

Abstract
Pancreatic cancer is one of the most lethal malignancies and the fourth leading cause of cancer-related deaths worldwide, because of its aggressive and highly metastatic ability even in its early stages. Cancer-associated fibroblasts (CAFs) have been reported to be vital to pancreatic cancer progression via regulating tumorigenesis and metastasis. Studies have shown that CAFs also mediate epithelial-to-mesenchymal transition (EMT) in tumor cells. Curcumin has recently been reported to have anticancer effects by modulating pathways involved in cancer progression. Therefore, we hypothesized that curcumin might inhibit EMT by targeting CAFs in pancreatic cancer. In this study, we show that the CAF-mediated enhancement of pancreatic cancer cell migration and metastasis was blocked by curcumin. In conclusion, our data provide the first evidence that curcumin inhibits the migration and metastasis of pancreatic cancer cells by reducing the mesenchymal characteristics of CAFs, which reverses the EMT phenotypes of pancreatic cancer cells.
Keywords: Pancreatic cancer, cancer-associated fibroblasts (CAFs), metastatic, curcumin, epithelial-mesenchymal transition (EMT)
Introduction
Pancreatic ductal adenocarcinoma (PDAC) is one of the most highly malignant tumors and has an extremely poor prognosis; It is resistant to chemotherapy and radiation [1]. Most patients are diagnosed at late stages, which contributes to its high death rate. Less than 20% of PDAC patients have the option of surgical resection; However, most patients incur reoccurrence even after resection, making pancreatic cancer a difficult disease to treat. Recently, researchers have shifted their focus to the influences of stromal components on PDAC. It has become obvious that the desmoplastic reaction is not simply secondary to tumor formation, but a driver of the cellular and molecular components that promote tumor progression and metastasis [2,3]. These cellular components include several heterotypic cells, such as cancer-associated fibroblasts (CAFs), macrophages and endothelial cells. CAFs are essential cells that exist in many malignant tumors, including PDAC, breast cancer and gastric cancer [4,5]. CAFs influence tumor behavior in many aspects, including growth, survival, metastasis, angiogenesis and immune surveillance [6]. Unlike resting fibroblasts, CAFs can acquire an activated phenotype, which can be identified by vimentin and α-smooth muscle actin (α-SMA) expression. Moreover, cancer cells can also transform into fibroblast-like cells via EMT [7]. EMT is a transcriptional and epigenetic program through which epithelial cells acquire mesenchymal features, such as loss of cell-cell junctions and increased motility, allowing them to escape the primary tumor and possibly colonize distant organs [8,9]. It has been shown that CAFs can affect cancer cell EMT, and that the fibroblast-like cancer cells have higher invasiveness and metastasis ability. A hallmark of EMT is loss of E-cadherin expression, which usually correlates with tumor grade and stage [10-12].
Curcumin, commonly known as turmeric, is a natural polyphenol found in the roots of Curcuma longa. Curcumin is non-toxic and has various therapeutic properties including anti-oxidant, anti-inflammatory, analgesic, and anti-septic activity, and thus has been used predominantly in Ayurvedic medicine for centuries [13,14]. Recently, curcumin has been shown to have anticancer activities through its effects on different biological pathways involved in oncogene expression, cell cycle regulation, apoptosis, tumorigenesis and metastasis [15]. However, whether curcumin suppresses the migration and metastasis of pancreatic cancer has not been fully elucidated. Based on previous studies, we hypothesized that curcumin might target CAFs and inhibit the migration ability of pancreatic cancer cells. In this study, we show that curcumin can attenuate the mesenchymal characteristics of CAFs, inhibiting the migration and metastasis of pancreatic cancer cells by reversing EMT phenotypes.
Materials and methods
Reagents
Curcumin (C21H20O6, molecular weight: 368.38, purity > 99%) powder was purchased from Nanjing Zelang Medical Technology Co., Ltd. (Nanjing, China). Curcumin is a lipophilic polyphenol that is insoluble in water but readily soluble in organic solvents such as dimethyl sulfoxide (DMSO), acetone and ethanol [16]. A stock solution of curcumin was dissolved at 10 mM in DMSO and stored at -20°C until use.
Cell lines and mice
The human pancreatic cancer cell lines Capan1 and Panc1 were obtained from American Type Culture Collection (Manassas, VA, USA). We regularly assessed their morphologies and tested for mycoplasma contamination using MycoAlert (Lonza, Rockland, ME, USA). Stromal fibroblasts were isolated from pancreatic cancer tissues obtained from two PDAC patients who underwent surgical resection. Detailed information regarding primary fibroblast isolation, culture and confirmation has been described in our previous publication [17]. Stromal fibroblasts were cultured in Dulbecco’s modified Eagle medium (DMEM; Gibco, Carlsbad, CA) high glucose supplemented with 10% fetal bovine serum (FBS) and 100 µg/mL each of penicillin and streptomycin at 37°C with 5% CO2 in a humidified atmosphere. Capan1 and Panc1 cells were used between the thirtieth to the fortieth passage. CAFs were used for 15 passages, during which there was no significant diminution in their ability to elicit EMT.
