

Abstract
Background
Korean mistletoe (Viscum album coloratum) is a semi-parasitic plant that grows on various trees and has a diverse range of effects on biological functions, being implicated in having anti-tumor, immunostimulatory, anti-diabetic, and anti-obesity properties. Recently, we also reported that Korean mistletoe extract (KME) improves endurance exercise in mice, suggesting its beneficial roles in enhancing the capacity of skeletal muscle.
Methods
We examined the expression pattern of several genes concerned with muscle physiology in C2C12 myotubes cells to identify whether KME inhibits muscle atrophy or promotes muscle hypertrophy. We also investigated these effects of KME in denervated mice model.
Results
Interestingly, KME induced the mRNA expression of SREBP-1c, PGC-1α, and GLUT4, known positive regulators of muscle hypertrophy, in C2C12 cells. On the contrary, KME reduced the expression of Atrogin-1, which is directly involved in the induction of muscle atrophy. In animal models, KME mitigated the decrease of muscle weight in denervated mice. The expression of Atrogin-1 was also diminished in those mice. Moreover, KME enhanced the grip strength and muscle weight in long-term feeding mice.
Conclusions
Our results suggest that KME has beneficial effects on muscle atrophy and muscle hypertrophy.
Background
Muscle atrophy is a feature of aging, starvation, cancer, diabetes, muscle denervation, and other physical conditions that limit muscle use [1]. The decrease of muscle amount in the human body is considered as a symptom of the disorder. With the increase of average life expectancy, the number of cancer and diabetes patients has also increased. Thus, the importance of maintaining muscle mass is increasing. Although muscle atrophy occurs frequently, there are no common clinical therapies to prevent or repress the muscle atrophy in human cases to date [2].
Over the last few decades, the mechanisms related to muscle mass decrease have not been clearly elucidated. Many research studies were undertaken to find the molecular mechanisms responsible for muscle mass decrease and increase. However, recent studies strongly support the idea that there are specific signaling pathways regulating the muscle weight increase and decrease [3]. Based on the recent reports, muscle mass regulation is closely related to the PI3K/Akt pathway [4]. In the case of muscle hypertrophy, it is generally known that increasing muscle mass is not because of an increased number of muscle cells but because of an increased amount of intracellular protein [5]. In the case of muscle atrophy, the decreasing muscle mass is the opposite of muscle hypertrophy—decreased proteins in muscle cells [6]. People called the genes that regulate this specific, muscle mass decreasing mechanism Murf1 and Atrogin-1 [7]. These genes are closely linked with protein ubiquitination [8, 9]. Nowadays, the aims of studies regarding muscle atrophy and therapy for muscle diseases are to find the unknown molecules or mechanisms regulating Murf1 and Atrogin-1. Murf1 and Atrogin-1 are members of the E3 ubiquitin ligase family [10, 11]. The targets of these E3 proteins are intimately involved with maintaining the structure of muscle cells and the construction of muscle [12]. It was also proven that these E3 ubiquitin ligases are regulated by the MAPK signaling pathway, including FoxO [13]. These genes stimulating muscle mass decrease are repressed by the MAPK signaling pathway and, are activated by FoxO [14]. In the case of muscle hypertrophy, previous studies verified that the Akt/mTOR signaling pathway is the main signaling pathway that regulates the muscle mass increase [15]. The Akt/mTOR pathway is affiliated with several other intracellular signaling pathways and key molecules for survival, proliferation, stimulation of the cell cycle, and many metabolic regulators [16, 17].
Mistletoe is a semi-parasitic plant that has been utilized as a traditional medicine in many countries to treat various human illnesses [18, 19]. In particular, Korean mistletoe (Viscum album coloratum) has been widely studied in the last decades since the report that it has a anticancer activity [20]. It has been reported that Korean mistletoe extract (KME) has a variety of effects on biological functions, demonstrating anti-tumor, anti-oxidant, anti-diabetes, and anti-obesity benefits, and promotes the extension of lifespan [21–23]. In addition, we have recently reported that KME improved the endurance capacity in mice by enhancing mitochondrial activity [24]. In the treadmill and swimming tests, KME-treated mice showed an increased exercise capacity compared to chow-fed control mice. Therefore, it is conceivable that KME also has beneficial functions in enhancing the capacity of skeletal muscle in mice.
In this study, we discovered that KME can induce the phosphorylation of Akt signaling pathway. Therefore, we hypothesized that the increase and decrease of muscle mass can be regulated by KME. Murf1 and Atrogin-1 were chosen as target genes to identify the inhibitory effect of KME in muscle atrophy. We treated mouse myoblast C2C12 cells with KME, and adopted an artificially induced muscle atrophy condition (denervation) [25] in mice to validate its effects in vivo.
Methods
Preparation of KME
Korean mistletoe extract (KME) was supplied by Daeho Corporation (Hwasung, Korea). Summarized manufacturing process is as follows. The leaves, twigs, and fruits of Korean Mistletoe plants were homogenized in 7 volumes of distilled water, using a blender. The homogenized Korean Mistletoe extracts were stirred for 8 h at 100 °C. Then, secondary extraction was performed with 4 volumes of distilled hot water and filtered with 1 μm Whatman™ filter paper. After filtration, the mistletoe extract was evaporated in vacuum condition and stored at 4 °C until use.
Reagents and antibodies
Dulbecco’s modified Eagle’s medium (DMEM), fetal bovine serum, and penicillin/streptomycin were purchased from GIBCO (Thermo Fisher Scientific, Waltham, MA, USA). Anti-phospho-Akt, anti-total Akt, anti-phospho-Foxo and anti-Foxo antibodies were purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA). Anti-mouse HRP and anti-rabbit HRP were purchased from Thermo Scientific. PVDF membranes and SuperSignal® West Pico and West Femto Chemiluminescent Substrate for western blotting analysis were purchased from Thermo Scientific, and X-ray films from AGFA (Mortsel, Belgium). SYBR Green Master Mix for the real-time PCR analysis was purchased from Applied Biosystems (Foster City, CA, USA).
