

Abstract
Background
The changes in the renal function leading to a reduction of medullary blood flow can have a great impact on sodium and water homeostasis and on the long-term control of arterial blood pressure. The RNA-Seq approach was used for transcriptome profiling of the renal medulla from hypertensive ISIAH and normotensive WAG rats to uncover the genetic basis of the changes underlying the renal medulla function in the ISIAH rats being a model of the stress-sensitive arterial hypertension and to reveal the genes which possibly may contribute to the alterations in medullary blood flow.
Results
Multiple DEGs specifying the function of renal medulla in ISIAH rats were revealed. The group of DEGs described by Gene Ontology term ‘oxidation reduction’ was the most significantly enriched one. The other groups of DEGs related to response to external stimulus, response to hormone (endogenous) stimulus, response to stress, and homeostatic process provide the molecular basis for integrated responses to homeostasis disturbances in the renal medulla of the ISIAH rats. Several DEGs, which may modulate the renal medulla blood flow, were detected. The reduced transcription of Nos3 pointed to the possible reduction of the blood flow in the renal medulla of ISIAH rats.
Conclusions
The generated data may be useful for comparison with those from different models of hypertension and for identifying the common molecular determinants contributing to disease manifestation, which may be potentially used as new pharmacological targets.
Electronic supplementary material
The online version of this article (doi:10.1186/s12863-016-0462-6) contains supplementary material, which is available to authorized users.
Keywords: Stress-sensitive hypertension, Renal medulla, Transcriptional profiling, RNA-Seq, ISIAH rats
Background
The study of molecular-genetic mechanisms of hypertension is an important task for biology and medicine. Multifactorial etiology of hypertension complicates the solution of this problem, and despite of numerous studies, the basal molecular mechanisms of essential hypertension remain not fully elucidated. However, it has long been known that renal dysfunction underlies the development of all forms of hypertension in experimental animals and humans [1–4].
The ISIAH (Inherited Stress-Induced Arterial Hypertension) rat strain was developed to study the genetic background of the stress-sensitive form of arterial hypertension and its complications. Selection of the ISIAH rats from an outbred Wistar stock was performed for a systolic arterial blood pressure (BP) elevation induced by 0.5 h emotional stress, which was caused by keeping the rats restricted in a small wire mesh cage. The ISIAH rats acquire the elevated basal systolic arterial BP at the age of 6 weeks (175.0 ± 3.5 mmHg in males and 165.0 ± 3.0 mmHg in females) and systolic arterial BP in these rats dramatically increases under the restriction conditions [5, 6]. The development of the hypertensive state in ISIAH rats is also accompanied by a hypertrophy of the left ventricle, increase in the wall thickness of the small arteries, and changes in the electrocardiographic pattern [6]. The studies on the kidney histology showed the alterations, which were indicative of an increase in filtration barrier functional load and of processes leading to the development of glomerular [7] and medullary sclerosis in ISIAH rats [8].
The unraveling of the genetic basis of the renal function in ISIAH rats may be useful for understanding the mechanisms underlying the stress-sensitive hypertension development and for identifying the molecular determinants, which may be potentially used as the therapeutic targets for pharmacological intervention.
Recently, using the next-generation RNA sequencing (RNA-Seq) approach, we analyzed the renal cortex transcriptome in ISIAH rats [9]. The results of this study showed that the functioning of the renal cortex in ISIAH rats is based on the changes in transcriptional activity of multiple genes related to different biological processes and metabolic pathways. However, in a number of studies it was demonstrated that the initial changes leading to the pathology of renal function begin from medullary blood flow reduction, and these changes may exert significant effects on sodium and water homeostasis and on the long-term control of arterial BP [2, 10, 11].
Thus, the goal of the current study was to uncover the genetic basis of the changes underlying the renal medulla function in the ISIAH rats and to reveal the features which possibly may be related to the alterations in medullary blood flow. To achieve this goal the RNA-Seq approach was used for transcriptome profiling of the renal medulla from hypertensive ISIAH and normotensive WAG rats. The differentially expressed genes (DEGs) related to stress-sensitive hypertension and possibly to the alterations in medullary blood flow as well as the metabolic pathways contributing to the inter-strain differences in renal medulla functions were detected. Several DEGs, which may modulate the renal medulla blood flow, were identified and their possible impact on the process was discussed.
Results
The expression of 13,646 genes was detected in the renal medulla of analyzed rats. The comparative analysis of their expression in the renal medulla of ISIAH and WAG rats revealed 960 DEGs (listed in the Additional file 1). The Additional file 2 represents the heatmap for the DEGs. Approximately a half of these genes (524 genes, i.e., 54.7%) were down-regulated in the renal medulla of ISIAH rats. The expression of 11 genes was detected in renal medulla of only one rat strain (Additional file 3). One of these genes (Retn, resistin) is known as associated with hypertension. Its expression was detected in renal medulla of ISIAH rats but not in WAG. The list of the top 40 genes with the highest differences in their expression in renal medulla of ISIAH and WAG rats included two genes (Acsm3, acyl-CoA synthetase medium-chain family member 3; and Ephx2, epoxide hydrolase 2, cytoplasmic), which were annotated in Rat Genome Database (RGD) as genes related to hypertension (Table 1).
Table 1.
Top 40 genes with the greatest difference in expression between ISIAH and WAG renal medulla
| Gene symbol | NCBI gene ID | Gene definition | log2 (fold_change) ISIAH/WAG |
|---|---|---|---|
| LOC102546948 | 102546948 | uncharacterized LOC102546948 | −5.73 |
| Galnt13 | 311039 | UDP-N-acetyl-alpha-D-galactosamine:polypeptide N-acetylgalactosaminyltransferase 13 (GalNAc-T13) | −5.56 |
| Pdilt | 293544 | protein disulfide isomerase-like, testis expressed | −5.17 |
| LOC501110 | 501110 | similar to Glutathione S-transferase A1 (GTH1) (HA subunit 1) (GST-epsilon) (GSTA1-1) (GST class-alpha) | −4.59 |
| Slc10a2 | 29500 | solute carrier family 10 (sodium/bile acid cotransporter), member 2 | −4.40 |
| Pcdh9 | 306091 | protocadherin 9 | −4.23 |
| LOC100909561 | 100909561 | nuclease-sensitive element-binding protein 1-like | −3.74 |
| Car5a | 54233 | carbonic anhydrase 5a, mitochondrial | −3.74 |
| LOC100361907 | 100361907 | complement factor H-related protein B | −3.73 |
| Sphkap | 316561 | SPHK1 interactor, AKAP domain containing | −3.65 |
| LOC102552001 | 102552001 | uncharacterized LOC102552001 | −3.62 |
| LOC102550987 | 102550987 | uncharacterized LOC102550987 | −3.57 |
| Kcnj5 | 29713 | potassium inwardly-rectifying channel, subfamily J, member 5 | −3.23 |
| Upk2 | 689093 | uroplakin 2 | 3.12 |
| LOC102555352 | 102555352 | uncharacterized LOC102555352 | 3.13 |
| LOC102551856 | 102551856 | uncharacterized LOC102551856 | 3.21 |
| Spta1 | 289257 | spectrin, alpha, erythrocytic 1 (elliptocytosis 2) | 3.28 |
| LOC686967 | 686967 | similar to olfactory receptor 1442 | 3.34 |
| Acsm3 a | 24763 | acyl-CoA synthetase medium-chain family member 3 | 3.40 |
| LOC100911960 | 100911960 | UDP-glucuronosyltransferase 1-9-like | 3.44 |
| Thrsp | 25357 | thyroid hormone responsive | 3.51 |
| Sprr1a | 499660 | small proline-rich protein 1A | 3.51 |
| Hpgd | 79242 | hydroxyprostaglandin dehydrogenase 15 (NAD) | 3.52 |
| LOC102553290 | 102553290 | collagen alpha-1(III) chain-like | 3.62 |
| Ly6al | 362935 | lymphocyte antigen 6 complex, locus A-like | 3.65 |
| Akr1b8 | 286921 | aldo-keto reductase family 1, member B8 | 3.70 |
| Shisa3 | 498356 | shisa family member 3 | 3.83 |
| Nefh | 24587 | neurofilament, heavy polypeptide | 3.90 |
| Tcerg1l | 361669 | transcription elongation regulator 1-like | 4.09 |
| Krt19 | 360626 | keratin 19 | 4.18 |
| Ephx2 a | 65030 | epoxide hydrolase 2, cytoplasmic | 4.52 |
| Ubd | 29168 | ubiquitin D | 4.61 |
| Resp18 | 50561 | regulated endocrine-specific protein 18 | 4.65 |
| Serpinb12 | 304692 | serpin peptidase inhibitor, clade B (ovalbumin), member 12 | 5.38 |
| Car3 | 54232 | carbonic anhydrase 3 | 5.54 |
| Fam111a | 499322 | family with sequence similarity 111, member A | 5.86 |
| LOC100362069 | 100362069 | ribosomal protein L28-like | 6.11 |
| Gys2 | 25623 | glycogen synthase 2 | 6.38 |
| Stk32c | 365381 | serine/threonine kinase 32C | 6.41 |
| RGD1565131 | 498143 | 60S ribosomal protein L15-like | 8.94 |
aGenes are annotated in Rat Genome Database (http://rgd.mcw.edu/) as associated with hypertension. ISIAH and WAG – rat strains used in the study
The genes making the strongest contribution to the inter-strain differences were detected by the partial-least squares discriminant analysis (PLS-DA). The distances between ISIAH and WAG rats are shown in Fig. 1a, and the results of the correlation analysis between gene expression and PLS-DA Axis 1 are presented in Fig. 1b. The DEGs are shown in red in Fig. 1b. The DEGs in the most polar position contribute the most to the inter-strain variations. The correlation coefficients for Acsm3 and Ephx2 were 0.996 and 0.994, correspondingly. Hence, Acsm3 and Ephx2 may be considered as genes making the strongest contribution to the inter-strain differences. The differential expression of these and several other genes was validated by real-time PCR (Fig. 2). The correlation between the gene expression estimated by two methods (RNA-Seq and real-time PCR) was 0.99 (Fig. 3).
Fig. 1.

a Axes maximizing the distances between ISIAH and WAG rats. b The distribution of expressed genes along the first axis. The data are based on the Pearson correlation coefficients between the first axis and the level of genes expression
Fig. 2.
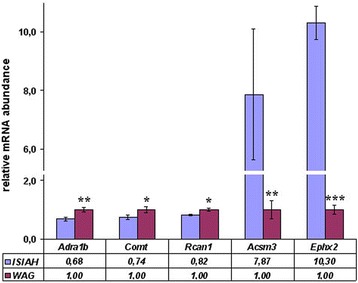
The relative mRNA abundance measured by qPCR. The normalized mRNA level in control samples of the WAG rats was assigned a value of 1. Vertical bars show the standard error of the mean, and significance of inter-strain difference is indicated by *p < 0.05, **p < 0.01, ***p < 0.001
Fig. 3.

