholar]
- Heinrich G. 1970 Isolierung, Struktur und Stoffwechsel von Insekten-Häutungshormonen. Dissertation, Mathemathische Naturwissenschaftliche Fakultät, University of Hamburg, Germany. [Google Scholar]
- Hennigan RF, Stambrook PJ. Dominant negative c-jun activation of the cyclin D1 and cyclin E kinase complexes. Molecular Biology of the Cell. 2001;12:2352–2363. doi: 10.1091/mbc.12.8.2352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hikino H, Ohizumi Y, Takemoto T. Absorption, distribution, metabolism and excretion of insect-metamorphosing hormone ecdysterone in mice. I. Yakugaku Zasshi. 1972a;92:945–950. doi: 10.1248/yakushi1947.92.8_945. [DOI] [PubMed] [Google Scholar]
- Hikino H, Ohizumi Y, Takemoto T. Absorption, distribution, metabolism and excretion of insect-metamorphosing hormone ecdysterone in mice. II. Chemical Pharmaceutical Bulletin. 1972b;20:2454–2458. doi: 10.1248/cpb.20.2454. [DOI] [PubMed] [Google Scholar]
- Hikino S, Nabetani S, Nomoto K, Arai T, Takemoto T, Otaka T, Uchiyama M. Effect of long-term administration of insect metamorphosing substances on higher animals. I. Yakugaku Zasshi. 1969;89:235–240. doi: 10.1248/yakushi1947.89.2_235. [DOI] [PubMed] [Google Scholar]
- Hirono I, Sasaoka I, Shimizu M. Effect of insect-molting hormones, ecdysterone and inokosterone, on tumor cells. Gann. 1969;60:341–342. [PubMed] [Google Scholar]
- Hoppe UC, Marban E, Johns DC. Adenovirus-mediated inducible gene expression in vivo by a hybrid ecdysone receptor. Molecular Therapy. 2000;1:159–164. doi: 10.1006/mthe.1999.0023. [DOI] [PubMed] [Google Scholar]
- Imhof MO, Rusconi S, Lezzi M. Cloning of a Chironomus tentans cDNA encoding protein (cEcRH) homologous to the Drosophila melanogaster ecdysteroid receptor (dEcR) Insect Biochemistry and Molecular Biology. 1993;23:115–124. doi: 10.1016/0965-1748(93)90089-b. [DOI] [PubMed] [Google Scholar]
- Inaoka Y, Yamamoto M, and Tsuji K. 1997 Psoriasis inhibitors containing ecdysteroid analogs. Jpn. Kokai Tokkyo Koho JP 09 02,955 [97 02,955]. (Chemical Abstracts 126:166507r). [Google Scholar]
- Iwata A, Maruyama M, Kanazawa I, Nukina N. α-Synuclein affects the MAPK pathway and accelerates cell death. Journal of Biological Chemistry. 2001;276:45320–45329. doi: 10.1074/jbc.M103736200. [DOI] [PubMed] [Google Scholar]
- Jana NR, Zemskov EA, Wang GH, Nukina N. Altered proteasomal function due to the expression of polyglutamine-expanded truncated N-terminal huntingtin induces apoptosis by caspase activation through mitochondrial cytochrome c release. Human Molecular Genetics. 2001;10:1049–1059. doi: 10.1093/hmg/10.10.1049. [DOI] [PubMed] [Google Scholar]
- Janowski BA, Grogan MF, Jones SA, Wisely GB, Kliewer SA, Corey EJ, Mangelsdorf DJ. Structural requirements of ligands for the oxysterol liver X receptors LXRalpha and LXRbeta. Proceedings of the National Academy of Sciences USA. 1999;69:266–271. doi: 10.1073/pnas.96.1.266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jepson I, Martinez A, Sweetman JP. Chemical-inducible gene expression systems for plants - a review. Pesticide Science. 1998;54:360–367. [Google Scholar]
- Jindra M, Huang JY, Malone F, Asahina M, Riddiford LM. Identification and mRNA developmental profiles of two ultraspiracle isoforms in the epidermis and wings of Manduca sexta. Insect Molecular Biology. 1997;6:41–53. doi: 10.1046/j.1365-2583.1997.00153.x. [DOI] [PubMed] [Google Scholar]
- Jones G, Jones D. Considerations on the structural evidence of a ligand-binding function of ultraspiracle, an insect homolog of vertebrate RXR. Insect Biochemistry and Molecular Biology. 2000;30:671–679. doi: 10.1016/s0965-1748(00)00038-2. [DOI] [PubMed] [Google Scholar]
- Jones SA, Moore LB, Shenk JL, Wisely GB, Hamilton GA, McKee DD, Tomlinson ACO, LeCluyse EL, Lamberft MH, Wilson TM, Kliewer SA, Moore JT. The pregnane X receptor : a promiscuous xenobiotic receptor that has diverged during evolution. Molecular Endocrinology. 2000;14:27–39. doi: 10.1210/mend.14.1.0409. [DOI] [PubMed] [Google Scholar]
- Kapitskaya M, Wang S, Cress DE, Dhadialla TS, Rhaikel AS. The mosquito ultraspiracle homologue, a partner of ecdysteroid receptor heterodimer: cloning and characterization of isoforms expressed during vitellogenesis. Molecular and Cellular Endocrinology. 1996;12:119–132. doi: 10.1016/0303-7207(96)03847-6. [DOI] [PubMed] [Google Scholar]
- Karns LR, Kisielewski A, Gulding KM, Seraj JM, and Theodorescu D. 2001 Manipulation of gene expression by an ecdysone-inducible gene switch in tumor xenografts. BioMed Central Biotechnology. 1(article 11):pp. 12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khalitova YuD, Syrov VN. Possible use of the extract of Ajuga turkestanica as a remedy contributing to lactation. Doklady Akademii Nauk Respubliki Uzbekistana. 1998;(8):35–38. [Google Scholar]
- Khimiko IN, Mitrokin YuI, Efremova OI, Sidorenko LI. The influence of ecdysterone on the biosynthesis of proteins and nucleic acids in mouse organs. Khimiko-Farmatsevticheskii Zhurnal. 2000;34(9):3–5. [Google Scholar]
- Kholodova YuD. Phytoecdysteroids : biological effects, application in agriculture and complementary medicine. Ukrainskii Biokhimicheskii Zhurnal. 2001;73:21–29. [PubMed] [Google Scholar]
- Kholodova YuD, Tuga VA, Zimina VP. Effects of vitamin D3 and 20-hydroxyecdysone on the content of ATP, creatine phosphate, carnosine and Ca2+ in skeletal muscles. Ukrainskii Biokhimicheskii Zhurnal. 1997;69:3–9. [PubMed] [Google Scholar]
- Khushbatkova ZA, Azizova SS, Syrov VN, Umarova FT, Mirsalikhova NT. The effect of celanide and ecdysterone on the activity of sodium-potassium ATP-ase in the myocardium. Medicinal'nii Zhurnal Uzbekistana. 1987;(5):68–70. [Google Scholar]
- Koelle MR, Talbot WS, Segraves WA, Bender MT, Cherbas P, Hogness DS. The Drosophila EcR gene encodes an ecdysone receptor, a new member of the steroid receptor superfamily. Cell. 1991;67:59–77. doi: 10.1016/0092-8674(91)90572-g. [DOI] [PubMed] [Google Scholar]
- Kondo M, Ji L, Kamibayashi C, Tomizawa Y, Randle D, Sekido Y, Yokota J, Kashuba V, Zabarovsky E, Kuzmin I, Lertman M, Roth J, Minna JD. Overexpression of candidate tumor suppressor gene FUS1 isolated from the 3p21.3 homozygous deletion region leads to G1 arrest and growth inhibition of lung cancer cells. Oncogene. 2001;20:6258–6262. doi: 10.1038/sj.onc.1204832. [DOI] [PubMed] [Google Scholar]
- Konovalova NP, Mitrokhin YI, Volkova LM, Sidorenko LI, Todorov IN. Ecdysterone modulates antitumor activity of cytostatics and biosynthesis of macromolecules in tumor-bearing mice. Biology Bulletin. 2002;29:530–536. [PubMed] [Google Scholar]