Cell culture
The mouse muscle cell line C2C12 (mouse myoblast cell line) was obtained from ATCC (American Type Culture Collection, Manassas, VA, USA). C2C12 cells were cultured in DMEM supplemented with 10% fetal bovine serum and 1% penicillin/streptomycin, at 37 °C, and in 5% CO2 and 95% humidified air. Before treatment of KME, C2C12 cells were moved to six-well plates and incubated overnight. Cells were then cultured, with the medium containing 2% horse serum, and incubated for 64 h to induce differentiation of the C2C12 cells. After differentiation, KME (100 μg/ml) was treated for 16 h to stimulate cells.
Animals
Five-week-old, specific-pathogen-free, male ICR mice were purchased from Dae-Han Laboratory Animal Research Center Co. Ltd (Seoul, Korea). Mice were cared for in the Laboratory of Animal Experiment, Institute of Bioscience, Handong Global University (Pohang, Korea). Mice were maintained in single cages with 12 h light–12 h dark cycles, and were used after 7 days had passed since their arrival. All of the animal experiments were approved by the Ethics Review Committee of the Handong Global University.
Preparation of cell lysate and western blot analysis
C2C12 cells were prepared for western blotting to detect phospho-Akt, total Akt, and β-actin expression. The cells were washed with cold PBS twice before harvesting. Cells were harvested in 150 μl of cell lysis buffer purchased from iNTRON Biotechnology Inc. (Daejeon, Korea). After collecting the cells, the samples were boiled for 10 min in 95 °C. After boiling, samples were centrifuged at 13,000 rpm for 10 min at 4 °C. The concentration of collected supernatants was determined by a BCA assay kit (Thermo Scientific).
The protein samples (30 μg) were loaded on the 8–10% polyacrylamide gel and blotted on a PVDF membrane in transfer buffer at 200 mA, 2 h after electrophoresis. The blots were blocked with 10% milk in PBST (0.1% Tween-20 in PBS) for 1 h, at room temperature. The blots were incubated with antibodies specific to phospho-Akt (1:1000), Akt(1:1000), phospho-Foxo(1:1000), Foxo(1:1000) and β-actin (1:2000), overnight at 4 °C. The blots were washed three times using PBST, and then incubated with the horseradish peroxidase-conjugated secondary antibody (1:10000) for 1 h, at room temperature. After incubation, the blots were washed again with PBST. After washing, the blots were detected with SuperSignal West Pico or West Femto Chemiluminescent Substrate, using X-ray film.
Denervation and KME treatment
Six-week-old, male ICR mice were used for the denervation experiments. The right leg was denervated by cutting the sciatic nerve in middle of the leg under anesthesia. KME (200 mg/kg or 500 mg/kg) was given to the mice twice a day, every day, for 2 weeks, through oral administration. After this period of time, whole body grip strength was determined using a grip strength machine, with tests repeated five times. Mice were sacrificed and the quadricepses of the mice were collected and directly used for the experiments or keep at −80 °C for future use.
Induction of hypertrophy
For hypertrophy, six-week–old, male ICR mice had access to water and the chow contained KME (0.25% KME extract was included) that was ad libitum for 4 weeks. After the 4 weeks, the whole body grip strength was tested using a grip strength machine, and tests were repeated five times. Mice were then sacrificed and quadricepses were harvested and directly used for the experiments or keep at −80 °C for the future experiments.
Preparing RNA and protein samples from quadricepses
The isolated quadricepses were used in preparing RNA samples for real-time PCR analysis and protein samples for western blotting analysis. PRO-PREP™ lysis buffer was used for preparing protein samples from the quadricepses: the muscle samples were homogenized in the lysis buffer. After homogenization, the samples were centrifuged at 13,000 rpm for 15 min at 4 °C. The concentrations of samples were determined by a BCA assay kit. Total RNA was extracted using the easy-spin™ RNA extraction kit (iNtRON Biotechnology Inc., Seongnam-si, Gyeonggi-do, Korea). Muscle samples were homogenized in the easy-spin™ RNA extraction buffer and the other procedures were performed according to the manufacturer’s instructions.
Real-time PCR analysis
Total RNA (1 μg) was used for cDNA synthesizing with SuperScript™ II Reverse Transcriptase (Invitrogen, Carlsbad, CA, USA) and an oligo(dT) primer. Real-time PCR was performed using the 7500 sequence detection system (Applied Biosystems) with SYBR Green Master Mix (Thermo Scientific) and ROX™ Reference Dye (Invitrogen). The amount of each reagent and the PCR condition used was according to the manufacturers’ instructions. Relative mRNA expression levels were determined using the ΔΔCt value. β-Actin was used for the endogenous loading control. Experiments were performed with specific primer sequences (Table 1).
Table 1.
Real-time PCR primer sequences
| Gene | Orientation | Primer sequence(5′ → 3′) |
|---|---|---|
| PGC-1α | Forward | TCGATGTGTCGCCTTCTTGC |
| Reverse | ACGAGAGCGCATCCTTTGG | |
| Atrogin-1 | Forward | ATTCTACACTGGCAGCAGCA |
| Reverse | TCAGCCTCTGCATGATGTTC | |
| Murf1 | Forward | ACCTGCTGGTGGAAAACATC |
| Reverse | AGGAGCAAGTAGGCACCTCA | |
| GLUT4 | Forward | AACCAGCATCTTCGAGTCGG |
| Reverse | CGAGACCAACGTGAAGACCG | |
| SREBP-1c | Forward | GGAGCCATGGATTGCACATT |
| Reverse | GGCCCGGGAAGTCACTGT |
Analysis of skeletal muscle fiber
Skeletal muscle tissues (quadriceps) were fixed overnight in 4% formaldehyde at room temperature. Then, skeletal muscle samples were frozen in optimal cutting temperature (OCT) and sectioned at 5 μm for hematoxylin and eosin (H&E) staining. Images were captured with Motic BA300 microscope and ImageJ software (developed by Wayne Rasband, National Institute of Health, Bethesda, MD) was used for analysis.