Comparison of gene expression level measurements obtained by RNA-seq and qPCR
According to the RGD annotations, 58 DEGs found in the current study are associated with hypertension (Table 2). Seven of them (Agtr1a, Fn1, Gja1, Lama2, Mmp2, Mmp9, Nos3) are known as being associated with renal hypertension. The functional annotation in Database for Annotation, Visualization and Integrated Discovery (DAVID) additionally revealed four DEGs (Col1a2, Guca2b, P2rx4, Pcsk5) related to BP regulation. The transcription of most of genes associated with hypertension (67.2%) was reduced in the renal medulla of ISIAH rats. About a half of the DEGs related to hypertension are also known as associated with diabetic nephropathy and insulin resistance, and two DEGs were found to be related to nephrosclerosis (Table 2). Many of DEGs listed in the Table 2 are related to the diseases of immune system. The study revealed 76 DEGs referred to in RGD as associated with different renal diseases, such as renal insufficiency, nephrosclerosis, diabetic nephropathy, and renal fibrosis (Table 3).
Table 2.
Genes differentially expressed in ISIAH and WAG renal medulla and referred to in Databases as associated with hypertension and blood pressure regulation
| Gene symbol | NCBI gene ID | Gene name | log2 fold_change ISIAH/WAG |
|---|---|---|---|
| Rat Genome Database | |||
| Acsm3 e | 24763 | acyl-CoA synthetase medium-chain family member 3 | 3.40 |
| Adipoq c,d,e | 246253 | adiponectin, C1Q and collagen domain containing | 1.17 |
| Adra1b c | 24173 | adrenoceptor alpha 1B | −1.31 |
| Adra2a | 25083 | adrenoceptor alpha 2° | −1.15 |
| Agtr1a a,c,d,e | 24180 | angiotensin II receptor, type 1° | −0.47 |
| Alas1 | 65155 | aminolevulinate, delta-, synthase 1 | 0.74 |
| Angpt2 e | 89805 | angiopoietin 2 | −0.79 |
| Aqp2 d | 25386 | aquaporin 2 (collecting duct) | −0.47 |
| Aqp4 e | 25293 | aquaporin 4 | −0.59 |
| Clu d,e | 24854 | clusterin, transcript variant X1 | −0.80 |
| Cnr1 c | 25248 | cannabinoid receptor 1 (brain) | −0.58 |
| Comt | 24267 | catechol-O-methyltransferase | −0.67 |
| Corin | 289596 | corin, serine peptidase | −1.04 |
| Cst3 c,d,e | 25307 | cystatin C | −0.50 |
| Cyp1a1 e | 24296 | cytochrome P450, family 1, subfamily a, polypeptide 1 | −0.51 |
| Cyp4f1 | 56266 | cytochrome P450, family 4, subfamily f, polypeptide 1 | 0.60 |
| Dio2 c | 65162 | deiodinase, iodothyronine, type II | −2.56 |
| Dusp1 e | 114856 | dual specificity phosphatase 1 | −0.71 |
| Ebag9 | 299864 | estrogen receptor binding site associated, antigen, 9 | 0.49 |
| Emilin1 | 298845 | elastin microfibril interfacer 1 | −0.65 |
| Ephx1 e | 25315 | epoxide hydrolase 1, microsomal (xenobiotic) | 0.70 |
| Ephx2 c,d | 65030 | epoxide hydrolase 2, cytoplasmic | 4.52 |
| F2 d, e | 29251 | coagulation factor II | −0.93 |
| Fbn1 d | 83727 | fibrillin 1, transcript variant X1 | −0.61 |
| Fn1 a,d,e | 25661 | fibronectin 1 | −0.98 |
| Gja1 a,e | 24392 | gap junction protein, alpha 1 | 0.99 |
| Gstm2 | 24424 | glutathione S-transferase mu 2 | −0.76 |
| Gstp1 e | 24426 | glutathione S-transferase pi 1 | 0.53 |
| Hgf c,d,e | 24446 | hepatocyte growth factor | −0.77 |
| Hsd17b4 e | 79244 | hydroxysteroid (17-beta) dehydrogenase 4 | 0.52 |
| Igf1 d,e | 24482 | insulin-like growth factor 1 | −0.75 |
| Itgav c | 296456 | integrin, alpha V | −0.60 |
| Lama2 a | 309368 | laminin, alpha 2 | −0.58 |
| Lox | 24914 | lysyl oxidase | −0.81 |
| Mif c,e | 81683 | macrophage migration inhibitory factor (glycosylation-inhibiting factor) | 0.55 |
| Mmp2 a,e | 81686 | matrix metallopeptidase 2 | −0.54 |
| Mmp9 ad,e | 81687 | matrix metallopeptidase 9 | −2.00 |
| Mthfr b,d,e | 362657 | methylenetetrahydrofolate reductase (NAD(P)H) | 0.49 |
| Nos3 a,c,d,e | 24600 | nitric oxide synthase 3, endothelial cell | −0.55 |
| Nox4 d | 85431 | NADPH oxidase 4 | 0.50 |
| Pappa | 313262 | pregnancy-associated plasma protein A | −0.68 |
| Pecam1 d,e | 29583 | platelet/endothelial cell adhesion molecule 1 | −0.45 |
| Ptgds b,c,d | 25526 | prostaglandin D2 synthase (brain) | −0.78 |
| Ptk2b c,e | 50646 | protein tyrosine kinase 2 beta | −0.79 |
| Retn c,d,e | 246250 | resistin | expression was detected only in ISIAH rats |
| Rnpep | 81761 | arginyl aminopeptidase (aminopeptidase B) | −0.47 |
| RT1-Bb e | 309622 | RT1 class II, locus Bb | −1.26 |
| Serpine2 | 29366 | serpin peptidase inhibitor, clade E (nexin, plasminogen activator inhibitor type 1), member 2 | −0.55 |
| Slc26a4 | 29440 | solute carrier family 26 (anion exchanger), member 4 | 0.59 |
| Slc2a4 c | 25139 | solute carrier family 2 (facilitated glucose transporter), member 4 | −0.76 |
| Slc5a2 | 64522 | solute carrier family 5 (sodium/glucose cotransporter), member 2 | 0.64 |
| Slc6a19 | 664630 | solute carrier family 6 (neutral amino acid transporter), member 19 | 0.48 |
| Slc8a1 | 29715 | solute carrier family 8 (sodium/calcium exchanger), member 1 | 0.53 |
| Slc9a3r2 | 116501 | solute carrier family 9, subfamily A (NHE3, cation proton antiporter 3), member 3 regulator 2 | −0.49 |
| Sod3 c,d | 25352 | superoxide dismutase 3, extracellular | 0.56 |
| Tacr3 | 24808 | tachykinin receptor 3 | −1.22 |
| Tf c,d,e | 24825 | transferrin | −1.43 |
| Vcam1 c,d,e | 25361 | vascular cell adhesion molecule 1 | −0.86 |
| DAVID | |||
| Col1a2 e | 84352 | collagen, type I, alpha 2 | −0.53 |
| Guca2b | 64055 | guanylate cyclase activator 2B | 2.03 |
| P2rx4 | 29659 | purinergic receptor P2X, ligand-gated ion channel 4 | −1.41 |
| Pcsk5 | 116548 | proprotein convertase subtilisin/kexin type 5 | 0.73 |
ISIAH and WAG – rat strains used in the study. DAVID – Database for Annotation, Visualization and Integrated Discovery (http://david.abcc.ncifcrf.gov/)
Genes associated with: arenal hypertension; bnephrosclerosis; cinsulin resistance; ddiabetic nephropathy; eimmune system diseases
Table 3.
Genes differentially expressed in ISIAH versus WAG renal medulla and annotated in Rat Genome Database as associated with kidney diseases
| Gene symbol | NCBI gene ID | Gene name | log2 fold change ISIAH/WAG |
|---|---|---|---|
| Acsm3 | 24763 | acyl-CoA synthetase medium-chain family member 3 | 3.40 |
| Adipoq c,d | 246253 | adiponectin, C1Q and collagen domain containing | 1.17 |
| Agtr1a a,c,d | 24180 | angiotensin II receptor, type 1a | −0.47 |
| Ak4 c | 29223 | adenylate kinase 4 | 0.63 |
| Alpl | 25586 | alkaline phosphatase, liver/bone/kidney | 0.53 |
| Amacr | 25284 | alpha-methylacyl-CoA racemase | 0.50 |
| Angpt2 a | 89805 | angiopoietin 2 | −0.79 |
| Anxa2 | 56611 | annexin A2 | −0.45 |
| Apoh d | 287774 | apolipoprotein H (beta-2-glycoprotein I) | −1.52 |
| Aqp2 d | 25386 | aquaporin 2 (collecting duct) | −0.47 |
| Baat | 29725 | bile acid CoA: amino acid N-acyltransferase (glycine N-choloyltransferase) | 0.78 |
| Car2 | 54231 | carbonic anhydrase 2 | 0.53 |
| Cftr | 24255 | cystic fibrosis transmembrane conductance regulator | −1.22 |
| Cldn19 | 298487 | claudin 19 | −0.53 |
| Clu a,c,d | 24854 | clusterin | −0.80 |
| Col3a1 a,c | 84032 | collagen, type III, alpha 1 | −0.64 |
| Col4a5 | 363457 | collagen, type IV, alpha 5 | −0.45 |
| Comt | 24267 | catechol-O-methyltransferase | −0.67 |
| Csf1r c,d | 307403 | colony stimulating factor 1 receptor | −0.73 |
| Cst3 d | 25307 | cystatin C | −0.50 |
| Cubn d | 80848 | cubilin (intrinsic factor-cobalamin receptor) | 0.68 |
| Cyp1a1 c | 24296 | cytochrome P450, family 1, subfamily a, polypeptide 1 | −0.51 |
| Cyp4a3 | 298423 | cytochrome P450, family 4, subfamily a, polypeptide 3 | 0.52 |
| Dusp1 | 114856 | dual specificity phosphatase 1 | −0.71 |
| Ebag9 | 299864 | estrogen receptor binding site associated, antigen, 9 | 0.49 |
| Ephx2 c,d | 65030 | epoxide hydrolase 2, cytoplasmic | 4.52 |
| F2 c,d | 29251 | coagulation factor II | −0.93 |
| Fbn1 a,d | 83727 | fibrillin 1 | −0.61 |
| Fga c | 361969 | fibrinogen alpha chain | 0.85 |
| Fhit | 60398 | fragile histidine triad | 1.58 |
| Fmod d | 64507 | fibromodulin | −1.09 |
| Fn1 a,c,d | 25661 | fibronectin 1 | −0.98 |
| Gatm c | 81660 | glycine amidinotransferase (L-arginine:glycine amidinotransferase) | 0.86 |
| Gja1 | 24392 | gap junction protein, alpha 1 | 0.99 |
| Gpc1 | 58920 | glypican 1 | −0.72 |
| Gstp1 c | 24426 | glutathione S-transferase pi 1 | 0.53 |
| Gtpbp4 c | 114300 | GTP binding protein 4 | 3.01 |
| Hao1 | 311446 | hydroxyacid oxidase (glycolate oxidase) 1 | −1.60 |
| Hgf d | 24446 | hepatocyte growth factor | −0.77 |
| Igf1 d | 24482 | insulin-like growth factor 1 | −0.75 |