- Kosar K, Opletal L, Vokac K, Harmatha J, Sovova M, Cerovsky J, Kratky F, Dvorak J. Embryotoxicity of 20-hydroxyecdysone and polypodine B from Leuzea carthamoides DC. Pharmazie. 1997;52:406–407. [PubMed] [Google Scholar]
- Kosovsky MI, Syrov VN, Mirakhmedov MM, Katkova SP, Khushbatkova ZA. The effect of nerobol and ecdysterone on processes related to insulin regulatory function in normal and in experimental insulin resistance. Problemy Endokrinology. 1989;35:77–81. [PubMed] [Google Scholar]
- Kothapalli R, Palli SR, Ladd TR, Sohi SS, Cress D, Dhadialla TS, Tzertzinis G, Rethakaran A. Cloning and developmental expression of the ecdysone receptor gene from the spruce budworm Choristoneura fumiferana. Developmental Genetics. 1995;17:319–330. doi: 10.1002/dvg.1020170405. [DOI] [PubMed] [Google Scholar]
- Kotsyuruba AV, Bukhanevich OM, Tuganova AV, Tarakanov SS, Berdishev AG. Mechanisms of early action of biologically active oxysterines - calcitriol and ecdysterone. Modulation of systems which generate low-molecular activators of guanylate cyclase. Ukrainskii Biokhimicheskii Zhurnal. 1995a;67:58–64. [PubMed] [Google Scholar]
- Kotsyuruba AV, Bukhanevich OM, Tuganova AV, Tarakanov SS. Mechanisms of early effect of biologically active oxysterones calcitriol and ecdysterone, modulation of intracellular pools of arachidonic acid and products of its oxidative metabolism. Ukrainskii Biokhimicheskii Zhurnal. 1995b;67:45–52. [PubMed] [Google Scholar]
- Kotsyuruba AV, Tuganova AV, Bukhanevich OM, Tarakanov SS. Mechanisms of early action of biologically active oxysterines - calcitriol and ecdysterone. Identification of sphingomyelin metabolism as the effector mechanism of early action. Ukrainskii Biokhimicheskii Zhurnal. 1995c;67:53–58. [PubMed] [Google Scholar]
- Kotsyuruba AV, Bukhanevich OM, Tarakanov SS, Tuganova AV, Berdyshev AG, Meged OF. C27-Steroid hormones calcitriol and ecdysterone activates hydrolysis of neutral lipids) cholesterol esters and triacylglycerols – in its early pregenomic phase of action. Ukrainskii Biokhimicheskii Zhurnal. 1998a;70(5):30–36. [PubMed] [Google Scholar]
- Kotsyuruba AV, Bukhanevich OM, Tarakanov SS, Tuganova AV, Berdyshev AG, Meged OF. C27-Steroid hormones calcitriol and ecdysterone in its early phase of action activates in target animals tissues the phosphatidylcholine hydrolysis. Ukrainskii Biokhimicheskii Zhurnal. 1998b;70(5):30–36. [PubMed] [Google Scholar]
- Kotsyuruba AV, Bukhanevich OM, Meged OF, Tarakanov SS, Berdyshev AG, Tuganova AV. C27-Steroid hormones ecdysterone and calcitriol activates phosphoinositide messenger cascade in its early membrane phase of action. Ukrainskii Biokhimicheskii Zhurnal. 1999;71(1):27–32. [Google Scholar]
- Koudela K, Tenora J, Bajer J, Mathova A, Sláma K. Stimulation of growth and development in Japanese quails after oral administration of ecdysteroid-containing diet. European Journal of Entomology. 1995;92:349–354. [Google Scholar]
- Kratky F, Opletal L, Hejhalek J, Kucharova S. Effect of 20-hydroxyecdysone on the protein synthesis of pigs. Zivocisna Vyroba. 1997;42:445–451. [Google Scholar]
- Kuate S, Wagner R, Überla K. Development and characterization of a minimal inducible packaging cell line for simian immunodeficincy virus-based lentiviral vectors. Journal of Gene Medicine. 2002;4:347–355. doi: 10.1002/jgm.290. [DOI] [PubMed] [Google Scholar]
- Kudo Y, Kitajima S, Sato S, Ogawa I, Miyauchi M, Takata T. Transfection of p27Kip1 Threonine residue 187 mutant type gene, which is not influenced by ubiquitin-mediated degradation, induces cell cycle arrest in oral sqamous cell carcinoma cells. Oncology. 2002;63:398–404. doi: 10.1159/000066222. [DOI] [PubMed] [Google Scholar]
- Kumar MB, Fujimoto T, Potter DW, Deng Q, and Palli SR. 2002 A single point mutation in ecdysone receptor leads to increased ligand specificity: implications for gene switch applications. Proceedings of the National Academy of Sciences, USA. (in press). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumar R, Thompson EB. The structure of the nuclear hormone receptors. Steroids. 1999;64:310–319. doi: 10.1016/s0039-128x(99)00014-8. [DOI] [PubMed] [Google Scholar]
- Kurmukov AG, Yermishina OA. Effect of ecdysterone on experimental arrhythmias, changes in hemodynamics and contractility of the myocardium produced by a coronary artery occlusion. Farmakologiya i Toksikologiya. 1991;54:27–29. [PubMed] [Google Scholar]
- Kurmukov AG, Syrov VN. Anti-inflammatory properties of ecdysterone. Medicinal'nii Zhurnal Uzbekistana. 1988;(10):68–70. [Google Scholar]
- Kutepova TA, Syrov VN, Khushbaktova ZA, Saatov Z. Hypoglycemic activity of the total extract from Ajuga turkestanica. Pharmaceutical Chemistry Journal. 2001;35:608–609. [Google Scholar]
- Kuzmenko AI, Morozova RP, Nikolenko IA, Koniets GV, Kholodova YuD. Effects of vitamin D3 and ecdysterone on free-radical lipid peroxidation. Biochemistry (Moscow) 1997;62(6):609–612. [PubMed] [Google Scholar]
- Kuzmenko AI, Niki E, Noguchi N. New functions of 20-hydroxyecdysone in lipid peroxidation. Journal of Oleo Science. 2001;50:497–506. [Google Scholar]
- Kuzmitsky BB, Golubeva MB, Konoplya NA, Kovganko NV, Achrem AA. New opportunities of search for immunomodulators among compounds with steroidal structure. Farmakologiya i Toksikologiya. 1990;53:20–22. [PubMed] [Google Scholar]
- Lafont R. Phytoecdysteroids in the World flora : diversity, distribution, biosynthesis and evolution. Russian Journal of Plant Physiology. 1998;45:276–295. [Google Scholar]
- Lafont R, Girault JP, Kerb U. Excretion and metabolism of injected ecdysone in the white mouse. Biochemical Pharmacology. 1988;37:1174–1177. doi: 10.1016/0006-2952(88)90529-1. [DOI] [PubMed] [Google Scholar]
- Lagova ND, Valueva IM. Effect of ecdysterone isolated from Rhaponticum carthamoides on the growth of experimental tumors. Eksperimental'naya Onkologiya. 1981;3(4):69–71. [Google Scholar]
- Laudet V. Evolution of the nuclear receptor superfamily: early diversification from an ancestral orphan receptor. Journal of Molecular Endocrinology. 1997;19:207–226. doi: 10.1677/jme.0.0190207. [DOI] [PubMed] [Google Scholar]
- Le Bizec B, Antignac JP, Monteau F, Andre F. Ecdysteroids : one potential new anabolic family in breeding animals. Analytica Chimica Acta. 2002;473:89–97. [Google Scholar]