Female BALB/c-nu/nu nude mice aged between 4-6 weeks were obtained from the Shanghai Institute of Materia Medica, Chinese Academy of Sciences (Shanghai, China). Mice were housed in laminar flow cabinets under specific pathogen-free conditions and provided food and water ad libitum.
Cell counting kit-8 (CCK-8)
The CAFs cells (5 × 103 cells in 0.1 mL) were plated in duplicate wells of 96 well plates. After overnight incubation, media were removed and replaced with different curcumin concentrations ranging from 0 to 50 µM in DMEM. After 48 h, cell proliferation indices were assessed daily using CCK-8 (Dojindo Molecular Technologies Inc., Gaithersburg, MD, USA), and growth curves were drawn according to OD values obtained from the assay. Subsequently, a series of curcumin concentrations lower than the IC50 value (1, 5 and 10 µM) were chosen for further studies.
Preparation of curcumin-pretreated CAF-conditioned medium (CAF-CM)
CAF-CM was obtained as previously described, with slight modifications [18]. Briefly, to obtain CM from curcumin-pretreated CAFs, isolated CAFs were cultured in DMEM containing 10% curcumin- or vehicle-containing FBS. After 48 h, the medium was removed, and cells were washed twice with PBS and incubated with fresh DMEM containing 10% FBS for an additional 24 h to obtain CM free of curcumin to further incubate pancreatic cancer cells. CAF-CM was subsequently harvested, cleared by centrifugation and transferred without storage.
Transwell migration assay
Transwell migration assays were carried out using 8.0-µm cell culture inserts (BD Biosciences, Franklin Lakes, NJ, USA) to test the migratory ability of Capan1 and Panc1 cells. Cells were seeded at 5 × 104 per well in 100 µL DMEM in the upper chamber of 24-well plates; CM were added in the lower chamber. Cells were allowed to migrate for 24 h (Panc1) or 36 h (Capan1) toward the underside of the membrane. After culturing, the cells that remained in the upper chamber were removed by wiping with a cotton swab and the migrated cells on the lower surface were fixed with ice-cold methanol and stained with crystal violet in 20% ethanol for 1 h. Images were acquired under light microscopy, and fully migrated cells were quantitated by counting cells in 5 predetermined fields. This process was repeated independently 3 times.
RNA isolation and quantitative real-time PCR
Total RNA was isolated using TRIzol reagent (Invitrogen Life Technologies, Carlsbad, CA, USA) according to the manufacturer’s protocol. Reverse transcription was performed using a PrimeScript RT reagent Kit (TaKaRa, Dalian, China). All primers were purchased from Sangon Biotech (Shanghai, China). The forward primer for human E-cadherin was 5’-GTCTCTCTCACCACCTCCACAG-3’, and the reverse primer was CAGACAGAGTGGGGAAAATGTA-3’. The forward primer for human vimentin was 5’-GAAGAGAACTTTGCCGTTGAAG-3’, and the reverse primer was 5’-ACGAAGGTGACGAGCCATT-3’. RT reactions were performed using the TaKaRa kit according to the manufacturer’s instructions. Quantitative PCR was performed using the SYBR green system (TaKaRa). All reactions were performed in triplicate. The 2-ΔΔCt method was used to determine relative gene expression.
Western blotting
Total protein was extracted from cultured cells and quantitated using the bicinchoninic acid assay kit (Pierce, Rockford, IL, USA). Western blotting was performed as previously reported. Equal amounts of protein from different samples were separated through 10% SDS-polyacrylamide gel electrophoresis and transferred to polyvinylidene fluoride membranes. Blocking buffer containing 5% non-fat milk was used to block and incubate membranes with primary antibodies. Target protein expression was examined using an enhance chemiluminescence kit (Amersham Pharmacia Biotech, Uppsala, Sweden), and quantitative analyses were performed using ImageJ software.
Xenograft tumor models
All experiments involving animals were in accordance with the Guidelines for the Care and Use of Laboratory Animals and were performed according to the institutional ethical guidelines of the NIH. The study protocol was approved by the Committee on the Use of Live Animals in Teaching and Research, Fudan University, Shanghai, China.
Panc1 cells were continuously cultured with CAF-CM for two weeks. Panc1 cells (2 × 106 cells/mouse) were collected, enzymatically dissociated and injected into the tail vein of each mouse. Mice were sacrificed 8 weeks after injection, and lungs were removed. The ratio of metastatic area was quantified for each mouse, with each group contained a minimum of 6 mice.