| Igfbp1 d | 25685 | insulin-like growth factor binding protein 1 | 1.26 |
| Il4r | 25084 | interleukin 4 receptor | 0.64 |
| Lama2 | 309368 | laminin, alpha 2 | −0.58 |
| Lgals1 | 56646 | lectin, galactoside-binding, soluble, 1 | 0.88 |
| Lrp1 a | 299858 | low density lipoprotein receptor-related protein 1 | −0.53 |
| Lrp5 | 293649 | low density lipoprotein receptor-related protein 5 | −0.47 |
| Ltbp1 | 59107 | latent transforming growth factor beta binding protein 1 | −0.57 |
| Mif | 81683 | macrophage migration inhibitory factor (glycosylation-inhibiting factor) | 0.55 |
| Mme | 24590 | membrane metallo-endopeptidase | 1.35 |
| Mmp2 a, c | 81686 | matrix metallopeptidase 2 | −0.54 |
| Mmp9 a, d | 81687 | matrix metallopeptidase 9 | −2.00 |
| Mok | 362787 | MOK protein kinase | −0.93 |
| Mthfr b, c, d | 362657 | methylenetetrahydrofolate reductase (NAD(P)H) | 0.49 |
| Nos3 c, d | 24600 | nitric oxide synthase 3, endothelial cell | −0.55 |
| Nox4 d | 85431 | NADPH oxidase 4 | 0.50 |
| Nphs2 a | 170672 | nephrosis 2, idiopathic, steroid-resistant | 0.57 |
| Pecam1 d | 29583 | platelet/endothelial cell adhesion molecule 1 | −0.45 |
| Pla2g4a | 24653 | phospholipase A2, group IVA (cytosolic, calcium-dependent) | −0.69 |
| Ptgds b, d | 25526 | prostaglandin D2 synthase (brain) | −0.78 |
| Ptk2b | 50646 | protein tyrosine kinase 2 beta | −0.79 |
| Rap1gap a | 313644 | Rap1 GTPase-activating protein | −0.60 |
| Retn c, d | 246250 | resistin | expression was detected only in ISIAH rats |
| RT1-Bb | 309622 | RT1 class II, locus Bb | −1.26 |
| Serpinf1 d | 287526 | serpin peptidase inhibitor, clade F (alpha-2 antiplasmin, pigment epithelium derived factor), member 1 | −2.10 |
| Serping1 | 295703 | serpin peptidase inhibitor, clade G (C1 inhibitor), member 1 | 0.56 |
| Sfrp1 | 84402 | secreted frizzled-related protein 1 | −0.61 |
| Slc17a2 | 306950 | solute carrier family 17, member 2 | 0.59 |
| Slc19a3 c | 316559 | solute carrier family 19 (thiamine transporter), member 3 | 1.00 |
| Slc5a2 | 64522 | solute carrier family 5 (sodium/glucose cotransporter), member 2 | 0.64 |
| Slc6a19 | 664630 | solute carrier family 6 (neutral amino acid transporter), member 19 | 0.48 |
| Slit2 | 360272 | slit homolog 2 (Drosophila) | 0.45 |
| Sod3 d | 25352 | superoxide dismutase 3, extracellular | 0.56 |
| Tf d | 24825 | transferrin | −1.43 |
| Timp2 d | 29543 | TIMP metallopeptidase inhibitor 2 | −0.52 |
| Vcam1 c,d | 25361 | vascular cell adhesion molecule 1 | −0.86 |
| Vtn d | 29169 | vitronectin | −1.18 |
Genes associated with: arenal fibrosis; bnephrosclerosis; crenal insufficiency, ddiabetic nephropathy; ISIAH and WAG – rat strains used in the study
The differential transcription of 59 transcription factor genes was found in the renal medulla of ISIAH and WAG rats (Table 4). No one of these genes is currently referred to in databases as associated with arterial hypertension or kidney diseases.
Table 4.
Transcription factor genes differentially expressed in ISIAH and WAG renal medulla
| Gene symbol | NCBI gene ID | Gene name | log2 fold_change ISIAH/WAG |
|---|---|---|---|
| Alx1 | 25401 | ALX homeobox 1 | −1.13 |
| Arntl | 29657 | aryl hydrocarbon receptor nuclear translocator-like | −1.01 |
| Bcl6 | 303836 | B-cell CLL/lymphoma 6 | −2.02 |
| Bcl6b | 360551 | B-cell CLL/lymphoma 6, member B | −0.80 |
| Bhlhe22 | 365748 | basic helix-loop-helix family, member e22 | −1.08 |
| Bnc2 | 298189 | basonuclin 2 | 0.61 |
| Btbd11 | 314675 | BTB (POZ) domain containing 11 | −0.65 |
| Ccrn4l | 310395 | CCR4 carbon catabolite repression 4-like (S, cerevisiae) | 0.64 |
| Cebpd | 25695 | CCAAT/enhancer binding protein (C/EBP), delta | −0.83 |
| Dab2 | 79128 | disabled 2, mitogen-responsive phosphoprotein | 0.49 |
| Dmrt2 | 309430 | doublesex and mab-3 related transcription factor 2 | −0.55 |
| Esrrb | 299210 | estrogen-related receptor beta | −0.58 |
| Etv5 | 303828 | ets variant 5 | −0.70 |
| Fhl1 | 25177 | four and a half LIM domains 1 | 1.09 |
| Gcfc2 | 312474 | GC-rich sequence DNA-binding factor 2 | −0.61 |
| Gne | 114711 | glucosamine (UDP-N-acetyl)-2-epimerase/N-acetylmannosamine kinase | −0.51 |
| Grhl1 | 313993 | grainyhead-like 1 (Drosophila) | 1.91 |
| Hand2 | 64637 | heart and neural crest derivatives expressed 2 | expression was detected only in ISIAH rats |
| Hes6 | 316626 | hes family bHLH transcription factor 6 | −0.72 |
| Id3 | 25585 | inhibitor of DNA binding 3 | −0.50 |
| Id4 | 291023 | inhibitor of DNA binding 4 | 0.47 |
| Ilf2 | 310612 | interleukin enhancer binding factor 2 | −0.46 |
| Irf7 | 293624 | interferon regulatory factor 7 | 0.59 |
| Irf8 | 292060 | interferon regulatory factor 8 | −0.58 |
| Khdrbs3 | 64015 | KH domain containing, RNA binding, signal transduction associated 3 | −1.23 |
| Lmo2 | 362176 | LIM domain only 2 | −0.79 |
| Map2 | 25595 | microtubule-associated protein 2 | −1.44 |
| Mcm7 | 288532 | minichromosome maintenance complex component 7 | 0.61 |
| Mef2c | 499497 | myocyte enhancer factor 2C | −0.59 |
| Mybl1 | 297783 | myeloblastosis oncogene-like 1 | −0.56 |
| Nfkbil1 | 361794 | nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor-like 1 | 0.98 |
| Notch3 | 56761 | notch 3 | −0.50 |
| Npas2 | 316351 | neuronal PAS domain protein 2 | −0.90 |
| Nr2f1 | 81808 | nuclear receptor subfamily 2, group F, member 1 | −1.31 |
| Nrip3 | 361625 | nuclear receptor interacting protein 3 | −0.83 |
| Osr1 | 298878 | odd-skipped related transciption factor 1 | −1.29 |
| Osr2 | 315039 | odd-skipped related transciption factor 2 | 1.17 |
| P8 | 113900 | nuclear proten 1 | −0.95 |
| Patz1 | 305471 | POZ (BTB) and AT hook containing zinc finger 1 | 0.57 |
| Paxbp1 | 681004 | PAX3 and PAX7 binding protein 1 | −0.53 |
| Pou2af1 | 690528 | POU class 2 associating factor 1 | 1.19 |
| Prdm6 | 307305 | PR domain containing 6 | 1.50 |
| Prox1 | 305066 | prospero homeobox 1 | −1.06 |
| Sim1 | 309888 | single-minded family bHLH transcription factor 1 | −0.50 |
| Smarcd3 | 296732 | SWI/SNF related, matrix associated, actin dependent regulator of chromatin, subfamily d, member 3 | −0.58 |
| Spry4 | 291610 | sprouty homolog 4 (Drosophila) | −0.61 |
| Stat5a | 24918 | signal transducer and activator of transcription 5A | 0.63 |
| Tcerg1l | 361669 | transcription elongation regulator 1-like | 4.09 |
| Tcf21 | 252856 | transcription factor 21 | −0.48 |
| Tcf4 | 84382 | transcription factor 4 | −0.66 |
| Tle2 | 299636 | transducin-like enhancer of split 2 (E(sp1) homolog, Drosophila) | −0.61 |
| Zbtb16 | 353227 | zinc finger and BTB domain containing 16 | −1.11 |
| Zdhhc2 | 246326 | zinc finger, DHHC-type containing 2 | 0.75 |
| Zfhx2 | 305888 | zinc finger homeobox 2 | −0.88 |
| Zfp189 | 313219 | zinc finger protein 189 | −0.62 |
| Zfp354a | 24522 | zinc finger protein 354A | 0.58 |
| Zfp449 | 684901 | zinc finger protein 449 | −0.71 |
| Zfp710 | 293044 | zinc finger protein 710 | −0.76 |
| Zfp958 | 100302405 | zinc finger protein 958 | −0.64 |
ISIAH and WAG – rat strains used in the study
Gene Ontology (GO) terms for biological processes defined as significantly enriched within the analysis in DAVID are represented in Additional file 4. GO terms which might be essential for stress-sensitive hypertension development in ISIAH rats are shown in bold in this file, and the details for DEGs from these groups are given in Additional file 5.
The most significantly enriched GO terms were those related to oxidation reduction, lipid metabolic process, response to external stimulus as well as regulation of response to external stimuli. The analysis showed that the differences in renal medulla function between ISIAH and WAG rats are under control of many genes participating in response to hormone stimulus including genes associated with response to steroid (glucocorticoid) hormone stimulus.
Multiple DEGs were related to transport, (including sodium ion transport, water transport, glucose transport, lipid transport) and regulation of transport. Many DEGs associated with transport are annotated also as responsible for homeostatic process, including ion homeostasis.
Several GO terms were related to BP control. These were the groups of DEGs associated with blood circulation, regulation of BP, and negative regulation of blood coagulation.
Two large groups of DEGs were associated with response to stress and regulation of response to stress. The specificity of the stress types was defined by GO terms related to responses to oxidative stress, to osmotic stress, and to salt stress.