- Levitskii EL, Gubskii YuI, Primak RG, Goriushko AG, Kholodova YuD, Vistunova IE, Marchenko AN. Chromatin-protective action of the biological preparation BTK-8L in tetrachloromethane and chlorphos. Ukrainskii Biokhimicheskii Zhurnal. 1996;68(5):76–84. [PubMed] [Google Scholar]
- Levitskii EL, Kholodova YuD, Gubskii YuI, Goriushko AG, Primak RG, Vistunova IE, Sachenko LG. Mechanism of the genoprotective action of a phytoecdysteroid drug (BTK-8L) in chromatin damage by chlorphos. Ukrainskii Biokhimicheskii Zhurnal. 1993a;65(6):84–91. [PubMed] [Google Scholar]
- Levitsky EL, Kholodova YuD, Gubski I, Primak RG, Chabanny VN, Kindruk NL, Mozzhukina TG, Lenchevskaya LK, Mironova VN, Saad LM, Vitsunova IE, Shabliy VI. Biochemical characteristics of rat liver fractionated chromatin under experimental D-hypovitaminosis and after treatment by steroidal preparations. Ukrainskii Biokhimicheskii Zhurnal. 1993b;65(1):28–36. [PubMed] [Google Scholar]
- Lin N, Lin W. 1989 β-Ecdysone containig skin-protecting cosmetics. Faming Zhuanli Shenqing Gonkkai Shuomingshu. CN 86,106,791 (Cl. A61K7/48), 13 Apr. 1988, Appl. 30 Sept 1986. 3. pp. (Chemical Abstracts 111: 239323e). [Google Scholar]
- Lin S, Yang Y, and Feng S. 1997 Effects of ecdysterone on proliferation of human umbilical vein endothelial cells. Zhongguo Yaolixue Tongbao. 13:176–179.(Chemical Abstracts 128 : 57260). [Google Scholar]
- Lüers GH, Jess N, Franz T. Reporter-linked monitoring of transgene expression in living cells using the ecdysone-inducible promoter system. European Journal of Cell Biology. 2000;79:653–657. doi: 10.1078/0171-9335-00086. [DOI] [PubMed] [Google Scholar]
- Lupien PJ, Hinse C, Chaudhary KD. Ecdysone as a hypocholesterolemic agent. Archives Internationales de Physiologie et de Biochimie. 1969;77:206–212. doi: 10.3109/13813456909109700. [DOI] [PubMed] [Google Scholar]
- Maimeskulova LA, Maslov LN. Anti-arrhythmic effect of phytoadaptogens. Eksperimental'naya i Klinicheskaya Farmakologiya. 2000;63:29–31. [PubMed] [Google Scholar]
- Martinez A, Sparks C, Hart CA, Thompson J, Jepson I. Ecdysone agonist inducible transcription in transgenic tobacco plants. Plant Journal. 1999a;19:97–106. doi: 10.1046/j.1365-313x.1999.00504.x. [DOI] [PubMed] [Google Scholar]
- Martinez A, Sparks C, Drayton P, Thompson J, Greenland A, Jepson I. Creation of ecdysone receptor chimeras in plants for controlled regulation of gene expression. Molecular and General Genetics. 1999b;261:546–552. doi: 10.1007/s004380050999. [DOI] [PubMed] [Google Scholar]
- Martinez A, Scanlon D, Gross B, Perara SC, Palli SR, Greenland AJ, Windass J, Pongs O, Broad P, Jepson I. Transcriptional activtion of the cloned Heliothis virescens (Lepidoptera) ecdysone receptor (HvEcR) by muristerone A. Insect Biochemistry and Molecular Biology. 1999c;29:915–930. doi: 10.1016/s0965-1748(99)00067-3. [DOI] [PubMed] [Google Scholar]
- Matsuda H, Kawaba T, Yamamoto Y. Pharmacological studies of insect metamorphosing steroids from Achyranthis radix. Nippon Yakubutsugaku Zasshi (Folia Pharmacologica Japonica) 1970;66:551–563. [PubMed] [Google Scholar]
- Matsuda H, Kawaba T, Yamamoto Y, Ogawa S. Effect of ecdysterone on experimental atherosclerosis in rabbit. Nippon Yakubutsugaku Zasshi (Folia Pharmacologica Japonica) 1974;70:325–339. doi: 10.1254/fpj.70.325. [DOI] [PubMed] [Google Scholar]
- Meents H, Enenkel B, Werner RG, Fussenegger M. p27Kip1-mediated controlled proliferation technology increases constiutive sICAM production in CHO-DUKX adapted for growth in suspension and serum-free media. Biotechnology and Bioengineering. 2002;79:619–627. doi: 10.1002/bit.10322. [DOI] [PubMed] [Google Scholar]
- Mellon I, Hock T, Reid R, Porter PC, States JC. Polymorphisms in the juman xeroderma pigmentosum group A gene and their impact on cell survival and nucleotide excision repair. DNA Repair. 2002;1:531–546. doi: 10.1016/s1568-7864(02)00053-8. [DOI] [PubMed] [Google Scholar]
- Meybeck A. 1999a Cosmetic emulsions containing hydrocarbons and phospholipids. Application FR 98-4542/19980410 (Chemical Abstracts 132:26660). [Google Scholar]
- Meybeck A. 1999b Cosmetic or dermatological water-in-oil emulsions containing liquid esters of a fatty acid or fatty alcohol and phospholipids. Application FR 98-4544/19980410 (Chemical Abstracts 132:26696). [Google Scholar]
- Meybeck A, Bonté F. 1990 Ecdysteroid-containing liposomes for wound healing and skin regeneration. Demande FR 2,637,182. (Chemical Abstracts 114:30138r). [Google Scholar]
- Meybeck A, Bonté F. 1993 Solid particles comprising a biologically active substance for cosmetics and agrochemicals. PCT Int. Appl. WO 93 12,761 (Chemical Abstracts 119:188275r). [Google Scholar]
- Meybeck A, Bonté F, and Redziniak G. 1994 Use of a cosmetic in dermatological composition of keratinocyte culture medium. PCT Int. Appl. WO 94 04,132 (Chemical Abstracts 120:253087y). [Google Scholar]
- Mironova VN, Kholodova YuD, Skatchkova TF, Bonda OP, Datsenko ZM, Govseeva NN. Hypocholesterolemic effects of phytoecdysones in rat experimental hypercholesterolemia. Voprosy Meditsinskoi Khimii. 1982;28(3):101–105. [PubMed] [Google Scholar]
- Mirzaev YuR, Syrov VN. Effect of phytoecdysteroids on the sexual activity of male rats. Doklady Akademii Nauk Respubliki Uzbekistana. 1992;(3):47–49. [Google Scholar]
- Mirzaev YuR, Syrov VN, Krushev SA, Iskanderova SD. Study of the effects of ecdysten on the sexual function under experimental and clinical conditions. Eksperimental'naya i Klinicheskaya Farmakologiya. 2000;63:35–37. [PubMed] [Google Scholar]
- Morgan WW, Richardson A, Sharp ZD, Walter CA. Application of exogenously regulatable promoter systems to transgenic models for the study of aging. Journal of Gerontology. 1999;54A:B30–B40. doi: 10.1093/gerona/54.1.b30. [DOI] [PubMed] [Google Scholar]
- Mouillet JF, Delbecque JP, Quennedy B, Delachambre J. Cloning of two putative ecdysteroid receptor isoforms from Tenebrio molitor and their developmental expression in the epidermis during metamorphosis. European Journal of Biochemistry. 1997;248:856–863. doi: 10.1111/j.1432-1033.1997.00856.x. [DOI] [PubMed] [Google Scholar]
- Mykhaylyk OM, Kotsuruba AV, Buchanevich OM, Gula NM, Bakai EA. Cell surface receptor interactions of C27-steroid hormone ecdysterone immobilized on nanodispersed magnetite. Journal of Magnetism and Magnetic Materials. 1999;194:113–119. [Google Scholar]
- Mykhaylyk OM, Kotsuruba AV, Buchanevich OM, Korduban AM, Mengel EF, Gulaya NM. Signal transduction of erythrocytes after specific binging of ecdysterone and cholesterol immobilized on nanodispersed magnetite. Journal of Magnetism and Magnetic Materials. 2001;225:226–234. [Google Scholar]