Statistical analysis
Data are expressed as the means ± standard deviations. Statistical analyses were performed using analysis of variance or Student’s t-tests. A P-value less than 0.05 was considered statistically significant. All statistical analyses were performed using SPSS v15.0 (SPSS Inc., Chicago, IL, USA).
Results
CAFs promote the migration capacity of pancreatic cancer cells by promoting EMT
Many studies have shown that CAFs can enhance cancer invasion, including studies in breast and prostate cancer. Molecularly, this process is thought to involve pathways such as paracrine TGF-β signaling, MAOA/mTOR/HIF-1α signaling or miRNAs that promote cancer migration and invasion [19-21]. In this study, we investigated whether CAF could affect EMT marker expression in human pancreatic cancer cells. Transwell assays demonstrated that CAF-CM significantly enhanced pancreatic cancer cell migration (Figure 1A). Next, we observed the morphology of pancreatic cancer cells after culture with CAF-CM. There were distinct morphological changes, including a depletion of cell-cell junctions and presence of needle-like extensions (Figure 2B). Furthermore, we evaluated EMT-associated marker expression after culture with CAF-CM, which showed increased Vimentin expression and E-cadherin downregulation by both RT-PCR and western blotting (Figure 1C, 1D). Thus, we confirmed CAFs promote the migration capacity of pancreatic cancer cells by promoting EMT.
Figure 1.
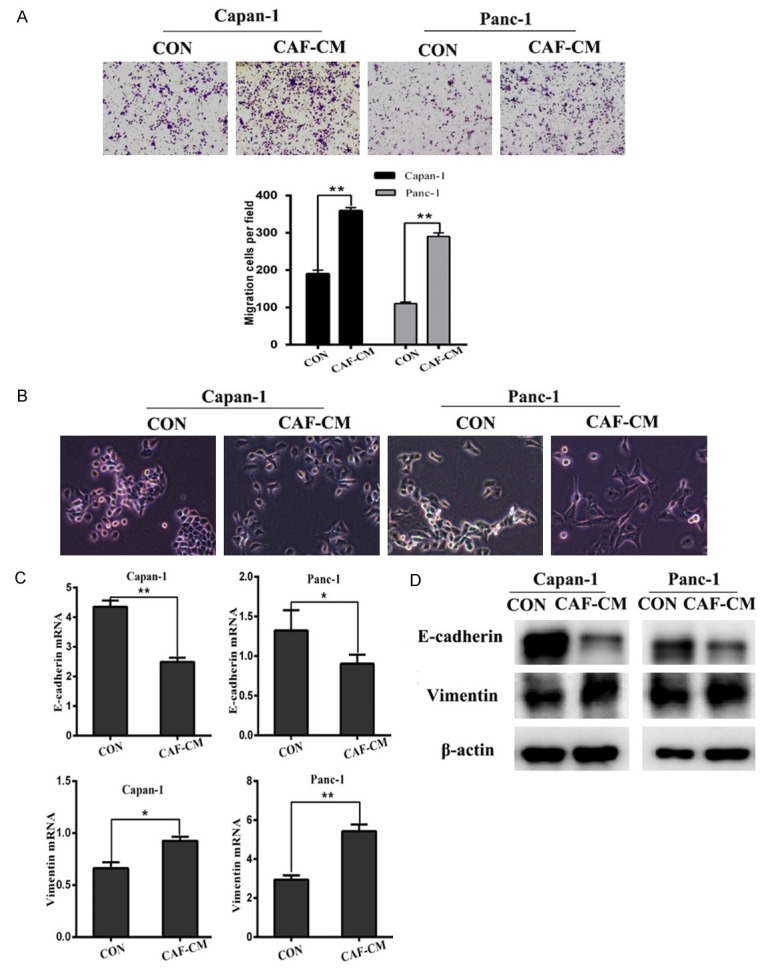
CAFs promote the migration capacity of pancreatic cancer cells by promoting EMT. A. The effect of CM from CAFs on pancreatic cancer cell migration was detected using Transwell chambers in Capan1 and Panc1 cells. B. Morphological changes in Panc1 and Capan1 cells after culture with pancreatic fibroblasts. C. QRT-PCR analysis of E-cadherin and Vimentin levels in Panc1 and Capan1 cells treated as described in A. D. E-cadherin and Vimentin levels determined by western blotting in pancreatic cancer cells culture with CAF-CM. *P < 0.05, **P < 0.01.
Figure 2.