Statistical analysis
Figures were expressed using GraphPad prism 5 software (GraphPad Software, Inc., CA, USA). Data were presented as mean ± S.D. and analyzed by one-way ANOVA t-test. P values < 0.05 were considered as significant.
Results
KME treatment induces the activation of the PI3K/Akt pathway in cells
To investigate the muscle atrophy inhibition and muscle hypertrophy stimulation by KME, we first checked whether the PI3K/Akt pathway is activated in C2C12 cell lines. KME treated cells, and a non-treated control cells were also established. In the KME-treated cells, higher phosphorylation of Akt was detected compared with non-treated control cells (Fig. 1). These data suggest that KME has an effect on the regulation of the muscle mass through the activation of the Akt/mTOR signaling pathway.
Fig. 1.
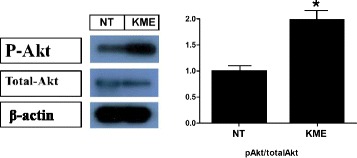
Activation of the Akt signaling pathway by KME (Left) Relative phosphorylation of Akt (P-Akt) is shown. The concentration of KME was 100 μg/ml. (Right) The relative amount of phospho-Akt is shown. All the samples were blotted against β-actin as a loading control. NT: non-treated cells. P-values of < 0.05 and < 0.01 are indicated by * and **, respectively
KME treatment regulates other metabolic pathways in cells
To investigate the effect of KME on muscle mass regulation further, we also examined the phosphorylation of FoxO in C2C12 cells. FoxO is a known key molecule of muscle atrophy due to its role in the activation of protein degradation [13]: FoxO induces muscle atrophy by stimulating the E3 ubiquitin ligases Murf1 and Atrogin-1 [26]. Compared to the non-treated control cells, the KME-treated cells showed increased phosphorylation of FoxO (Fig. 2). These data also support our previous observation that KME could induce the phosphorylation of AMPK, which is a repressor of FoxO.
Fig. 2.
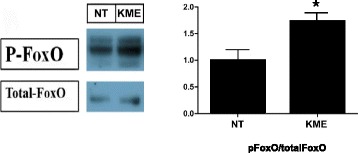
The effect of KME on the phosphorylation of FoxO. Phosphorylation of FoxO (P-FoxO) by KME(100 μg/ml) treatment in C2C12 cells. (Right) The relative amount of phospho-FoxO is indicated. P-values of < 0.05 and < 0.01 are indicated by * and **, respectively
KME treatment regulates muscle atrophy related gene expression in cells
To determine the direct effect of KME in the regulation of muscle atrophy, we checked the mRNA expression of genes related to muscle atrophy in C2C12 cells. A real-time PCR method was used to measure the expression levels of Murf1 and Atrogin-1, so-called “atrogenes”. In addition, the mRNA expression levels of PGC-1α, GLUT4, and SREBP-1c, that regulate the expression of Atrogin-1 and Murf1, were also examined. After 16 h, the KME-treated group showed a decreased gene expression of Atrogin-1, compared to the non-treated group (Fig. 3). On the contrary, the KME-treated group showed increased mRNA expression of PGC-1α, GLUT4, and SREBP-1c, compared to the non-treated group (Fig. 4). These genes are related to the inhibition of muscle atrophy and are related to the induction of muscle hypertrophy [27–29]. These analyses demonstrate the multifunctional effects of KME in the activation of Akt and AMPK signaling pathways.
Fig. 3.
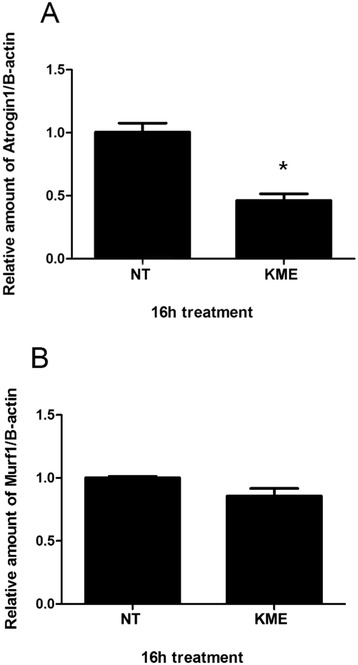
The inhibition of muscle atrophy related gene expression by KME (a) The relative amount of Atrogin-1 was measured after KME treatment. The final concentration of KME was 100 μg/ml. b The relative amount of Murf1 is shown. The mRNA expression levels were compared to β-actin. Data are shown as means of the relative expression levels ± SEM. P-values of < 0.05 and < 0.01 are indicated by * and **, respectively
Fig. 4.