- Najmutdinova DK, Saatov Z. Lung local defense in experimental diabetes mellitus and the effect of 11,20-dihydroxyecdysone in combination with manilil. Archives of Insect Biochemistry and Physiology. 1999;41:144–147. doi: 10.1002/(SICI)1520-6327(1999)41:3<144::AID-ARCH5>3.0.CO;2-0. [DOI] [PubMed] [Google Scholar]
- Nicolai M, Bouhin H, Quennedey B, Delachambre J. Molecular cloning and expression of Tenebrio molitor ultraspiracle during metamorphosis and in vivo induction of its phosphorylation by 20-hydroxyecdysone. Insect Molecular Biology. 2000;9:241–249. doi: 10.1046/j.1365-2583.2000.00181.x. [DOI] [PubMed] [Google Scholar]
- Niikura T, Murayama N, Hashimoto Y, Ito Y, Yamagishi Y, Matsuoka M, Takeuchi Y, Aiso S, Nishimoto I. V642I APP-inducible neuronal cells: a model system for investigating Alzheimer's disorders. Biochemical and Biophysical Research Commununications. 2000;274:445–454. doi: 10.1006/bbrc.2000.3143. [DOI] [PubMed] [Google Scholar]
- No D, Yao TP, Evans RM. Ecdysone-inducible gene expression in mammalian cells and transgenic mice. Proceedings of the National Academy of Sciences, USA. 1996;93:3346–3351. doi: 10.1073/pnas.93.8.3346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Odero-Marah VA, Khalkali-Ellis Z, Schneider GB, Seftor EA, Seftor REB, Koland JG, Hendrix MJC. Tyrosine phosphorylation of maspin in normal mammary epithelia and breast cancer cells. Biochemical and Biophysical Research Communications. 2002;295:800–805. doi: 10.1016/s0006-291x(02)00764-7. [DOI] [PubMed] [Google Scholar]
- Ogawa S, Nishimoto N, and Matsuda H. 1974 Pharmacology of ecdysones in Vertebrates. In: Burdette, WJ, editor. Invertebrate Endocrinology and Hormonal Heterophylly. 341–344.Springer-Verlag, Berlin. [Google Scholar]
- Okada M, Ishihara K, Sasa M, Izumi R, Yajin K, Harada Y. Enhancement of GABA-mediated inhibition of rat medial vestibular nucleus neurons by the neurosteroid 20-hydroxyecdysone. Acta Otolaryngologia. 1998;118:11–16. doi: 10.1080/00016489850155062. [DOI] [PubMed] [Google Scholar]
- Okui S, Otaka T, Uchiyama M, Takemoto T, Hikino H, Ogawa S, Nishimoto N. Stimulation of protein synthesis in mouse liver by insect-moulting steroids. Chemical and Pharmaceutical Bulletin. 1968;16:384–387. doi: 10.1248/cpb.16.384. [DOI] [PubMed] [Google Scholar]
- Oro AE, McKeown M, Evans RM. Relationship between the product of the Drosophila ultraspiracle locus and the vertebrate retinoid X receptor. Nature. 1990;347:298–301. doi: 10.1038/347298a0. [DOI] [PubMed] [Google Scholar]
- Osynskaya LF, Saad LM, Kholodova YuD. Antiradical properties and antioxidative activity of ecdysterone. Ukrainskii Biokhimicheskii Zhurnal. 1992;64(1):114–117. [PubMed] [Google Scholar]
- Otaka T, Uchiyama M, Okui S, Takemoto T, Hikino H, Ogawa S, Nishimoto N. Stimulatory effect of insect metamorphosing steroids from Achyranthes and Cyathula on protein synthesis in mouse liver. Chemical and Pharmaceutical Bulletin. 1968;16:2426–2429. doi: 10.1248/cpb.16.2426. [DOI] [PubMed] [Google Scholar]
- Otaka T, Okui S, Uchiyama M. Stimulation of protein synthesis in mouse liver by ecdysterone. Chemical and Pharmaceutical Bulletin. 1969a;17:75–81. doi: 10.1248/cpb.17.75. [DOI] [PubMed] [Google Scholar]
- Otaka T, Uchiyama M, Takemoto T, Hikino H. Stimulatory effect of insect metamorphosing steroids from ferns on protein synthesis in mouse liver. Chemical and Pharmaceutical Bulletin. 1969b;17:1352–1355. doi: 10.1248/cpb.17.1352. [DOI] [PubMed] [Google Scholar]
- Padidam M. Chemically regulated gene expression in plants. Current Opinion in Plant Biology. 2003;6:1–9. doi: 10.1016/s1369-5266(03)00005-0. [DOI] [PubMed] [Google Scholar]
- Padidam M, Gore M, Lu DL, and Smirnova O. 2002 Chemical-inducible, ecdysone receptor-based gene expression system for plants. Transgenic Research. 11(in press). [DOI] [PubMed] [Google Scholar]
- Palli SR, Kapitskaya MZ, Kumar MB, and Cress DE. 2003 Improved ecdysone receptor based inducible gene regulation system. European Journal of Biochemistry. (in press). [DOI] [PubMed] [Google Scholar]
- Palmer MJ, Harmon MA, Laudet V. Characterization of EcR and RXR homologues in the Ixodid tick, Amblyomma americanum (L.) American Zoologist. 1999;39:747–757. [Google Scholar]
- Patrick CW, Zheng B, Wu X, Gurtner G, Barlow M, Koutz C, Chang D, Schmidt M, Evans GRD. Muristerone A-induced nerve growth factor release form genetically engineered human dermal fibroblasts for peripheral nerve tissue engineering. Tissue Engineering. 2001;7:303–311. doi: 10.1089/10763270152044161. [DOI] [PubMed] [Google Scholar]
- Perera SC, Ladd TR, Dhadialla TS, Krell PJ, Sohi SS, Retnakaran A, Palli SR. Studies on two ecdysone receptor isoforms of the spruce budworm, Choristoneura fumiferana. Molecular and Cellular Endocrinology. 1999;152:73–84. doi: 10.1016/s0303-7207(99)00058-1. [DOI] [PubMed] [Google Scholar]
- Plotnikov MB, Zibareva LN, Koltunov AA, Aliev OI, Yakimova TV, Maslov YuM. The hemorheologic properties of extracts from some ecdysteroids-containing plants. Rastitel'niye Resursy. 1998;34:91–97. [Google Scholar]
- Plows D, Briassouli P, Owen C, Zoumpourlis V, Garrett MD, Pintzas A. Ecdysone-inducible expression of oncogenic Ha-Ras in NIH 3T3 cells leads to transient nuclear localization of activated extracellular signal-regulated kinase regulated by mitogen-activated protein kinase phosphatase-1. Biochemical Journal. 2002;362:305–315. doi: 10.1042/0264-6021:3620305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Politova NK, Kovler LA, Volodin VV, Lushka VG, Pshunetleva EA. Chemical modification of 20-hydroxyecdysone and study of membranotropic properties of its derivatives. Khimiya Rastitel'nogo Syr'ya. 2001;(2):69–81. [Google Scholar]
- Prabhu VKK, Nayar KK. Crustecdysone is without estrogenic or antiestrogenic activity in the rat. Experientia. 1974;30:821. doi: 10.1007/BF01924207. [DOI] [PubMed] [Google Scholar]
- Purser DB, Baker SK. 1994 Ecdysones used to improve productivity of ruminants. PCT Int. Appl. WO 94 18,984, AU Appl. 93/7,397 (Chemical Abstracts 121:254587). [Google Scholar]
- Ramazanov N, Saatov Z, Syrov VN. Study of ecdysterone metabolites isolated from rat urine. Khimiia Prirodnykh Soedineniï. 1996;(4):558–564. [Google Scholar]
- Rampazzo C, Johansson M, Gallinaro L, Ferraro P, Hellman U, Karlsson A, Reichard P, Bianchi V. Mammalian 5′(3′)-deoxyribonucleotidase, cDNA cloning, and overexpression of the enzyme in Escherichia coli and mammalian cells. Journal of Biological Chemistry. 2000;275:5409–5415. doi: 10.1074/jbc.275.8.5409. [DOI] [PubMed] [Google Scholar]