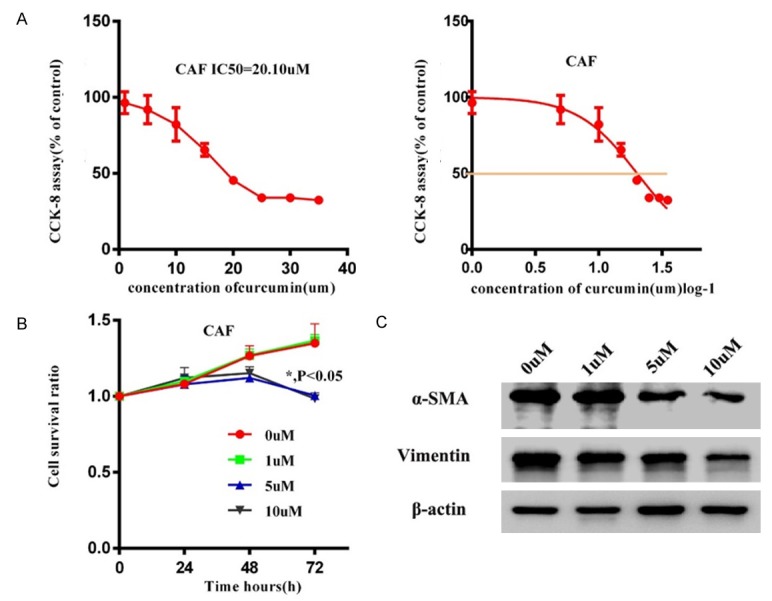
Cytotoxicity and effects of curcumin on CAFs. A. The cytotoxicity of curcumin at various concentrations (1 to 50 µm) in pancreatic cancer cells. B. Effect of curcumin on CAFs growth as detected by CCK-8 assay after treatment with curcumin at 0, 1, 5 and 10 µm for 0, 24, 48 and 72 h. C. Α-SMA and Vimentin levels were determined by western blotting in CAFs cells after curcumin treatment.
Cytotoxicity and other curcumin effects in CAFs
To test the cytotoxicity of herbal curcumin in CAFs, CAFs were treated with curcumin at various concentrations (1 to 50 µm) for 24 h, and the CCK-8 assay was used to evaluate cell viability. The results demonstrated that the proliferation rates of CAFs decreased in response to curcumin in a dose dependent fashion. High-concentration curcumin exhibited cytotoxic effects on CAFs, and we confirmed that the IC50 of curcumin is 20.10 µm (Figure 2A). CAFs were also subjected to curcumin at concentrations lower than the IC50 (0, 1, 5 and 10 µm) for 0, 24, 48 and 72 h. CAF proliferation rates were uninfluenced at 48 h treatment (P > 0.05); However, 72 h curcumin treatment showed notable inhibitory effects (P < 0.05) (Figure 2B). Therefore, curcumin was applied at concentrations no higher than 10 µm and the incubation time was no longer than 48 h in subsequent experiments to avoid cytotoxic effects. The expression of α-SMA and Vimentin in CAFs after curcumin treatment were determined by western blotting. We found that α-SMA and Vimentin (fibrosis active proteins) in CAFs decreased compared with untreated controls (Figure 2C), demonstrating that curcumin can reduce the mesenchyme characteristics of CAFs.
Curcumin attenuated the cancer promoting capacity of CAFs by reducing their mesenchymal characteristics
Studies have demonstrated that curcumin inhibits invasion by altering EMT in prostate and breast cancer [21,22]. However, studies focusing on the effect of curcumin in PDAC by affecting CAFs are rare. The migration capacity of pancreatic cancer cells treated with curcumin-pretreated CAF-CM decreased with increasing curcumin concentrations (Figure 3A). Then we observed whether curcumin-pretreated CAF-CM influenced EMT in pancreatic cancer cells. A depletion of cell-cell junctions and presence of needle-like extensions were obvious in pancreatic cancer cells cultured with CAF-CM, but in the curcumin-pretreated (5 or 10 µm) groups these events were receded to the different extent (Figure 3B). Next, E-cadherin and Vimentin expression in pancreatic cancer cells treated with CAF-CM were evaluated using real-time PCR, which showed that Vimentin was reduced and E-cadherin increased (Figure 3C). We also performed western blotting to assess EMT marker expression at the protein level. Increased E-cadherin expression and a downregulation of Vimentin were observed in pancreatic cancer cells cultured in CAF-CM from curcumin-pretreated CAFs (Figure 3D). Furthermore, combined with Figure 2C, the data suggest that curcumin inhibited the migration-promoting capacity of CAFs by reducing the mesenchymal characteristic of CAFs.
Figure 3.