The other processes which might be involved in hypertension development in ISIAH rats were: neurogenesis, regulation of action potential in neuron, angiogenesis, regulation of smooth muscle cell differentiation, cell adhesion and regulation of cell adhesion.
The functional annotation underlined the important role of the immune system process and its regulation. The analysis in the Kyoto Encyclopedia of Genes and Genomes Pathway Database (KEGG) performed within the DAVID highlighted sixteen pathways that were significantly (p < 0.05) enriched (Additional file 6). Several of them were also related to immune system functioning. All of the pathways contained genes annotated in RGD as associated with hypertension or with kidney diseases.
Discussion
The transcriptome profiling of the renal medulla from ISIAH and WAG rats let to identify multiple DEGs and several metabolic pathways contributing to differences between the renal medulla functions in ISIAH rats with stress-sensitive hypertension and normotensive controls. The functional annotation of DEGs within the analysis in Databases demonstrated that many of them are associated with hypertension and regulation of BP. One of these, Retn, was expressed in renal medulla of hypertensive rats but not in controls. However, the low level of expression of this gene was reported in kidney from Fischer 344 male rats, too [12]. So, the inter-strain differences in transcriptional activity of Retn shouldn’t be essential for hypertension development in ISIAH rats.
In recent years, it has become clear that a key determinant of the set point of the renal pressure-natriuresis curve is the balance of reactive oxygen and nitrogen species within the renal medullary region [11]. The experimentally induced elevations of either superoxide or hydrogen peroxide in the renal medulla result in reduction of medullary blood flow, enhanced sodium reabsorption, and hypertension [13].
In the current study, the group of DEGs described by GO term ‘oxidation reduction’ was the most significantly enriched one (Additional file 5). This group contained several genes associated with hypertension and some of them (Cyp1a1, cytochrome P450, family 1, subfamily a, polypeptide 1; Cyp4f1, cytochrome P450, family 4, subfamily f, polypeptide 1; Nos3, nitric oxide synthase 3, endothelial cell; Nox4, NADPH oxidase 4, Sod3, superoxide dismutase 3, extracellular) are known as involved in modulation of vascular tone and renal tubular function.
Cyp1a1 knockout mice are hypertensive. Cyp1a1 metabolizes omega-3 polyunsaturated fatty acids to vasodilators and the loss of these vasodilators may lead to increases in BP [14]. CYP1A1 contributes to eNOS (same as Nos3) activation, nitric oxide (NO) bioavailability, and NO-dependent BP regulation [15]. So, the decreased level of Cyp1a1 transcription in ISIAH renal medulla suggests its contribution to hypertension development in ISIAH rats.
CYP4F1 was characterized as the most critical 4 F isoform involved in the production of 20-hydroxyeicosatetraenoic acid (20-HETE), a potent eicosanoid that modulates vascular tone and renal tubular function [16]. 20-HETE has been shown to play a complex role in BP regulation. In the kidney tubules, 20-HETE inhibits sodium reabsorption and promotes natriuresis, thus, contributing to antihypertensive mechanisms. In contrast, in the microvasculature, 20-HETE plays a pressor role by sensitizing smooth muscle cells to constrictor stimuli and increasing vascular myogenic tone, and by acting on the endothelium to further promote endothelial dysfunction and endothelial activation. In addition, 20-HETE induces endothelial angiotensin-converting enzyme, thus, setting forth a potential feed forward prohypertensive mechanism by stimulating the renin-angiotensin-aldosterone system (RAAS) [17].
Nox4 is a hydrogen peroxide-producing NADPH oxidase isoform highly expressed in the kidney. NAD(P)H oxidase was recognized as the major source of reactive oxygen species (ROS) contributing to salt-induced hypertension [18]. Dahl salt-sensitive hypertensive rats with knockout of Nox4 exhibited a reduced renal injury and attenuated BP response to high salt [19]. Thus, we may suggest that the increased transcription of Nox4 in the renal medulla of ISIAH rats may contribute to increased rate of ROS production and hypertension development.
Superoxide dismutases (SODs) convert superoxide to hydrogen peroxide, which is then removed by glutathione peroxidase or catalase [20]. The oxidative stress appears to be a common feature of hypertensive disorders from diverse origins [21], and it has been long known that oxidative stress induces or enhances the activity of SODs [22]. Thus, the enhanced expression of Sod3 provides a rationale to suggest the presence of the oxidative stress in the renal medulla of ISIAH rats which may require the enhanced activity of SOD3 to prevent the formation of highly aggressive ROS.
Nos3 is known as related to the diversity of biological processes. In the current research, Nos3 was related to GO term groups such as regulation of BP, oxidation reduction, response to stress and to steroid hormone stimulus, regulation of calcium and sodium ion transport, and angiogenesis (Additional file 5). Endothelial nitric oxide synthase (eNOS; NOS3) is physiologically important for vascular homeostasis [23]. It is expressed predominantly in the endothelium of blood vessels where it catalyzes the production of NO. NO regulates vascular tone and local blood flow, platelet aggregation and adhesion, and leukocyte-endothelial cell interactions. Abnormalities in NO production by the vascular endothelium result in endothelial dysfunction, which occurs in hypertension, diabetes, aging, and as a prelude to atherosclerosis [24].
Nos3 transcription was found to be reduced in the renal medulla of ISIAH rats. The reduced eNOS expression was earlier reported for different models of hypertension: spontaneously hypertensive rats (SHR) [25], Dahl salt-sensitive rats [26], two-kidney, one-clip hypertensive rats [27]. The reduced level of eNOS and NO production in the renal medulla is associated with a reduction in medullary blood flow and have considerable importance in sodium and water homeostasis and the long-term control of arterial pressure [27–29]. Recently, the reduced renal blood flow measured by magnetic resonance angiography was reported in ISIAH rats as compared to normotensive Wistar rats [30]. So, the reduced transcription of the Nos3 in the renal medulla of ISIAH rats may be considered as a key feature possibly leading to the medullary blood flow reduction. This, and the decreased NO availability in serum of ISIAH rats and its negative correlation with the BP reported earlier [31], might indicate the impact of this mechanism on stress-sensitive hypertension development.
One more gene which may participate in regulation of NO availability in the renal medulla of ISIAH rats is Ephx2 encoding the soluble epoxide hydrolase (sEH). In the current study, Ephx2 was one of the DEGs associated with hypertension and showing the highest differences in expression in ISIAH and WAG renal medulla. sEH metabolizes the epoxyeicosatrienoic acids (EETs) having antihypertensive properties. EETs also possess anti-inflammatory actions that could protect the kidney vasculature from injury during renal and cardiovascular diseases [32]. It was shown that P450 eicosanoids are vasodilatory, largely through their ability to activate eNOS and NO release [33]. sEH is a main effector of angiotensin II-induced hypertension [34]. Ephx2 was also recognized as a gatekeeper gene contributing to programmed hypertension [35]. In the current study, the PLS regression method, which is commonly used for biomarker selection in metabolomic [36] and gene expression [37] studies, reckoned Ephx2 as one of the genes making the most significant contribution to the inter-strain differences. All this and the increased transcription of Ephx2 found earlier in hypothalamus [38] and renal cortex [9] of ISIAH rats prompt us to consider Ephx2 as a key candidate for further studies of the mechanisms underlying the stress-sensitive hypertension in ISIAH rats. The sEH was already considered as a suitable target for pharmaceutical intervention in the hypertension treatment [39].
The other groups of DEGs, being among the most significantly enriched and described by GO terms ‘response to external stimulus’, ‘response to hormone (endogenous) stimulus’, ‘response to stress’, and ‘homeostatic process’, provide the molecular basis for integrated responses to homeostasis disturbances in the renal medulla of the ISIAH rats (Additional file 5). Most DEGs associated with BP regulation (Adipoq, Adra1b, Agtr1a, Aqp2, Ephx2, Guca2b, Mif, Nos3, P2rx4, Tacr3) were related to these groups. As it may be seen from the following discussion, some of these genes may contribute to vascular tone and renal blood flow regulation.
Adiponectin (Adipoq gene) stimulates production of nitric oxide in vascular endothelial cells [40] and possess anti-atherogenic properties [41]. The increased expression of Adipoq in renal medulla of ISIAH rats may be protective against the reduction in medullary blood flow.
Agtr1a (angiotensin II receptor, type 1a) encodes the receptor for angiotensin II, which is the main effector of RAAS [42]. AT1A receptors expressed on the renal vasculature and/or renal tubular epithelia play a critical role in sodium and volume homeostasis [43]. AGTR1a is required for mineralocorticoid receptor stimulation to induce vascular remodeling, inflammation and endothelial dysfunction [44]. It was shown that in the euvolemic state the mean arterial pressure was significantly lower in the AT1A receptor-deficient mice compared with wild-type controls [45]. So, the reduced transcription of Agtr1a in ISIAH renal medulla may be considered as a compensatory mechanism against the complications mentioned above.
The alpha1B-adrenoceptors (Adra1b) are involved in blood vessel remodeling [46] and mediate the vasoconstrictor actions of the renal sympathetic nerves in rats with renal failure [47, 48]. Adra1b is one of the sympathetic nervous system (SNS) components [49]. It is known that increased renal SNS activity reduces urinary sodium and water excretion, renal blood flow and glomerular filtration rate [49]. Thus, the reduced transcription of Adra1b in ISIAH renal medulla may be considered as a protective mechanism against the excessive effects of SNS activation reported earlier in ISIAH rats [50].
One more DEG, which may cause the differences in SNS activity in ISIAH and WAG rats is Comt (catechol-O-methyltransferase), which encodes the enzyme involved in the degradation of catecholamines. The inhibition of COMT induces dopamine-dependent natriuresis [51]. The Comt-gene-disrupted mice were resistant to salt-induced hypertension [52]. So, the decreased expression of Comt in the renal medulla of ISIAH rats may lead to increase in renal dopaminergic effects and sodium excretion, and may be considered as an adaptive or protective mechanism for control of hypertension development in ISIAH rats. Earlier, the significantly decreased transcription of Comt was also detected in kidney of 6-month old ISIAH rats [53].
MIF (Macrophage migration inhibitory factor) possesses the ability to directly regulate the immunosuppressive actions of glucocorticoids and thus plays a critical role in the host control of inflammation and immunity [54]. MIF is a key factor in atherogenesis [55, 56]. The upregulation of the podocyte-expressed MIF induces an injury of podocytes and accelerates the progression of glomerulosclerosis [57]. In ISIAH rats, the upregulation of Mif gene was found both in renal cortex [9] and in renal medulla (the current study), and possibly may contribute to development of atherosclerosis as well as to glomerulo- and medullary sclerosis histologically determined earlier [7, 8].
The changes in expression of DEGs associated with renal hypertension (Agtr1a, Fn1, Gja1, Lama2, Mmp2, Mmp9, Nos3) may also be suggested to have a significant impact on disease development in ISIAH rats (Table 2).