- Ravi M, Hopfinger AJ, Hormann RE, Dinan L. 4D-QSAR of a set of ecdysteroids and a comparison to CoMFA modeling. Journal of Chemical Information and Computer Sciences. 2001;41:1587–1604. doi: 10.1021/ci010076u. [DOI] [PubMed] [Google Scholar]
- Rinehart JP, Cikra-Ireland R, Flannagan RD, Denlinger DL. Expression of the ecdysone receptor is unaffected by pupal diapause in the flesh fly, Sarcophaga crassipalpis, while its dimerization partner, USP, is downregulated. Journal of Insect Physiology. 2001;47:915–921. [Google Scholar]
- Saatov Z, Agzamhodjaeva DA, Syrov VN. The prevalence of phytoecdysteroids in plants of Uzbekistan and the possibility of using the preparations created and their basis in nephrology to practice. Khimiya Prirodnykh Soedinenii. 1999;(2):209–215. [Google Scholar]
- Saez E, Nelson MC, Eshelman B, Banayo E, Koder A, Cho GJ, Evans RM. Identification of ligands and coligands for the ecdysone-regulated gene switch. Proceedings of the National Academy of Sciences, USA. 2000;97:14512–14517. doi: 10.1073/pnas.260499497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saez E, No D, West A, Evans RM. Inducible gene expression in mammalian cells and transgenic mice. Current Opinion in Biotechnology. 1997;8:608–616. doi: 10.1016/s0958-1669(97)80037-7. [DOI] [PubMed] [Google Scholar]
- Sakhibov AD, Syrov VI, Usmanova AS, Abakumova OYu. Experimental analysis of the immunotropic action of phytoecdysteroids. Doklady Akademii Nauk Uzbeckoy SSR. 1989;(8):55–57. [Google Scholar]
- Saleh DS, Zhang J, Wyatt GR, Walker VK. Cloning and characterization of an ecdysone receptor cDNA from Locusta migratoria. Molecular and Cellular Endocrinology. 1998;143:91–99. doi: 10.1016/s0303-7207(98)00131-2. [DOI] [PubMed] [Google Scholar]
- Sasa M, Tsujiyama S, Ishihara K, Hanaya r, Fujita M, Kurisu K, Yajin K, and Serikawa T. 1996 Enhancement of GABA-induced current by 20-hydroxyecdysone in cultured cortical neurons. In GABA : Receptors, Transport and Metabolism (Tanaka C and Bowery NG Eds), pp. 185–194.Birkhaeuser, Basel, Ch. [Google Scholar]
- Sawicki JA, Monks B, Morris RJ. Cell-specific ecdysone-inducible expression of FLP recombinase in mammalian cells. BioTechniques. 1998;25:868–875. doi: 10.2144/98255rr01. [DOI] [PubMed] [Google Scholar]
- Schmidt M, Fan Z. Protection against chemotherapy-induced cytotoxicity by cyclin-dependent kinase inhibitors (CKI) in CKI-responsive cells compared with CKI-unresponsive cells. Oncogene. 2001;20:6164–6171. doi: 10.1038/sj.onc.1204814. [DOI] [PubMed] [Google Scholar]
- Schneider S, Wünsch S, Schwab A, Oberleithner H. Rapid activation of calcium-sensitive Na+/H+ exchange induced by 20-hydroxyecdysone in salivary gland cells of Drosophila melanogaster. Molecular and Cellular Endocrinology. 1996;116:73–79. doi: 10.1016/0303-7207(95)03699-7. [DOI] [PubMed] [Google Scholar]
- Schroepfer GJ Jr. Oxysterols: modulators of cholesterol metabolism and other processes. Physiological Reviews. 2000;80:361–554. doi: 10.1152/physrev.2000.80.1.361. [DOI] [PubMed] [Google Scholar]
- Selepcova L, Jalc D, Javorsky P, Baran M. Influence of Rhaponticum carthamoides Wild on the growth of ruminal bacteria in vitro and on fermentation in an artificial rumen (Rusitec) Archives of Animal Nutrition. 1993a;43:147–156. doi: 10.1080/17450399309386030. [DOI] [PubMed] [Google Scholar]
- Selepcova L, Magic D, and Vajda V. 1993b Use of Rhaponticum carthamoides Wild. in animals nutrition. Cultivation, Harvesting and Processing of Herbs, Meeting held in The High Tatras, Slovak Republic, June 15–17. Book of Abstracts, p. 76. [Google Scholar]
- Senner V, Sotoodeh A, Paulus W. Regulated gene expression in glioma cells: a comparison of three inducible systems. Neurochemical Research. 2001;26:521–524. doi: 10.1023/a:1010965013113. [DOI] [PubMed] [Google Scholar]
- Shi YJ, Simmons MN, Seki T, Oh SP, Sugrue SP. Change in gene expression subsequent to induction of Pnn/DRS/memA: increase in p21 (cip1/waf1) Oncogene. 2001;20:4007–4018. doi: 10.1038/sj.onc.1204507. [DOI] [PubMed] [Google Scholar]
- Shibatani J, Okada M, Inaoka Y, and Tsuji K. 1996 Preparation of novel steroid and its use for cosmetics and anticancer agents. Application JP 94-271515/19941104 (Chemical Abstracts 125:143132). [Google Scholar]
- Simon P, Koolman J. 1989 Ecdysteroids in vertebrates : pharmalogical aspects. In Koolman J., editor. Ecdysone - from chemistry to mode of action. 254–259.Georg Thieme Verlag, Stuttgart. [Google Scholar]
- Sláma K, Koudela K, Tenora J, Mathova A. Insect hormones in vertebrates : anabolic effects of 20-hydroxyecdysone in Japanese quails. Experientia. 1996;52:702–706. doi: 10.1007/BF01925578. [DOI] [PubMed] [Google Scholar]
- Sláma K, Lafont R. Insect hormones - ecdysteroids : their presence and actions in vertebrates. European Journal of Entomology. 1995;92:355–377. [Google Scholar]
- Sorensen PW, Hara TJ, Stacey NE, Dulka JG. Extreme olfactory specificity of male goldfish to the preovulatory steroidal pheromone 17α,20β-dihydroxy-4-pregnen-3-one. Journal of Comparative Physiology, Part A. 1990;166:373–383. [Google Scholar]
- Sparacio S, Pfeiffer T, Schaal H, Bosch V. Generation of a flexible cell line with regulatable, high-level expression of HIV Gag/Pol particles capable of packaging HIV-derived vectors. Molecular Therapy. 2001;3:602–612. doi: 10.1006/mthe.2001.0296. [DOI] [PubMed] [Google Scholar]
- Stauffer DR, Howard TL, Nyun T, Hollenberg SM. CHMP1 is a novel nuclear matrix protein affecting chromatin structure and cell-cycle progression. Journal of Cell Science. 2001;114:2383–2393. doi: 10.1242/jcs.114.13.2383. [DOI] [PubMed] [Google Scholar]
- Stolarov J, Chang K, Reiner A, Rogers L, Hannon GJ, Wigler MH, Mittal V. Design of a retroviral-mediated ecdysone-inducible system and its application to the expression profiling of the PTEN tumor suppressor. Proceedings of the National Academy of Sciences, USA. 2001;98:13043–13048. doi: 10.1073/pnas.221450598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stopka P, Stancl J, Sláma K. Effect of insect hormone, 20-hydroxyecdysone on growth and reproduction in mice. Acta Societatis Zoologicae Bohemicae. 1999;63:367–378. [Google Scholar]
- Suhr ST, Gil EB, Senut MC, Gage FH. High level transactivation by a modified Bombyx ecdysone receptor in mammalian cells without exogenous retinoid X receptor. Proceedings of the National Academy of Sciences, USA. 1998;95:7999–8004. doi: 10.1073/pnas.95.14.7999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swevers L, Cherbas L, Cherbas P, Iatrou K. Bombyx EcR (BmEcR) and Bombyx USP (BmCF1) combine to form a functional ecdysone receptor. Insect Biochemistry and Molecular Biology. 1996;26:217–221. doi: 10.1016/0965-1748(95)00097-6. [DOI] [PubMed] [Google Scholar]