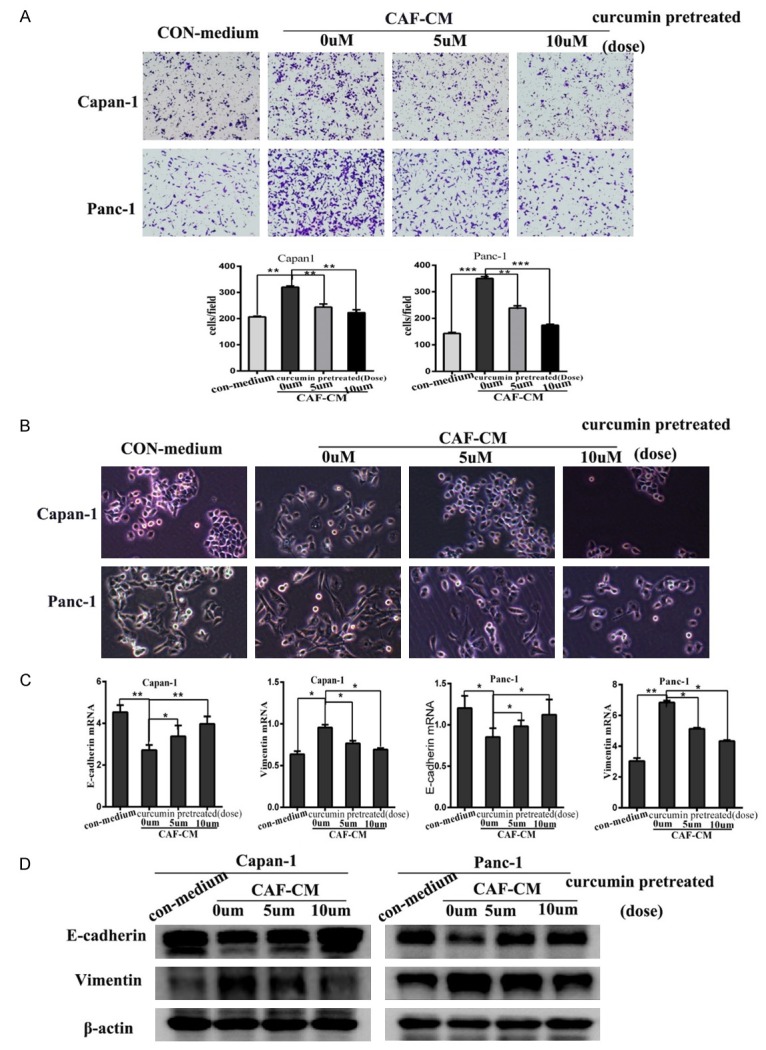
Curcumin attenuated the migration-promoting capacity of CAFs by reducing their mesenchymal characteristics. A. The inhibitory effect of CAF-CM (0, 5 or 10 µm curcumin-pretreated) on pancreatic cancer cell migration was detected using the transwell assay with Capan1 and Panc1 cells. B. Morphological changes in Panc1 and Capan1 cells after culture with pancreatic fibroblasts treated as described in A. C. QRT-PCR analysis of E-cadherin and Vimentin mRNA levels in Panc1 and Capan1 cells treated as described in A. D. E-cadherin and Vimentin expression were measured in pancreatic cancer cells cultured with CAF-CM as described in A. *P < 0.05, **P < 0.01, ***P < 0.001.
Curcumin treatment inhibited metastasis in vivo
To further explore the effects of curcumin on CAFs and metastasis in vivo, we next examined the metastatic nodules formed in the lungs of Female BALB/c-nu/nu nude mice injected in the tail vein with Panc1 cells cultured with DMEM containing vehicle (control-medium) or CAF-CM. Mice were sacrificed 8 weeks after injection and lungs were harvested. The control and CAF-CM groups formed lungs tumors following tail vein injection. However, pancreatic cancer cells exposed to CAF-CM (10 µm curcumin pretreated) showed significantly fewer and smaller lung tumors than CAF-CM (0 µM). Moreover, addition of the CAF-CM (10 µM) prior to the control generates the formation and number of lung metastases (Figure 4A-C). Taken together, these results indicate that curcumin may inhibit pancreatic cancer metastasis by targeting CAFs.
Figure 4.
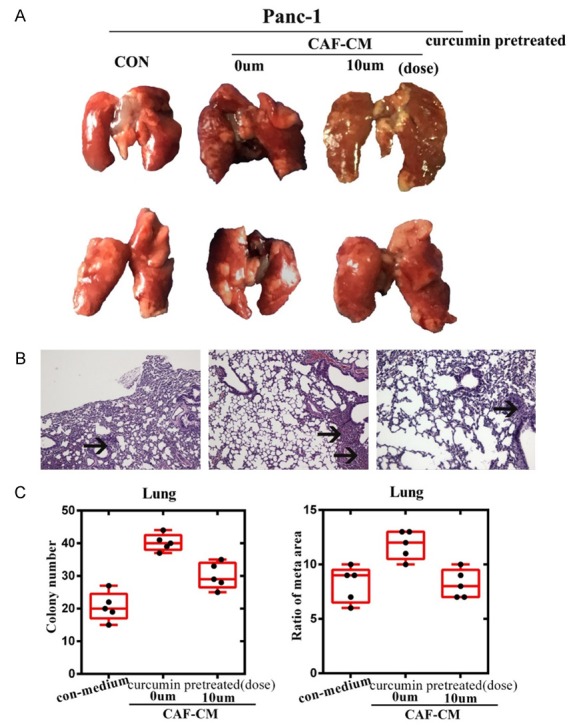
Curcumin treatment inhibited metastasis in vivo. A. Representative lungs from Female BALB/c-nu/nu nude mice are shown. B. Hematoxylin and eosin staining was performed on metastatic tumor and normal lung sections. Metastatic nodules are indicated by arrows at 200 × magnification. C. Metastases were quantified by counting the number of metastatic colonies in one histological section of the mid-portion of each lung sample from each mouse and determining the ratio of metastatic area to total area in histological sections from the mid-portion of each lung (n = 6 per group). A Student’s t-test was used to determine the statistical significance of differences between the groups.