Fibronectin 1 is a glycoprotein involved in cell–matrix and cell–cell adhesion [58]. Fibronectin 1 may be induced by different agents including angiotensin II [59] and aldosterone [44]. Both of these stimuli may lead to vascular remodeling and vascular inflammation [44, 60]. So, the reduction in Fn1 expression may be directed against the excessive vascular complications development in ISIAH renal medulla. It is worth also to mention that almost all (33 gene out of 37) of the multiple DEGs related to GO terms ‘cell adhesion’ and ‘cell–matrix adhesion’ were downregulated in ISIAH renal medulla (Additional file 5), and it is of high probability that those changes, at least in a part, are directed towards the prevention of vascular complications.
The protein encoded by Gja1 (or Cx43, connexin 43) is a component of gap junctions, which permit the passage of ions and chemical mediators from cell to cell [61]. The expression of connexins in renal arterioles is believed to have a profound impact on conducted responses, regulation of arteriolar tonus and renal blood flow. Besides, the cell-to-cell communication mediated by Cx43 channels may contribute to regulating the elasticity of the vascular wall [62]. However, no evidence for an increased abundance of Cx43 in renal arterioles of SHR when compared with normotensive counterparts was found [63]. In the ISIAH renal medulla the increased transcriptional level of Gja1 was detected. Earlier a differential regulation of aortic Cx43 in different models of hypertension was reported [62], and this may also be true for kidney. This may be one of the features distinguishing the mechanism of hypertension development in ISIAH rats from that in SHR.
LAMA2 is an extracellular matrix protein [64] involved in regulation of cell adhesion. Matrix metalloproteinases (MMP2 and MMP9) belong to a family of metalloendopeptidases that cleave the protein components of the extracellular matrix and thereby play a central role in tissue remodeling [65]. MMP2 (Mmp2) and MMP9 (Mmp9) are involved in the vascular smooth muscle cell activation and neointimal formation that characterize arterial tissue remodeling after injury [66]. The overexpression of MMP9 and MMP2 have been observed within plaques [67]. The elevated urine values of MMP-9 was recognized as a marker of atherosclerotic disease [68], and, alternatively, the loss of MMP9 reduces atherosclerotic burden [69]. MMP-2 also contributes to the development of atherosclerosis [70]. Taking that into consideration, the decrease in Mmp2 and Mmp9 expression may be considered as protective against vascular remodeling in ISIAH renal medulla.
So, in ISIAH renal medulla, several DEGs associated with renal hypertension are related to cell–matrix and cell–cell adhesion processes. It is known that in hypertension the increase in arterial stretch stimulates vessel thickening to normalize the tensile forces. This process requires modification of the extracellular matrix and of cell–matrix interactions [71]. The results of the current study are in a good agreement with this conception and highlighted the major DEGs contributing to the modification of the extracellular matrix and of cell–matrix interactions in renal medulla of rats with stress-sensitive hypertension.
Multiple genes with the changed level of transcription related to ion transport and to regulation of ion transport were found in the renal medulla of ISIAH rats. Many of these genes should be essential for maintaining the sodium and water balance in hypertensive state. However, it is not possible to discuss the role of all the DEGs contributing to these processes within the current paper. We just would like to mention that all the processes including such a basic one as sodium and water balance may undergo the strain-specific regulation. This may be seen, for example, in the case of the Aqp2 being regulated by vasopressin [72] and playing a role in body water balance [73]. Mice with reduced expression of Aqp2 have severe polyuria and very low urine osmolality [74]. Aqp2 transcription was reduced in renal medulla of ISIAH rats. The reduced AQP2 in the renal medulla was reported earlier in the clipped kidney of rats with experimental two-kidney, one clip hypertension [75] and in Milan hypertensive rats [76]. However, the medullary expression of AQP2 protein was increased in DOCA-salt hypertensive rats [77] and in SHR [78]. The existence of the strain-specific differences in the molecular mechanisms of the hypertension development points out the importance of the search for the common features contributing to disease manifestation which may be potentially used as new pharmacological targets.
Conclusions
The current study helped to identify a number of DEGs specifying the function of renal medulla in ISIAH rats being a model of the stress-sensitive arterial hypertension. The genes already known as associated with hypertension development were mostly discussed. However, the other genes also may have indirect influence on disease development and maintenance. The discussion demonstrated that the changes in expression of many genes may be considered as protective or adaptive and directed toward the integration and coordination of the general homeostasis of the organism. Several DEGs, which may modulate the renal medulla blood flow were detected. The reduced transcription of the Nos3 points to the possible reduction of the blood flow in the renal medulla of ISIAH rats.
The generated data may be useful for comparison with those from different models of hypertension and for identifying the common molecular determinants contributing to disease manifestation which may be potentially used as new pharmacological targets. Based on discussion, Ephx2 seems to be a promising candidate for further studies of its potential as therapeutic target.
Methods
Animals
The hypertensive ISIAH/Icgn and normotensive WAG/GSto-Icgn rat strains were used in the current study. Rats were bred in the Center for Genetic Resources of Laboratory Animals at the Institute of Cytology and Genetics, Siberian Branch of the Russian Academy of Sciences, (Novosibirsk, Russia, RFMEFI61914X0005 and RFMEFI62114X0010). All rats were kept in standard laboratory conditions and had free access to food and water. The 3-month old ISIAH, and WAG male rats were used in RNA-seq experiments. Each group of experimental animals contained 3 rats. Rats were individually caged one week before the measurement of the systolic arterial BP, which was measured indirectly by the tail-cuff method as it was described earlier [9]. The mean systolic arterial BP was 171.7 ± 1.22 mmHg in the experimental group of ISIAH males and 116.33 ± 1.86 mmHg in WAG. Six days after BP measurement, rats were decapitated and their kidneys were rapidly removed and sectioned. The samples of renal medulla were stored in RNA Later (Qiagen, Chatsworth, CA) at −70 °C until use. The animal experiments were conducted with approval of the Institute’s Animal Care and Use Committee.
RNA-seq analysis
The samples of renal medulla were sent to JSC Genoanalytica (Moscow, Russia), where the technological part of the RNA-seq analysis was conducted as it was described earlier {9}. Dynabeads mRNA Purification Kit (Ambion, USA) was used for mRNA extraction and NEBNext mRNA Library Prep Reagent Set for Illumina (NEB, USA) was used for cDNA libraries construction. All kits were used according to the manufacturer’s protocol. cDNA libraries were sequenced using a HiSeq1500 Sequencing System (Illumina Sequencing, San Diego, USA) in a single end mode with a read length of 50 bases. Three biological replicates were analyzed for each rat strain. The sequencing data were subjected to adapter trimming and low-quality sequence removal and mapped to the rat reference genome (RGSC Rnor_5.0, rn5) with the use of Tophat2 [79]. Quality metrics of the mapped libraries (Additional file 7) were collected using the Picard ‘CollectRnaSeqMetrics’ tool (http://broadinstitute.github.io/picard/). The differential expression analysis was performed using the Cufflinks workflow [80]. A gene was defined as being expressed if it matched the Cufflinks criteria on suitability for statistical testing (test status ‘OK’). Genes with a Benjamini-Hochberg adjusted p-value (q-value) <0.05 were considered to be differentially expressed. Heatmap of the genes differentially expressed (q value < 0.01) in the renal medulla of the ISIAH and WAG rats was built using the core functions of R statistical language (https://cran.r-project.org/); a hierarchical ‘complete linkage’ clustering by Euclidean distance was used to construct the dendrograms. The raw RNA-Seq data are available at the NCBI Short Read Archive database under the Accession number: PRJNA299102.
Functional annotation
The DEGs were functionally annotated in DAVID (The Database for Annotation, Visualization and Integrated Discovery) (http://david.abcc.ncifcrf.gov/) [81, 82] with the use of Rattus norvegicus genome as the background list for the over-representation analysis. The significantly enriched biological processes (p < 0.05) were identified using the Gene Ontology option. The most significant to the data set pathways were identified in Kyoto Encyclopedia of Genes and Genomes Pathway Database (KEGG, http://www.genome.jp/kegg/). The annotation of DEGs in the Rat Genome Database Disease Portals (RGD, http://rgd.mcw.edu/wg/portals?100) was used to detect the genes related to arterial hypertension and renal diseases. The DEGs encoding the transcription factors were detected using the atlas of combinatorial transcriptional regulation in mouse and man [83], GenBank (http://www.ncbi.nlm.nih.gov/gene/), and Panther classification system (http://www.pantherdb.org/) [84].
Quantitative real-time PCR (qPCR)
To estimate the relative amount of target mRNA, qPCR analysis was performed on the renal medulla samples from ISIAH (n = 5) and WAG (n = 5) male rats aged as 3-month old. TRI reagent (Molecular research center, USA) was used for the extraction of the total RNA, and DNase I (Promega, USA) treatment was performed to remove the residual genomic DNA. The kits were used according to the manufacturer’s recommendations.
The protocol for the reverse transcription was the following: the reaction was carried out with 40 units of MoMLV (Vektor-Best, Russia) in 50 μl of reaction mixture, which contained reverse transcription (RT) buffer, 0.4 mM dNTPs, 0.25 nmol of random nonanucleotide primers (Biosan, Russia), and 3 μg of total RNA. The cDNA synthesis was performed at 37 °C (1 h), 42 °C (30 min), and 50 °C (10 min) with the following inactivation of the enzyme by heating at 75 °C for 5 min.
The reaction volume for qPCR was 20 μl. It contained a master mix with SYBR Green, 0.15 mM of each forward and reverse primers, 1 unit of HotStart Taq polymerase (Vektor-Best, Russia), and the cDNA template. The Rpl30 (ribosomal protein L30) stably expressed in different tissues from ISIAH and WAG rats was used as a reference gene. The primers sequences and characteristics are given in Additional file 8.
qPCR was performed in an iCycler iQ4 Real-Time PCR Detection System (Bio-Rad Laboratories, USA) using the protocol described earlier [9]. Relative transcript levels were determined by standard-curve quantitation method [85]. The mixture of the aliquots from all synthesized cDNA samples was used as a standard cDNA for calibration curves plotting. The value obtained for the target gene was normalized against the value for reference gene and then the relative mRNA abundance was calculated as a ratio of the normalized mRNA level in the experimental ISIAH samples to the normalized mRNA level in the samples from WAG rats. A value of 1 was assigned to the normalized mRNA level obtained for the samples from control WAG rats.
Statistical methods
Mann-Whitney U-test (Statistica v.8.0, Statsoft, USA) was used for calculations of the statistical significance for qPCR data. Statistical significance was set at p < 0.05. The data were expressed as M ± S.E.M. (means and their standard errors).