- Swevers L, Drevet JR, Lunke MD, Iatrou K. The silkmoth homolog* of the Drosophila ecdysone receptor (B1 isoform): cloning and analysis of expression during follicular cell differentiation. Insect Biochemistry and Molecular Biology. 1995;25:857–866. doi: 10.1016/0965-1748(95)00024-p. [DOI] [PubMed] [Google Scholar]
- Syrov VN. Mechanism of the anabolic action of phytoecdysteroids in mammals. Biologichekie Nauki (Moscow) 1984;(11):16–20. [PubMed] [Google Scholar]
- Syrov VN. Phytoecdysteroids: their biological effects in the body of higher animals and the outlook for their use in medicine. Eksperimental'naya i Klinicheskaya Farmakologiya. 1994;57:61–66. [PubMed] [Google Scholar]
- Syrov VN. Comparative experimental investigations of the anabolic activity of ecdysteroids and steranabols. Pharmaceutical and Chemical Journal. 2000;34:193–197. [Google Scholar]
- Syrov VN, Aizikov MI, Kurmukov AG. Effect of ecdysterone on the content of protein, glycogen, and fat in white rat liver, heart and muscle. Doklady Akademii Nauk Uzbeckoy SSR. 1975a;(8):37–38. [Google Scholar]
- Syrov VN, Khushbaktova ZA, Abzalova MKh, Sultanov MB. On the hypolipidemic and antiatherosclerotic action of phytoecdysteroids. Doklady Akademii Nauk Uzbeckoy SSR. 1983;(9):44–45. [Google Scholar]
- Syrov VN, Khushbaktova ZA, Mirzaev YuR, Baltaev UA. Antiulcer activity of phytoecdysteroids. Khimiko Farmatsevticheskii Zhurnal. 1989;23:441–445. [Google Scholar]
- Syrov VN, Khushbaktova ZA, Nabiev AN. An experimental study of the hepatoprotective properties of phytoecdysteroids and nerobol in carbon tetrachloride - induced liver injury. Eksperimental'naya i Klinicheskaya Farmakologiya. 1992;55:61–65. [PubMed] [Google Scholar]
- Syrov VN, Khushbaktova ZA, Tashmukhamedova MA. Hypoglycemic action of phytoecdysteroids and some aspects of its mechanism of realization in experimental animals. Doklady Akademii Nauk Respubliki Uzbekistana. 1997a;(4):46–49. [Google Scholar]
- Syrov VN, Khushbaktova ZA. Wound-healing effects of ecdysteroids. Doklady Akademii Nauk Respubliki Uzbekistana. 1996;(12):47–50. [Google Scholar]
- Syrov VN, Khushbaktova ZA. The pharmacokinetics of phytoecdysteroids and nerobol on animals with experimental toxic renal damage. Eksperimental'naya i Klinicheskaya Farmakologiya. 2001;64:56–58. [PubMed] [Google Scholar]
- Syrov VN, Kurmukov AG, Sakhibov AD. Effect of turkesterone and nerobol on the activity of the protein synthesizing system in mice liver. Voprosy Meditsinskoi Khimii. 1978;24:456–460. [PubMed] [Google Scholar]
- Syrov VN, Kurmukov AG, Sultanov MB. Evaluation of the anabolic effect of phytoecdysones and their 6-keto analogs in tests with female rats. Doklady Akademii Nauk Uzbeckoy SSR. 1981a;(3):31–33. [Google Scholar]
- Syrov VN, Kurmukov AG, Usmanov BZ. Anabolic effects of turkesterone and turkesterone tetraacetate. Doklady Akademii Nauk Uzbeckoy SSR. 1975b;32:32–34. [Google Scholar]
- Syrov VN, Kurmukov AG. On the anabolic effect of viticosterone E. Doklady Akademii Nauk Uzbeckoy SSR. 1975a;(6):31–32. [Google Scholar]
- Syrov VN, Kurmukov AG. Effect of viticosterone E on the weight of organs and their protein content in rats of different ages. Doklady Akademii Nauk Uzbeckoy SSR. 1975b;(9):40–41. [Google Scholar]
- Syrov VN, Kurmukov AG. Anabolic properties of the phytoecdysone turkesterone and turkesterone tetraacetate in experiments on male rats. Problemy Endokrinology. 1976a;22:107–112. [PubMed] [Google Scholar]
- Syrov VN, Kurmukov AG. Biological activity of cyasterone in experiments on male rats. Biologichekie Nauki (Moscow) 1976b;19:72–74. [Google Scholar]
- Syrov VN, Kurmukov AG. Anabolic activity of phytoecdysone-ecdysterone isolated from Rhaponticum carthamoides. Farmakologiya i Toksikologiya. 1976c;39:690–693. [PubMed] [Google Scholar]
- Syrov VN, Matveev SB, Kurmukov AG, Islambekov US. Effect of ecdysterone and nerobol on the healing of experimental bone fractures. Medicinal'nii Zhurnal Uzbekistana. 1986a;(3):67–69. [Google Scholar]
- Syrov VN, Mel'nikova EV, Sultanov MB. Effects of the phytoecdysteroid ecdysterone on the course of heliotrine-induced toxic hepatitis in rats. Doklady Akademii Nauk Uzbeckoy SSR. 1981b;(5):36–38. [Google Scholar]
- Syrov VN, Nabiev AN, Sultanov MB. The effect of phytoecdysteroids on the bile secretion function of the liver in normal rats and in animals with experimental hepatitis. Farmakologiya i Toksikologiya. 1986b;49:100–103. [PubMed] [Google Scholar]
- Syrov VN, Nasyrova SS, Khushbaktova ZA. The results of experimental study of phytoecdysteroids as erythropoiesis stimulators in laboratory animals. Eksperimental'naya i Klinicheskaya Farmakologiya. 1997b;60:41–44. [PubMed] [Google Scholar]
- Syrov VN, Osipova S, Khushbaktova ZA. Influence of prolonged administration of ecdysteron on the spontaneous infection of rabbits with Lamblia duodenalis. Bulletin de la Société Française de Parasitologie. 1990;8(Suppl. 1):466. [Google Scholar]
- Syrov VN, Saatov V, Sagdullaev ShSh, Mamatkhanov AU. Study of the structure – anabolic activity relationship for the phytoecdysteroids extracted from some plants of central Asia. Pharmaceutical Chemistry Journal. 2001;35:667–671. [Google Scholar]
- Takahashi H, Nishimoto K. 1992 Antidiabetic agents containing ecdysterone or inokosterone. Jpn Kokai Tokkyo Koho J.P. 04,125,135 [92 124,135]. (Chem. Abstr. 117:84874b). [Google Scholar]
- Takei M, Endo K, Nishimoto N, Shiobara Y, Inoue S, Matsuo S. Effect of ecdysterone on histamine release from rat peritoneal mast cells. Journal of Pharmaceutical Sciences. 1991;80:309–310. doi: 10.1002/jps.2600800403. [DOI] [PubMed] [Google Scholar]
- Takemoto T, Nishimoto K, Shiobara Y, Fujino M, and Inoue S. 1988 Analgesics containing 20-hydroxyecdysone and its extraction. Jpn Kokai Tokkyo Koho JP 63 02,928 [88 02,928] Appl 86/142,959 (Chemical Abstracts 109:79730p). [Google Scholar]
- Talbot WS, Swyryd EA, Hogness DS. Drosophila tissues with different metamorphic responses to ecdysone express different ecdysone receptor isoforms. Cell. 1993;73:1323–1337. doi: 10.1016/0092-8674(93)90359-x. [DOI] [PubMed] [Google Scholar]
- Tanaka Y, Asaoka K, Takeda S. Different feeding and gustatory responses to ecdysone and 20-hydroxyecdysone by larvae of the silkworm, Bombyx mori. Journal of Chemical Ecology. 1994;20:125–133. doi: 10.1007/BF02065995. [DOI] [PubMed] [Google Scholar]