Discussion
In this study, we revealed the curcumin can inhibit the migration and metastasis of pancreatic cancer cells by targeting CAFs, confirming that the tumor microenvironment could be targeted by Traditional Chinese Medicine (TCM) [23].
Recently, many countries have accepted TCM, particularly for cancer treatment [24,25]. We have stressed the holistic concept of TCM; In the tumor microenvironment, CAFs play an important role in cancer invasion and metastasis [6]. It has been demonstrated that CAFs can affect cancer cell EMT [26,27]. Cancer treatment should not be confined to the cancer itself, but should include the tumor environment. The overall concept of treatment in TCM emphasizes local links with the surrounding environment; Both have the same view. We think TCM may provide a suitable and effective treatment for pancreatic cancer, especially for the tumor microenvironment.
Curcumin, a naturally occurring polyphenolic compound that derives from turmeric (Curcuma longa), has been commonly used as a food additive or dietary pigment and in traditional medicines [28-30]. Several preclinical studies have demonstrated that curcumin has anticancer activities against different types of cancer, including PDAC, by modulating many molecular targets [31-33]. Moreover, in the tumor microenvironment, curcumin was reported to suppress crosstalk between cancer cells and stromal fibroblasts in colon and head and neck cancers [34,35]. In our study, using in vitro functional assays, we demonstrated that CAFs can promote PDAC cell migration and enhance EMT in pancreatic cancer cells. Upon the treatment of CAFs with curcumin, the invasive ability of cultured pancreatic cancer cells decreased. Further results showed that the expression of mesenchymal markers in CAFs receded, and EMT in pancreatic cancer was reversed. Further in vivo studies confirmed these results. Mice injected with tumor cells treated with curcumin-pretreated CAF-CM (0 µm) were more inclined to lung metastasis compared with the control group, but CAF-CM treated with (10 µm) were reduced compared with CAF-CM.
In conclusion, targeting CAFs and cancer cells through pharmacological inhibition might provide new insight into the therapeutic effects of curcumin for PDAC. In this study, we showed that curcumin significantly suppressed the mesenchymal features of CAFs and reversed pancreatic cancer cell EMT, thereby preventing pancreatic cancer cell migration and metastasis. However, the concrete pathways involved in EMT that curcumin may target in PDAC remain to be elucidated.
Acknowledgements
This work is supported by National Science Fundation of China (81622049, 81572376, 81370068); National Young Top-notch Talent program and Fudan University Zhuo-Xue program. We thank Dr. Haixia Bi (Department of Pathology, Jing’an District Center Hospital of Shanghai (Huashan Hospital, Fudan University, Jing’an Branch) for analyzing pancreatic cancer H&E samples.
Disclosure of conflict of interest
None.