The acquired RNA-seq data (FPKM values) were log transformed, centered and normalized to run the partial-least squares discriminant analysis (PLS-DA). First, the scaling of the data sets was performed using principal coordinates method based on Euclidean metric distances. Then, the pattern of co-variation for linear combinations between two blocks of variables [86] was explored to get the separation of groups in PLS-DA. PLS-DA and the following calculation of Pearson correlation coefficients were performed to find a set of variables (expressed genes) that maximize the covariance between gene expression in ISIAH and WAG rat strains and fixed dummy matrix representing group membership [86] for WAG and ISIAH rats, correspondingly. As a result of these procedures the PLS-DA Axes maximizing the distances between hypertensive and normotensive rats were constructed, the correlation between gene expression and PLS-DA Axis 1 was calculated, and the genes contributing the most to differences between ISIAH and WAG transcriptomes were revealed.
Acknowledgements
The authors are grateful to JSC Genoanalytica (Moscow, Russia) for conducting the technological part of the experiment and the primary statistical analysis. This work has been supported by the Russian Science Foundation, grant No. 14-15-00118. The Siberian Branch of the Russian Academy of Sciences (SB RAS) Siberian Supercomputer Center is gratefully acknowledged for providing supercomputer facilities.
Declarations
This article has been published as part of BMC Genetics Vol 17 Suppl 3, 2016: Selected articles from BGRS\SB-2016: genetics. The full contents of the supplement are available online at https://bmcgenet.biomedcentral.com/articles/supplements/volume-17-supplement-3.
Funding
Publication of this article has been funded by the Russian Science Foundation (grant 14-15-00118).
Availability of data and materials
All data generated or analyzed during this study are included in this published article [and its supplementary information files]. The RNA-Seq data were deposited in the NCBI Short Read Archive database with Accession number: PRJNA299102.
Authors’ contributions
MR and LF performed quantitative real-time PCR, participated in interpretation of data, and helped to draft the manuscript; NE has made substantial contribution to bioinformatics analysis; VE has made substantial contributions to conception and use of multivariate statistical analysis; AM has made substantial contributions to conception and design of the study and participated in interpretation of data; OR participated in interpretation of data and drafted the manuscript. All authors read and approved the final version of the manuscript.
Competing interests
The authors declare that they have no competing interests. The authors alone are responsible for the content and writing of the paper.
Consent for publication
Not applicable.
Ethics approval and consent to participate
All animal experiments were approved by the Institute’s Animal Care and Use Committee.
Abbreviations
- 20-HETE
20-hydroxyeicosatetraenoic acid
- BP
Blood pressure
- DAVID
Database for Annotation, Visualization and Integrated Discovery
- DEGs
Differentially expressed genes
- EETs
Epoxyeicosatrienoic acids
- eNOS or NOS3
endothelial nitric oxide synthase
- FPKM
Fragments per kilobase of transcript per million mapped reads
- GO
Gene Ontology
- ISIAH
Inherited Stress-Induced Arterial Hypertension
- KEGG
Kyoto Encyclopedia of Genes and Genomes Pathway Database
- NO
Nitric oxide
- PLS-DA
Partial-least squares discriminant analysis
- qPCR
quantitative real time polymerase chain reaction
- RAAS
Renin-angiotensin-aldosterone system
- RGD
Rat Genome Database
- RNA-seq
RNA sequencing
- ROS
Reactive oxygen species
- SABP
Systolic arterial blood pressure
- sEH
soluble epoxide hydrolase
- SHR
Spontaneously hypertensive rats
- SNS
Sympathetic nervous system
- SODs
Superoxide dismutases
- WAG
Wistar Albino Glaxo rats
Additional files
Genes differentially expressed in renal medulla of hypertensive ISIAH and normotensive control WAG rats. (XLS 197 kb)
Heatmap of the differentially expressed genes in the renal medulla of the ISIAH and WAG rats. (PDF 50 kb)
Genes with detected expression in renal medulla of only one rat strain. (DOC 51 kb)
Functional annotation of differentially expressed genes (DEGs) found in 3-month old ISIAH and WAG renal medulla. (XLS 95 kb)
Differentially expressed genes related to Gene Ontlogy (GO) terms for biological processes in the renal medulla of 3-month old ISIAH and WAG male rats. (XLS 160 kb)
Metabolic pathways enriched with genes differentially expressed in ISIAH and WAG renal medulla. (XLS 41 kb)
The summary statistics for the sequenced libraries from renal medulla of ISIAH and WAG rats. (XLS 17 kb)
Primers used in real-time PCR. (DOC 33 kb)
Contributor Information
Marina A. Ryazanova, Email: ocean-2006@yandex.ru
Larisa A. Fedoseeva, Email: frumgarts@ngs.ru
Nikita I. Ershov, Email: nikotinmail@mail.ru
Vadim M. Efimov, Email: efimov@bionet.nsc.ru
Arcady L. Markel, Email: markel@bionet.nsc.ru
Olga E. Redina, Email: oredina@ngs.ru
References
- 1.Guyton AC. Dominant role of the kidneys and accessory role of whole-body autoregulation in the pathogenesis of hypertension. Am J Hypertens. 1989;2:575–585. doi: 10.1093/ajh/2.7.575. [DOI] [PubMed] [Google Scholar]
- 2.Cowley AWJ, Mattson DL, Lu S, Roman RJ. The renal medulla and hypertension. Hypertension. 1995;25:663–673. doi: 10.1161/01.HYP.25.4.663. [DOI] [PubMed] [Google Scholar]
- 3.Hall JE. The kidney, hypertension, and obesity. Hypertension. 2003;41:625–633. doi: 10.1161/01.HYP.0000052314.95497.78. [DOI] [PubMed] [Google Scholar]
- 4.Mullins LJ, Bailey MA, Mullins JJ. Hypertension, kidney, and transgenics: a fresh perspective. Physiol Rev. 2006;86:709–746. doi: 10.1152/physrev.00016.2005. [DOI] [PubMed] [Google Scholar]
- 5.Markel AL. Development of a new strain of rats with inherited stress-induced arterial hypertension. In: Sassard J, editor. Genetic hypertension. Paris: Colloque INSERM; 1992. pp. 405–407. [Google Scholar]
- 6.Markel AL, Maslova LN, Shishkina GT, Bulygina VV, Machanova NA, Jacobson GS. Developmental influences on blood pressure regulation in ISIAH rats. In: McCarty R, Blizard DA, Chevalier RL, editors. Development of the hypertensive phenotype: basic and clinical studies. Amsterdam- Lausanne- NewYork- Oxford- Shannon- Singapore- Tokyo: Elsevier; 1999. pp. 493–526. [Google Scholar]
- 7.Shmerling MD, Filiushina EE, Lazarev VA, Buzueva II, Markel’ AL, Iakobson GS. Ultrastructural changes of kidney corpuscles in rats with hereditary stress-induced arterial hypertension [Article in Russian] Morfologiia. 2001;120:70–74. [PubMed] [Google Scholar]
- 8.Filyushina EE, Shmerling MD, Buzueva II, Lazarev VA, Markel AL, Yakobson GS. Structural characteristics of renomedullary interstitial cells of hypertensive ISIAH rats. Bull Exp Biol Med. 2013;155:408–412. doi: 10.1007/s10517-013-2164-7. [DOI] [PubMed] [Google Scholar]
- 9.Fedoseeva LA, Ryazanova MA, Ershov NI, Markel AL, Redina OE. Comparative transcriptional profiling of renal cortex in rats with inherited stress-induced arterial hypertension and normotensive Wistar Albino Glaxo rats. BMC Genet. 2016;17(Suppl 1):12. doi: 10.1186/s12863-015-0306-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Miyata N, Cowley AWJ. Renal intramedullary infusion of L-arginine prevents reduction of medullary blood flow and hypertension in Dahl salt-sensitive rats. Hypertension. 1999;33:446–450. doi: 10.1161/01.HYP.33.1.446. [DOI] [PubMed] [Google Scholar]
- 11.O’Connor PM, Cowley AWJ. Modulation of pressure-natriuresis by renal medullary reactive oxygen species and nitric oxide. Curr Hypertens Rep. 2010;12:86–92. doi: 10.1007/s11906-010-0094-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Yu Y, Fuscoe JC, Zhao C, Guo C, Jia M, Qing T, Bannon DI, Lancashire L, Bao W, Du T, Luo H, Su Z, Jones WD, Moland CL, Branham WS, Qian F, Ning B, Li Y, Hong H, Guo L, Mei N, Shi T, Wang KY, Wolfinger RD, Nikolsky Y, Walker SJ, Duerksen-Hughes P, Mason CE, Tong W, Thierry-Mieg J, Thierry-Mieg D, Shi L, Wang C. A rat RNA-Seq transcriptomic BodyMap across 11 organs and 4 developmental stages. Nat Commun. 2014;5:3230. doi: 10.1038/ncomms4230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Cowley AWJ, Abe M, Mori T, O’Connor PM, Ohsaki Y, Zheleznova NN. Reactive oxygen species as important determinants of medullary flow, sodium excretion, and hypertension. Am J Physiol Renal Physiol. 2015;308:F179–F197. doi: 10.1152/ajprenal.00455.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Agbor LN, Walsh MT, Boberg JR, Walker MK. Elevated blood pressure in cytochrome P4501A1 knockout mice is associated with reduced vasodilation to omega-3 polyunsaturated fatty acids. Toxicol Appl Pharmacol. 2012;264:351–360. doi: 10.1016/j.taap.2012.09.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Agbor LN, Wiest EF, Rothe M, Schunck WH, Walker MK. Role of CYP1A1 in modulating the vascular and blood pressure benefits of omega-3 polyunsaturated fatty acids. J Pharmacol Exp Ther. 2014;351:688–698. doi: 10.1124/jpet.114.219535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kalsotra A, Cui X, Anakk S, Hinojos CA, Doris PA, Strobel HW. Renal localization, expression, and developmental regulation of P450 4 F cytochromes in three substrains of spontaneously hypertensive rats. Biochem Biophys Res Commun. 2005;338:423–431. doi: 10.1016/j.bbrc.2005.08.101. [DOI] [PubMed] [Google Scholar]