- Taniguchi SF, Bersani-Amado CA, Sudo LS, Assel SMC, Oga S. Effect of Pfaffia iresinoides on the experimental inflammatory process in rats. Phytotherapy Research. 1997;11:568–571. [Google Scholar]
- Tashmukhamedova MA, Almatov KT, Syrov VN, Sultanov MB, Abidov AA. Effect of phytoecdysteroids and anabolic steroids on respiration and oxidative phosphorylation of rat liver mitochondria under alloxan-diabetes. Biologicheskie Nauki. 1985;9:37–39. [PubMed] [Google Scholar]
- Thomas HE, Stunnenberg HG, Stewart AF. Heterodimerization of the Drosophila ecdysone receptor with retinoid X receptor and ultraspiracle. Nature. 1993;362:471–475. doi: 10.1038/362471a0. [DOI] [PubMed] [Google Scholar]
- Tice CM, Hormann RE, Thompson CS, Friz JL, Cavanaugh CK, Michelotti EL, Garcia J, Nicolas E, Albericio F. Synthesis and SAR of α-acylaminoketone ligands for control of gene expression. Bioorganic and Medicinal Chemistry Letters. 2003;13:475–478. doi: 10.1016/s0960-894x(02)00980-0. [DOI] [PubMed] [Google Scholar]
- Todorov IN, Mitrokhin YuI, Efremova OI, Sidorenko LI. The influence of ecdysterone on the biosynthesis of proteins and nucleic acids in mouse organs. Khimiko Farmatsevticheskii Zhurnal. 2000;34:3–5. [Google Scholar]
- Tomaschko KH. Nongenomic effects of ecdysteroids. Archives of Insect Biochemistry and Physiology. 1999;41:89–98. [Google Scholar]
- Tran HT, Askari HB, Shaaban S, Price L, Palli SR, Dhadialla TS, Carlson GR, Butt TR. Reconstruction of ligand-dependent transactivation of Choristoneura fumiferana ecdysone receptor in yeast. Molecular Endocrinology. 2001;15:1140–1153. doi: 10.1210/mend.15.7.0660. [DOI] [PubMed] [Google Scholar]
- Trenin DS, Volodin VV, Beikin YaB, Shiykova AB. The influence of the ecdysteroid fraction from shoots of Serratula coronata on E-rosette formation and agar migration tests in vitro. Eksperimental'naya i Klinicheskaya Farmakologiya. 1996;59:55–57. [PubMed] [Google Scholar]
- Trenin DS, Volodin VV. 20-Hydroxyecdysone as a human lymphocyte and neutrophil modulator : in vitro evaluation. Archives of Insect Biochemistry and Physiology. 1999;41:156–161. doi: 10.1002/(SICI)1520-6327(1999)41:3<156::AID-ARCH7>3.0.CO;2-Q. [DOI] [PubMed] [Google Scholar]
- Tsitsimpikou C, Tsamis GD, Siskos PA, Spyridaki MH, Georgakopoulos CG. Study of excretion of ecdysterone in human urine. Rapid Communications in Mass Spectrometry. 2001;15:1796–1801. doi: 10.1002/rcm.433. [DOI] [PubMed] [Google Scholar]
- Tsuji K, Hirose T, Okada M, Shibatani J, Hirai Y, Muramatsu N, Inaoka Y, Fukuda T, and Yagi M. 1995a Skin cosmetics or bath preparations containing steroid derivatives as skin metabolism-activating and anti-wrinkling agents. Application JP 94-109471/19940523 (Chemical Abstracts 124:155687). [Google Scholar]
- Tsuji K, Shibatani J, Okada M, and Inaoka Y. 1995b Blood flow improver and cosmetics. PCT International Application. Application JP 94-02075/19941209 (Chemical Abstracts 123:265775). [Google Scholar]
- Tsujiyama S, Ujihara H, Ishihara K, Sasa M. Potentiation of GABA-induced inhibition by 20-hydroxyecdysone, a neurosteroid, in cultured rat cortical neurons. Japanese Journal of Pharmacology. 1995;68:133–136. doi: 10.1254/jjp.68.133. [DOI] [PubMed] [Google Scholar]
- Tuganova AV, Kotsyuruba AV. The in vitro interaction of C27-steroids with the erythrocyte membranes depends on the sterol structure and concentration. Cellular and Molecular Biology Letters. 1996;1:129–135. [Google Scholar]
- Uchiyama M, Ogawa S. 1970 Hypoglycemic formulation containing insect hormones. Application JP 19690506 S. (Chemical Abstracts 74:24985). [Google Scholar]
- Uchiyama M, Otaka T. 1974 Phytoecdysones and protein metabolism in Mammalia. In Burdette WJ, editor. Invertebrate Endocrinology and Hormonal Heterophylly. 375–398.Springer-Verlag, Berlin. [Google Scholar]
- Uchiyama M, Yoshida T. 1974 Effect of ecdysterone on carbohydrate and lipid metabolism. In Burdette WJ, editor. Invertebrate Endocrinology and Hormonal Heterophylly. 401–416.Springer-Verlag, Berlin. [Google Scholar]
- Unger E, Cigan AM, Trimnell M, Xu R-j, Kendall T, Roth B, Albertsen M. A chimeric ecdysone receptor facilitates methoxyfenozide-dependent restoration of male fertility in ms45 maize. Transgenic Research. 2002;11:455–465. doi: 10.1023/a:1020350208095. [DOI] [PubMed] [Google Scholar]
- Van Craenenbroeck K, Vanhoenacker P, Leysen JE, Haegeman G. Evaluation of the tetracycline- and ecdysone-inducible systems for expression of neurotransmitter receptors in mammalian cells. European Journal of Neuroscience. 2001;14:968–976. doi: 10.1046/j.0953-816x.2001.01719.x. [DOI] [PubMed] [Google Scholar]
- Vargas Gonzalez J. 1986 Isolation of ecdysones from plants. Application ES 85-547554 19851003 (Chemical Abstracts 107 : 172800). [Google Scholar]
- Verras M, Mavroidis M, Kokolakis G, Gourzi P, Zacharopoulou A, Mintzas AC. Cloning and characterization of CcEcR: an ecdysone receptor homolog from the Mediterranean fruit fly, Ceratitis capitata. Europen Journal of Biochemistry. 1999;265:798–808. doi: 10.1046/j.1432-1327.1999.00788.x. [DOI] [PubMed] [Google Scholar]
- Vickers ER, Sharrocks AD. The use of inducible engrailed fusion proteins to study the cellular functions of eukaryotic transcription factors. Methods. 2002;26:270–280. doi: 10.1016/S1046-2023(02)00031-2. [DOI] [PubMed] [Google Scholar]
- Vögtli M, Imhof MO, Brown NE, Rauch P, Spindler-Barth M, Lezzi M, Henrich VC. Functional characterization of two Ultraspiracle forms (CtUSP-1 and CtUSP-2) from Chironomus tentans. Insect Biochemistry and Molecular Biology. 1999;29:931–942. doi: 10.1016/s0965-1748(99)00068-5. [DOI] [PubMed] [Google Scholar]
- Volodin VV, Shirshova TI, Burtseva SA, Melnik MV. Biological activity of 20-hydroxyecdysone and its acetates. Rastitel'nye Resursy, Vyp. 1999;2:76–81. [Google Scholar]
- Vongher JL, Frye CA. Progesterone in conjunction with estradiol has neuroprotective effects in an animal model of neurodegeneration. Pharmacology Biochemistry and Behavior. 1999;64:777–785. doi: 10.1016/s0091-3057(99)00140-9. [DOI] [PubMed] [Google Scholar]
- Wakita K, McCormick F, Tetsu O. Method for screening ecdysone-inducible stable cell lines. BioTechniques. 2001;31:414–418. doi: 10.2144/01312rr03. [DOI] [PubMed] [Google Scholar]