References
- 1.Siegel R, Ma J, Zou Z, Jemal A. Cancer statistics, 2014. CA Cancer J Clin. 2014;64:9–29. doi: 10.3322/caac.21208. [DOI] [PubMed] [Google Scholar]
- 2.Waghray M, Yalamanchili M, di Magliano MP, Simeone DM. Deciphering the role of stroma in pancreatic cancer. Curr Opin Gastroenterol. 2013;29:537–543. doi: 10.1097/MOG.0b013e328363affe. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Feig C, Gopinathan A, Neesse A, Chan DS, Cook N, Tuveson DA. The pancreas cancer microenvironment. Clin Cancer Res. 2012;18:4266–4276. doi: 10.1158/1078-0432.CCR-11-3114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Kasashima H, Yashiro M, Kinoshita H, Fukuoka T, Morisaki T, Masuda G, Sakurai K, Kubo N, Ohira M, Hirakawa K. Lysyl oxidase-like 2 (LOXL2) from stromal fibroblasts stimulates the progression of gastric cancer. Cancer Lett. 2014;354:438–446. doi: 10.1016/j.canlet.2014.08.014. [DOI] [PubMed] [Google Scholar]
- 5.Sun X, Mao Y, Wang J, Zu L, Hao M, Cheng G, Qu Q, Cui D, Keller ET, Chen X, Shen K, Wang J. IL-6 secreted by cancer-associated fibroblasts induces tamoxifen resistance in luminal breast cancer. Oncogene. 2014;33:4450. doi: 10.1038/onc.2014.158. [DOI] [PubMed] [Google Scholar]
- 6.Lunardi S, Muschel RJ, Brunner TB. The stromal compartments in pancreatic cancer: Are there any therapeutic targets? Cancer Lett. 2014;343:147–155. doi: 10.1016/j.canlet.2013.09.039. [DOI] [PubMed] [Google Scholar]
- 7.Kalluri R, Zeisberg M. Fibroblasts in cancer. Nat Rev Cancer. 2006;6:392–401. doi: 10.1038/nrc1877. [DOI] [PubMed] [Google Scholar]
- 8.Thiery JP. Epithelial-mesenchymal transitions in tumour progression. Nat Rev Cancer. 2002;2:442–454. doi: 10.1038/nrc822. [DOI] [PubMed] [Google Scholar]
- 9.Friedl P. Prespecification and plasticity: shifting mechanisms of cell migration. Curr Opin Cell Biol. 2004;16:14–23. doi: 10.1016/j.ceb.2003.11.001. [DOI] [PubMed] [Google Scholar]
- 10.Christofori G. New signals from the invasive front. Nature. 2006;441:444–450. doi: 10.1038/nature04872. [DOI] [PubMed] [Google Scholar]
- 11.Cowin P, Rowlands TM, Hatsell SJ. Cadherins and catenins in breast cancer. Curr Opin Cell Biol. 2005;17:499–508. doi: 10.1016/j.ceb.2005.08.014. [DOI] [PubMed] [Google Scholar]
- 12.Franci C, Takkunen M, Dave N, Alameda F, Gomez S, Rodriguez R, Escriva M, Montserrat-Sentis B, Baro T, Garrido M, Bonilla F, Virtanen I, García de Herreros A. Expression of Snail protein in tumor-stroma interface. Oncogene. 2006;25:5134–5144. doi: 10.1038/sj.onc.1209519. [DOI] [PubMed] [Google Scholar]
- 13.Aggarwal BB, Sundaram C, Malani N, Ichikawa H. Curcumin: the indian solid gold. Adv Exp Med Biol. 2007;595:1–75. doi: 10.1007/978-0-387-46401-5_1. [DOI] [PubMed] [Google Scholar]
- 14.Ammon HP, Wahl MA. Pharmacology of curcuma longa. Planta Med. 1991;57:1–7. doi: 10.1055/s-2006-960004. [DOI] [PubMed] [Google Scholar]
- 15.Wilken R, Veena MS, Wang MB, Srivatsan ES. Curcumin: a review of anti-cancer properties and therapeutic activity in head and neck squamous cell carcinoma. Mol Cancer. 2011;10:12. doi: 10.1186/1476-4598-10-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Anand P, Sundaram C, Jhurani S, Kunnumakkara AB, Aggarwal BB. Curcumin and cancer: an “old-age” disease with an “age-old” solution. Cancer Lett. 2008;267:133–164. doi: 10.1016/j.canlet.2008.03.025. [DOI] [PubMed] [Google Scholar]
- 17.Chen L, Qu C, Chen H, Xu L, Qi Q, Luo J, Wang K, Meng Z, Chen Z, Wang P, Liu L. Chinese herbal medicine suppresses invasion-promoting capacity of cancer-associated fibroblasts in pancreatic cancer. PLoS One. 2014;9:e96177. doi: 10.1371/journal.pone.0096177. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 18.Muerkoster S, Wegehenkel K, Arlt A, Witt M, Sipos B, Kruse ML, Sebens T, Kloppel G, Kalthoff H, Folsch UR, Schafer H. Tumor stroma interactions induce chemoresistance in pancreatic ductal carcinoma cells involving increased secretion and paracrine effects of nitric oxide and interleukin-1beta. Cancer Res. 2004;64:1331–1337. doi: 10.1158/0008-5472.can-03-1860. [DOI] [PubMed] [Google Scholar]