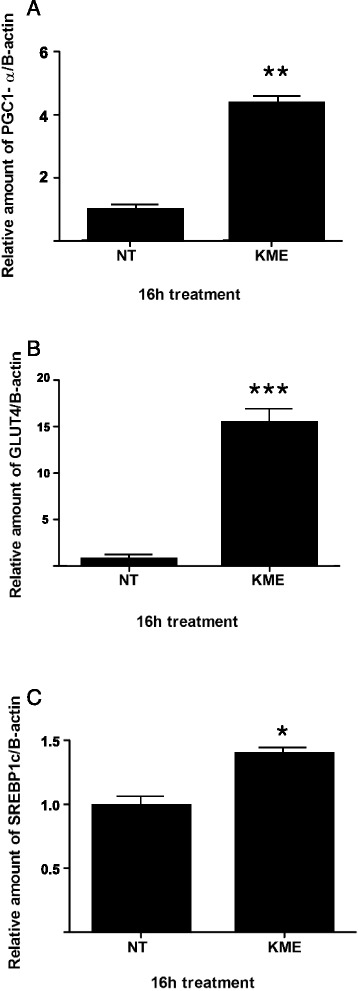
The induction of muscle atrophy and hypertrophy related gene expression by KME. a The relative amount of PGC-1α is shown. b The relative amount of GLUT4 is shown. c The relative amount of SREBP-1c is shown. All the analyses were performed after treatment of KME at a final concentration of 100 μg/ml. The mRNA expression levels were compared to β-actin. Data are shown as means of the relative expression levels ± SEM. P-values of < 0.05 and < 0.01 are indicated by * and **, respectively
KME lessens the decrease of muscle mass in denervated mouse models
Based on our data from real-time RT-PCR in C2C12 cells, we hypothesized that KME might reduce muscle atrophy in vivo, using the animal model. To validate this hypothesis, we tried to selectively denervate the right quadricepses of mice by amputating each right leg’s sciatic nerve. Denervation is a common method to induce artificial muscle atrophy in the specific skeletal muscle. In our study, we designated control group the PBS-treated denervated group. After denervation, we treated KME (200 mg/kg and 500 mg/kg) to the mice every 12 h by oral gavage for 15 days. After 15 days of treatment, KME treated mice showed decreased denervation induced by muscle atrophy (Fig. 5a). In the mRNA expression levels monitored, Murf1 expression was not affected by KME treatment. However, in the case of Atrogin-1, KME (low fed) decreased the expression of Atrogin-1 compared to the KME non-treated group (Fig. 5b and c). These data suggest that KME might repress the artificial muscle decrease, and this effect is closely related to the regulation of Atrogin-1 gene expression.
Fig. 5.
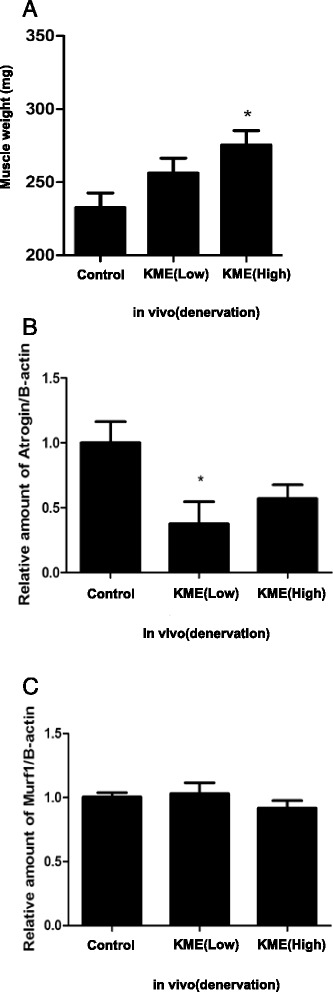
Decreased muscle atrophy in KME-fed denervated mice (a) The weight of quadricepses is shown. b The relative amount of Atrogin-1 is shown. c The relative amount of Murf1 is shown. Data are shown as means of the relative expression levels ± SEM; n = 5 per group. P-values of < 0.05 and < 0.01 are indicate by * and **, respectively. KME(low) : 0.3%, KME(high) : 1.5%
KME increases muscle mass and grip strength in native conditions
The reduction of muscle atrophy by KME led us to hypothesize that KME might induce muscle hypertrophy. This hypothesis is based on the observation that KME induces the upregulation of muscle atrophy related genes and the activation of the Akt signaling pathway in C2C12 cells. To evaluate the muscle hypertrophic effects of KME in vivo, mice were allowed free access to control chow or chow containing 0.3% or 1.5% KME for 4 weeks. Compared to the control mice, the KME-containing chow-fed mice showed increased whole body weights, a higher weight of quadricepses, and increased grip strengths in high KME-fed group (Fig. 6). These data present the morphological indication that KME might induce skeletal muscle hypertrophy in mice.
Fig. 6.
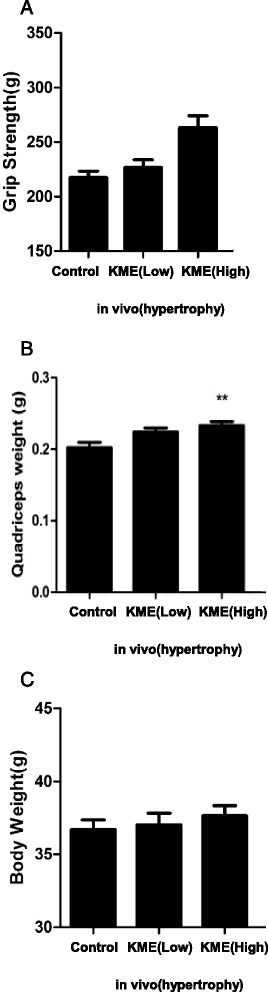
Physical changes in KME-fed mice (a) The grip strength of each chow-fed mouse is presented. b The weights of quadricepses are shown. c Whole body weights are shown. All the mice were fed chow for 4 weeks and analyses were done after the 4 weeks. Data shown are mean ± S.D.; n = 5 per group. P-values of < 0.05 and < 0.01 are indicate by * and **, respectively. KME(low) : 0.3%, KME(high) : 1.5%
KME showed increased endurance in vivo
To validate the muscle hypertrophic effect of KME, the treadmill and swimming pool tests were performed after the feeding of control chow or KME-containing chow over the 4 weeks. KME-containing chow-fed mice showed increased swimming activity and elevated running times on the treadmill compared to the control chow-fed mice (Fig. 7). In addition, skeletal muscle area and diameter were also increased in KME-fed mice (Fig. 8).