- 17.Wu CC, Gupta T, Garcia V, Ding Y, Schwartzman ML. 20-HETE and blood pressure regulation: clinical implications. Cardiol Rev. 2014;22:1–12. doi: 10.1097/CRD.0b013e3182961659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Taylor NE, Glocka P, Liang M, Cowley AWJ. NADPH oxidase in the renal medulla causes oxidative stress and contributes to salt-sensitive hypertension in Dahl S rats. Hypertension. 2006;47:692–698. doi: 10.1161/01.HYP.0000203161.02046.8d. [DOI] [PubMed] [Google Scholar]
- 19.Cowley AWJ, Yang C, Zheleznova NN, Staruschenko A, Kurth T, Rein L, Kumar V, Sadovnikov K, Dayton A, Hoffman M, Ryan RP, Skelton MM, Salehpour F, Ranji M, Geurts A. Evidence of the Importance of Nox4 in Production of Hypertension in Dahl Salt-Sensitive Rats. Hypertension. 2016;67:440–450. doi: 10.1161/HYPERTENSIONAHA.115.06280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Afonso V, Champy R, Mitrovic D, Collin P, Lomri A. Reactive oxygen species and superoxide dismutases: role in joint diseases. Joint Bone Spine. 2007;74:324–329. doi: 10.1016/j.jbspin.2007.02.002. [DOI] [PubMed] [Google Scholar]
- 21.Uddin M, Yang H, Shi M, Polley-Mandal M, Guo Z. Elevation of oxidative stress in the aorta of genetically hypertensive mice. Mech Ageing Dev. 2003;124:811–817. doi: 10.1016/S0047-6374(03)00135-0. [DOI] [PubMed] [Google Scholar]
- 22.Scandalios JG. Oxygen Stress and Superoxide Dismutases. Plant Physiol. 1993;101:7–12. doi: 10.1104/pp.101.1.7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Li H, Forstermann U. Nitric oxide in the pathogenesis of vascular disease. J Pathol. 2000;190:244–254. doi: 10.1002/(SICI)1096-9896(200002)190:3<244::AID-PATH575>3.0.CO;2-8. [DOI] [PubMed] [Google Scholar]
- 24.Huang PL. Endothelial nitric oxide synthase and endothelial dysfunction. Curr Hypertens Rep. 2003;5:473–480. doi: 10.1007/s11906-003-0055-4. [DOI] [PubMed] [Google Scholar]
- 25.Zhou XJ, Vaziri ND, Zhang J, Wang HW, Wang XQ. Association of renal injury with nitric oxide deficiency in aged SHR: prevention by hypertension control with AT1 blockade. Kidney Int. 2002;62:914–921. doi: 10.1046/j.1523-1755.2002.00516.x. [DOI] [PubMed] [Google Scholar]
- 26.Yoshihara F, Suga S, Yasui N, Horio T, Tokudome T, Nishikimi T, Kawano Y, Kangawa K. Chronic administration of adrenomedullin attenuates the hypertension and increases renal nitric oxide synthase in Dahl salt-sensitive rats. Regul Pept. 2005;128:7–13. doi: 10.1016/j.regpep.2004.12.028. [DOI] [PubMed] [Google Scholar]
- 27.Wickman A, Andersson IJ, Jia J, Hedin L, Bergstrom G. Endothelial nitric oxide synthase protein is reduced in the renal medulla of two-kidney, one-clip hypertensive rats. J Hypertens. 2001;19:1665–1673. doi: 10.1097/00004872-200109000-00020. [DOI] [PubMed] [Google Scholar]
- 28.Cowley AWJ, Mori T, Mattson D, Zou AP. Role of renal NO production in the regulation of medullary blood flow. Am J Physiol Regul Integr Comp Physiol. 2003;284:R1355–R1369. doi: 10.1152/ajpregu.00701.2002. [DOI] [PubMed] [Google Scholar]
- 29.Mount PF, Power DA. Nitric oxide in the kidney: functions and regulation of synthesis. Acta Physiol (Oxf) 2006;187:433–446. doi: 10.1111/j.1748-1716.2006.01582.x. [DOI] [PubMed] [Google Scholar]
- 30.Seryapina AA, Shevelev OB, Moshkin MP, Markel’ AL. Parameters of Blood Flow in Great Arteries in Hypertensive ISIAH Rats with Stress-Dependent Arterial Hypertension. Bull Exp Biol Med. 2016;161:468–471. doi: 10.1007/s10517-016-3440-0. [DOI] [PubMed] [Google Scholar]
- 31.Bobko AA, Sergeeva SV, Bagryanskaya EG, Markel AL, Khramtsov VV, Reznikov VA, Kolosova NG. 19 F NMR measurements of NO production in hypertensive ISIAH and OXYS rats. Biochem Biophys Res Commun. 2005;330:367–370. doi: 10.1016/j.bbrc.2005.02.166. [DOI] [PubMed] [Google Scholar]
- 32.Imig JD. Epoxide hydrolase and epoxygenase metabolites as therapeutic targets for renal diseases. Am J Physiol Renal Physiol. 2005;289:F496–F503. doi: 10.1152/ajprenal.00350.2004. [DOI] [PubMed] [Google Scholar]
- 33.Hercule HC, Schunck WH, Gross V, Seringer J, Leung FP, Weldon SM, da Costa Goncalves AC, Huang Y, Luft FC, Gollasch M. Interaction between P450 eicosanoids and nitric oxide in the control of arterial tone in mice. Arterioscler Thromb Vasc Biol. 2009;29:54–60. doi: 10.1161/ATVBAHA.108.171298. [DOI] [PubMed] [Google Scholar]
- 34.Jung O, Brandes RP, Kim IH, Schweda F, Schmidt R, Hammock BD, Busse R, Fleming I. Soluble epoxide hydrolase is a main effector of angiotensin II-induced hypertension. Hypertension. 2005;45:759–765. doi: 10.1161/01.HYP.0000153792.29478.1d. [DOI] [PubMed] [Google Scholar]
- 35.Tain YL, Huang LT, Chan JY, Lee CT. Transcriptome analysis in rat kidneys: importance of genes involved in programmed hypertension. Int J Mol Sci. 2015;16:4744–4758. doi: 10.3390/ijms16034744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Szymanska E, Saccenti E, Smilde AK, Westerhuis JA. Double-check: validation of diagnostic statistics for PLS-DA models in metabolomics studies. Metabolomics. 2012;8(Suppl 1):3–16. doi: 10.1007/s11306-011-0330-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Perez-Enciso M, Tenenhaus M. Prediction of clinical outcome with microarray data: a partial least squares discriminant analysis (PLS-DA) approach. Hum Genet. 2003;112:581–592. doi: 10.1007/s00439-003-0921-9. [DOI] [PubMed] [Google Scholar]
- 38.Klimov LO, Ershov NI, Efimov VM, Markel AL, Redina OE. Genome-wide transcriptome analysis of hypothalamus in rats with inherited stress-induced arterial hypertension. BMC Genet. 2016;17(Suppl 1):13. doi: 10.1186/s12863-015-0307-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Chiamvimonvat N, Ho CM, Tsai HJ, Hammock BD. The soluble epoxide hydrolase as a pharmaceutical target for hypertension. J Cardiovasc Pharmacol. 2007;50:225–237. doi: 10.1097/FJC.0b013e3181506445. [DOI] [PubMed] [Google Scholar]
- 40.Chen H, Montagnani M, Funahashi T, Shimomura I, Quon MJ. Adiponectin stimulates production of nitric oxide in vascular endothelial cells. J Biol Chem. 2003;278:45021–45026. doi: 10.1074/jbc.M307878200. [DOI] [PubMed] [Google Scholar]
- 41.Ouchi N, Kobayashi H, Kihara S, Kumada M, Sato K, Inoue T, Funahashi T, Walsh K. Adiponectin stimulates angiogenesis by promoting cross-talk between AMP-activated protein kinase and Akt signaling in endothelial cells. J Biol Chem. 2004;279:1304–1309. doi: 10.1074/jbc.M310389200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Fyhrquist F, Saijonmaa O. Renin-angiotensin system revisited. J Intern Med. 2008;264:224–236. doi: 10.1111/j.1365-2796.2008.01981.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Oliverio MI, Best CF, Smithies O, Coffman TM. Regulation of sodium balance and blood pressure by the AT(1A) receptor for angiotensin II. Hypertension. 2000;35:550–554. doi: 10.1161/01.HYP.35.2.550. [DOI] [PubMed] [Google Scholar]
- 44.Briet M, Barhoumi T, Mian MO, Coelho SC, Ouerd S, Rautureau Y, Coffman TM, Paradis P, Schiffrin EL. Aldosterone-Induced Vascular Remodeling and Endothelial Dysfunction Require Functional Angiotensin Type 1a Receptors. Hypertension. 2016;67:897–905. doi: 10.1161/HYPERTENSIONAHA.115.07074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Cervenka L, Mitchell KD, Oliverio MI, Coffman TM, Navar LG. Renal function in the AT1A receptor knockout mouse during normal and volume-expanded conditions. Kidney Int. 1999;56:1855–1862. doi: 10.1046/j.1523-1755.1999.00757.x. [DOI] [PubMed] [Google Scholar]
- 46.Zhang H, Cotecchia S, Thomas SA, Tanoue A, Tsujimoto G, Faber JE. Gene deletion of dopamine beta-hydroxylase and alpha1-adrenoceptors demonstrates involvement of catecholamines in vascular remodeling. Am J Physiol Heart Circ Physiol. 2004;287:H2106–H2114. doi: 10.1152/ajpheart.00290.2004. [DOI] [PubMed] [Google Scholar]
- 47.Khan AH, Sattar MA, Abdullah NA, Johns EJ. Influence of cisplatin-induced renal failure on the alpha(1)-adrenoceptor subtype causing vasoconstriction in the kidney of the rat. Eur J Pharmacol. 2007;569:110–118. doi: 10.1016/j.ejphar.2007.04.063. [DOI] [PubMed] [Google Scholar]
- 48.Hye Khan MA, Sattar MA, Abdullah NA, Johns EJ. Influence of combined hypertension and renal failure on functional alpha(1)-adrenoceptor subtypes in the rat kidney. Br J Pharmacol. 2008;153:1232–1241. doi: 10.1038/bjp.2008.13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Zhu H, Poole J, Lu Y, Harshfield GA, Treiber FA, Snieder H, Dong Y. Sympathetic nervous system, genes and human essential hypertension. Curr Neurovasc Res. 2005;2:303–317. doi: 10.2174/156720205774322575. [DOI] [PubMed] [Google Scholar]
- 50.Markel AL, Redina OE, Gilinsky MA, Dymshits GM, Kalashnikova EV, Khvorostova YV, Fedoseeva LA, Jacobson GS. Neuroendocrine profiling in inherited stress-induced arterial hypertension rat strain with stress-sensitive arterial hypertension. J Endocrinol. 2007;195:439–450. doi: 10.1677/JOE-07-0254. [DOI] [PubMed] [Google Scholar]
- 51.Eklof AC, Holtback U, Sundelof M, Chen S, Aperia A. Inhibition of COMT induces dopamine-dependent natriuresis and inhibition of proximal tubular Na+, K + −ATPase. Kidney Int. 1997;52:742–747. doi: 10.1038/ki.1997.390. [DOI] [PubMed] [Google Scholar]
- 52.Helkamaa T, Mannisto PT, Rauhala P, Cheng ZJ, Finckenberg P, Huotari M, Gogos JA, Karayiorgou M, Mervaala EM. Resistance to salt-induced hypertension in catechol-O-methyltransferase-gene-disrupted mice. J Hypertens. 2003;21:2365–2374. doi: 10.1097/00004872-200312000-00026. [DOI] [PubMed] [Google Scholar]