- Wang SF, Ayer S, Segraves WA, Williams DR, Raikhel AS. Molecular determinants of differential ligand sensitivities of insect ecdysteroid receptors. Molecular and Cellular Biology. 2000;20:3870–3879. doi: 10.1128/mcb.20.11.3870-3879.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang X, Bauer JH, Li Y, Shao ZH, Zetoune FS, Cattaneo E, Vincenz C. Characterization of a p75(NTR) apoptotic signaling pathway using a novel cellular model. Journal of Biological Chemistry. 2001;276:33812–33820. doi: 10.1074/jbc.M010548200. [DOI] [PubMed] [Google Scholar]
- Wang XH, Jin DY, Ng RWM, Feng HC, Wong YC, Cheung ALM, Tsao SW. Significance of MAD2 expression to mitotic checkpoint control in ovarian cancer cells. Cancer Research. 2002;62:1662–1668. [PubMed] [Google Scholar]
- Whiteman EL, Cho H, Birnbaum MJ. Role of Akt/protein kinase B in metabolism. Trends in Endocrinology and Metabolism. 2002;13:444–451. doi: 10.1016/s1043-2760(02)00662-8. [DOI] [PubMed] [Google Scholar]
- Wing K.D. RH 5849, a nonsteroidal ecdysone agonist: effects on a Drosophila cell line. Science. 1988;241:467–469. doi: 10.1126/science.3393913. [DOI] [PubMed] [Google Scholar]
- Wolter S, Mushinski JF, Saboori AM, Resch K, Kracht M. Inducible expression of a constitutively active mutant of mitogen-activated protein kinase kinase 7 specifically activates c-JUN NH2-terminal protein kinase, alters expression of at least nine genes, and inhibits cell proliferation. Journal of Biological Chemistry. 2002;277:3576–3584. doi: 10.1074/jbc.M105800200. [DOI] [PubMed] [Google Scholar]
- Wu X. 2001 Use of ecdysteroids in preparing medicine for angiocardiopathy. Application CN 2000-12119/200000731 (Chemical Abstracts 135:147422). [Google Scholar]
- Wu X, Jiang Y, Fan S. Effect of ecdysterone on lung contusion from impact. Chinese Journal of Traumatology. 1997;13:295–296. [Google Scholar]
- Wu X, Jiang Y, Fan S, Wang R, Xiang M, Niu H, Li T. Effects of ecdysterone on rat lung reperfusion injury. Zhongguo Yaolixue Tongbao (Chinese Pharmaceutical Bulletin) 1998a;14:256–258. [Google Scholar]
- Wu X, Lin S, Yang Y, Feng S. Effects of ecdysterone on human umbilical vein endothelial cells injuried by tumor necrosis factor. Chinese Journal of Pathophysiology. 1998b;14:58–62. [Google Scholar]
- Wurtz JM, Guillot B, Fagart J, Moras D, Tietjen K, Schindler M. A new model for 20-hydroxyecdysone and dibenzoylhydrazine binding: a homology modeling and docking approach. Protein Science. 2000;9:1073–1084. doi: 10.1110/ps.9.6.1073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wyborski DL, Bauer JC, Vaillancourt P. Bicistronic expression of ecdysone-inducible receptors in mammalian cells. BioTechniques. 2001;31:618–620. 622, 624. doi: 10.2144/01313pf02. [DOI] [PubMed] [Google Scholar]
- Xiao Y-y, Beilstein MA, Wang M-c, Purintrapiban J, Forsberg NE. Development of a ponasterone A-inducible gene expression system for application in cultured skeletal muscle cells. International Journal of Biochemistry and Cell Biology. 2003;35:79–85. doi: 10.1016/s1357-2725(02)00122-x. [DOI] [PubMed] [Google Scholar]
- Xu N, Guo Y, Li X. Advances in pharmacological resarch on ecdysterone. Shenyang Yaoke Daxue Xuebao (Journal of Shenyang Pharmaceutical University) 1997;14:300–302. [Google Scholar]
- Xu N, Guo Y, Rui W, Li Z, Li X. Protective effect of ecdysterone on amnesia induced by diazepam and alcohol. Zhongguo Yaolixue Yu Dilixue Zazhi. 1999;13:119–122. [Google Scholar]
- Xu WM, Liu LZ, Charles IG. Microencapsulated iNOS-expressing cells cause tumor suppression in mice. FASEB Journal. 2001;15:U131–U148. doi: 10.1096/fj.01-0590fje. [DOI] [PubMed] [Google Scholar]
- Xu Y, Mellgren RL. Calpain inhibition decreases the growth rate of mammalian cell colonies. Journal of Biological Chemistry. 2002;277:21474–21479. doi: 10.1074/jbc.M111689200. [DOI] [PubMed] [Google Scholar]
- Yam JWP, Chan KW, Hsiao WLW. Suppression of the tumorigenicity of mutant p53-transformed rat embryo fibroblasts through expression of a newly cloned rat nonmuscle myosin heavy chain-B. Oncogene. 2001;20:58–68. doi: 10.1038/sj.onc.1203982. [DOI] [PubMed] [Google Scholar]
- Yang C, Xu J, Dong Y, Liu X. Studies on the isolation and identification of ?-ecdysone from Zebrina pendula Schnizl. and its antiarrhythmic effect. Tianran Chanwu Yanjiu Yu Kaifa. 1996;8:17–19. [Google Scholar]
- Yang C, Zhang G, Liu X, and Wang C. 2001 Oral antidiabetic compositions containing β-ecdysone from Cyanothis arachnoides. Appl. CN-2000-10637/20000612 (Chem. Abstr. 135:127188). [Google Scholar]
- Yang G, Hannan GN, Lockett TJ, Hill RJ. Functional transfer of an elementary ecdysone gene regulatory system to mammalian cells: transient transfections and stable cell lines. European Journal of Entomology. 1995;92:379–389. [Google Scholar]
- Yao TP, Forman BM, Jiang Z, Cherbas L, Chen JD, McKeown M, Cherbas P, Evans RM. Functional ecdysone receptor is the product of EcR and Ultraspiracle genes. Nature. 1993;366:476–479. doi: 10.1038/366476a0. [DOI] [PubMed] [Google Scholar]
- Yao TP, Segraves WA, Oro AE, McKeown M, Evans RM. Drosophila Ultraspiracle modulates ecdysone receptor function via heterodimer formation. Cell. 1992;71:63–72. doi: 10.1016/0092-8674(92)90266-f. [DOI] [PubMed] [Google Scholar]
- Yarovoi SV, Pederson T. Human cell lines expressing hormone regulated T7 RNA polymerase localized at distinct intranuclear sites. Gene. 2001;275:73–81. doi: 10.1016/s0378-1119(01)00652-7. [DOI] [PubMed] [Google Scholar]
- Yoshida T, Otaka T, Uchiyama M, Ogawa S. Effect of ecdysterone on hyperglycemia in experimental animals. Biochemical Pharmacology. 1971;20:3263–3268. doi: 10.1016/0006-2952(71)90431-x. [DOI] [PubMed] [Google Scholar]
- Zhang MY, Huang NN, Clawson GA, Osmani SA, Pan WH, Xin P, Razzaque MS, Miller BA. Involvement of the fungal nuclear migration gene nudC human homolog in cell proliferation and mitotic spindle formation. Experimental Cell Research. 2002;273:73–84. doi: 10.1006/excr.2001.5414. [DOI] [PubMed] [Google Scholar]
- Zhu B, Benjamin D, Zheng Y, Angliker H, Thiry S, Segmann M, Jost JP. Overexpression of 5-methylcytosine DNA glycosylase in human embryonic kidney cells EcR293 demethylates the promoter of a hormone-regulated reporter gene. Proceedings of the National Academy of Sciences, USA. 2001;98:5031–5036. doi: 10.1073/pnas.091097298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu Y, Rice CD, Pang Y, Pace M, and Thomas P. 2003 Cloning, expression, and characterization of a membrane progestin receptor and evidence it is an intermediary in meiotic maturation in fish oocytes. Proceedings of the National Academy of Sciences, USA. (in press). [DOI] [PMC free article] [PubMed] [Google Scholar]