Fig. 7.
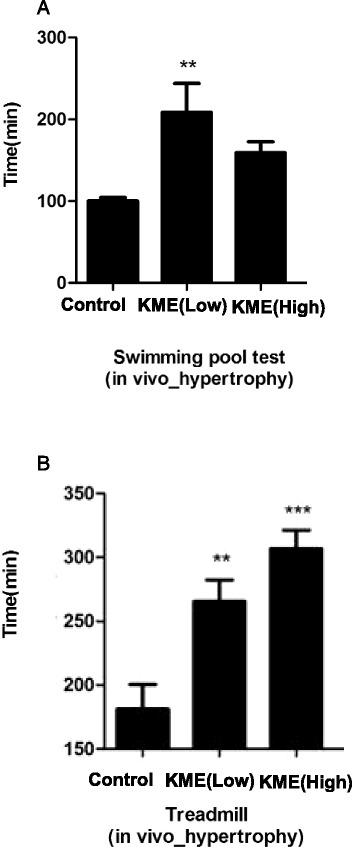
The increased endurance in the KME-fed mice (a) The endurance results of the mice measured by swimming pool tests are shown. b The endurance results of the mice measured by the treadmill tests are shown. All the mice were fed chow for 4 weeks. Data signifies means ± S.D.; n = 5 per group. P-values of < 0.05 and < 0.01 are indicate by * and **, respectively. KME(low) : 0.3%, KME(high) : 1.5%
Fig. 8.
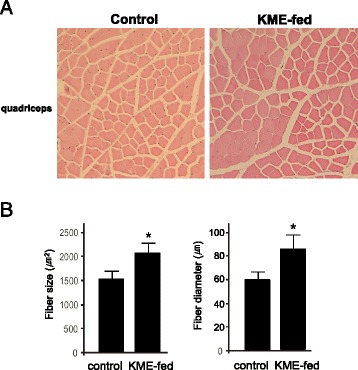
The increased muscle fiber size and diameter in the KME-fed(1.5%) mice (a) Cross-sections of quadriceps muscle stained with H & E. b Fiber size and diameters were measured from quadriceps muscle sections from a total of 500 fibers (n = 5 mice per group). All the mice were fed chow for 4 weeks. Data signifies means ± S.D.; P-values of < 0.05 is indicate by *
Discussion
Previous reports regarding muscle atrophy and hypertrophy have shown that muscle atrophy induced by various kinds of mechanisms—such as fasting, illness (cancer and diabetes), and muscle disuse—all result in similar specific increases of mRNA expression [30, 31]. These similar specific genes are called “atrogenes” [32]. In this present study, we showed that these genes (Murf1 and Atrogin-1) are downregulated by KME treatment, suggesting its effect on the inhibition of muscle atrophy. Atrogin-1 is known to promote the degradation of MyoD and elF3f [33]: MyoD is an important transcription factor of muscle and elF3f is one of the enhancer proteins of protein synthesis [34, 35]. Atrogin-1 degrades both of these proteins resulting in muscle atrophy. Murf1 regulates muscle structural proteins like troponin 1 and myosin [11, 36].
Mistletoe extracts were already used in broad spectrums as remedying agents [37, 38]. Based on the activation of the Akt signaling pathway and AMPK, we speculated that mistletoe extract could regulate cellular signaling involved in the regulation of muscle mass because these pathways are closely related to muscle atrophy and hypertrophy. Consistent with our prediction, we found that KME inhibits “atrogene” expression in C2C12 cells. Thus, we adopted an in vivo animal model to validate our observation in myotubes. In a subsequent study, we used a well-known method to induce muscle atrophy called denervation [39, 40]. After denervation of the sciatic nerves of the control mice, Murf1 and Atrogin-1 mRNA expression were increased, and the muscle mass were decreased. Grip strength was also reduced in the control mice. Compared with these control mice, KME treated mice showed a decreased expression of Atrogin-1 and Murf1, increased muscle mass, and greater grip strength.
We also investigated other signaling molecules involved in muscle atrophy repression and muscle hypertrophy in KME-treated cells and mice. However, there were no significant differences in the phosphorylation of mTOR and S6k by KME [41, 42]. Recently, it we reported that mTOR promotes muscle atrophy in denervated mice through the activation of FoxO and E3 ubiquitin ligases [43]. This is exactly opposite to the current paradigm depicting the function of mTOR in muscle hypertrophy and muscle atrophy [44]. The current dogma of mTOR functionality is the stimulation of muscle hypertrophy in skeletal muscle [45]. Therefore, there are still controversies about the role of mTOR in muscle physiology. Further studies are needed to figure out the relationship between the mTOR signaling pathway and the induction of muscle atrophy.
Another well-known signaling pathway involved in muscle atrophy is the NF-κB one [46]. This signaling pathway is also closely related with inflammation [47]. It was also reported that inflammation was repressed by mistletoe extract [48]. Further studies are required to elucidate whether KME treatment affects inflammation and NF-κB signaling in the inhibition of muscle atrophy.
Extending from the induced muscle atrophy model investigated, we also tried to test whether KME induce muscle hypertrophy in mice skeletal muscle. It is proven that an inhibition of muscle atrophy is associated with muscle hypertrophy [7]. We also proved this kind of hypertrophic effect with KME treatment in this study. Without any artificial induction of muscle atrophy, mice that fed on KME for 4 weeks showed increased muscle mass and grip strength. These results suggest that KME affects not only muscle atrophy but also muscle hypertrophy. Furthermore, our results also imply that KME can regulate both atrophy and hypertrophy independently. It was confirmed by the endurance tests that KME regulates muscle hypertrophy independent of muscle atrophy [49]. KME-containing chow-fed mice showed increased endurance compared to the control chow-fed mice. Muscle hypertrophy and increased endurance were closely related with the muscle fiber type switching [50, 51]. It is noteworthy that the study on the type switching of the muscle fiber might give us another molecular clue of the effects of KME in muscle regulation. In addition, a comparative study on the effects of KME in cardiac muscle and skeletal muscle is required—several reports demonstrated that the function of the mTOR signaling pathway is different between skeletal muscle and cardiac muscle [52].