- 53.Redina OE, Smolenskaya SE, Abramova TO, Ivanova LN, Markel AL. Differential transcriptional activity of kidney genes in hypertensive ISIAH and normotensive WAG rats. Clin Exp Hypertens. 2015;37:249–259. doi: 10.3109/10641963.2014.954711. [DOI] [PubMed] [Google Scholar]
- 54.Flaster H, Bernhagen J, Calandra T, Bucala R. The macrophage migration inhibitory factor-glucocorticoid dyad: regulation of inflammation and immunity. Mol Endocrinol. 2007;21:1267–1280. doi: 10.1210/me.2007-0065. [DOI] [PubMed] [Google Scholar]
- 55.Bernhagen J, Krohn R, Lue H, Gregory JL, Zernecke A, Koenen RR, Dewor M, Georgiev I, Schober A, Leng L, Kooistra T, Fingerle-Rowson G, Ghezzi P, Kleemann R, McColl SR, Bucala R, Hickey MJ, Weber C. MIF is a noncognate ligand of CXC chemokine receptors in inflammatory and atherogenic cell recruitment. Nat Med. 2007;13:587–596. doi: 10.1038/nm1567. [DOI] [PubMed] [Google Scholar]
- 56.Santos LL, Morand EF. Macrophage migration inhibitory factor: a key cytokine in RA, SLE and atherosclerosis. Clin Chim Acta. 2009;399:1–7. doi: 10.1016/j.cca.2008.09.014. [DOI] [PubMed] [Google Scholar]
- 57.Sasaki S, Nishihira J, Ishibashi T, Yamasaki Y, Obikane K, Echigoya M, Sado Y, Ninomiya Y, Kobayashi K. Transgene of MIF induces podocyte injury and progressive mesangial sclerosis in the mouse kidney. Kidney Int. 2004;65:469–481. doi: 10.1111/j.1523-1755.2004.00394.x. [DOI] [PubMed] [Google Scholar]
- 58.Yamada KM, Olden K. Fibronectins—adhesive glycoproteins of cell surface and blood. Nature. 1978;275:179–184. doi: 10.1038/275179a0. [DOI] [PubMed] [Google Scholar]
- 59.Pan Z, Zhao W, Zhang X, Wang B, Wang J, Sun X, Liu X, Feng S, Yang B, Lu Y. Scutellarin alleviates interstitial fibrosis and cardiac dysfunction of infarct rats by inhibiting TGF?1 expression and activation of p38-MAPK and ERK1/2. Br J Pharmacol. 2011;162:688–700. doi: 10.1111/j.1476-5381.2010.01070.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Touyz RM, Tabet F, Schiffrin EL. Redox-dependent signalling by angiotensin II and vascular remodelling in hypertension. Clin Exp Pharmacol Physiol. 2003;30:860–866. doi: 10.1046/j.1440-1681.2003.03930.x. [DOI] [PubMed] [Google Scholar]
- 61.Fishman GI, Spray DC, Leinwand LA. Molecular characterization and functional expression of the human cardiac gap junction channel. J Cell Biol. 1990;111:589–598. doi: 10.1083/jcb.111.2.589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Haefliger JA, Meda P. Chronic hypertension alters the expression of Cx43 in cardiovascular muscle cells. Braz J Med Biol Res. 2000;33:431–438. doi: 10.1590/S0100-879X2000000400009. [DOI] [PubMed] [Google Scholar]
- 63.Braunstein TH, Sorensen CM, Holstein-Rathlou NH. Connexin abundance in resistance vessels from the renal microcirculation in normo- and hypertensive rats. APMIS. 2009;117:268–276. doi: 10.1111/j.1600-0463.2009.02432.x. [DOI] [PubMed] [Google Scholar]
- 64.Masaki T, Matsumura K, Hirata A, Yamada H, Hase A, Arai K, Shimizu T, Yorifuji H, Motoyoshi K, Kamakura K. Expression of dystroglycan and the laminin-alpha 2 chain in the rat peripheral nerve during development. Exp Neurol. 2002;174:109–117. doi: 10.1006/exnr.2001.7856. [DOI] [PubMed] [Google Scholar]
- 65.Stamenkovic I. Extracellular matrix remodelling: the role of matrix metalloproteinases. J Pathol. 2003;200:448–464. doi: 10.1002/path.1400. [DOI] [PubMed] [Google Scholar]
- 66.Bendeck MP, Zempo N, Clowes AW, Galardy RE, Reidy MA. Smooth muscle cell migration and matrix metalloproteinase expression after arterial injury in the rat. Circ Res. 1994;75:539–545. doi: 10.1161/01.RES.75.3.539. [DOI] [PubMed] [Google Scholar]
- 67.Pasterkamp G, Schoneveld AH, Hijnen DJ, de Kleijn DP, Teepen H, van der Wal AC, Borst C. Atherosclerotic arterial remodeling and the localization of macrophages and matrix metalloproteases 1, 2 and 9 in the human coronary artery. Atherosclerosis. 2000;150:245–253. doi: 10.1016/S0021-9150(99)00371-8. [DOI] [PubMed] [Google Scholar]
- 68.Fitzsimmons PJ, Forough R, Lawrence ME, Gantt DS, Rajab MH, Kim H, Weylie B, Spiekerman AM, Dehmer GJ. Urinary levels of matrix metalloproteinase 9 and 2 and tissue inhibitor of matrix metalloproteinase in patients with coronary artery disease. Atherosclerosis. 2007;194:196–203. doi: 10.1016/j.atherosclerosis.2006.07.027. [DOI] [PubMed] [Google Scholar]
- 69.Luttun A, Lutgens E, Manderveld A, Maris K, Collen D, Carmeliet P, Moons L. Loss of matrix metalloproteinase-9 or matrix metalloproteinase-12 protects apolipoprotein E-deficient mice against atherosclerotic media destruction but differentially affects plaque growth. Circulation. 2004;109:1408–1414. doi: 10.1161/01.CIR.0000121728.14930.DE. [DOI] [PubMed] [Google Scholar]
- 70.Kuzuya M, Nakamura K, Sasaki T, Cheng XW, Itohara S, Iguchi A. Effect of MMP-2 deficiency on atherosclerotic lesion formation in apoE-deficient mice. Arterioscler Thromb Vasc Biol. 2006;26:1120–1125. doi: 10.1161/01.ATV.0000218496.60097.e0. [DOI] [PubMed] [Google Scholar]
- 71.Lemarie CA, Tharaux PL, Lehoux S. Extracellular matrix alterations in hypertensive vascular remodeling. J Mol Cell Cardiol. 2010;48:433–439. doi: 10.1016/j.yjmcc.2009.09.018. [DOI] [PubMed] [Google Scholar]
- 72.Kortenoeven ML, Fenton RA. Renal aquaporins and water balance disorders. Biochim Biophys Acta. 2014;1840:1533–1549. doi: 10.1016/j.bbagen.2013.12.002. [DOI] [PubMed] [Google Scholar]
- 73.Moeller HB, Fuglsang CH, Fenton RA. Renal aquaporins and water balance disorders. Best Pract Res Clin Endocrinol Metab. 2016;30:277–288. doi: 10.1016/j.beem.2016.02.012. [DOI] [PubMed] [Google Scholar]
- 74.Yang B, Zhao D, Qian L, Verkman AS. Mouse model of inducible nephrogenic diabetes insipidus produced by floxed aquaporin-2 gene deletion. Am J Physiol Renal Physiol. 2006;291:F465–F472. doi: 10.1152/ajprenal.00494.2005. [DOI] [PubMed] [Google Scholar]
- 75.Lee J, Oh Y, Kim SW. Altered renal expression of aquaporin-2 water channels in rats with experimental two-kidney, one clip hypertension. J Korean Med Sci. 2001;16:462–466. doi: 10.3346/jkms.2001.16.4.462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Procino G, Romano F, Torielli L, Ferrari P, Bianchi G, Svelto M, Valenti G. Altered expression of renal aquaporins and alfa-adducin polymorphisms may contribute to the establishment of salt-sensitive hypertension. Am J Hypertens. 2011;24:822–828. doi: 10.1038/ajh.2011.47. [DOI] [PubMed] [Google Scholar]
- 77.Lee J, Kang DG, Kim Y. Increased expression and shuttling of aquaporin-2 water channels in the kidney in DOCA-salt hypertensive rats. Clin Exp Hypertens. 2000;22:531–541. doi: 10.1081/CEH-100100089. [DOI] [PubMed] [Google Scholar]
- 78.Lee J, Kim S, Kim J, Jeong MH, Oh Y, Choi KC. Increased expression of renal aquaporin water channels in spontaneously hypertensive rats. Kidney Blood Press Res. 2006;29:18–23. doi: 10.1159/000092483. [DOI] [PubMed] [Google Scholar]
- 79.Kim D, Pertea G, Trapnell C, Pimentel H, Kelley R, Salzberg SL. TopHat2: accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013;14:R36. doi: 10.1186/gb-2013-14-4-r36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Trapnell C, Hendrickson DG, Sauvageau M, Goff L, Rinn JL, Pachter L. Differential analysis of gene regulation at transcript resolution with RNA-seq. Nat Biotechnol. 2013;31:46–53. doi: 10.1038/nbt.2450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Huang DW, Sherman BT, Lempicki RA. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat Protoc. 2009;4:44–57. doi: 10.1038/nprot.2008.211. [DOI] [PubMed] [Google Scholar]
- 82.Huang DW, Sherman BT, Lempicki RA. Bioinformatics enrichment tools: paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009;37:1–13. doi: 10.1093/nar/gkn923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Ravasi T, Suzuki H, Cannistraci CV, Katayama S, Bajic VB, Tan K, Akalin A, Schmeier S, Kanamori-Katayama M, Bertin N, Carninci P, Daub CO, Forrest AR, Gough J, Grimmond S, Han JH, Hashimoto T, Hide W, Hofmann O, Kamburov A, Kaur M, Kawaji H, Kubosaki A, Lassmann T, van Nimwegen E, MacPherson CR, Ogawa C, Radovanovic A, Schwartz A, Teasdale RD, Tegner J, Lenhard B, Teichmann SA, Arakawa T, Ninomiya N, Murakami K, Tagami M, Fukuda S, Imamura K, Kai C, Ishihara R, Kitazume Y, Kawai J, Hume DA, Ideker T, Hayashizaki Y. An atlas of combinatorial transcriptional regulation in mouse and man. Cell. 2010;140:744–752. doi: 10.1016/j.cell.2010.01.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Mi H, Muruganujan A, Casagrande JT, Thomas PD. Large-scale gene function analysis with the PANTHER classification system. Nat Protoc. 2013;8:1551–1566. doi: 10.1038/nprot.2013.092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Ginzinger DG. Gene quantification using real-time quantitative PCR: an emerging technology hits the mainstream. Exp Hematol. 2002;30:503–512. doi: 10.1016/S0301-472X(02)00806-8. [DOI] [PubMed] [Google Scholar]
- 86.Barker M, Rayens W. Partial least squares for discrimination. J Chemom. 2003;17:166–173. doi: 10.1002/cem.785. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All data generated or analyzed during this study are included in this published article [and its supplementary information files]. The RNA-Seq data were deposited in the NCBI Short Read Archive database with Accession number: PRJNA299102.