- 19.Du Y, Long Q, Zhang L, Shi Y, Liu X, Li X, Guan B, Tian Y, Wang X, Li L, He D. Curcumin inhibits cancer-associated fibroblast-driven prostate cancer invasion through MAOA/mTOR/HIF-1alpha signaling. Int J Oncol. 2015;47:2064–2072. doi: 10.3892/ijo.2015.3202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Gandellini P, Giannoni E, Casamichele A, Taddei ML, Callari M, Piovan C, Valdagni R, Pierotti MA, Zaffaroni N, Chiarugi P. miR-205 hinders the malignant interplay between prostate cancer cells and associated fibroblasts. Antioxid Redox Signal. 2014;20:1045–1059. doi: 10.1089/ars.2013.5292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Yu Y, Xiao CH, Tan LD, Wang QS, Li XQ, Feng YM. Cancer-associated fibroblasts induce epithelial-mesenchymal transition of breast cancer cells through paracrine TGF-beta signalling. Br J Cancer. 2014;110:724–732. doi: 10.1038/bjc.2013.768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Gallardo M, Calaf GM. Curcumin inhibits invasive capabilities through epithelial mesenchymal transition in breast cancer cell lines. Int J Oncol. 2016;49:1019–1027. doi: 10.3892/ijo.2016.3598. [DOI] [PubMed] [Google Scholar]
- 23.Chen Z, Chen LY, Wang P, Dai HY, Gao S, Wang K. Tumor microenvironment varies under different TCM ZHENG models and correlates with treatment response to herbal medicine. Evid Based Complement Alternat Med. 2012;2012:635702. doi: 10.1155/2012/635702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Chen Z, Gu K, Zheng Y, Zheng W, Lu W, Shu XO. The use of complementary and alternative medicine among Chinese women with breast cancer. J Altern Complement Med. 2008;14:1049–1055. doi: 10.1089/acm.2008.0039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Chang KH, Brodie R, Choong MA, Sweeney KJ, Kerin MJ. Complementary and alternative medicine use in oncology: a questionnaire survey of patients and health care professionals. BMC Cancer. 2011;11:196. doi: 10.1186/1471-2407-11-196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kikuta K, Masamune A, Watanabe T, Ariga H, Itoh H, Hamada S, Satoh K, Egawa S, Unno M, Shimosegawa T. Pancreatic stellate cells promote epithelial-mesenchymal transition in pancreatic cancer cells. Biochem Biophys Res Commun. 2010;403:380–384. doi: 10.1016/j.bbrc.2010.11.040. [DOI] [PubMed] [Google Scholar]
- 27.Alba-Castellon L, Olivera-Salguero R, Mestre-Farrera A, Pena R, Herrera M, Bonilla F, Casal JI, Baulida J, Pena C, García de Herreros A. Snail1-Dependent activation of cancer-associated fibroblast controls epithelial tumor cell invasion and metastasis. Cancer Res. 2016;76:6205–6217. doi: 10.1158/0008-5472.CAN-16-0176. [DOI] [PubMed] [Google Scholar]
- 28.Kanai M. Therapeutic applications of curcumin for patients with pancreatic cancer. World J Gastroenterol. 2014;20:9384–9391. doi: 10.3748/wjg.v20.i28.9384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Strimpakos AS, Sharma RA. Curcumin: preventive and therapeutic properties in laboratory studies and clinical trials. Antioxid Redox Signal. 2008;10:511–545. doi: 10.1089/ars.2007.1769. [DOI] [PubMed] [Google Scholar]
- 30.Perrone D, Ardito F, Giannatempo G, Dioguardi M, Troiano G, Lo Russo L, DE Lillo A, Laino L, Lo Muzio L. Biological and therapeutic activities, and anticancer properties of curcumin. Exp Ther Med. 2015;10:1615–1623. doi: 10.3892/etm.2015.2749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Pattanayak R, Basak P, Sen S, Bhattacharyya M. Interaction of KRAS G-quadruplex with natural polyphenols: a spectroscopic analysis with molecular modeling. Int J Biol Macromol. 2016;89:228–237. doi: 10.1016/j.ijbiomac.2016.04.074. [DOI] [PubMed] [Google Scholar]
- 32.Bimonte S, Barbieri A, Palma G, Luciano A, Rea D, Arra C. Curcumin inhibits tumor growth and angiogenesis in an orthotopic mouse model of human pancreatic cancer. Biomed Res Int. 2013;2013:810423. doi: 10.1155/2013/810423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Bimonte S, Barbieri A, Palma G, Rea D, Luciano A, D’Aiuto M, Arra C, Izzo F. Dissecting the role of curcumin in tumour growth and angiogenesis in mouse model of human breast cancer. Biomed Res Int. 2015;2015:878134. doi: 10.1155/2015/878134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Dudas J, Fullar A, Romani A, Pritz C, Kovalszky I, Hans Schartinger V, Mathias Sprinzl G, Riechelmann H. Curcumin targets fibroblast-tumor cell interactions in oral squamous cell carcinoma. Exp Cell Res. 2013;319:800–809. doi: 10.1016/j.yexcr.2012.12.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Hendrayani SF, Al-Khalaf HH, Aboussekhra A. Curcumin triggers p16-dependent senescence in active breast cancer-associated fibroblasts and suppresses their paracrine procarcinogenic effects. Neoplasia. 2013;15:631–640. doi: 10.1593/neo.13478. [DOI] [PMC free article] [PubMed] [Google Scholar]