In conclusion, the results of this study suggest that KME has positive regulatory effects on the maintenance of muscle mass. Our data shows the possibilities that KME could be used as a potential therapy for muscle atrophy in human cases.
Conclusions
Korean mistletoe extracts have inhibitory effects on muscle atrophy and stimulatory effects on muscle hypertrophy in C2C12 cells and denervated mice. These findings suggest that Korean mistletoe might be a potential candidate as a functional food promoting the restoration from muscle atrophy.
Acknowledgements
This research was supported by High Value-added Food Technology Development Program (112084–2, 2012), Ministry for Food, Agriculture, Forestry and Fisheries, Republic of Korea.
Availability of data and materials
The datasets supporting the conclusions of this article are included within the article.
Authors’ contributions
Conceived and designed the experiments: JSJ. Performed the experiments: JSJ and IBK. Analyzed the data: JSJ, CHP, YHK, JMY, KSK, and JBK. Wrote the paper: JSJ, CHP and JBK. All authors read and approved the final manuscript.
Competing interest
The authors declare that there is no conflict of interest regarding the publication of this article.
Consent for publication
Not applicable.
Ethics approval and consent to participate
All of the animal experiments were approved by the Ethics Review Committee of the Handong Global University.
Abbreviations
- ATCC
American Type Culture Collection
- DMEM
Dulbecco’s modified Eagle’s medium
- FBS
Fetal bovine serum
- KME
Korean mistletoe extract
- MAPK
Mitogen-activated protein kinase
- NC
Nitrocellulose
- PBS
Phosphate-buffered saline.
References
- 1.Lecker SH, Solomon V, Mitch WE, Goldberg AL. Muscle protein breakdown and the critical role of the ubiquitin-proteasome pathway in normal and disease states. J Nutr. 1999;129:227S–237S. doi: 10.1093/jn/129.1.227S. [DOI] [PubMed] [Google Scholar]
- 2.Ruegg MA, Glass DJ. Molecular mechanisms and treatment options for muscle wasting diseases. Annu Rev Pharmacol Toxicol. 2011;51:373–395. doi: 10.1146/annurev-pharmtox-010510-100537. [DOI] [PubMed] [Google Scholar]
- 3.Velloso CP. Regulation of muscle mass by growth hormone and IGF-I. Br J Pharmacol. 2008;154:557–568. doi: 10.1038/bjp.2008.153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Glass DJ. Skeletal muscle hypertrophy and atrophy signaling pathways. Int J Biochem Cell Biol. 2005;37:1974–1984. doi: 10.1016/j.biocel.2005.04.018. [DOI] [PubMed] [Google Scholar]
- 5.Goldberg AL, Etlinger JD, Goldspink DF, Jablecki C. Mechanism of work-induced hypertrophy of skeletal muscle. Med Sci Sports. 1975;7:185–198. [PubMed] [Google Scholar]
- 6.Bodine SC, Latres E, Baumhueter S, Lai VK, Nunez L, et al. Identification of ubiquitin ligases required for skeletal muscle atrophy. Science. 2001;294:1704–1708. doi: 10.1126/science.1065874. [DOI] [PubMed] [Google Scholar]
- 7.Sacheck JM, Ohtsuka A, McLary SC, Goldberg AL. IGF-I stimulates muscle growth by suppressing protein breakdown and expression of atrophy-related ubiquitin ligases, atrogin-1 and MuRF1. Am J Physiol Endocrinol Metab. 2004;287:E591–601. doi: 10.1152/ajpendo.00073.2004. [DOI] [PubMed] [Google Scholar]
- 8.Foletta VC, White LJ, Larsen AE, Leger B, Russell AP. The role and regulation of MAFbx/atrogin-1 and MuRF1 in skeletal muscle atrophy. Pflugers Arch. 2011;461:325–335. doi: 10.1007/s00424-010-0919-9. [DOI] [PubMed] [Google Scholar]
- 9.Jackman RW, Kandarian SC. The molecular basis of skeletal muscle atrophy. Am J Physiol Cell Physiol. 2004;287:C834–843. doi: 10.1152/ajpcell.00579.2003. [DOI] [PubMed] [Google Scholar]
- 10.Gomes MD, Lecker SH, Jagoe RT, Navon A, Goldberg AL. Atrogin-1, a muscle-specific F-box protein highly expressed during muscle atrophy. Proc Natl Acad Sci U S A. 2001;98:14440–14445. doi: 10.1073/pnas.251541198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Clarke BA, Drujan D, Willis MS, Murphy LO, Corpina RA, et al. The E3 Ligase MuRF1 degrades myosin heavy chain protein in dexamethasone-treated skeletal muscle. Cell Metab. 2007;6:376–385. doi: 10.1016/j.cmet.2007.09.009. [DOI] [PubMed] [Google Scholar]
- 12.Lokireddy S, Wijesoma IW, Sze SK, McFarlane C, Kambadur R, et al. Identification of atrogin-1-targeted proteins during the myostatin-induced skeletal muscle wasting. Am J Physiol Cell Physiol. 2012;303:C512–529. doi: 10.1152/ajpcell.00402.2011. [DOI] [PubMed] [Google Scholar]
- 13.Sandri M, Sandri C, Gilbert A, Skurk C, Calabria E, et al. Foxo transcription factors induce the atrophy-related ubiquitin ligase atrogin-1 and cause skeletal muscle atrophy. Cell. 2004;117:399–412. doi: 10.1016/S0092-8674(04)00400-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Stitt TN, Drujan D, Clarke BA, Panaro F, Timofeyva Y, et al. The IGF-1/PI3K/Akt pathway prevents expression of muscle atrophy-induced ubiquitin ligases by inhibiting FOXO transcription factors. Mol Cell. 2004;14:395–403. doi: 10.1016/S1097-2765(04)00211-4. [DOI] [PubMed] [Google Scholar]
- 15.Vandenburgh H, Kaufman S. In vitro model for stretch-induced hypertrophy of skeletal muscle. Science. 1979;203:265–268. doi: 10.1126/science.569901. [DOI] [PubMed] [Google Scholar]
- 16.Altomare DA, Testa JR. Perturbations of the AKT signaling pathway in human cancer. Oncogene. 2005;24:7455–7464. doi: 10.1038/sj.onc.1209085. [DOI] [PubMed] [Google Scholar]
- 17.Wu YT, Tan HL, Huang Q, Ong CN, Shen HM. Activation of the PI3K-Akt-mTOR signaling pathway promotes necrotic cell death via suppression of autophagy. Autophagy. 2009;5:824–834. doi: 10.4161/auto.9099. [DOI] [PubMed] [Google Scholar]
- 18.Adesina SK, Illoh HC, Johnny II, Jacobs IE. African mistletoes (Loranthaceae); ethnopharmacology, chemistry and medicinal values: an update. Afr J Tradit Complement Altern Med. 2013;10:161–170. doi: 10.4314/ajtcam.v10i4.26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Moghadamtousi SZ, Kamarudin MN, Chan CK, Goh BH, Kadir HA. Phytochemistry and biology of Loranthus parasiticus Merr, a commonly used herbal medicine. Am J Chin Med. 2014;42:23–35. doi: 10.1142/S0192415X14500025. [DOI] [PubMed] [Google Scholar]
- 20.Khwaja TA, Varven JC, Pentecost S, Pande H. Isolation of biologically active alkaloids from Korean mistletoe Viscum album, coloratum. Experientia. 1980;36:599–600. doi: 10.1007/BF01965825. [DOI] [PubMed] [Google Scholar]
- 21.Lee SH, An HS, Jung YW, Lee EJ, Lee HY, et al. Korean mistletoe (Viscum album coloratum) extract extends the lifespan of nematodes and fruit flies. Biogerontology. 2014;15:153–164. doi: 10.1007/s10522-013-9487-7. [DOI] [PubMed] [Google Scholar]
- 22.Lee CH, Kim JK, Kim HY, Park SM, Lee SM. Immunomodulating effects of Korean mistletoe lectin in vitro and in vivo. Int Immunopharmacol. 2009;9:1555–1561. doi: 10.1016/j.intimp.2009.09.011. [DOI] [PubMed] [Google Scholar]
- 23.Kim KW, Yang SH, Kim JB. Protein fractions from Korean mistletoe (Viscum album coloratum) extract induce insulin secretion from pancreatic beta cells. Evid Based Complement Alternat Med. 2014;2014:703624. doi: 10.1155/2014/703624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Jung HY, Lee AN, Song TJ, An HS, Kim YH, et al. Korean mistletoe (Viscum album coloratum) extract improves endurance capacity in mice by stimulating mitochondrial activity. J Med Food. 2012;15:621–628. doi: 10.1089/jmf.2010.1469. [DOI] [PubMed] [Google Scholar]
- 25.Shavlakadze T, White JD, Davies M, Hoh JF, Grounds MD. Insulin-like growth factor I slows the rate of denervation induced skeletal muscle atrophy. Neuromuscul Disord. 2005;15:139–146. doi: 10.1016/j.nmd.2004.10.013. [DOI] [PubMed] [Google Scholar]
- 26.Glass DJ. Signaling pathways perturbing muscle mass. Curr Opin Clin Nutr Metab Care. 2010;13:225–229. doi: 10.1097/MCO.0b013e32833862df. [DOI] [PubMed] [Google Scholar]
- 27.Stuart CA, Howell ME, Baker JD, Dykes RJ, Duffourc MM, et al. Cycle training increased GLUT4 and activation of mammalian target of rapamycin in fast twitch muscle fibers. Med Sci Sports Exerc. 2010;42:96–106. doi: 10.1249/MSS.0b013e3181ad7f36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Lecomte V, Meugnier E, Euthine V, Durand C, Freyssenet D, et al. A new role for sterol regulatory element binding protein 1 transcription factors in the regulation of muscle mass and muscle cell differentiation. Mol Cell Biol. 2010;30:1182–1198. doi: 10.1128/MCB.00690-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Pedersen BK, Febbraio MA. Muscles, exercise and obesity: skeletal muscle as a secretory organ. Nat Rev Endocrinol. 2012;8:457–465. doi: 10.1038/nrendo.2012.49. [DOI] [PubMed] [Google Scholar]
- 30.Kunkel SD, Suneja M, Ebert SM, Bongers KS, Fox DK, et al. mRNA expression signatures of human skeletal muscle atrophy identify a natural compound that increases muscle mass. Cell Metab. 2011;13:627–638. doi: 10.1016/j.cmet.2011.03.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Glass DJ. Molecular mechanisms modulating muscle mass. Trends Mol Med. 2003;9:344–350. doi: 10.1016/S1471-4914(03)00138-2. [DOI] [PubMed] [Google Scholar]
- 32.Lecker SH, Jagoe RT, Gilbert A, Gomes M, Baracos V, et al. Multiple types of skeletal muscle atrophy involve a common program of changes in gene expression. FASEB J. 2004;18:39–51. doi: 10.1096/fj.03-0610com. [DOI] [PubMed] [Google Scholar]
- 33.Lagirand-Cantaloube J, Cornille K, Csibi A, Batonnet-Pichon S, Leibovitch MP, et al. Inhibition of atrogin-1/MAFbx mediated MyoD proteolysis prevents skeletal muscle atrophy in vivo. PLoS One. 2009;4:e4973. doi: 10.1371/journal.pone.0004973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Lassar AB, Buskin JN, Lockshon D, Davis RL, Apone S, et al. MyoD is a sequence-specific DNA binding protein requiring a region of myc homology to bind to the muscle creatine kinase enhancer. Cell. 1989;58:823–831. doi: 10.1016/0092-8674(89)90935-5. [DOI] [PubMed] [Google Scholar]
- 35.Weintraub H, Davis R, Tapscott S, Thayer M, Krause M, et al. The myoD gene family: nodal point during specification of the muscle cell lineage. Science. 1991;251:761–766. doi: 10.1126/science.1846704. [DOI] [PubMed] [Google Scholar]
- 36.Kedar V, McDonough H, Arya R, Li HH, Rockman HA, et al. Muscle-specific RING finger 1 is a bona fide ubiquitin ligase that degrades cardiac troponin I. Proc Natl Acad Sci U S A. 2004;101:18135–18140. doi: 10.1073/pnas.0404341102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Grossarth-Maticek R, Kiene H, Baumgartner SM, Ziegler R. Use of Iscador, an extract of European mistletoe (Viscum album), in cancer treatment: prospective nonrandomized and randomized matched-pair studies nested within a cohort study. Altern Ther Health Med. 2001;7:57–66. [PubMed] [Google Scholar]
- 38.Janssen O, Scheffler A, Kabelitz D. In vitro effects of mistletoe extracts and mistletoe lectins. Cytotoxicity towards tumor cells due to the induction of programmed cell death (apoptosis) Arzneimittelforschung. 1993;43:1221–1227. [PubMed] [Google Scholar]
- 39.Kohn RR. Mechanism of protein loss in denervation muscle atrophy. Am J Pathol. 1964;45:435–447. [PMC free article] [PubMed] [Google Scholar]
- 40.Goldberg AL. Protein turnover in skeletal muscle. II. Effects of denervation and cortisone on protein catabolism in skeletal muscle. J Biol Chem. 1969;244:3223–3229. [PubMed] [Google Scholar]
- 41.Marcotte GR, West DW, Baar K. The Molecular Basis for Load-Induced Skeletal Muscle Hypertrophy. Calcif Tissue Int. 2015;96:196–210. [DOI] [PMC free article] [PubMed]
- 42.Bonaldo P, Sandri M. Cellular and molecular mechanisms of muscle atrophy. Dis Model Mech. 2013;6:25–39. doi: 10.1242/dmm.010389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Tang H, Inoki K, Lee M, Wright E, Khuong A, et al. mTORC1 promotes denervation-induced muscle atrophy through a mechanism involving the activation of FoxO and E3 ubiquitin ligases. Sci Signal. 2014;7:ra18. doi: 10.1126/scisignal.2004809. [DOI] [PubMed] [Google Scholar]
- 44.Goodman CA, Frey JW, Mabrey DM, Jacobs BL, Lincoln HC, et al. The role of skeletal muscle mTOR in the regulation of mechanical load-induced growth. J Physiol. 2011;589:5485–5501. doi: 10.1113/jphysiol.2011.218255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Sandri M, Barberi L, Bijlsma AY, Blaauw B, Dyar KA, et al. Signalling pathways regulating muscle mass in ageing skeletal muscle: the role of the IGF1-Akt-mTOR-FoxO pathway. Biogerontology. 2013;14:303–323. doi: 10.1007/s10522-013-9432-9. [DOI] [PubMed] [Google Scholar]
- 46.Schakman O, Dehoux M, Bouchuari S, Delaere S, Lause P, et al. Role of IGF-I and the TNFalpha/NF-kappaB pathway in the induction of muscle atrogenes by acute inflammation. Am J Physiol Endocrinol Metab. 2012;303:E729–739. doi: 10.1152/ajpendo.00060.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Tak PP, Firestein GS. NF-kappaB: a key role in inflammatory diseases. J Clin Invest. 2001;107:7–11. doi: 10.1172/JCI11830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Hegde P, Maddur MS, Friboulet A, Bayry J, Kaveri SV. Viscum album exerts anti-inflammatory effect by selectively inhibiting cytokine-induced expression of cyclooxygenase-2. PLoS One. 2011;6:e26312. doi: 10.1371/journal.pone.0026312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Riedy M, Moore RL, Gollnick PD. Adaptive response of hypertrophied skeletal muscle to endurance training. J Appl Physiol. 1985;59:127–131. doi: 10.1152/jappl.1985.59.1.127. [DOI] [PubMed] [Google Scholar]
- 50.Handschin C, Chin S, Li P, Liu F, Maratos-Flier E, et al. Skeletal muscle fiber-type switching, exercise intolerance, and myopathy in PGC-1alpha muscle-specific knock-out animals. J Biol Chem. 2007;282:30014–30021. doi: 10.1074/jbc.M704817200. [DOI] [PubMed] [Google Scholar]
- 51.Parsons SA, Millay DP, Wilkins BJ, Bueno OF, Tsika GL, et al. Genetic loss of calcineurin blocks mechanical overload-induced skeletal muscle fiber type switching but not hypertrophy. J Biol Chem. 2004;279:26192–26200. doi: 10.1074/jbc.M313800200. [DOI] [PubMed] [Google Scholar]
- 52.Lee CH, Inoki K, Guan KL. mTOR pathway as a target in tissue hypertrophy. Annu Rev Pharmacol Toxicol. 2007;47:443–467. doi: 10.1146/annurev.pharmtox.47.120505.105359. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets supporting the conclusions of this article are included within the article.